Embed Size (px)
Citation preview

INSTITUTO POLITECNICO NACIONAL
CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
EL PAPEL DEL MERO ROJO (Epinephelus morio,
VALENCIENNES, 1828) EN EL ECOSISTEMA DE
PLATAFORMA CONTINENTAL DE YUCATÁN,
MÉXICO
QUE PARA OBTENER EL GRADO DE
MAESTRÍA EN CIENCIAS
EN
MANEJO DE RECURSOS MARINOS
PRESENTA:
BIOL. MAR. ANIA YARAZETH CHAMÚ MARTÍNEZ
La Paz, Baja California Sur,



Dedicatoria
A la bella familia que me tocó coincidir Chamú-Martínez.

Agradecimientos
A mi director de tesis Dr. Francisco Arreguín Sánchez, por su respaldo en el desarrollo de
esta tesis y por su predisposición siempre amena a entablar espacios de discusión, que me
permitieron desarrollar una visión más amplia sobre la forma de abordar los procesos
ecosistémicos.
Agradezco también las observaciones y de la comisión de evaluación de profesores: Dr.
Manuel Zetina Rejón, Pablo Del Monte Luna, Arturo Tripp Valdez y Germán Ponce Díaz.
A Juan Carlos Hernández Padilla por su gran apoyo profesional y personal.
A CONACyT por la beca otorgada para la realización de mis estudios de maestría.
Al Programa Institucional de Formación de Investigadores del Instituto Politécnico Nacional
(BEIFI -IPN) por el apoyo económico otorgado.
A todos mis compañeros de generación 2014-2016, gracias por los momentos compartidos.

Índice
Lista de figuras ...................................................................................................................... i
Lista de tablas ....................................................................................................................... ii
Glosario ................................................................................................................................ iv
Resumen ............................................................................................................................... vi
Abstract ............................................................................................................................... vii
1. Introducción ................................................................................................................... 1
2. Antecedentes ..................................................................................................................... 2
3. Justificación ....................................................................................................................... 4
4. Hipótesis ............................................................................................................................ 4
5. Objetivo general ................................................................................................................ 5
5. 1. Objetivos específicos ................................................................................................. 5
6. Área de estudio ................................................................................................................. 5
7. Materiales y métodos ........................................................................................................ 7
7.1 Descripción del modelo Ecopath ............................................................................... 7
7.2 Construcción del modelo ............................................................................................ 8
7.2.1 Estimación de los datos de entrada ................................................................... 11
7.2.2 Biomasa (B) ......................................................................................................... 11
7.2.3 Producción biomasa (P/B) ................................................................................. 11
7.2.4 Consumo/biomasa (Q/B) .................................................................................... 13
7.2.5 Flotas pesqueras y capturas (Yi) ....................................................................... 17
7.2.6 Eficiencia Ecotrófica (EEi) ................................................................................ 18
7.2.7 Matriz de predador/presa .................................................................................. 19
7.3 Incertidumbre del modelo ........................................................................................ 22
7.4 Atributos tróficos de los grupos funcionales .......................................................... 22

7.4.1 Nivel trófico ......................................................................................................... 22
7.4.2 Índice de omnivoría ............................................................................................ 23
7.5 Atributos del ecosistema ........................................................................................... 23
7.5.1 Flujos del sistema ............................................................................................... 23
7.5.2 Índice de conectividad ........................................................................................ 24
7.5.3 Impactos tróficos ................................................................................................ 24
7.5.4 Indicadores de crecimiento y desarrollo .......................................................... 24
7.6 Indicadores estructurales ......................................................................................... 26
7.7 Simulación temporal ................................................................................................. 27
7.8 Interpretación del papel del mero ........................................................................... 28
8. Resultados ....................................................................................................................... 29
8.1 Construcción del modelo: estimación de los datos de entrada ............................. 29
8.2 Balanceo del modelo ................................................................................................. 40
8.3 Atributos de los grupos funcionales ........................................................................ 41
8.4 Atributos del ecosistema ........................................................................................... 43
8.4.1 Flujos del sistema ............................................................................................... 43
8.4.2 Índice de conectividad ........................................................................................ 45
8.4.3 Impactos tróficos mixtos .................................................................................... 46
8.5 Indicadores estructurales ......................................................................................... 48
8.6 Simulación temporal ................................................................................................. 51
8.7 Interpretación del papel del mero ........................................................................... 52
9. Discusión .......................................................................................................................... 62
9. 1 Construcción del modelo: estimación de los datos de entrada ............................ 62
9.2 Balanceo del modelo ................................................................................................. 63
9.3 Atributos de los grupos funcionales ........................................................................ 65

9.4 Atributos del ecosistema ........................................................................................... 65
9.5 Indicadores de crecimiento y desarrollo ................................................................. 68
9.6 Indicadores estructurales ......................................................................................... 68
9.7 Simulación temporal ................................................................................................. 69
9.8 Interpretación del papel del mero ........................................................................... 70
10. Conclusión ..................................................................................................................... 73
11. Refencias ........................................................................................................................ 74
12. Anexos ............................................................................................................................ 87

i
Lista de figuras
Figura 1. Área de estudio, plataforma continental de Yucatán ............................................. 6
Figura 2. Representación de la estructura trófica y biomasa de los grupos funcionales de la
plataforma continental de Yucatán. Cada círculo corresponde a un grupo funcional y su
tamaño, es proporcional a su biomasa. Las líneas representan las relaciones predador- presa.
.............................................................................................................................................. 29
Figura 3. Tendencias de los cocientes producción/consumo (P/Q) y respiración/asimilación
(R/A), con respecto al nivel trófico de los grupos funcionales ............................................ 40
Figura 4. Impactos tróficos entre los grupos funcionales y flotas pesqueras del ecosistema
de la plataforma continental de Yucatán, México ................................................................ 47
Figura 5. Gráfica de índice de grado de la plataforma continental de Yucatán; México .... 49
Figura 6. Gráfica de índice de cercanía de la plataforma continental de Yucatán, México 49
Figura 7. Gráfica de índice de intermediación de la plataforma continental de Yucatán,
México .................................................................................................................................. 50
Figura 8. Tendencia de la anomalía de temperatura superficial del mar expresada por la
Oscilación Multidecadal del Atlántico. ................................................................................ 51
Figura 9. Tendencias entre el orden y biomasa, y variaciones de la biomasa a través del
tiempo. a) mero, b) sierra, c) corvina y d) pastos marinos. Solo se representaron un grupo
por cada patrón encontrad. Las flechas indican el año de inicio de la serie de tempo. ........ 54

ii
Lista de tablas
Tabla 1. Familias y/o especies incluidas en cada grupo funcional presentes en modelo
Ecopath en la plataforma continental de Yucatán, México .................................................... 9
Tabla 2. Parámetros utilizados para la estimación de los cocientes P/B y Q/B de la especie
representativa de cada grupo funcional de peces del ecosistema de la plataforma continental
de Yucatán. ........................................................................................................................... 14
Tabla 3. Principales flotas y recurso de la plataforma continental de Yucatán ................... 17
Tabla 4. Referencias usadas para obtener las dietas de las especies representantes de los
grupos funcionales incluidos en el modelo........................................................................... 20
Tabla 5. Datos de entrada del modelo del ecosistema de la plataforma continental de
Yucatán. Los valores en negrita fueron calculados por Ecopath. ......................................... 30
Tabla 6. Matriz de dietas, en las columnas se encuentran los predadores y en las filas las
presas. ................................................................................................................................... 32
Tabla 7. Índices clave estimados para los grupos funcionales de la plataforma continental de
Yucatán ................................................................................................................................. 42
Tabla 8. Resumen de estadísticos del ecosistema de la plataforma continental de Yucatán
.............................................................................................................................................. 45
Tabla 9. Índices de flujo de desarrollo del ecosistema de la plataforma continental de
Yucatán ................................................................................................................................. 48
Tabla 10. Resultados del análisis de componentes principales ........................................... 55
Tabla 11. Resultados del análisis de componentes principales, con la clasificación y
extracción de los componentes de los atributos del ecosistema ........................................... 55
Tabla 12. Resultados del análisis de componentes principales, con la clasificación y
extracción de los componentes de los atributos de cada grupo funcional ............................ 55

