Embed Size (px)
Citation preview

Exp. Brain Res. 11, 128--144 (1970) �9 by Springer-Verlag 1970
Effects of Foveal Prestriate and Inferotemporal Lesions on Visual Discrimination by Rhesus Monkeys 1'2
A. C o w r y
Institute of Experimental Psychology, University of Oxford (England) and
C.G. G~oss
Department of Psychology, Princeton University (USA)
Received February 2, 1970
Summary . Ab la t ion of in fe ro tempora l cor tex in monkeys impai rs visual dis- c r iminat ion learning, and in fe ro tempora l cor tex receives v isual in format ion from s t r ia te cor tex b y way of the c i rcumst r ia te bel t . Ye t mos t previous s tudies have fai led to f ind a n y d isc r imina t ion impa i rmen t a f te r pa r t i a l ab la t ions of the cireum- s t r ia te bel t .
I n this exper imen t severe impa i rmen t s in pos t -opera t ive acquis i t ion and reten- t ion of visual d i sc r imina t ion problems were found af ter lesions of " foveal p res t r i a te cor tex" , i.e. the por t ion of the c i rcumst r ia te bel t which receives a p ro jec t ion f rom the cort ical r epresen ta t ion of the fovea in s t r ia te cor tex and which lies, la rgely bur ied, in the ven t ro la t e ra l por t ion of p res t r i a t e cortex. Al though foveal pre- s t r ia te lesions p roduced a grea te r impa i rmen t on ind iv idua l p a t t e r n d i sc r imina t ion tasks t h a n in fe ro tempora l lesions, the opposi te was t rue of concurrent visual dis- c r iminat ion tasks in which several different pai rs of d i sc r iminanda are p resen ted in each tes t ing session unt i l the an imal learns to d iscr iminate every pair .
The results are re la ted to a two-s tage model of d i scr imina t ion learning and i t is suggested t h a t foveal p res t r i a t e lesions impa i r visual a t t en t i on or percept ion, whereas in fero tempora l lesions d i s turb the associat ive or mnemonic s tage of visual d i scr imina t ion learning.
Key Words : Monkeys - - Visual d i scr imina t ion - - Tempora l lobe - - Visual cor tex
1 This work was supported by National Institute of Mental Health Grant MH-14471, National Science Foundation Grant GB-6999 and United Cerebral Palsy Research Grant R/213/67. For providing travel expenses to the United States A. Cowey wishes to thank the H.E. Durham Fund of King's College, Cambridge, and the Royal Society. The authors are particularly indebted to D.B. Bender for advice and technical assistance. We wish to thank Dr. Mortimer Mishkin for sharing with us his ideas, enthusiasm and unpublished data.
2 The terminology for the subdivisions of the non-striate visual areas of the occipital and temporal lobes of the monkey is still rather confusing. This is hardly surprising, for the subdivision of these areas on cytoarchitectonic grounds by different authorities is contradictory and the study of the properties of single units in these areas has only begun. Although the recent demonstrations by Zeki (1969b) and Cragg and Ainsworth (1969) that lateral striate cortex has two topographic and a third non-topographic projection onto prestriate cortex is a major step forward, the exact boundaries of these projections and their detailed relations to the various cytoarchitectonic subdivisions and subdivisions based on electrophysiological data are not yet entirely clear. Since the terminology used in behavioural studies of lesions of the non-striate visual areas is also inconsistent, it may be helpful to explain the terminology we have used in this report.

Cortical Lesions and Visual Discrimination 129
We have called our posterior lesions "foveal prestriate lesions" because they include the entire area of prestriate cortex to which foveal striate cortex selectively projects (Zeki, 1969b; Cragg and Ainsworth, 1969). Simply to call them prestriate cortex lesions or posterior inferotemporal cortex lesions is misleading because they include far less than the former and far more than the latter. Our inferotemporal lesions correspond closely to yon Bonin and Bailey's "Area TE". However, numerous publications on the behavioural effects of removing "inferotemporal" cortex illustrate lesions which often extend posterior to "Area TE" as far as the inferior occipital sulcus and thus may include part of foveal prestriate cortex. We there- fore wish to stress that our inferotemporal lesions are restricted to area TE and may not be comparable to some "inferotemporal" lesions in other studies.
Introduction
I t is well known tha t the cortex on the inferior convexity of the temporal lobe of rhesus monkeys is involved in vision, l~emoving it impairs visual, and only visual, discrimination tasks (see reviews by Mishkin, 1966; Gross, 1970). Single neurons in inferotemporal cortex have visual receptive fields and respond differ- entially to visual stimuli (Gross et al., 1969). Furthermore the primary visual cortex (area 17) provides afferent connexions to prestriate cortex which in turn projects to the inferotemporal region (Kuypers et al., 1965), and this occipital-temporal pathway appears to be necessary for normal visual discrimination (Mishkin, 1966). I t is therefore puzzling that several previous investigations found little or no im- pairment on visual tasks after lesions confined to parts of prestriate cortex or the "cireumstriate belt" (see review by Zeki, 1969a), although this area is specifically responsive to visual stimulation (Cowey, 1964; t tubel and Wiesel, 1968; Gross, Rocha-Miranda and Bender, in preparation) besides providing a prominent input to the inferotemporal region.
Two recent developments help to clarify this apparent paradox. The first is the demonstration by Iwai and Mishkin (1968, 1969) tha t lesions confined to the vicinity of the inferior occipital sulcus (and thus in lateroventral prestriate cortex) do produce a severe impairment on visual discrimination learning. The second is tha t the lateroventral prestriate region receives a projection from macular striate cortex (Zeki, 1969b; Cragg and Ainsworth, 1969). Indeed, striate cortex has a three-fold projection onto prestriate cortex and Iwai and Mishkin's lesion included the foveal portions of all three. Iwai and Mishkin's findings are interesting for an- other reason: they found tha t the visual discrimination deficit caused by their lateroventral prestriate lesion (including "foveal prestriate" cortex) was qualita- tively different from tha t following inferotemporal cortical lesions.
In a series of investigations we have studied the contrasting behavioural effects of removing these two non-striate visual areas. In this initial report we present a confirmation and further analysis of Iwai and Mishkin's findings.
Surgical and Histological Procedures
Surgical Procedures
Nineteen experimentally naive Macaca mulatta were used. They weighed 3- -4 kg at surgery. Six animals received inferotemporal lesions; in seven animals the foveal prestriate region and adjacent tissue was removed, and six animals served as unoperated controls.
10"

130 A. Cowey and C.G. Gross:
The inferotemporal lesions ~ were intended to extend rostrally to the anterior limit of area TE as defined by yon Bonin and Bailey (1947), i.e. almost to the tem- poral pole, caudally to about 10 mm anterior to the ascending limb of the inferior occipital sulcus, dorsally to the bottom of the inferior bank of the superior tempo- ral sulcus, and ventrally to the lateral bank of the occipitotemporal sulcus. This lesion corresponded closely to the anterior lesion in Iwai and Mishkin's study (1968).
