Embed Size (px)
Citation preview

Acta physiol. scand. 1973. 87. 158-167 From the Department of Pharmacology, Karolinska Institutet, Stockholm, Sweden
Effect of Divalent Cations on the Interaction of Adenosine 5'-Triphosphate (ATP) with Histamine Release Induced
by Compound 48/80 BY
RUNE DAHLQUIST, BERTIL DIAMANT and KERSTIN ELWIN
Received 21 December 1971
Abstract
DAHLQUIST, R., B. DIAMANT and K. ELWIN. Effect of divalent cations on the inter- action of adenosine 5'-triphosphate ( A T P ) with histamine release induced by compound 48/80. Acta physiol. scand. 1973. 87. 158-167.
The inhibition exerted by ATP on histamine release caused by compound 48/80 from isolated rat mast cells was counteracted by Ca'+, Mg", Bait, and Sr++, but not by Be++. In addition, after the inhibitory effect had been obtained by pretreatment of mast cells with ATP, addition of !tlg++ restored the sensitivity of the cells towards 48/80. The spontaneous restoration of the sensitivity of the mast cells towards 48/80 observed after prolonged pretreatment of the cells with ATP (< 10 pM) was enhanced by the presence of glucose. Glucose did not effect the inhibition per se. EDTA blocked 1 ) the protecting effect of divalent cations on the action of ATP on mast cells in regard to their sensitivity towards 48/80, 2 ) the action of Mg++ to restore the sensitivity of mast cells towards 48/80 after rendered insensitive by treatment with .%TP, 3) the spontaneous restoration of the sensitivity of ATP-treated mast cells towards 48/80 observed after prolonged incubation alone, as well as the enhanced restoration observed in the presence of glucose. The results suggest an interaction between divalent cations and ATP on the plasma membrane.
Adenosine 5'-triphosphate (ATP) exerts various effects on isolated rat mast cells. In the presence of Ca" histamine release is induced (Diamant and Kruger 1967, Sugiyama and Yamasaki 1969). In the absence of divalent cations ATP renders mast cells insensitive towards the degranulating and histamine releasing effect of compound 48/80 (Dahlquist et al. 1973 a ) . In addition, ATP causes changes in the distribution of sodium and potassium over the mast cell membrane, effects that are inhibited by increasing concentrations of divalent cations (Dahlquist et al. To be publ.). Furthermore, ATP stimulates the uptake of 45Ca to the cells from the incuba- tion medium (Dahlquist and Diamant 1972). Also, ATP causes swelling of the mast cells together with certain specific intracellular morphological changes (Diamant and Kruger 1968, Bloom et al. 1970, Sugiyama 1971). This swelling is counteracted by the presence of Cat+ 1 mM and above (Kriiger et al. to be publ.). In addition,
158

INTERACTION OF ATP WITH 48/80 159 in the presence of 0.25 mM of Ca" swelling and intracellular morphological changes of the mast cells precedes a slowly occurring degranulating process which in time is related to the histamine release (Sugiyama 1971, Kruger ct al. to be publ.).
The present study was performed in order to further study the characteristics of the inhibition of 48/80-induced histamine release from rat mast cells caused by extra- cellularly applied ATP. as well as the mechanism behind the spontaneous cessation of this inhibition observed after prolonged preincubation of the cells with A T P i< 10 m M ) (Dahlquist ct al. 1973 a ) .
.4 preliminary report on parts of the present study has been presented earlier (Diamant and Dahlquist 1970).
Methods and Materials Isolatiori of rut must cells Vast cells from the peritoneal and pleural cavities of Sprague-Dawley rats (male, weight 350- 450 g ) were isolated by density gradient centrifugation in Ficoll according to the method of Thon and Uvnas ( 1966). Mast cells collected from the Ficoll were centrifuged (350 x g) and washed three times in room temperature, each time with 5 ml of a buffered salt solution con- taining NaCl 145 mM. KCI 2.7 mhl and which was buffered to p H 7.0 with 10 70 ( v / v ) Sorensen phosphate buffer (NazHPOJ + KH2p0-1, 67 m M ) . In addition, the buffered salt solu- tion conwined human serum albumin, 1 mglml. After washing, the mast cells were finally resuspended in 0.3-1.0 ml of the buffered salt solution giving a final concentration of 1.2- 3 . 8 ~ 10" cells/nil. The cells were counted in a Biirker chamber.
In cu bu tion p roced tire Isolated rat mast cells were incubated in 2 ml of the buffered salt solution ( v i d e supra) a t 37" C giving a final concentration varying in different experiments between 2-5 x lo3 cells/ml.