iii
Anexos
Anexo 1. Gráfica de frecuencia de índice de grado ............................................................. 87
Anexo 2. Gráfica de frecuencia del índice de cercanía ........................................................ 87
Anexo 3. Gráfica de frecuencia del índice de intermediación ............................................. 87
Anexo 4. Cambios en la composición de las capturas del mero y pulpo a través del periodo
1990 – 2007. Tomadas de SAGARPA- Cinvestav 2008 ...................................................... 88
Anexo 5. Composiciónde las capturas en Yucatán. Tomadas de SAGARPA – Cinvestav 2008
.............................................................................................................................................. 88

iv
Glosario
Ascendencia: es un índice ecológico que mide el crecimiento y desarrollo de un ecosistema.
Este índice está basado en el tamaño y organización, considerando los flujos totales y
contenido de información en la trama trófica (Ulanowicz, 1986)
Capacidad de desarrollo: es el límite (teórico) de la ascendencia y mide la capacidad de
crecimiento de un ecosistema (Ulanowicz, 1986).
Cociente Consumo/Biomasa: esta relación representa la cantidad de alimento ingerido por
un organismo respecto a su propia biomasa en un periodo de tiempo determinado (Pauly,
1989).
Cociente Producción/Biomasa: es equivalente a la tasa instantánea de mortalidad total
(Allen, 1971) y representa la producción biológica de un organismo por unidad de biomasa.
Cociente Producción/Consumo: corresponde a la eficiencia bruta de conversión de
alimento.
Ecopath: es un programa para construir modelos (simples o complejos) de los flujos tróficos
del ecosistema.
Ecosim: modelo dinámico; consiste en una trasformación dinámica de la ecuación en
equilibrio del modelo de Ecopath.
Ecosistema: unidad funcional de varias especies localizadas en un sitio determinado y sujetas
a las mismas condiciones ambientales (Tansley, 1935).
Eficiencia ecotrófica: proporción de la producción que se utiliza en el ecosistema a través
de la depredación, migración, capturas y acumulación de biomasa.
Eficiencia de transferencia: es una proporción que expresa cuan óptimo es el transporte de
energía de un nivel trófico al siguiente.
Entropía: Es un término de termodinámica que se refiere a una medida de la energía no libre
del sistema o a la cantidad de desorden que guarda un objeto o un sistema.

v
Flujo: es un vector de energía (biomasa) que describe una trayectoria y una magnitud para
cada compartimento dentro de la red trófica.
Grupo funcional: especie o grupo de especies que pueden presentar afinidades taxonómicas
y/o hábitos alimenticios similares, aunque también pueden ser aquellas especies de mayor
importancia comercial para una pesquería o las más abundantes en el medio.
Impacto trófico: es el efecto positivo o negativo ocasionado por la depredación o pesca
como resultado de las interacciones tróficas.
Madurez: estado final en la sucesión ecológica de los ecosistemas que se alcanza después
de haber pasado por diferentes etapas intermedias, que comprenden cambios en su estructura.
Se caracteriza por tener un grado máximo de biomasa por unidad de energía presente en
concordancia con el ambiente en que se desarrolla (Odum 1969).
Modelo: es una representación simple o compleja de un sistema ecológico, que enfatizan
ciertos aspectos del sistema investigado para entender su funcionamiento.
Nivel trófico: es la posición que ocupa en la trama trófica una especie o grupo funcional de
acuerdo a sus hábitos alimenticios.
Orden: es la ganancia o pérdida de entropía
Overhead: potencial de reserva del ecosistema ante perturbaciones (Ulanowicz, 1986), y se
calcula como la diferencia entre la capacidad de desarrollo y la ascendencia.
Resiliencia: es un atributo de la estabilidad y es el que describe la capacidad de los
ecosistemas de regresar rápidamente al punto inicial, antes de que se presentara el disturbio
que provocó el cambio

vi
Resumen
El objetivo del presente estudio fue conocer el papel del mero rojo dentro del ecosistema de
la plataforma continental de Yucatán, para lo cual se construyó un modelo trófico empleando
la plataforma de programas Ecopath con Ecosim. El modelo está compuesto por 37 grupos
funcionales, siendo los grupos de niveles tróficos primarios y secundarios los que aportan
mayor biomasa al ecosistema. La mayoría de los flujos de niveles tróficos menores estuvieron
dirigidos hacia detritus; y una fracción alta de los flujos totales se destina para el
mantenimiento de la estructura de la red trófica. El índice de conectividad fue igual a 0.2,
esto significa que sólo existen 20 % de las conexiones totales posibles en la trama alimentaria.
De acuerdo con los índices de desarrollo, el ecosistema resulto ser inmaduro, pero con una
alta capacidad de responder ante perturbaciones externas. De acuerdo con los atributos
evaluados para el mero rojo, se identificó que el nivel trófico, Q/B y A/C fueron los atributos
más característicos y significativos del grupo, lo cual sugiere el mero es un grupo que
contribuye de manera importante a mantener la estabilidad del ecosistema, caracterizando al
mero como un predador tope, con hábitos generalistas, capaz de regular la abundancia de sus
presas en el ecosistema. Por otro lado, los experimentos de simulación basados en la
respuesta del ecosistema ante el forzamiento ambiental del AMO, mostraron que los atributo
de biomasa y orden (A/C) del mero rojo, presenta un patrón de variación de tipo cíclico,
cuando la biomasa tiende a decaer el A/C decrece, a niveles bajos de biomasa, esta se
mantiene estable y el orden del ecosistema aumenta. Posteriormente la biomasa aumenta y el
orden tiende a decrecer, y a niveles altos de biomasa, esta decrece gradualmente
manteniéndose el orden relativamente estable para posteriormente disminuir, completándose
el ciclo. Este patrón parece reflejar la respuesta del ecosistema a las variaciones de la
abundancia del mero como reflejo del efecto ambiental.
Palabras clave: mero rojo, ecosistema, modelo trófico, Ecopath, Ecosim, grupos
funcionales, flujos, red trófica, conexiones, simulación, desarrollo, inmaduro, análisis de
componente principales, orden