The prestriate lesions included at least the entire region onto which foveal striate cortex projects. They were intended to extend rostra]ly to about 10 mm in front of the ascending limb of the inferior occipital sulcus, caudally about 5 mm into the anterior bank of the lunate suleus and as far as a line extending this sulcus down to the inferior occipital sulcus, dorsally to the depth of the inferior bank of the superior temporal sulcus, and ventrally to the lateral bank of the occipito- temporal sulcus. They were also intended to include both banks of the inferior occipital suleus in its anterior ascending portion in front of the lunate sulcus, and the inferior bank for 3--4 mm posterior to the tip of the lunate sulcus. This lesion corresponded closely to the posterior lesion in Iwai and Mishkin's study.
All surgery was performed with aseptic procedures and Nembutal anaesthesia. Recovery was uneventful.
Histological Procedures
At the end of the experiments the monkeys were anaesthetized with Nembutal and perfused through the dorsal aorta with 0.9 % saline and then l0 ~ formalin. The head was placed in a stereotaxie machine and the dorsal skull and dura removed. A cut was made in front of and behind the lesion in stereotaxic vertical planes and a steel pin was passed horizontally through each hemisphere, making a small hole which provided a baseline for reconstructing the brain from serial coronal sections. The brain was then removed, photographed and placed in 10% formalin for about 7 days followed by sucrose formalin (10 % formalin, 30 ~ sucrose) until it sank. Fif ty/z frozen sections were cut except in the region of the lateral gcniculate bodies, where 25/~ sections were made. Every tenth section was stained with thionin or cresyl violet. Reconstructions of the lesions were made from sec- tions 1 mm apart, with an examination of additional intermediate sections where necessary.
Histological Results
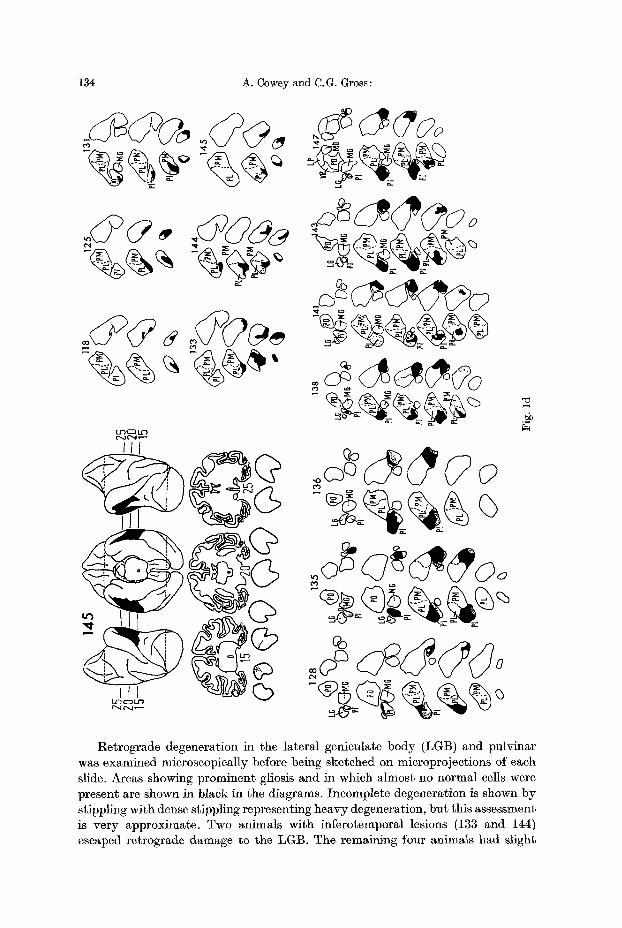
Reconstructions of the surface of the brain, representative coronal sections, and drawings of the lateral genieu]ate bodies and pulvinar are shown in Fig. 1.
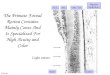
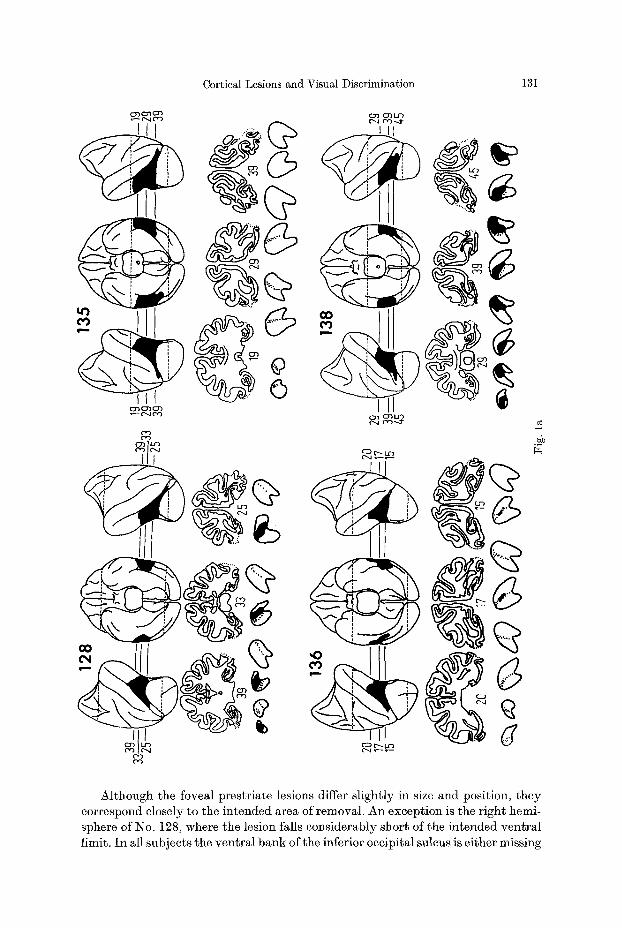
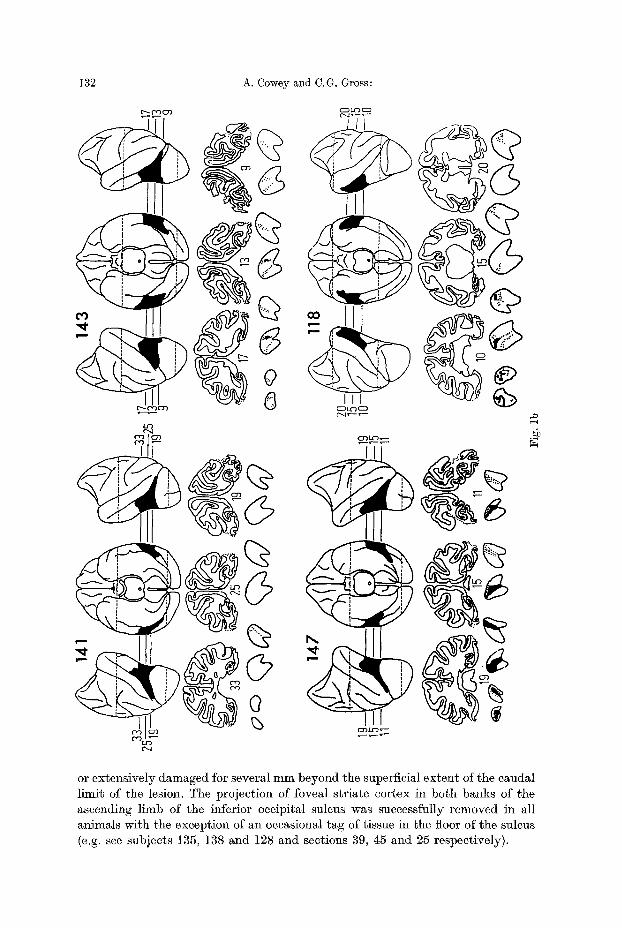
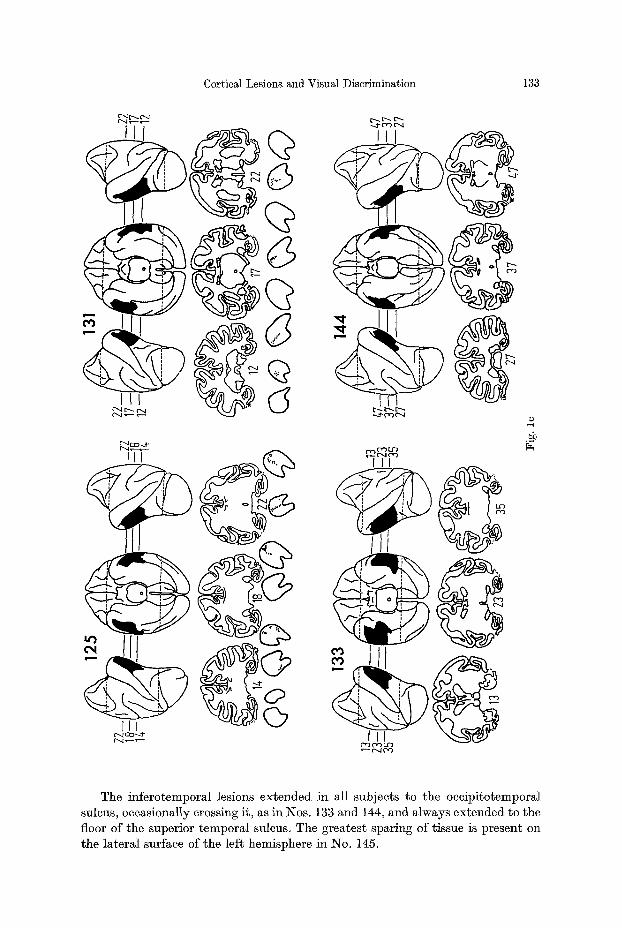
Fig. 1 a--d. Reconstructions of lateral and ventral views of the brains of the experimental animals, together with representative coronal sections. On the reconstructions the lesions are shown in black. On the coronal sections the borders of the lesions are indicated by dotted lines. With the excep- tion of two animals where no degeneration was present in the LGB, sections through the po- sterior three quarters of the LGB are shown immediately beneath the brain sections, with the most caudal sections on the left in each case. Total retrograde degeneration is shown in black, partial degeneration is shown by stippling. Two representative series of sections through the pulvinar are shown in 2d. Labelling of thalamic nuclei: LP ~ lateralis posterior; VPL ~ ven- tralis posterior lateralis; MD ~ medialis dorsalis; MG = medial geniculate; LG ~ lateral genieulate; PO, PI, PL and PiK = pulvinar oralis, inferior, lateralis and medialis respectively
Cortical Lesions and Visual Discrimination 131
O'~ I : ~ ~r3 CD ( :~ I.g3
m m
Although the foveal prestriate lesions differ slightly in size and position, they correspond closely to the intended area of removal. An exception is the right hemi- sphere of No. 128, where the lesion falls considerably short of the intended ventral limit. In all subjects the ventral bank of the inferior occipital sulcus is either missing