.4fter preincubation of the cells with .4TP for various lengths of time compound 48/80 was added. The incubation was continued for an additional 10 min and was terminated by placing the tubes in an ice-bath.
After incubation the tubes were centrifuged for 10 min at 350 x g at 2-4" C. T h e super- natants were decanted into new tubes and 2 ml of distilled water were added to the cell sedi- ment to extract the remaining histamine.
Histamine determination Histamine was determined by the fluorescence method of Shore e t a1 (1959), omitting the extraction steps (Bergendorff 1965).
The fluorescence was measured in a Farrand fluoronieter Model .42 or a Farrand ratio fluorometcr (Farrand Optical Co. Ltd, USA).
The histamine release is expressed as percentage appearing in the supernatant of the total histamine content of the cells.
The spontaneous release of histamine (i.e. in the absence of compound 48/80 and ATP) arnounted to < 7 54 and was deducted from all values presented. T h e release obtained by .4TP in the absence of 48/80 was subtracted from the histamine release values observed in cell samples exposed to both ATP and 48/80.
.411 samples were run in duplicate.
Materials Compound 48/80 was kindly supplied by AB Leo, Helsingborg, Sweden, and human serum albumin by AB Kabi, Stockholm, Sweden. Ficoll was purchased from AB Pharmacia, Uppsala, Sweden. .4denosine 5'-triphosphate ( A T P ) (sodium salt) was obtained from Sigma Chemical Comp., St. Louis, USA. Stock solution of ATP (about 0.1 M ) was neutralized by the addition of NaOH and assayed spectrophotometrically. The stock solution was stored a t -20" C. The divalent cations investigated were all chloride salts.
When necessary, the pH of solutions to be used was adjusted to 7.0.
160 RUNE DAHLQUIST, BERTIL DIAMANT AND KERSTIN ELWIN
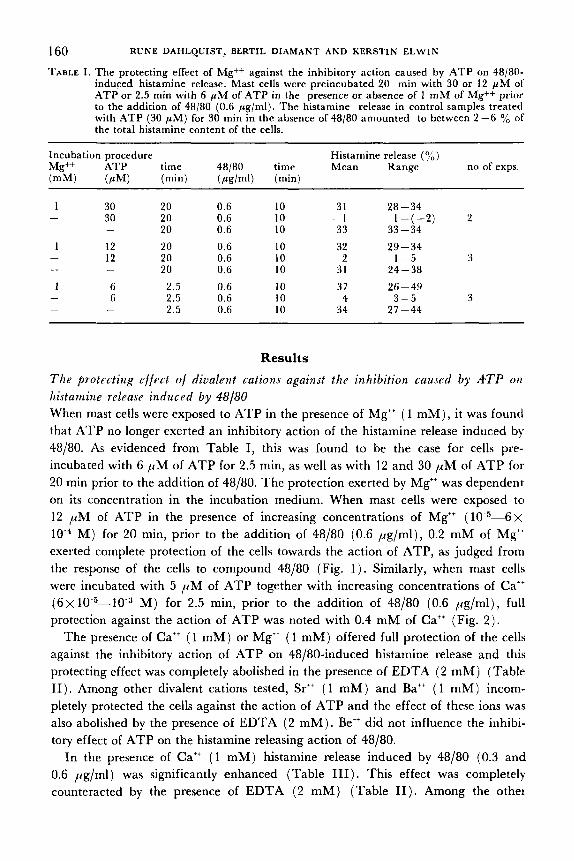
TABLE I . The protecting effect of Mg++ against the inhibitory action caused by ATP on 48/80- induced histamine release. Mast cells were preincubated 20 min with 30 or 12 p M of ATP or 2.5 min with 6 pM of ATP in the presence or absence of 1 mM of Mg++ prior to the addition of 48/80 (0.6 pg/ml). The histamine release in control samples treated with ATP (30 pM) for 30 min in the absence of 48/80 amounted to between 2 -6 yo of the total histamine content of the cells.