vii
Abstract
The study seek to examine the role of red grouper in the ecosystem of the continental shelf
of Yucatan, for which a trophic model using the suite of programs Ecopath with Ecosim was
used. The model consists of 37 functional groups; from which, those pertaining to primary
and secondary trophic levels contribute with higher biomasses to the ecosystem. Main flows
of lower trophic levels were directed to detritus; and a high fraction of total flows is intended
to maintain the structure of the food web. The connectivity index suggests only 20 % of total
possible connections in the food web are realized. According to the index of ecosystem
development, the ecosystem turned out to be immature, but with a relative high resilience.
According to the attributes evaluated for the red grouper, it was identified that the trophic
level, Q/B and A/C were the most characteristic and significant attributes of the group,
suggesting the red grouper is a group that contributes importantly to the to ecosystem
maintenance and stability; characterizing the red grouper as a top predator, generalist in its
feeding behavior, able to regulate the abundance of its preys in the ecosystem. On the other
hand, the simulation experiments based on the ecosystem response to environmental forcing,
the AMO, show that the biomass and order (A/C) attributes relative to the red grouper present
a cyclical pattern of variation; when biomass tends to decay, A/C (order) decreases; at low
levels of biomass, it remains stable and ecosystem order increases. Subsequently, biomass
increases and the order tends to decrease slightly, and at high levels of biomass, it decreases
gradually, maintaining the ecosystem order relatively stable, decreasing abruptly when
biomass also decrease notably, completing the cycle. This pattern seems to reflect the
response of the ecosystem to the variations of the red grouper abundance as response to the
environmental effect.
Keywords: red grouper, ecosystem, trophic model, Ecopath, Ecosim, functional groups
flows, food web, connections, simulation, development, immature, principal component
analysis, order

1
1. Introducción
El papel ecológico que desempeñan cada una de las especies, de acuerdo a su composición,
patrones de comportamiento y hábitos alimenticios, generan las rutas indispensables en el
flujo energético que sostiene al ecosistema (Sánchez-Gil & Yáñez- Arancibla, 1999). Los
procesos ecosistémicos, la naturaleza y magnitud de las contribuciones individuales de cada
especie varían considerablemente en el ecosistema, por ende, es importante identificar la
composición específica o la identidad de las especies en una comunidad. (Naemm et al.,
1999).
Por todo lo anteriormente expuesto, se infiere que los estudios de interacción
ecológica, tales como la depredación y competencia, junto con otros factores de régimen
ambiental y alteración del hábitat están adquiriendo una importancia creciente de cara a los
modelos de evaluación multiespecífica. Y de igual manera, están siendo considerados en la
implementación del enfoque de la pesca basado en el ecosistema, ya que en su conjunto afecta
a la dinámica de las poblaciones en términos de abundancia y composición (Christensen,
1996; Pauly et al., 2000; Link, 2002; Stergiou y Karpouzi, 2002; Bascompte et al., 2005;
Cury et al., 2005). Ante tal panorama, los modelos de balance de masa, como el modelo
Ecopath with Ecosim (Pauly & Christensen, 1992; Walter et al., 1997; 1999, 2000) han sido
una de las alternativas más utilizadas en la cuantificación de las interacciones tróficas en
sistemas explotados. Estas aproximaciones permiten cuantificar y predecir los cambios en
tiempo y espacio y con la alternativa de explorar los impactos pasados y futuros que ejercen
la pesca y los factores forzantes (Walters, 1997, 1999, 2004, Christensen & Walters, 2004).
En el caso específico del mero rojo (Epinephelus morio), desempeña el papel de
máximo predador en la trama trófica, pues es un carnívoro activo que se alimenta de peces
(entre los cuales predominan lutjánidos, hemúlidos y jureles), así como también varios tipos
de invertebrados, incluyendo taxas de interés comercial, como: moluscos y crustáceos (Brulé
& Rodríguez-Canché, 1993; Brulé & Déniel, 1994; Giménez-Hurtado et al., 2001). También
se caracteriza por tener hábitos demersales en zonas tropicales y subtropicales con
temperaturas mayores de los 20° C, la incidencia más alta de esta especie se presenta en aguas
de 23° a 25 ° C. Vive sobre fondos duros, arrecifes coralinos o cualquier cueva o refugio del
fondo. A los juveniles y pre-adultos se les encuentra en aguas bajas cerca de la costa, entre

2
10 y 30 metros de profundidad y a los adultos en profundidades hasta de 130 metros
(González et al., 1974). Esta especie es uno de los serránidos más comunes y de importancia
comercial del Golfo de México, con dos centros de abundancia: uno en la costa occidental
de la Florida y otro en la plataforma continental de Yucatán, conocida como Sonda o Banco
de Campeche (Giménez et al., 2001). Además esta especie ha representado una fuente
importante de empleos y de captación de divisas, e históricamente ha sido la pesquería más
importante del Golfo de México después del camarón. A inicios de los años 1950 y principios
de los 1970, la pesquería de este recurso tuvo su fase de crecimiento, alcanzando el máximo
rendimiento histórico en 1972 (19, 886 toneladas); declinando hasta la época actual, con
rendimientos anuales de alrededor de las 8,000 t, mostrando fluctuaciones (DOF, 2015). En
general, los estudios que se han realizado sobre esta especie han sido enfocados en su pesca
y biología, principalmente en: reproducción, parámetros de crecimiento y hábitos
alimenticios (esto como medida estratégica en recuperar la abundancia del recurso), no
obstante, los trabajos desarrollados no contemplan el efecto del papel de la especie en el
ecosistema, lo cual podría ser un elemento útil en la caracterización del funcionamiento del
ecosistema y por ende en el desarrollo de medidas de manejo eficientes para la toma de
decisiones del recurso.
2. Antecedentes
En 1993 se publicaron los primeros modelos tróficos representativos de ecosistema del Golfo
de México. Entre ellos se puede citar el de Browder para el norte del Golfo de México
(plataforma continental de USA). Así mismo De la Cruz-Agüero construyó un modelo
preliminar de flujos tróficos entre las comunidades bióticas en laguna de Mandigna,
Veracruz. Mientras que Abarca-Arenas y Valero-Pacheco modelaron las interacciones
tróficas de la laguna costera de Tamiahua, Veraccruz. Por otro lado Chávez et al., realizaron
una extensión del estudio de la estructura trófico-dinámica de la Laguna Celestun, Yucatán
en el sur del Golfo de México. Arreguín-Sánchez et al. (1993) construyeron un modelo de
flujos tróficos de la comunidad de peces costeros del suroeste del Golfo de México. Por
último Vega-Cendejas et al., analizaron los flujos tróficos del ecosistema de la región costera
del Banco de Campeche. Todos los trabajos publicados resaltan la importancia de los grupos