132 A. Cowey and C.G. Gross:
g
% c ~
f--4
or extensively damaged for several mm beyond the superficial extent of the caudal limit of the lesion. The projection of foveal striate cortex in both banks of the ascending limb of the inferior occipital sulcus was successfully removed in all animals with the exception of an occasional tag of tissue in the floor of the suleus (e.g. see subjects 135, 138 and 128 and sections 39, 45 and 25 respectively).
Cortical Lesions and Visual Discrimination 133
d ~
m f,e.) 1..~
The inferotemporal lesions extended in all subjects to the oceipitotemporal sulcus, occasionally crossing it, as in Nos. 133 and 144, and always extended to the floor of the superior temporal sulcus. The greatest sparing of tissue is present on the lateral surface of the left hemisphere in No. 145.
134 A. Cowey and C.G. Gross:
L r ~ O t / ' )
Ill
t f ~ D t ~
e o
oo
~0
Retrograde degeneration in the lateral geniculate body (LGB) and pulvinar was examined microscopically before being sketched on microprojections of each slide. Areas showing prominent gliosis and in which almost no normal cells were present are shown in black in the diagrams. Incomplete degeneration is shown by stippling with dense stippling representing heavy degeneration, but this assessment is very approximate. Two animals with inferotemporal lesions (133 and 144) escaped retrograde damage to the LGB. The remaining four animals had slight

Cortical Lesions and Visual Discrimination 135
cell loss which was b i l a te ra l in two of t hem (118 and 145). Because of the pro- x i m i t y of the v isual rad ia t ions to the foveal p res t r i a te cortex, the re t rograde degenera t ion was more severe in the LGB of this group. Three had p rominen t bi- l a t e ra l degenera t ion (128, 138 and 147); the remain ing four had a cell loss roughly equiva len t to t h a t p resen t in the in fe ro tempora l group. The relevance of this degenera t ion to the observed visual d i sc r imina t ion deficits is considered la ter .
Re t rog rade degenera t ion in the pu lv ina r was clear wi th conspicuous gliosis and a lmos t t o t a l absence of cells in the affected regions. I t was p lo t t ed wi th respect to the divisions of the pu lv ina r p resen ted b y Olszewski (1952). I n the infcrotempo- ral group the degenera t ion was confined to the ven t ra l aspect of the caudal t h i rd of the pulv inar , t h a t is, i t fell en t i re ly wi th in the caudal por t ion of pu lv ina r la tcra l is (see Fig. ld ) . The degenera t ion in animals wi th foveal p res t r i a te lesions was more extens ive b u t was confined to the inferior pulv inar , wi th poss ib ly a sl ight involve- m e n t of the la te ra l pu lv ina r in the region of the b o u n d a r y between the two divi- sions. The mos t ros t ra l t ip of the infer ior pulv inar , ly ing i m m e d i a t e l y benea th the caudal pole of the la te ra l genicula te body, was e i ther comple te ly spared (Nos. 136, 141, 128, 143) or only s l ight ly invo lved (Nos. 147, 138 and 135), bu t from t h a t po in t to i ts caudM l imi t the inferior pu lv ina r was ex tens ive ly degenera ted (80--90 per cent) except in No. 128 in the r ight hemisphere where only abou t 20% of the inferior pu lv ina r was affected. This spar ing re la tes well to the incomple te cort ical lesion in the same hemisphere .