Incubation procedure Histamine release (yo) Mg++ ATP time 48/80 time Mean Range no of exps. (mM) (PM) (min) (pcglml) (min)
30 30
12 12
6 6
-
-
20 0.6 20 0.6 20 0.6 20 0.6 20 0.6 20 0.6
2.5 0.6 2.5 0.6 2.5 0.6
10 10 10
10 10 10
10 10 10
31 28-34
33 33-34 32 29-34
- 1 I - ( - 2 ) 2
2 1-5 3 31 24-38 37 26-49
34 27-44 4 3-5 3
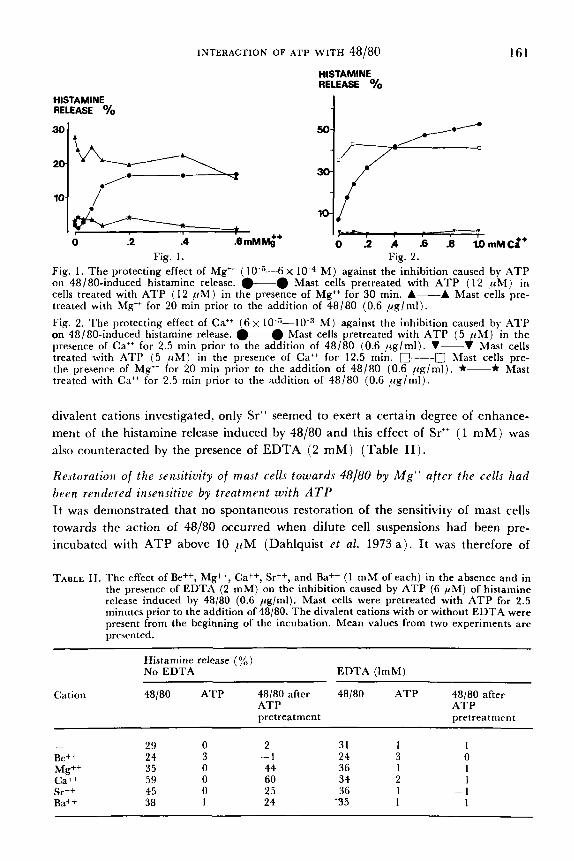
Results T h e protecting effect of diualent cations against the inhibition rauJed by ATP ori histamine release induced by 48/80 When mast cells were exposed to ATP in the presence of Mg" ( 1 mM), it was found that ATP no longer exerted an inhibitory action of the histamine release induced by 48/80. As evidenced from Table I, this was found to be the case for cells pre- incubated with 6 pM of ATP for 2.5 min, as well as with 12 and 30 pM of ATP for 20 rnin prior to the addition of 48/80. The protection exerted by Mg" was dependent on its concentration in the incubation medium. When mast cells were exposed to 12 pM of ATP in the presence of increasing concentrations of Mg" (10 5-6 x 10 M ) for 20 min, prior to the addition of 48/80 (0.6 pg/ml), 0.2 mM of Mg" exerted complete protection of the cells towards the action of ATP, as judged from the response of the cells to compound 48/80 (Fig. 1 ) . Similarly, when mast cells were incubated with 5 p M of ATP together with increasing concentrations of Ca" ( 6 ~ 10 5-10 M ) for 2.5 min, prior to the addition of 48/80 (0.6 pg/ml), full protection against the action of ATP was noted with 0.4 mM of Cat+ (Fig. 2 ) .
The presence of Cat+ ( 1 mM) or Mg" (1 mM) offered full protection of the cells against the inhibitory action of .4TP on 48/80-induced histamine release and this protecting effect was completely abolished in the presence of EDTA ( 2 mM) (Table 11). Among other divalent cations tested, Sr++ (1 mM) and Ba" ( 1 mM) incom- pletely protected the cells against the action of ATP and the effect of these ions was also abolished by the presence of EDTA ( 2 mM) . Be++ did not influence the inhibi- tory effect of ATP on the histamine releasing action of 48/80.
In the presence of Ca" ( 1 mM) histamine release induced by 48/80 (0.3 and 0.6 pg/ml) was significantly enhanced (Table 111). This effect was completely counteracted by the presence of EDTA ( 2 mM) (Table 11). Among the othei
161
HISTAMINE RELEASE '/o
50- =- A ! -
10 + 30
.L 20- \AA\./-----A
10- / /* &J.----t
i /
/ .- Fig. 1. Fig. 2.
Fig. 1. The protecting effect of Mg++ (10 5-6 X M ) against the inhibition caused by ATP on 48/80-induced histamine release. *@ Mast cells pretreated with ATP ( 1 2 p M ) in cells treated with ATP ( 1 2 p M ) in the presence of Mg++ for 30 min. A--A Mast cells pre- treated with Mg" for 20 min prior to the addition of 48/80 (0.6 ,ug/ml). Fig. 2. The protecting effect of Cat+ (6x 10 "10 M ) against the inhibition caused by .4TP on 48/80-induced histamine release. -0 Mast cells pretreated with ATP ( 5 p M ) in the presence of Ca++ for 2.5 min prior to the addition of 48/80 (0.6 pg/ml) . V-V Mast cells treated with ATP ( 5 pM) in the presence of Ca++ for 12.5 min. ~ - - - - ~ Mast cells pre- the presence of Mg" for 20 niin prior to the addition of 48/80 (0.6 pglml) . *-* Mast treated with Cat+ for 2.5 niin prior to the addition of 48/80 (0.6 rcglml).
divalent cations investigated, only Sr" seemed to exert a certain degree of enhance- ment of the histamine release induced by 48/80 and this effect of Sr" (1 mM) was also counteracted by the presence of EDTA ( 2 mM) (Table 11).