3
funcionales dentro de cada ecosistema, así como el funcionamiento del ecosistema en
términos de flujos de energía.
Concretamente para la zona de la plataforma continental de Yucatán, Arreguín-Sánchez et
al. (1993) encontraron que cerca de la mitad de la energía proviene de los productores
primarios (bentónicos y fitoplancton) y el resto se importa como detritus, así mismo los
grupos asociados al bentos como microcrustáceos y anélidos fueron de los que más aportaron
energía al sistema. Por otro lado, se encontró que la pesca tiene una alta influencia dentro de
ecosistema (equivalente a un depredar que ocupa un nivel trófico medio 5.1), siendo los
grupos más afectados el mero rojo, tiburones, truchas de mar y corvina.
Vega-Cendejas y Arreguín-Sánchez (2001) evaluaron los flujos de energía del ecosistema de
manglar de una laguna costera en la Península de Yucatán. 19 grupos funcionales fueron los
que se integraron al modelo. Se incluyeron a dos grupos de productores primarios, diez de
invertebrados marinos carnívoros y tres grupos de carnívoros incluyendo a peces y
microcrustáceos, el grupo que aporto mayor biomasa al ecosistema fue el detritus (2,400 g
m-2 año-1), seguido del grupo de macrofitas (153.42 g m-2 año-1), mientras que el grupo de
zooplancton obtuvo la mayor tasa de consumo y desechos metabólicos. La transferencia de
energía del ecosistema fue 1924 kcal m-2 año-1. Los atributos del ecosistema determinaron
que se encuentra en un estado inmaduro.
En el 2000 Arreguín-Sánchez describió la interacción de explotación del pulpo y mero rojo
en el ecosistema de la parte norte de la plataforma continental de Yucatán. Este trabajo se
evaluó los cambios en la vulnerabilidad del pulpo debido a la depredación por mero rojo bajo
tres escenarios de mortalidad por pesca (F). El primer escenario la F fue baja y constante; en
el segundo escenario la F incrementaba gradualmente; el tercer escenario prevalecieron altos
valores de F y por último se presenta un cuarto escenario, basado en el escenario 2, para
observar los patrones de biomasa de otros grupos. Para todos los escenarios de observó que
cuando la vulnerabilidad del pulpo a la depredación redujo, su biomasa aumenta, y en caso
contrario disminuye cuando la depredación aumenta. Este mismo patrón se encontró en altos
niveles de vulnerabilidad cuando los pulpos fueron sometidos a un alto esfuerzo de pesca.
Arreguín-Sánchez et al. (2015). Realizaron un modelo ecosistémico del sureste del Golfo de
México y evaluando un caso en particular el manejo de la pesquería del camarón rosado,

4
considerando como indicadores ambientales la Oscilación Multidecadal del Atlántico (AMO
por sus siglas en inglés) y la Temperatura Superficial del Mar (TSM). Se demostró que las
variaciones climáticas dentro del Golfo de México tienen efecto sobre los procesos
oceanográficos y atmosféricos locales, y por consiguiente en el ecosistema. Tal es caso del
AMO durante los últimos 50 años. Y se encontró que el AMO explica alrededor del 50 % de
la variación del reclutamiento del camarón rosado. Además, se encontró que los niveles más
altos de la capacidad de carga fueron a mediados de la década de 1970 y declinando después
de eso para todos los grupos considerados incluyendo el camarón rosado.
3. Justificación
Los modelos poblacionales que de manera convencional son utilizados para asesorar al sector
pesquero, no son suficientes para explicar y representar la variación de la abundancia de las
poblaciones explotadas, y se ha sugerido adoptar un enfoque de manejo basado en el
ecosistema (FAO 2008). En este sentido, el estado actual de la aproximación de manejo
basado en el ecosistema se lleva a cabo desde la perspectiva del recurso objetivo y como este
se vincula al resto del ecosistema. En el caso del mero rojo (E. morio), los antecedentes en
literatura hablan de efectos de la pesca y ambientales, ambos incidiendo sobre la abundancia
del recurso, lo cual lleva implícito la re-organización del ecosistema. Sin embargo, se carece
de información sobre el papel de la especie en la estructura, función y organización del
ecosistema que explique otras fuentes de variación aparte de la pesca.
Por otro lado, se ha destacado en la literatura el papel de los predadores tope en el
mantenimiento o control de la estructura trófica del ecosistema (Baum & Worm, 2009,
Ritchie & Johnson, 2009).
4. Hipótesis
El estado actual de la pesquería del mero es resultado de dos fuentes de variación
significativas, la pesca y el ambiente; de tal suerte que el conocimiento de su papel ecológico
resulta de relevancia para la toma de decisiones orientadas al manejo de la pesquería en el
contexto ecosistémico.

5
5. Objetivo general
Conocer el papel del mero rojo (Epinephelus morio) en la estructura, función y organización
del ecosistema de la plataforma continental de Yucatán.
5. 1. Objetivos específicos
Construir un modelo trófico tipo Ecopath del ecosistema de la plataforma continental
de Yucatán
Estimar atributos en función de organización del ecosistema de la plataforma
continental de Yucatán
Identificar el papel del mero rojo en la organización del ecosistema de la plataforma
continental de Yucatán
6. Área de estudio
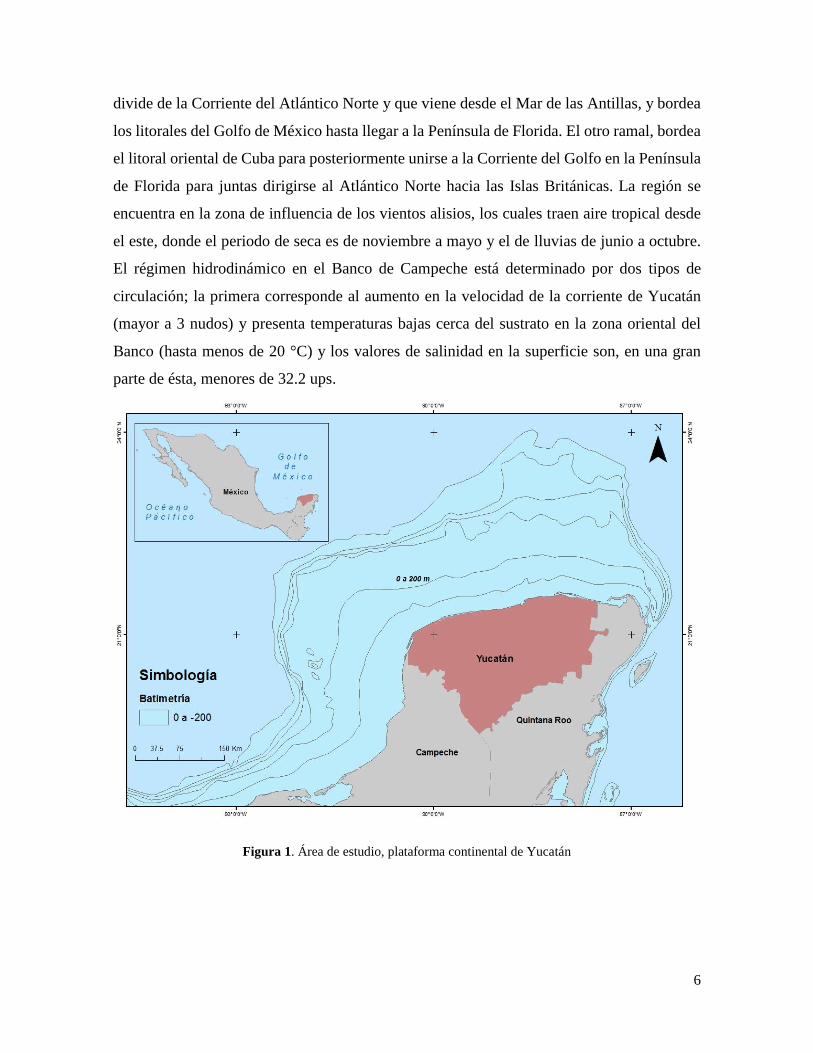
El área de estudio se encuentra localizada en la plataforma continental de Yucatán, México;
que junto con la plataforma de Campeche constituyen el denominado Banco de Campeche,
los límites geográficos de la plataforma continental de Yucatán son 21° 35’ N, 86° 43’ W
(Figura 1), y comprende un área de 129, 499.5 km2 aproximadamente, delimitada por la
isóbata de 200 metros y la línea de costa. El sustrato está caracterizado por sedimentos
constituidos por proporciones variables de calizas, arena y conchas, además de existir algunas
irregularidades formadas por cayos y arrecifes coralinos (García 1980). Por su situación
geográfica, la plataforma de Yucatán se encuentra en la zona tropical. Bogdanov (1965)
sugiere que dentro de las condiciones oceanográficas de importancia, el proceso que domina
el ambiente costero en la Península de Yucatán es la Corriente de Yucatán, que fluye con
fuerte intensidad por el Caribe y penetra en el Golfo de México a través del estrecho de
Yucatán. Esta corriente presenta un flujo de 25 a 35 x 106 m3 s-1 y una velocidad promedio
de 0.80 m s-1 en la superficie, y hasta de 1.50 m s-1 a una profundidad de 300 m. Por debajo
de la Corriente de Yucatán circula una contracorriente que es parte importante del mecanismo
de surgencia en el Banco de Campeche (Reyes, 2005). Al respecto, Tamayo (1974) reporta
que la corriente que baña las costas de la Península de Yucatán es uno de los ramales que se
6
divide de la Corriente del Atlántico Norte y que viene desde el Mar de las Antillas, y bordea
los litorales del Golfo de México hasta llegar a la Península de Florida. El otro ramal, bordea
el litoral oriental de Cuba para posteriormente unirse a la Corriente del Golfo en la Península
de Florida para juntas dirigirse al Atlántico Norte hacia las Islas Británicas. La región se
encuentra en la zona de influencia de los vientos alisios, los cuales traen aire tropical desde
el este, donde el periodo de seca es de noviembre a mayo y el de lluvias de junio a octubre.
El régimen hidrodinámico en el Banco de Campeche está determinado por dos tipos de
circulación; la primera corresponde al aumento en la velocidad de la corriente de Yucatán
(mayor a 3 nudos) y presenta temperaturas bajas cerca del sustrato en la zona oriental del
Banco (hasta menos de 20 °C) y los valores de salinidad en la superficie son, en una gran
parte de ésta, menores de 32.2 ups.
Figura 1. Área de estudio, plataforma continental de Yucatán