Experiment 1
The three groups of an imals ( inferotemporal , foveal p res t r i a te and normal con- trols) were t e s ted for r e ten t ion of a s imul taneous pa t t e rn d iscr imina t ion acqui red pr ior to surgery.
Methods Apparatus
The animals were trained in a sound insulated box. Two translucent Gerbrands keys 1.1/s inches in diameter and 12 inches apart were mounted in one wall of the box. The dis- criminanda were back projected onto the keys by rear-projection readout units (series 10 In- dustrial Electronic Engineers, Inc.), modified so that the keys were in the plane of the image of the stimulus. A liquid dispenser was mounted in the centre of the same wall 12 inches below a line connecting the two keys. The interior of the box was illuminated from above and a mask- ing noise was present. Stimulus presentation and reinforcement were programmed automatic- ally.
Procedures The monkeys were tested when water deprived and for 100 trials per day, seven days each
week. On all tasks, the spatial position of the stimuli was randomized. The inter-trial interval was 5 see and any press during the inter-trial interval reset the interval. Response to either or both keys turned the stimuli off. A correct response was followed by presentation of .25 ce. orange juice. Simultaneous presses on both keys counted as an error. Correction trials were not used in this or in subsequent experiments.
Preliminary Training. The animals were first trained to respond to the keys when illu- minated with a white square (Grason-Stadler stimulus 156-4), and then were trained to dis- criminate the white square (positive) from an unlighted blank key to a criterion of 45 correct responses in 50 trials.
Experimental Training. The animals were trained to discriminate two horizontal white lines (positive) from two vertical ones (Grason-Stadler 154-5, 6) to a criterion of 90 correct responses in 100 trials on each of two successive days. On the basis of their performance on this
136 A. Cowcy and C.G. Gross:
t a sk t h e y were d iv ided into th ree m a t c h e d groups and, af ter th ree days w i t h o u t wa te r depri- va t ion , t he expe r imen t a l g roups received the i r lesions. F o u r t e e n days la te r all an ima l s were retrained to the pre-operative criterion unless they failed to relearn in 1,500 trials, in which case testing was stopped.
Results
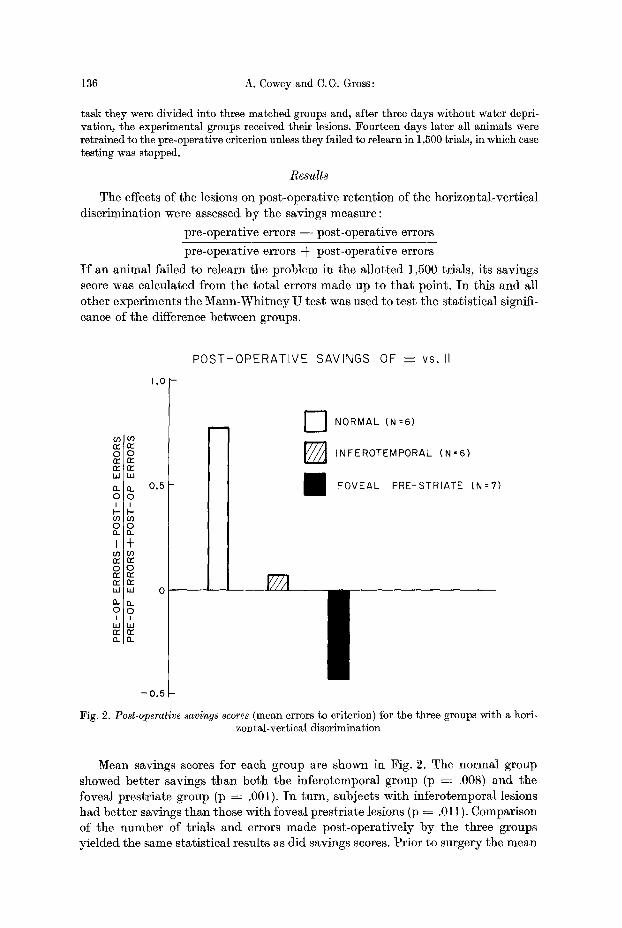
The effects of the lesions on pos t -opera t ive re ten t ion of the hor izonta l -ver t ica l d i sc r imina t ion were assessed b y the savings measure :
p re -opera t ive errors - - pos t -opera t ive errors
p re -opera t ive errors -k pos t -opera t ive errors
I f an an imal fai led to re learn the p rob lem in the a l lo t t ed 1,500 tr ials , i ts savings score was ca lcula ted f rom the t o t a l errors made up to t h a t point . I n th is and all o ther exper iments the M a n n - W h i t n e y U tes t was used to t es t the s ta t i s t ica l signifi- cance of the difference be tween groups.
1,0
n--
oo re
rr I~ 1,1 UJ
a- a. 0.5
o 0 I I
a . i~_
I +
0 0
w u 0 n ~
0 0 I I
w w 0.. n
-0.5
POST-OPERATIVE SAVINGS OF ---- vs. ll
rl IJ
D NORMAL (N=6)
F ~ INFEROTEMPORAL (N=6)
FOVEAL PRE-STRIATE (N=7)
Fig. 2. Post-operative savings scores (mean errors to criterion) for the three groups with a hori- zontal-vertical discrimination
Mean savings scores ibr each group are shown in Fig. 2. The norma l group showed be t t e r savings t h a n bo th the in fe ro tempora l group (p ---- .008) and the foveal p res t r i a t e group (p ~ .001). I n turn , subjects wi th in fe ro tempora l lesions had be t t e r savings t h a n those wi th foveal p res t r i a te lesions (p ~ .011). Comparison of the number of t r ia ls and errors made pos t -ope ra t ive ly b y the three groups y ie lded the same s ta t i s t ica l resul ts as d id savings scores. Pr io r to surgery the mean

Cortical Lesions and Visual Discrimination 137
trials and errors required to reach criterion were 146 and 333 respectively for the normal controls, 148 and 343 for foveal prestriates, 162 and 350 for infcrotempo- rals.
Animal No. 128, which had an incomplete foveal prestriate lesion and slight pulvinar degeneration on the r ight side, was much less impaired than the other subjects in its group with savings of .09 compared to a mean o f - - . 4 2 for the re- maining subjects.