Restoration o/ the sensitivity of mast cells towards 48/80 by Mg" after the cells had been rendered insensitive by treatment wi th ATP It was demonstrated that no spontaneous restoration of the sensitivity of mast cells towards the action of 48/80 occurred when dilute cell suspensions had been pre- incubated with ATP above 10 pM (Dahlquist et al. 1973 a ) . It was therefore of
TABLE 11. The effect of Be++, Mg++, Ca++, Sr++, and Bat+ ( 1 m M of each) in the absence and in the presence of EDTA (2 m M ) on the inhibition caused by ATP (6 p M ) of histamine release induced by 48/80 (0.6 pg/ml). Mast cells were pretreated with ATP for 2.5 minutes prior to the addition of 48/80, The divalent cations with or without EDTA were present from the beginning of the incubation. Mean values from two experiments are presented.
~
Histamine release (%) No EDTA EDTA (ImM)
Cation 48/80 ATP 48/80 after 48/80 ATP 48/80 after ATP ATP pretreatment pretreatment
- 29 0 2 31 1 1 Be++ 24 3 - I 24 3 0
Cat+ 59 0 60 34 2 1 Sr++ 45 0 25 36 1 - 1 Ba++ 38 1 24 '35 1 1
Mg++ 35 0 44 36 1 I
162 RUNE DAHLQUIST, BERTIL DIAMANT AND KERSTIN ELWIN
30-
20-
HISTAMINE RELEASE '/o
Ab7=\*
Fig. 3. Fig. 4.
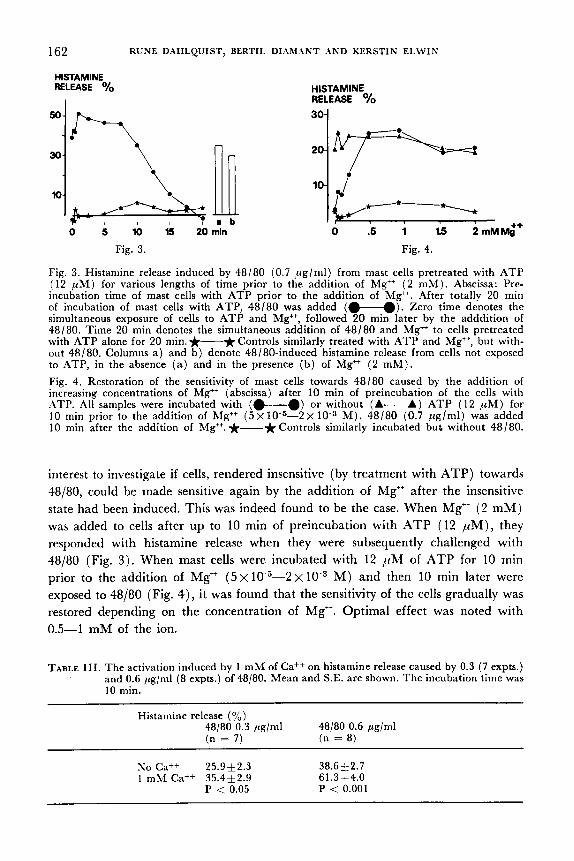
Fig. 3. Histamine release induced by 48/80 (0.7 pglml) from mast cells pretreated with ATP (12 pM) for various lengths of time prior to the addition of Mg++ ( 2 mM). Abscissa: Pre- incubation time of mast cells with ATP prior to the addition of Mg". .4fter totally 20 min of incubation of mast cells with ATP, 48/80 was added (w). Zero time denotes the simultaneous exposure of cells to ATP and Mg++, followed 20 min later by the adddition of 48/80. Time 20 min denotes the simultaneous addition of 48/80 and Mg++ to cells pretreated with ATP alone for 20 min. *-* Controls similarly treated with ATP and Mg++, but with- out 48/80. Columns a ) and b ) denote 48180-induced histamine release from cells not exposed to ATP, in the absence (a) and in the presence (b ) of Mg++ ( 2 mM). Fig. 4. Restoration of the sensitivity of mast cells towards 48/80 caused by the addition of increasing concentrations of Mg" (abscissa) after 10 min of preincubation of the cells with ATP. .411 samples were incubated with (M) or without (A-A) ATP (12 p M ) for 10 min prior to the addition of Mg++ ( 5 ~ 1 0 - s - 2 ~ M ) . 48/80 (0.7 pglml) was added 10 min after the addition of Mg++. *-* Controls similarly incubated but without 48/80.