7
7. Materiales y métodos
7.1 Descripción del modelo Ecopath
Originalmente el enfoque Ecopath fue propuesto por Polovina (1984) y posteriormente
modificado por Christensen y Pauly (1992). Ecopath es un modelo que permite caracterizar
la estructura trófica y funcionalidad de un ecosistema con base en el balance de masas (donde
para cada grupo funcional o especie la producción es igual a las pérdidas), que permite
también describir atributos de su organización a través del análisis de redes (Ulanowicz
1986).
Las interacciones tróficas entre los grupos funcionales del ecosistema son descritos
por un conjunto de ecuaciones lineales que representa cada uno de los grupos funcionales en
el ecosistema. Dichas ecuaciones están basadas en dos ecuaciones maestras, que suponen un
sistema de balance de masas entre los diferentes componentes (especies o grupos
funcionales), una describe la producción (Ecuación 1) en términos de masa y la segunda se
describe el balance de energía de cada grupo (Ecuación 4).
La ecuación de producción para cada grupo (i) puede ser dividida en componentes:
𝑃𝑟𝑜𝑑𝑢𝑐𝑐𝑖ó𝑛 (𝑃) = 𝑐𝑎𝑝𝑡𝑢𝑟𝑎𝑠 (𝐶) + 𝑚𝑜𝑟𝑡𝑎𝑙𝑖𝑑𝑎𝑑 𝑝𝑜𝑟 𝑑𝑒𝑝𝑟𝑒𝑑𝑎𝑐𝑖ó𝑛(𝐷) + 𝑜𝑡𝑟𝑎𝑠 𝑚𝑜𝑟𝑡𝑎𝑙𝑖𝑑𝑎𝑑𝑒𝑠
Y formalmente se representa como:
𝑃𝑖 = 𝑌𝑖 + 𝐵𝑖 ∙ 𝑀2𝑖 + 𝑃𝑖 ∙ (1 − 𝐸𝐸𝑖)
donde para un grupo i, Pi es la producción, Bi es la biomasa, M2i es la mortalidad por
depredación y EEi es la eficiencia ecotrófica (proporción la producción que se emplea en el
sistema). Así, que el término de la izquierdo de la ecuación representa la producción (Pi), en
el lado derecho el primer termino representa las capturas (Yi), el segundo las perdidas por
depredación (Bi · M2i) y el tercero las pérdidas que no son debidas a depredación ni a
exportación (Pi · (1-EEi)).
Dado que la transferencia de materia entre los grupos se da a través de las relaciones
tróficas, la anterior ecuación es expresada de la siguiente manera para incluir a la predación:
Ecuación 1
Ecuación 2