Experiment 2
Exper iment 1 confirmed the previous finding (Iwai and Mishkin, 1968) t ha t post-operative retention of a fairly difficult pa t te rn discrimination is worse follow- ing foveal prestriate than inferotemporal lesions. I n Exper iment 2 we investigated whether the foveal prestriate group was also more impaired on the post-operative acquisition of similar visual tasks. The animals were tested successively on three pa t te rn discrimination problems.
Methods Subjects and Apparatus
These were the same as in Experiment 1.
Procedures Problem a was the discrimination of �9 versus ~ (Grason-Stadler 150-1, 2), problem b of
W versus T (Grason-Stadler 159-3, 4), and problem c of ~-O-~ versus @, positive stimulus indicated first in each case. Procedures were identical to Experiment 1 except that testing on problem a was terminated after 2,000 trials if the animal had not reached criterion by that time, and problems b and c were each terminated after a maximum of 4,000 trials. The problems were given in the order a, b, c.
Results
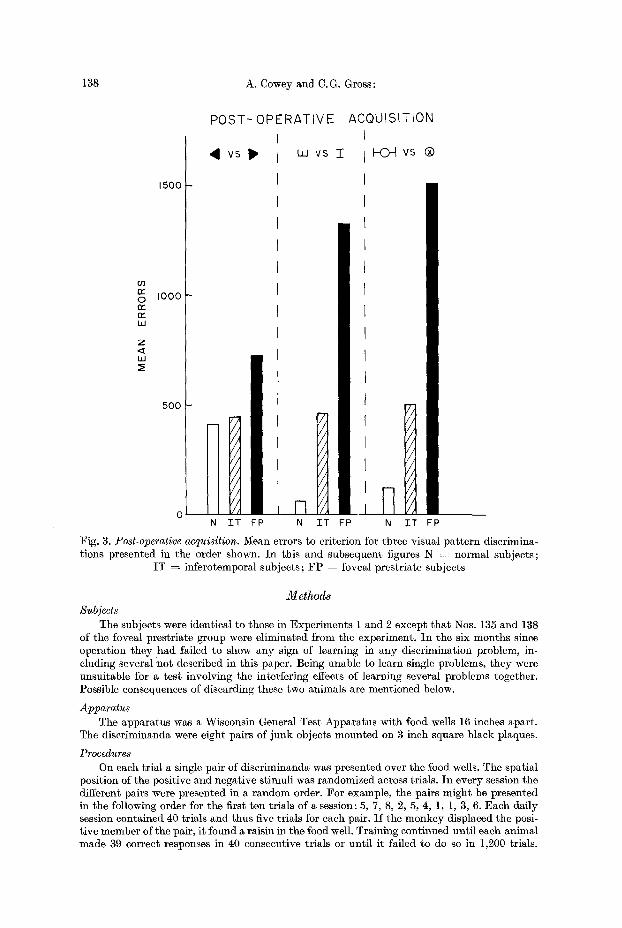
As can be seen from Fig. 3, on all three problems the normal subjects made fewer errors than either operated group, and the inferotemporal group made fewer errors than the foveal prcstriate group. The superiori ty of the normal group to the foveal prestriate group reached statistical significance on all problems (a, p = .037 ; b, p = .002 ; e, p = .00]). The normal group was significantly superior to the infero- temporal group on two of the problems (a, p = .35; b, p = .02; c, p = .013). Finally, the inferotemporal group was significantly superior to the foveal pre- striate group on two of the three problems (a, p = .05; b, p ~ .069; e, p = .011). Animal No. 128, with an incomplete foveal prestriate lesion, was again superior to all other animals in t ha t group (a, 236 errors; b, 59 errors; e, 288 errors).
I n summary, the foveal prestriate group was more impaired than the anterior inferotemporal group not only on post-operative retent ion of a visual discrimina- tion (Experiment 1), but also on the post-operat ive acquisition of new discrimina- t ion tasks (Experiment 2).
Experiment 3
The groups were now tested on a ra ther different type of visual discrimination task in which eight different and relatively simple object discriminations are pre- sented intermingled in each testing session, such tha t the animal is learning several visual problems in a single session. This paradigm has been called a "concurrent discrimination" (Iwai and Mishkin, 1968).
138 A. Cowey and C.G. Gross:
P O S T - O P E R A T I V E ACQUIS IT ION
(./3 n - O rr r~" W
Z
I..I.I z~
1500
IO00
500
~1 vs Ip,
I I
/ 2
I I / / N TT
L U v s I
II ,nH FP N TT FP
I-K~q vs |
7, / /
/ 1 / ]
/ /
/ /
/ /
/ /
N T T FP
Fig. 3. Post-operative acquisition. Mean errors to criterion for three visual pa t t e rn discrimina- tions presented in the order shown. In this and subsequent figures N = normal subjects;
IT = inferotemporal subjects; F P = foveal prestriate subjects
Methods Subjects
The subjects were identical to those in Experiments ] and 2 except t ha t Nos. 135 and 138 of the foveal prestriate group were eliminated from the experiment. In the six months since operation they had failed to show any sign of learning in any discrimination problem, in- eluding several ho t described in this paper. Being unable to learn single problems, they were unsuitable for a test involving the interfering effects of learning several problems together. Possible consequences of discarding these two animals are mentioned below.
Apparatus The apparatus was a Wisconsin General Test Apparatus with food wells 16 inches apart .
The discriminanda were eight pairs of junk objects mounted on 3 inch square black plaques.
Procedures On each trial a single pair of discriminanda was presented over the food wells. The spatial
position of the positive and negative stimuli was randomized across trials. In every session the different pairs were presented in a random order. :For example, the pairs might be presented in the following order for the first ten trials of a session: 5, 7, 8, 2, 5, 4, 1, 1, 3, 6. Each daffy session contained 40 trials and thus five trims for each pair. I f the monkey displaced the posi- t ive member of the pair, i t found a raisin in the food well. Training continued until each animal made 39 correct responses in 40 consecutive trials or unti l it failed to do so in 1,200 trials.