interest to investigate if cells, rendered insensitive (by treatment with ATP) towards 48/80, could be made sensitive again by the addition of Mg" after the insensitive state had been induced. This was indeed found to be the case. When Mg" ( 2 mM) was added to cells after up to 10 rnin of preincubation with .4TP (12 pM) , they responded with histamine release when they were subsequently challenged with 48/80 (Fig. 3). When mast cells were incubated with 12 ,rtM of ATP for 10 rnin prior to the addition of Mg" ( 5 ~ 1 0 - ~ - - 2 ~ M ) and then 10 rnin later were exposed to 48/80 (Fig. 4), it was found that the sensitivity of the cells gradually was restored depending on the concentration of Mg". Optimal effect was noted with 0.5-1 mM of the ion.
TABLE 111. The activation induced by 1 mM of Caf+ on histamine release caused by 0.3 (7 expts.) and 0.6 pg/ml (8 expts.) of 48/80. Mean and S.E. are shown. The incubation time was 10 min.
Histamine release (yo) 48/80 0.3 pg/ml (n = 7)
48/80 0.6 pg/ml (n = 8)
No Ca++ 25.912.3 38.612.7 1 mM Ca++ 35.412.9 61.3 1 4 . 0
P < 0.001 P < 0.05
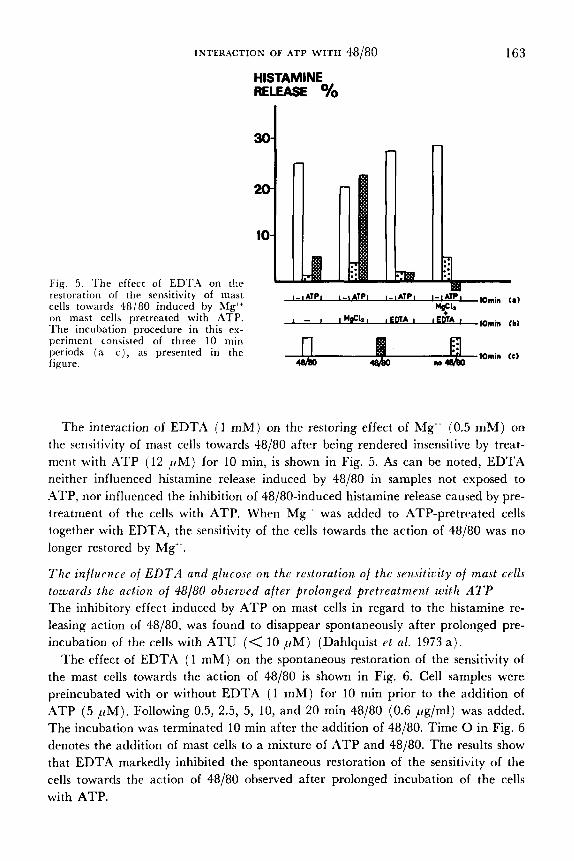
INTERACTION OF ATP WITH 48/80 163
HISTAMINE RELEASE '/I
n
Fig. 5. 'l'he effect of EDTA on t!:e testoration of the sensitivity of mast cells towards 48/80 induced by hfg" on mast cells pretreated with ATP. - I I M & ~ I # E M A I IFMA I 1Omin (bl The incubation procedure in this ex- periment consisted of three 10 min periods (a-c), as presented in the 1Omin Cc) figure.
The interaction of EDT.4 (1 m M ) on the restoring effect of Mg" (0.5 m M ) on the sensitivity of mast cells towards 48/80 after being rendered insensitive by treat- ment with ATP (12 pM) for 10 min, is shown in Fig. 5. As can be noted, EDTA neither influenced histamine release induced by 48/80 in samples not exposed to .4TP, nor influenced the inhibition of 48/80-induced histamine release caused by pre- treatment of the cells with ATP. When Mg" was added to ATP-pretreated cells together with EDTA, the sensitivity of the cells towards the action of 48/80 was no longer restored by Mg".