8
𝐵𝑖 (𝑃
𝐵)
𝑖∙ 𝐸𝐸𝑖 = 𝑌𝑖 + 𝐵𝑖 ∙ (
𝑃
𝐵)
𝑖∙ (1 − 𝐸𝐸𝑖) + ∑ 𝐵𝑗 ∙
𝑛
𝑗=1
(𝑄
𝐵)
𝑗∙ 𝐷𝐶𝑗𝑖
donde el subíndice j representa a los predadores; Bj es la biomasa, Q es el consumo y DCji
es la proporción de la especie i en la dieta del predador j. La producción y el consumo se
representa como proporciones de sus biomasas.
La segunda ecuación maestra de Ecopath se encarga del balance de energía de cada grupo en
donde las entradas y salidas de energía deben ser balanceadas. El balance de energía es
definido dentro de cada grupo por medio de la siguiente ecuación:
𝐶𝑜𝑛𝑠𝑢𝑚𝑜 = 𝑝𝑟𝑜𝑑𝑢𝑐𝑐𝑖ó𝑛 (𝑃) + 𝑟𝑒𝑠𝑝𝑖𝑟𝑎𝑐𝑖ó𝑛 (𝑅) + 𝑎𝑙𝑖𝑒𝑚𝑡𝑜 𝑛𝑜 𝑎𝑠𝑖𝑚𝑖𝑙𝑎𝑑𝑜 (𝐴𝑁𝐴)
Se incluye el balance de energía de los grupos con la finalidad de estimar la respiración a
partir de las diferencias entre los términos de consumo, producción y alimento no asimilado.
7.2 Construcción del modelo
El modelo construido se basa en el modelo preliminar de Arreguín-Sánchez et al. (1993) y
Arreguín-Sánchez (2000), cabe destacar que el modelo se actualizó, integrando 16 grupos
funcionales, además, se actualizó la información de datos de entrada al modelo de cada grupo
funcional a un periodo más reciente (2006 - 2014). El primer paso para la construcción del
modelo fue definir los grupos funcionales, estos fueron conformados de acuerdo a su
importancia como recurso pesquero, ecológica y su abundancia en la zona. Se consideraron
37 grupos funcionales, de los cuales tres son productores primarios (macroalgas, pastos
marinos y fitoplancton), uno de detritus, nueve de invertebrados (pulpo, microcrustáceos,
calamar, camarón rojo, langosta, jaiba azul, poliquetos, zooplancton y equinodermos), 21 de
peces, y uno para los siguientes: delfín, ave marina, tortuga marina. En la tabla 1 se muestran
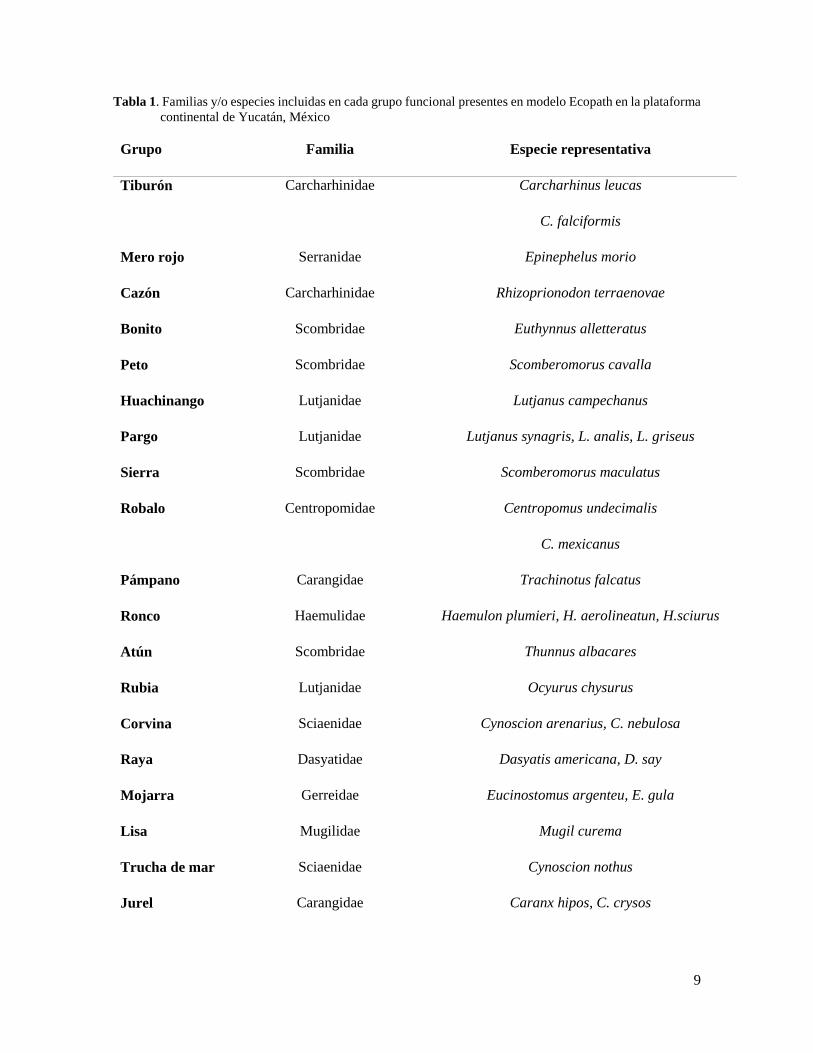
las familias y especies más representativas de los grupos funcionales.
Ecuación 4
Ecuación 3
9
Tabla 1. Familias y/o especies incluidas en cada grupo funcional presentes en modelo Ecopath en la plataforma
continental de Yucatán, México
Grupo Familia Especie representativa
Tiburón Carcharhinidae Carcharhinus leucas
C. falciformis
Mero rojo Serranidae Epinephelus morio
Cazón Carcharhinidae Rhizoprionodon terraenovae
Bonito Scombridae Euthynnus alletteratus
Peto Scombridae Scomberomorus cavalla
Huachinango Lutjanidae Lutjanus campechanus
Pargo Lutjanidae Lutjanus synagris, L. analis, L. griseus
Sierra Scombridae Scomberomorus maculatus
Robalo Centropomidae Centropomus undecimalis
C. mexicanus
Pámpano Carangidae Trachinotus falcatus
Ronco Haemulidae Haemulon plumieri, H. aerolineatun, H.sciurus
Atún Scombridae Thunnus albacares
Rubia Lutjanidae Ocyurus chysurus
Corvina Sciaenidae Cynoscion arenarius, C. nebulosa
Raya Dasyatidae Dasyatis americana, D. say
Mojarra Gerreidae Eucinostomus argenteu, E. gula
Lisa Mugilidae Mugil curema
Trucha de mar Sciaenidae Cynoscion nothus
Jurel Carangidae Caranx hipos, C. crysos

10
Anchoveta Engraulidae Anchoa mitchilli
Cetengraulis edentulus
Sardina Clupeidae Opisthonema oglinum, Harengula jaguana
Delfín Delphinidae Tursiops truncatus
Aves marinas Fregatidae Fregata magnificens
Tortugas
marinas
Cheloniidae Chelonia mydas, Lepidochelis olivacea
Pulpo Octopodidae Octopus maya
Microcrustáceos Melongenidae Melongena corona bispinosa
Calamar Loliginidae Doryteuthis pealeii
Camarón rojo Penaeidae Farfantepenaeus sp.
Langosta Palinuridae Panulirus argus
Jaiba azul Portunidae Callinectes sapidus
Poliquetos Tomopteridae
Zooplancton Centropagidae, Palinuridae
(larvas), Portunidae (larvas),
Mugilidae (larvas),
Engraulidae (larvas),
Clupeidae (larvas),
Palinuridae (larvas).
Equinodermos Psolidae
Stichopodidae
Holothuria floridana
Isostichopus badionotus
Macroalgas Chlorophyceae Cladophorasp.
Pastos marino Thalassia Thalassia testudinum
Fitoplancton Goniodomateceae Phyrodinium sp.

11
7.2.1 Estimación de los datos de entrada
Los datos de entrada utilizados para la construcción del modelo fueron la biomasa (B), tasa
de producción biomasa (P/B), tasa de consumo biomasa (Q/B), captura (Y), eficiencia
ecotrófica (EE) y la matriz de predador-presa construida con los datos de composición de la
dieta de las presas y predadores (DCij). Para estimar estos parámetros se utilizó información
sobre los hábitos alimentarios, edad y crecimiento de cada grupo funcional para la zona de
estudio. En algunos casos, debido a la falta de información para la zona, fue necesario
complementarla con datos publicados de las mismas especies en otros ecosistemas.
7.2.2 Biomasa (B)
Para el caso de los grupos de peces, pulpo y langosta, se estimó la biomasa a partir con datos
de avisos de arribo (2006 - 2011), asumiendo que existe una relación entre la CPUE y la
abundancia del stock, se utilizaron los coeficientes de capturabilidad obtenidos con el modelo
de Leslie y Davies (1939), expresado de la siguiente forma:
𝐶𝑡
𝑓𝑡= 𝑞𝐵𝑡 = 𝑞𝑁0 − 𝑞𝐾𝑡
donde t, es un tiempo dado, Ct y ft representan la CPUE durante un determinado tiempo, q
es la capturabilidad, qN0 equivalen a la pendiente e intercepto de regresión lineal; Kt es la
captura acumulada y N es el tamaño poblacional. Donde 𝐵𝑡 =(
𝐶𝑡𝑓𝑡
)
𝑞
Para los productores primarios y detritus las biomasas se obtuvieron del trabajo de Arreguín-
Sánchez 2000. Y para el grupo de tortuga marina ave marina y delfín se utilizaron los datos
de trabajos de Cabrera-Neri (2005). En el caso del resto de los grupos de invertebrados se
dejó que Ecopath estimara su biomasa.
7.2.3 Producción biomasa (P/B)
El cociente P/B es equivalente a la tasa instantánea de mortalidad total (Z año-1) en
condiciones de equilibrio (Allen 1971). Esta tasa está compuesta principalmente por la
mortalidad natural (M) y la mortalidad por pesca (F). Para los peces, expresado como:
𝑍 = 𝑀 + 𝐹
Ecuación 5
Ecuación 6