Cortical Lesions and Visual Discrimination
CONCURRENT DISCRIMINATION I
139
O0 rY 0 n - r~ L~
5 0 0
25O LU
M N
777 / / / / / t / / /
; / / , / / / ' / / /
"rT
/ FP
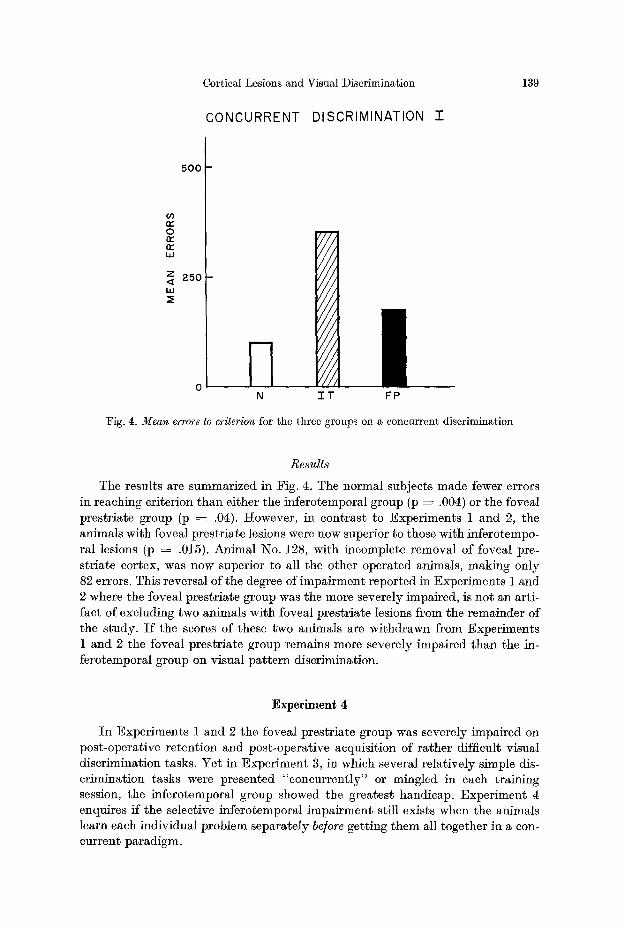
Fig. 4. Mean errors to criterion for the three groups on a concurrent discrimination
Resu l t s
The results are summarized in Fig. 4. The normal subjects made fewer errors in reaching criterion than either the inferotemporal group (p ---- .004) or the foveal prestriate group (p ~ .04). However, in contrast to Experiments 1 and 2, the animals with foveal prestriate lesions were now superior to those with inferotempo- ral lesions (p -~ .015). Animal No. 128, with incomplete removal of foveal pre- striate cortex, was now superior to all the other operated animals, making only 82 errors. This reversal of the degree of impairment reported in Experiments 1 and 2 where the foveal prestriate group was the more severely impaired, is not an arti- fact of excluding two animals with foveal prestriate lesions from the remainder of the study. Is the scores of these two animals are withdrawn from Experiments 1 and 2 the foveal prestriate group remains more severely impaired than the in- ferotemporal group on visual pat tern discrimination.
Experimen~ 4
In Experiments 1 and 2 the foveal prestriate group was severely impaired on post-operative retention and post-operative acquisition of rather difficult visual discrimination tasks. Yet in Experiment 3, in which several relatively simple dis- crimination tasks were presented "concurrently" or mingled in each training session, the inferotemporal group showed the greatest handicap. Experiment 4 enquires ff the selective inferotemporal impairment still exists when the animals learn each individual problem separately be]ore getting them all together in a con- current paradigm.

140 A. Cowey and C.G. Gross:
Method8 Subjects and Apparatus
Subjects were the same as in Experiment 3 except for the absence of No. 133, an infero- temporal animal which died, leaving 6 normal subjects, 5 inferotemporal subjects and 5 foveal prestriate subjects.
The apparatus was the same Wisconsin General Test Apparatus used in Experiment 3. The discriminanda were ten new pairs of junk objects mounted on 3 inch square black plaques.
Procedures Stage a. Animals were trained for 50 trials per day to discriminate the first pair of stimuli
to a criterion of 45 correct responses in 50 consecutive trials. The day following criterion they began training on the second pair with the same procedures. Similarly, they were then trained on a third, fourth and fifth pair of diseriminanda. Each experimental group was divided into 2 sub-groups, one being given pairs 1--5, and the other pairs 6--10, in that order.
Stage b. The subjects were tested for retention of the five individual problems, to ensure that they still knew them individually before receiving them concurrently in the next stage. They were tested for 50 trials per day and given one pair per day until the original criterion was reached for all pairs on five successive days.
Stage c. In this stage, the five pairs of objects learned by each animal in Stages a and b were presented in concurrent form (as described in Experiment 3), each pair appearing 10 times in a random order in each daily testing session of 50 trials. Animals were trained to a criterion of 49 correct responses in any 50 consecutive trials.
Stage d. The day after completing Stage c, the animals were given a concurrent discrimi- nation consisting of the five object pairs previously presented in Stages a, b and c, together with the five pairs of stimuli each sub-group had never seen. Subjects that had learned pairs 1--5 or pairs 6--10 now received pairs 1--10. The monkeys were trained to a criterion of 49 correct responses in 50 consecutive trials. Each pair of objects appeared five times in 50 daily trials. Testing was stopped if an animal failed to reach criterion in 1,200 trials.
Results
Stage a. There were no significant differences among the groups in the total numbers of errors on the five problems. The median tota l error score for the normal animals was 46, for the inferotemporals 79, and for the foveal prestriates 66. The groups also did no t differ significantly on any individual problem.
Stage b. All groups showed excellent re ten t ion of the indiv idual problems;
each group had a median error score of zero. Similarly, in Stage c, presenting all five problems concurrent ly in a single
session did no t alter the performance of any group. The median errors scores for the normal , inferotcmporal, and foveal prestr iate groups were, respectively 0, 5
and 0. The effects of presenting the five previously learned discriminations concur-
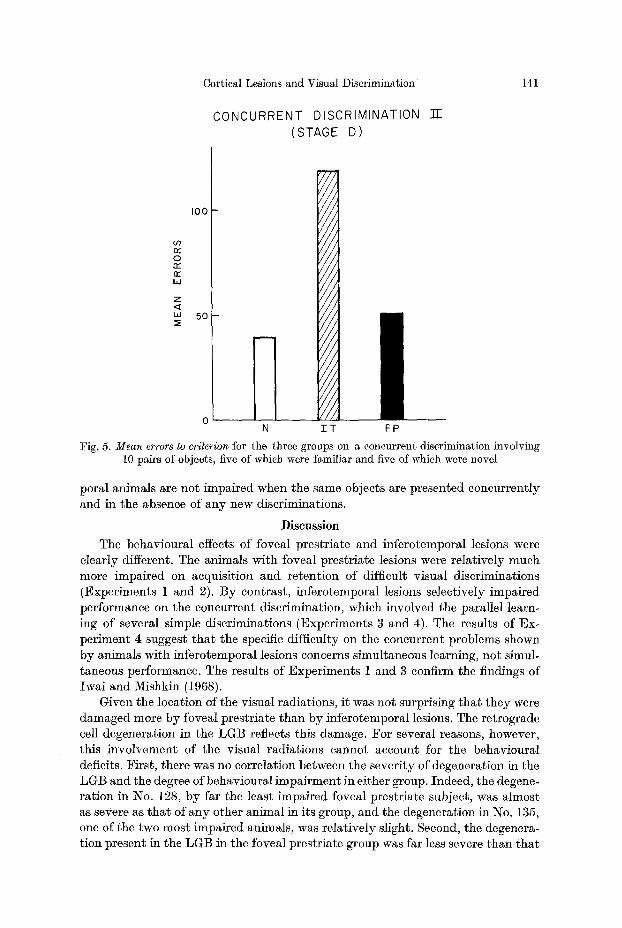
rent ly with the five new problems in Stage d are shown in Fig. 5. The inferotempo- ral group was markedly impaired relative to bo th the normal group (p ---- .004) and the foveai prestriate group (p ~- .016) as had been the case in the previous con- current acquisi t ion task (Exper iment 3). Indeed, the slightly poorer performance of the foveal prestriate group relative to the normal controls did no t reach the .05 level of significance. All the groups made some errors on the indiv idual discrimina- tions which they had previously learned in Stages a, b and c, bu t every group made more errors on the new problems t h a n on the old (p -~ .02) for M1 groups.