T h e influence of E D T A and glucose on the restoration of the senritivity of mast cells towards the action of 48/80 observrd after prolonged pretreatment with ATP The inhibitory effect induced by ATP on mast cells in regard to the histamine re- leasing action of 48/80, was found to disappear spontaneously after prolonged pre- incubation of the cells with ATU (< 10 p M ) (Dahlquist et al. 1973 a ) .
The effect of EDTA (1 mM) on the spontaneous restoration of the sensitivity of the mast cells towards the action of 48/80 is shown in Fig. 6. Cell samples were preincubated with or without EDTA (1 mM) for 10 rnin prior to the addition of ATP (5 ,MM). Following 0.5, 2.5, 5, 10, and 20 rnin 48/80 (0.6 pg/ml) was added. The incubation was terminated 10 rnin after the addition of 48/80. Time 0 in Fig. 6 denotes the addition of mast cells to a mixture of ATP and 48/80. The results show that EDTA markedly inhibited the spontaneous restoration of the sensitivity of the cells towards the action of 48/80 observed after prolonged incubation of the cells with ATP.
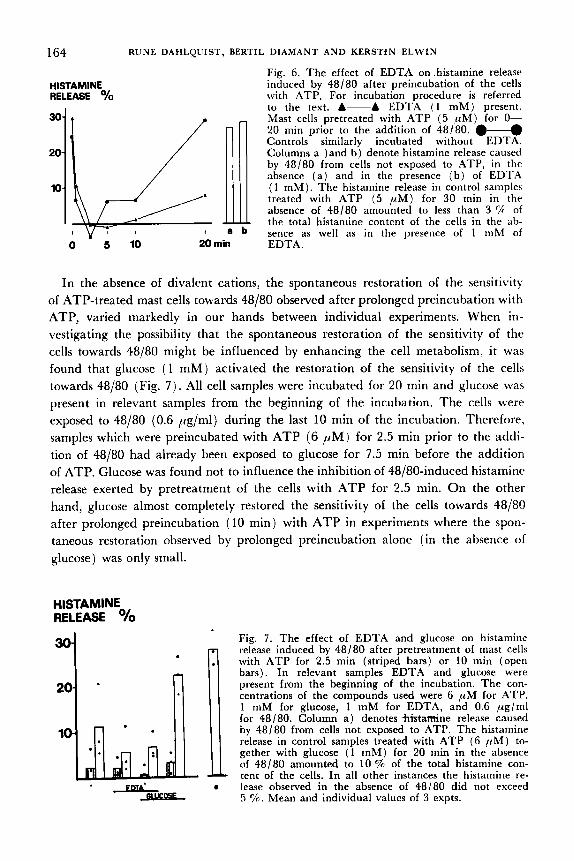
164 R U N E DAHLQUIST, BERTIL D I A M A N T A N D K E R S T i N E L W I N
Fig. 6. The effect of EDTA on histamine release HISTAMINE induced by 48/80 after preincubation of the cells RELEASE '/o with ATP. For incubation procedure is referred
to the text. A-A EDTA ( 1 mM) present. Mast cells pretreated with ATP ( 5 p M ) for 0- 20 niin prior to the addition of 48/80. Controls similarly incubated without EDTA. Columns a )and b ) denote histamine release caused by 48/80 from cells not exposed to ATP, in the absence ( a ) and in the presence ( b ) of EDTA
10 ( 1 mM). The histamine release in control samples treated with ATP ( 5 p M ) for 30 min in the absence of 48/80 amounted to less than 3 % of the total histamine content of the cells in the ab-
a sence as well as in the presence of 1 mhl of I~LlIl 0 5 10 20 rnin EDTA.
In the absence of divalent cations, the spontaneous restoration of the sensitivity of ATP-treated mast cells towards 48/80 observed after prolonged preincubation with ATP, varied markedly in our hands between individual experiments. When in- vestigating the possibility that the spontaneous restoration of the sensitivity of the cells towards 48/80 might be influenced by enhancing the cell metabolism, it was found that glucose (1 mM) activated the restoration of the sensitivity of the cells towards 48/80 (Fig. 7 ) . All cell samples were incubated for 20 rnin and glucose was present in relevant samples from the beginning of the incubation. The cells were exposed to 48/80 (0.6 pglml) during the last 10 rnin of the incubation. Therefore, samples which were preincubated with ATP (6 pM) for 2.5 rnin prior to the addi- tion of 48/80 had already been exposed to glucose for 7.5 rnin before the addition of ATP. Glucose was found not to influence the inhibition of 48/80-induced histamine release exerted by pretreatment of the cells with ATP for 2.5 min. O n the other hand, glucose almost completely restored the sensitivity of the cells towards 48/80 after prolonged preincubation (10 min) with ATP in experiments where the spon- taneous restoration observed by prolonged preincubation alone (in the absence of glucose) was only small.