12
Para la estimación de M se utilizó el modelo propuesto por Taylor (1960). Este
modelo supone que la longevidad de una especie se relaciona con la tasa de crecimiento (K)
(Longhurst & Pauly 1987) y es inversamente proporcional a su mortalidad natural. De esta
manera, sí se considera la edad límite como el tiempo requerido para alcanzar 95 % de
longitud máxima promedio, se tiene que le índice de longevidad (A0.95) de expresa de la
siguiente manera:
𝐴0.95 = 𝑡0 +2.996
𝐾
donde, t0 es una constante que representa la edad que debería de tener el organismo para que
su longitud sea igual a cero; K es el coeficiente de crecimiento, y la mortalidad natural según
el modelo de Taylor queda de la siguiente forma:
𝑀 =2.996
𝐴0.95− 𝑡0
Los parámetros t0, K de la mayoría de los grupos de peces fueron tomados de Fishbase
(Froese & Pauly 1995).
Para los grupos de peces explotados, la mortalidad por pesca F se estimó mediante la
siguiente formula:
𝐹 = (𝐶
𝐵)
Se utilizó la base de datos FishBase (www.fishbase.org) (Frose y Pauly, 2000) como parte
complementaria en la construcción de la base de datos.
Los datos para los grupos de invertebrados fueron tomados de lo reportado en el
modelo de Arreguín-Sánchez et al. (1993), Cabrera-Neri (2005) y Criales- Hernández (2006)
(Tabla 5).
Ecuación 7
Ecuación 8
Ecuación 9

13
7.2.4 Consumo/biomasa (Q/B)
El cociente Q/B se refiere a la cantidad de alimento ingerido por un grupo expresado con
respecto a su propia biomasa en el periodo de tiempo considerado. Para los grupos de peces
se empleó la ecuación 10 propuesta por Palomares y Pauly (1998):
𝑄𝐵⁄ = 10⋀ (
7.964−0.204∗𝑙𝑜𝑔𝑊∞−1.965 ∗1000
𝑇+273.15+ 0.083 ∗ 𝐴 + 05.32 ∗ ℎ + 0.398 ∗ 𝑑)
donde W∞ es el peso asintótico del modelo de crecimiento von Bertalanffy (g), T es la
temperatura promedio del sistema donde habita la especie, A es una variable morfométrica
que representa el aspecto de la aleta caudal (A=h2/s, donde h es la altura de la aleta caudal y
s es la superficie de la aleta caudal) y d es el tipo de alimentación que consume el organismo
(toma valores de cero para carnívoros, uno para herbívoros u detritívoros) ( ver Tabla 2).
Para los grupos que nos son peces, los cocientes Q/B se obtuvieron de los trabajos de
Arreguín-Sánchez et al. (1993), Cabrera-Neri (2005) y Criales- Hernández (2006) (Tabla 5).
Ecuación 10
14
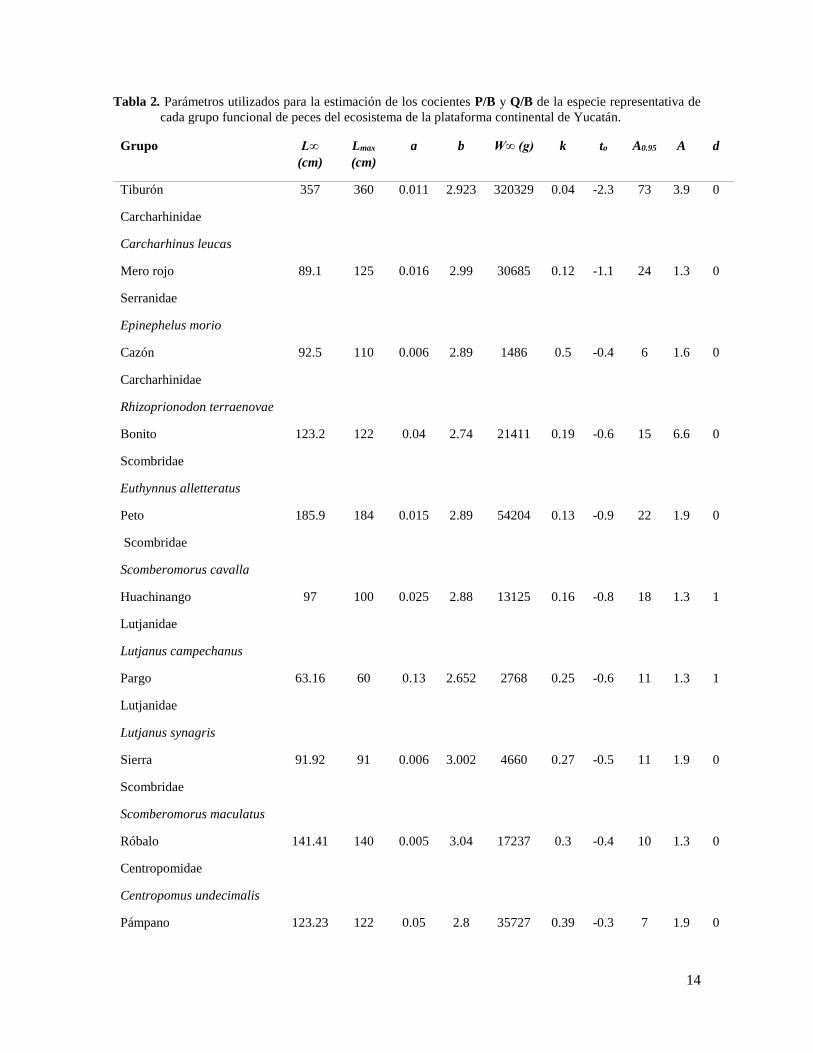
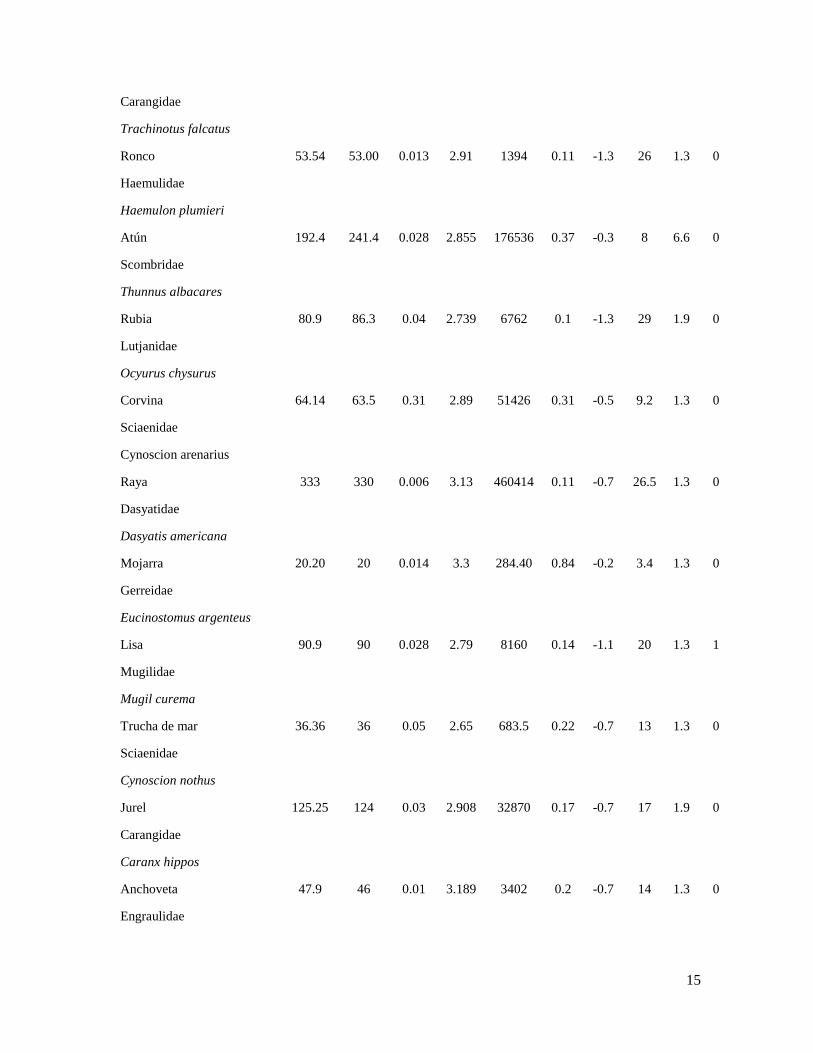
Tabla 2. Parámetros utilizados para la estimación de los cocientes P/B y Q/B de la especie representativa de
cada grupo funcional de peces del ecosistema de la plataforma continental de Yucatán.
Grupo L∞
(cm)
Lmax
(cm)
a b W∞ (g) k to A0.95 A d
Tiburón 357 360 0.011 2.923 320329 0.04 -2.3 73 3.9 0
Carcharhinidae
Carcharhinus leucas
Mero rojo 89.1 125 0.016 2.99 30685 0.12 -1.1 24 1.3 0
Serranidae
Epinephelus morio
Cazón 92.5 110 0.006 2.89 1486 0.5 -0.4 6 1.6 0
Carcharhinidae
Rhizoprionodon terraenovae
Bonito 123.2 122 0.04 2.74 21411 0.19 -0.6 15 6.6 0
Scombridae
Euthynnus alletteratus
Peto 185.9 184 0.015 2.89 54204 0.13 -0.9 22 1.9 0
Scombridae
Scomberomorus cavalla
Huachinango 97 100 0.025 2.88 13125 0.16 -0.8 18 1.3 1
Lutjanidae
Lutjanus campechanus
Pargo 63.16 60 0.13 2.652 2768 0.25 -0.6 11 1.3 1
Lutjanidae
Lutjanus synagris
Sierra 91.92 91 0.006 3.002 4660 0.27 -0.5 11 1.9 0
Scombridae
Scomberomorus maculatus
Róbalo 141.41 140 0.005 3.04 17237 0.3 -0.4 10 1.3 0
Centropomidae
Centropomus undecimalis
Pámpano 123.23 122 0.05 2.8 35727 0.39 -0.3 7 1.9 0
15
Carangidae
Trachinotus falcatus
Ronco 53.54 53.00 0.013 2.91 1394 0.11 -1.3 26 1.3 0
Haemulidae
Haemulon plumieri
Atún 192.4 241.4 0.028 2.855 176536 0.37 -0.3 8 6.6 0
Scombridae
Thunnus albacares
Rubia 80.9 86.3 0.04 2.739 6762 0.1 -1.3 29 1.9 0
Lutjanidae
Ocyurus chysurus
Corvina 64.14 63.5 0.31 2.89 51426 0.31 -0.5 9.2 1.3 0
Sciaenidae
Cynoscion arenarius
Raya 333 330 0.006 3.13 460414 0.11 -0.7 26.5 1.3 0
Dasyatidae
Dasyatis americana
Mojarra 20.20 20 0.014 3.3 284.40 0.84 -0.2 3.4 1.3 0
Gerreidae
Eucinostomus argenteus
Lisa 90.9 90 0.028 2.79 8160 0.14 -1.1 20 1.3 1
Mugilidae
Mugil curema
Trucha de mar 36.36 36 0.05 2.65 683.5 0.22 -0.7 13 1.3 0
Sciaenidae
Cynoscion nothus
Jurel 125.25 124 0.03 2.908 32870 0.17 -0.7 17 1.9 0
Carangidae
Caranx hippos
Anchoveta 47.9 46 0.01 3.189 3402 0.2 -0.7 14 1.3 0
Engraulidae