I n summary, the inferotemporal animals are more impaired t h a n the foveal prestriate animals in learning object discr iminat ions presented in a concurrent paradigm. However, ff the objects are previously learned individual ly , infcrotem-
Cortical Lesions and Visual Discrimination
CONCURRENT DISCRIMINATION (STAGE D)
"O-
141
0 n," n"-
W
Z
U.I
lO0
50
, i l l
r
i / / J
, l l J
I / / j
/ / / , / / / / r
t / / ,
7/ /
N ET FP
Fig. 5. M e a n errors to criterion for the three groups on a concurrent discrimination involving 10 pairs of objects, five of which were familiar and five of which were novel
poral animals are not impaired when the same objects are presented concurrently and in the absence of any new discriminations.
Discussion
The behavioural effects of foveal prestriate and inferotemporal lesions were clearly different. The animals with foveal prestriate lesions were relatively much more impaired on acquisition and retention of difficult visual discriminations (Experiments 1 and 2). By contrast, inferotemporal lesions selectively impaired performance on the concurrent discrimination, which involved the parallel learn- ing of several simple discriminations (Experiments 3 and 4). The results of Ex- periment 4 suggest tha t the specific difficulty on the concurrent problems shown by animals with inferotemporal lesions concerns simultaneous learning, not simul- taneous performance. The results of Experiments 1 and 3 confirm the findings of Iwai and Mishkin (1968).
Given the location of the visual radiations, it was not surprising that they were damaged more by foveal prestriate than by inferotemporal lesions. The retrograde cell degeneration in the LGB reflects this damage. For several reasons, however, this involvement of the visual radiations cannot account for the behavioural deficits. First, there was no correlation between the severity of degeneration in the LGB and the degree of behavioural impairment in either group. Indeed, the degene- ration in No. 128, by far the least impaired foveal prestriate subject, was almost as severe as tha t of any other animal in its group, and the degeneration in No. 135, one of the two most impaired animals, was relatively slight. Second, the degenera- tion present in the LGB in the foveal prestriate group was far less severe than tha t

142 A. Cowey and C.G. Gross:
following lateral striate lesions, which have slight, if any, effect, on pattern dis- criminations similar to those of the present study (Wilson and Mishkin, 1959; Butter and Gekoski, 1966; Cowey and Weiskrantz, 1967).
By contrast, the retrograde cell degeneration in the inferior pulvinar did corre- late with the extent of the behavioural impairment in the foveal prestriate group. Animal 128 was superior to all other animals in that group on every discrimination, and the degeneration in the right inferior pulvinar was substantially less than in any other animal. Animals 135, 138 and 147 were consistently poorer than the other four animals of their group and only they showed degeneration in the most rostral portion of the inferior pulvinar (see Fig. 1, d).
The lesion of animal No. 128, the least impaired foveal prestriate subject, was interesting for another reason. I t spared the ventral portion of the inferior occipital gyrus in the right hemisphere as well as the depth of the inferior occipital sulcus on the left side (see Fig. 2, a). I t is precisely one or the other or both of these areas that were spared in all the previous investigations in which little or no deficit on visual discrimination tasks was found after prestriate or circumstriate lesions (Lashley, 1948; Chow, 1951; l~iopelle et al., 1951; Evarts, 1952; Pribram et al., 1969). Furthermore, both areas were included in the prestriatc lesions of Iwai and Mishkin (1968, 1969), which severely impaired visual pattern discrimination. A possible explanation for this critical role of the inferior occipital sulcus and the in- ferior occipital gyrus is provided by the recent anatomical studies of Zeki (1969b) and Cragg and Ainsworth (1969). Both found that the region of striate cortex onto which the fovea projects sends fibres to the ventral prehinate gyms, both banks of the tip of the lunate suleus, the inferior bank of the superior temporal sulcus, the inferior occipital gyrns and the banks of the ascending portion of the inferior oeci- pital sulcus. This entire region was removed in all subjects of our foveal prestriate group except for )~o. 128, whereas it was partially spared in this subject and in the studies mentioned above.
Why should foveal prestriate cortex be crucial for pattern discrimination learn- ing, when foveal striate cortex is not ? When a monkey or man has no foveal striate cortex, it uses the extra-foveal part of its retina for visually-guided behaviour by fixating eccentrically (Cowey, 1967; Cowey and Weiskrantz, 1963; Teuber et al., 1960). Thus, the visual information is processed by extra-foveal striate cortex and then presumably by extra-foveal prestriate cortex. However, if a monkey has no foveal prestriate cortex it may nevertheless continue to fixate foveally. I f so, in- formation arriving at foveal striate cortex could not be processed in the missing prestriate cortex to which it normally projects, thereby resulting in a pattern dis- crimination deficit. This explanation assumes (a) that after removing foveal pre- striate cortex, there is no functional alteration in the projection of striate onto prestriate cortex and that information from foveal striate cortex therefore no lon- ger reaches the prestriate area, and (b) that monkeys with foveal prestriate lesions continue to fixate foveally. Both assumptions are reasonable and testable. I t also follows that the ablating foveal striate cortex in an animal with an existing foveal prestriate lesion should improve pattern discrimination by making him fixate eccentrically and so use extra-foveal striate and prestriate cortex. This also is test- able although it is probably best done by removing foveal striate and foveal pre- striate together, in which case no impairment would be predicted.

Cortical Lesions and Visual Discrimination 143
What could be the functions of foveal prestriate cortex and inferotemporal cortex such that removing them produces two different behavioural disorders ? Behavioural experiments provide a tentative suggestion. Many studies of dis- crimination learning in normal animals indicate that discrimination learning in- volves two distinct stages, an attentional or perceptual stage and an associative or mnemonic stage (Krechevsky, 1938 ; Sutherland, 1959, 1964; Mackintosh, 1965; Lovejoy, 1968; Bower and Trebasso, 1968). Perhaps inferotemporal lesions chiefly disturb the associative stage since their maximum effect was on the concurrent discrimination in which proactive and retroactive interference among the problems must have been high. On the other hand, foveal prestriate lesions may impair an attentional or perceptual mechanism which is necessary for detecting small differ- ences between the pairs of patterns used in Experiments 1 and 2. I t is interesting in this connexion that in Experiment 2 the normal subjects found the three pattern discriminations increasingly easy whereas the foveal prestriate subjects found them increasingly difficult (see Fig. 3). The complexity of the patterns (as measured by number, orientation, and intersection of lines) also increased from pair 1 to pair 3.