HISTAMINE RELEASE '10
Fig. 7. The effect of EDTA and glucose on histamine release induced by 48/80 after pretreatment of mast cells with ATP for 2.5 min (striped bars) or 10 min (open bars). I n relevant samples EDTA and glucose were present from the beginning of the incubation. 'The con- centrations of the compounds used were 6 pM for ATP, 1 mM for glucose, 1 mM for EDTA, and 0.G ,ug/nil for 48/80. Column a ) denotes -histamine release caused by 48/80 from cells not exposed to ATP. The histamine release in control samples treated with ATP (6 p M ) to- gether with glucose ( 1 mM) for 20 min in the absence of 48/80 amounted to 10 76 of the total histamine con- tent of the cells. I n all other instances the histamine re- lease observed in the absence of 48/80 did not exceed 5 %. Mean and individual values of 3 expts.

INTERACTION OF ATP WITH 48/80 165
In addition the effect of EDTA (1 m M ) is presented regarding histamine release induced by 48/80 after pretreatment of the cells with ATP for 2.5 and 10 min in the absence and presence of glucose (Fig. 7 ) . The experiments show that 1 m M of EDTA counteracted the spontaneous restoration of the sensitivity of the mast cells towards 48/80 observed after I0 min of pretreatment of the cells with ATP, as well as the enhanced restoration caused by the presence of glucose.
Discussion
The present investigation has shown that certain divalent cations (Mg"? Ca++, Sr++, Ra") completely or partly protected the mast cells towards the inhibitory action of :ITP in regard to histamine release exerted by compound 48/80. This could indicate that these ions modify the action of .4TP on the plasma membrane. An interaction of divalent cations with ATP on the mast cell membrane is also suggested from other experimental data. Firstly, the swelling of mast cells induced by ATP was counter- acted by the presence of 1 mM of Ca" (Kruger c,t al. to be publ.). Secondly, the changes in distribution of sodium and potassium over the plasma membrane in- duced by ATP were counteracted by the presence of 1 m M of Ca" or Mg" (Dahl- quist P t al. to be publ.). Further support for an interaction between certain divalent cations and extracellularly applied ATP on the plasma membrane have been presented by Gasic and Stewart (1968) and by Stewart, Gasic and Hempling (1969). They found that the volume of TA3 ascites tuniour cells increased up to 50 % when exposed to ATP and that the increase in cell volume was prevented by the presence of Ca" or Mg". Furthermore, cells already swollen by the action of ATP (in the absence of divalent cations) began to shrink back to control size upon the addition of Ca" and Mg". The effect of ATP was specific and could not be mimicked by other triphosphonucleotides or by EDTA. Parallel to the swelling of ascites tumor cells, ATP caused a dramatic drop in cell potassium and an even greater rise in cell sodium (Hempling, Stewart and Gasic 1969). Thus, the findings reported regarding TA3 ascites tumour cells seem to well parallel observations on isolated rat mast cells.
Restoration of the sensitivity of the mast cells towards 48/80 was shown to be obtained when Mg" was added to cells rendered insensitive by treatment with ATP. This indicates, that at least after comparative short periods of exposure of the cells to ATP, the suggested configurational changes that ATP would induce on the plasma membrane and which resulted in resistance as to the effect of 48/80, could be reversed by Mg". This effect of Mg" was completely abolished by the presence of EDTA.
The insensitive state induced by ATP towards the action of 48/80 on mast cells was found to spontaneously disappear upon prolonged pretreatment of the cells with ATP (< I0 p M ) prior to the addition of 48/80. This effect was not due to ATP decreasing to ineffertive concentrations in the medium (Dahlquist ~t a/ . 1973 a ) .

166 RUNE DAHLQUIST, BERTIL DIAMANT AND KERSTIN ELWIN
The spontaneous restoration of the mast cells towards 48/80 observed after pro- longed incubation with ATP was found to be facilitated in the presence of glucose. On the other hand, glucose did not influence the inhibition observed after short periods of ATP pretreatment. Both the spontaneous restoration of the sensitivity of the mast cells towards 48/80 observed after prolonged preincubation of the cells with .4TP, as well as the enhanced restoration caused by glucose was found to be in- hibited by EDTA.