A simpler interpretation of the deficit produced by foveal prestriate lesions might be that it is a "sensory deficit" rather than an impairment in some percep- tual, attentional or other complex process. We know that this is an unlikely possibility for the inferotemporal lesions. Inferotemporal lesions do not impair visual acuity or the integrity of the visual fields (Cowey and Weiskrantz, 1967), or thresholds for brightness discrimination (Ettlinger, 1959), or detection of a brief visual stimulus (D. B. Bender, unpublished observations). Some preliminary results suggest that foveal prestriate lesions also do not alter thresholds for detecting brief visual stimuli (D.B. Bender, unpublished observations). Interpreting the visual impairment in terms of a sensory loss is also unsatisfactory in view of the fact that ablating the macular projection area in striate cortex has negligible effects on visual pattern discrimination despite producing field defects. However, more studies are required to establish that neither lesion produces changes in psychophysical functions.
Several previous investigators have debated whether the "classical" infero- temporal lesion (which usually included all of our hfferotemporal lesion but often the anterior part of our foveal prestriate lesion) produced impairments in "visual at tent ion" or in "visual learning". As Iwai and Mishkin (1968) pointed out, these large inferotemporal lesions may produce both types of impairment, which may be dissoeiable by the smaller lesions of their study and of ours. We are now in the process of further testing this hypothesis that foveal prestriate lesions primarily interfere with the first or attentional/perceptual stage of discrimination learning and that inferotemporal lesions primarily interfere with the second or associative/ mnemonic stage.
References
Bower, G.H., Trebasso, T. : Attention in learning. New York: Wiley 1968. Butter, C.M., Gekoski, W. L. : Alterations in pattern equivalence following inferotemporal and
lateral striate lesions in rhesus monkeys. J. eomp. physiol. Psychol. 61, 309--312 (1966). Chow, K. L. : Effects of partial extirpation of posterior association cortex on visually mediated
behavior in monkeys. Comp. Psyehol. Monogr. 20, 187~217 (1951).
10a Exp. Brain l~es. Vol. 11

144 A. Cowey and C.G. Gross: Cortical Lesions and Visual Discrimination
Cowey, A. : Projection of the retina onto striate and prestriate cortex in the squirrel monkey, Saimiri sciureus. J. Neurophysiol. 27, 366--393 (1964).
- - Perimetric study of visual field defects in monkeys after cortical and retinal ablations. Quart. J. exp. Psychol. 19, 232--245 (1967).
- - Weiskrantz, L. : A perimetric study of visual field defects in monkeys. Quart. J. exp. Psyehol. 1 5 , 91--115 (1963).
- - - - A comparison of the effects of inferotemporal and striate cortex lesions on the visual behaviour of rhesus monkeys. Quart. J. exp. Psychol. 19, 246--253 (1967).
Cragg, B.G., Ainsworth, A.: The topography of the afferent projections in the circumstriate visuM cortex of the monkey studied by the Nauta method. Vision Res. 9, 737--747 (1969).
Ettlinger, G. : Visual discrimination with a single manipulandum following temporal ablations in the monkey. Quart. J. exp. Psychol. 11, i64--174 (i959).
Evarts, E.V. : Effects of ablation of prestriate cortex on auditory-visuM association in monkey. J. Neurophysiol. 15, 191--200 (1952).
Gross, C.G. : Visual functions of inferotemporal cortex. In: Handbook of Sensory Physiology, Vol. 7, Par t 3. Ed. by R. Jung. Berlin-Heidelberg-New York: Springer I970 [In press].
- - Bender, D.B., Rocha-l~Iiranda, C.E. : Visual receptive fields of neurons in inferotemporM cortex of the monkey. Science 166, 1303--1305 (1969).
Hubel, D.H., Wiesel, T.N.: Receptive fields and functional architecture of monkey striate cortex. J. Physiol. (Lond.) 195, 215--243 (1968).
Iwai, E., Mishkin, M.: Two visual foei in the temporal lobe of monkey. Japan-U.S. Joint Seminar on Neurophysiological Basis of Learning and Behavior. Kyoto, Japan, 1968.
- - - - Further evidence on the locus of the visual area in the temporal lobe of the monkey. Exp. Neurol. 25, 585--.594 (1969).
Krechevski, I. : A study of the continuity of the problem-solving process. Psychol. Rev. 45, 107--133 (1938).
Kuypers, It . G. J.M., Szwarebart, M. A., lVIishkin, M., Rosvold, H. E. : OccipitotemporM eortieo- cortical connections in the rhesus monkey. Exp. Neurol. 11, 24~5--262 (1965).
Lashley, K. S. : The mechanism of vision. XVIII . Effects of destroying the visual 'associative areas' of the monkey. Genet. Psychol. lV[onogr. 37, 107--166 (1948).
Lovejoy, E. : Attention in discrimination learning. London: Holden-Day 1968. Mackintosh, N.J . : Selective attention in animal discrimination learning. Psychol. Bull. 64,
124-150 (1965). Mishkin, M.: Visual mechanisms beyond the striate cortex, pp. 93--119. In: Frontiers in
physiological psychology. Ed. by R.W. Russell. New York: Academic Press 1966. Olszewski, J. : The thalamus of )s mulatta. Basel, Switzerland: Karger 1952. Pribram, K.H. , Spinelli, D.N., Reitz, S. L. : The effects of radical disconnexion of occipital and
temporal cortex on visual behaviour of monkeys. Brain 92, 301--312 (1969). Riopelle, A.J . , Harlow, H.F. , Settlage, P.H., Ades, H.W.: Performance of normal and
operated monkeys on visual learning tests. J. comp. physiol. Psychol. 44, 283--289 (1951). Sutherland, N.S.: Stimulus analyzing mechanisms, pp. 575--609. In: Proceedings of a sym-
posium on the mechanism of thought processes. London: Her Majesty's Stationery Office i959.
- - The learning of discriminations by animals. Endeavour 23, 148--152 (1964). Teuber, H.-L., Battersby, W.S., Bender, M.B. : Visual field defects after penetrating missile
wounds of the brain. Cambridge/Mass. : Harvard University Press 1960. Von Bonin, G., Bailey, P. : The neocortex of Maeaca mulatta. Urbana/Ill. : University of
Illinois Press 1947. Wilson, W.A., Mishkin, M. : Comparison of the effects of inferotemporal and lateral occipital
lesion on visually guided behavior in monkeys. J. comp. physiol. Psychol. 52, 10--17 (1959). Zeki, S.M. : The secondary visual areas of the monkey. Brain Res. 13, 197--226 (1969a). - - Representation of central visual fields in prestriate cortex of monkey. Brain Res. 14,
271--291 (1969b).
Dr A. Cowey Institute of Experimental Psychology 1 South Parks Road Oxford (England)