In summary the present study has revealed the following features in regard to tlie interaction of ATP with histamine release induced by 48/80:
a ) Divalent cations like Mg", Ca", Sr++, and Ba++, but not Be , when present together with ATP, counteracted the interaction of ATP with the effect of 48/80. EDTA blocked this effect of the divalent cations.
bl Mg" restored the sensitivity of ATP-treated mast cells towards the action of 48/80 after inhibition had occurred. EDTA abolished this effect of Mg".
c ) Provided the concentration of ATP was less than 10 pM, the sensitivity of ATP-treated mast cells towards the action of 48/80 was spontaneously restored by prolonged incubation. This spontaneous restoration was blocked by EDTA.
d ) The spontaneous restoration of the sensitivity of ATP-treated mast cells to- wards the action of 48/80 observed after prolonged incubation was enhanced by tlie presence of glucose. This effect of glucose was abolished by EDTA.
The experiments might therefore suggest that the spontaneous restoration of the sensitivity of ATP-treated mast cells towards the action of 48/80 is an energy- dependent process involving a factor which seems to react with EDTA and thus become inactivated. The function of this factor might be to counteract configu- rational changes of the plasma membrane induced by ATP, so that it again becomes reactive with 48/80, which would lead to degrada t ion and histamine release from the cell. Such a factor could be a divalent cation present in the mast cell.
This study was supported by the Swedish Medical Research Council (grant nr B71-14X-1004- 06A), Karolinska Institutet, Stockholm, Sweden, and Hesselmans Research Foundation. Sweden.
References BERCENDORFF, A, , Unpublished observation. 1965. BLOOM, G. D., B. DIAMANT, c). HAECERMARK and M. R I T Z ~ N , The effects Of adenosine-5'-tri-
phosphate (ATP) on structure and amine content of rat peritoneal mast cells. E.wp. Cell. Res. 1970. 62. 61-75.
DAHLQUIST, R. and B. DIAMANT, Relation of uptake of sodium and calcium to ATP-induced histamine release from rat mast cells. Proc. F i f t h I n t . ConEress on Phnrmacol. (Sari
I
Francisco) 1972. 50. DAHLQUIST, R., B. DIAMANT and K. ELWIN, Interaction of adenosine-5'-triphosphate (ATP)
with histamine release induced by compound 48/80. Acta physiol. scand. 1973 a. 87. 145- 157.
DAHLQUIST, R., B. DIAMANT and P. G. KRUCER, To be published. DIAMANT, B. and P. G. K R ~ ~ G E R , Histamine release from isolated rat peritoneal mast cells in-
duced by adenosine-5'-triphosphate. Acta physiol. scand. 1967. 71. 291-302. DIAMANT, B. and P. G. KRUGER, Structural changes of isolated rat peritoneal mast cells in-
duced by adenosine triphopshate and adenosine diphosphate. J . If istocliem. Cytoc/re?n. 1968. 16. 707-716.

RUNE DAHLQUIST, BERTIL DIAMANT AND KERSTIN ELWIN 167
DIAMANT, B. and R. DAHLQUIST, The inhibitory action of adenosine-5'-triphosphate (ATP) on histamine release and mast cell degranulation induced by compound 48/80. Acta pharrrra- col. (Kbh.) 1970. 28. Suppl. 1. 44.
G~src , G. and C. STEWART, Cell volume regulation in mouse TA3 ascites tumor cells by exogenous ATP as measured by the Coulter Counter. J. g e n . Physiol. 1968. 71. 239- 242.
HEMPLING, H. G., C. C. STEWAFU and G. CAW, The effect of exogenous ATP on the electro- lyte content of TA3 ascites tumor cells. J. gen . Physiol. 1969. 73. 133-140.
K R ~ G E R , P. G., B. DIARIANT and R. DAHLQUIST, To be published. SHORE, P. A., A. BURKHALTER and V. H. COIIN, A method for the fluorometric assay of
STEWART, C. C., G . GASIC and H. G. HEMPLING, Effect of cxogenous ATP on the volume of
SLIGIYAMA, K., Calcium-dependent histamine release with degranulation from isolated rat mast
SUGIYAMA, K. and H. YAMASAKI, Calcium-dependent histamine release by .4TP from isolated
THON, I. L. and B. Uv?;.is. hfode of storage of histamine in mast cells. A c f a ph]'siol. scand.
histamine in tissues. /. Pharmacol exp. Ther . 1959. 127. 182-186.
T.43 ascites tumor cells. J . gen. Physiol. 1969. 73. 125--132.
cells by adenosine-5'-triphosphate. Jap. J. Pharmacol. 1971. 21. 209-226.
rat mast cells. Jap. J . Pharmcol. 1969. 19. 175-176.
1966. 67. 455-470.