16
Cetengraulis edentulus
Sardina 25.9 38 0.02 2.92 283 0.65 -0.3 4 1.3 0
Clupeidae
Opisthonema oglinum
Donde L∞ es longitud promedio asintótico; Lmax es la máxima longitud observada de la especie; a y b son
constantes de la relación longitud-peso; K es el coeficiente de crecimiento; t0 es una constante que representa
la edad que debería de tener el organismo para que su longitud sea igual a cero A0.95 corresponde a la edad
límite, en el cual el individuo alcanza una longitud correspondiente al 95 % de L∞; A es una variable
morfométrica que representa el aspecto de la aleta caudal y d es el tipo de alimentación que consume el
organismo.

17
7.2.5 Flotas pesqueras y capturas (Yi)
La flota pesquera en Yucatán por sus características se puede dividir en dos tipos: flota menor
(6 -12 m de eslora) y flota mayor (mayor a 12 m de eslora). La flota mayor dirige su esfuerzo
hacia seis tipos de pesquerías: camarón, atún, mero rojo, langosta, pulpo y de escama o de
pesca múltiple. Para el caso de la flota menor o ribereña el esfuerzo está dirigido
principalmente a la pesca de pulpo, langosta y mero (que a su vez incide sobre múltiples
especies de peces) y escama, referido el término a paces de diferentes especies (ver tabla 3)
(Moreno et al., 2002; Mexicano-Cíntora et al., 2007; DOF, 2014). Los artes y métodos de
pesca que las flotas menor y mayor utilizan varían de acuerdo a la especie objetivo,
incluyendo líneas de mano, redes y buceo con compresor entre otros. Por ejemplo en la pesca
del pulpo se utiliza en forma casi exclusiva el “gareteo” con jimbas y líneas pulperas. La
captura de langosta se ha realizado mediante buceo libre o apnea y buceo con compresor.
Para el caso del mero rojo es esencialmente multiespecífica; las capturas de esta flota son
denominadas de manera genérica como “escama” en la cual se emplean líneas de anzuelos y
anzuelos a mano y palangres (Giménez-Hurtado 2005; Mexicano-Cíntora et al., 2007).
Todos los datos de capturas de los grupos explotados fueron obtenidos de los registros
en las estadísticas pesqueras, correspondientes al periodo de 2006 a 2011 (Secretaría de
Pesca), éstas fueron promediadas para representar la época para la cual fueron incorporados
los datos de abundancia (biomasa) del modelo.
Tabla 3. Principales flotas y recurso de la plataforma continental de Yucatán
Flota Tipo Recurso
Mayor Camaronera Camarón
Atunera Atún
Escama Huachinango, rubia, pargo, mojarra, bonito, cazón, tiburón, lisa, trucha,
robalo, sardina, anchoveta, corvina, jurel, pámpano, peto, ronco
Mero Mero
Pulpo pulpo
Langosta Langosta
Menor Pulpo Pulpo
Mero Mero, rubia, pargo, mojarra, bonito
Langosta Langosta

18
Escama Sardina, sierra, cazón, tiburón, lisa, trucha de mar, jurel, ronco, corvina,
robalo, rayas, peto, anchoveta, pámpano
7.2.6 Eficiencia Ecotrófica (EEi)
El balanceo del modelo debe entenderse como una forma de compensar (para cada grupo
particular) los flujos de energía entre sus componentes más que en el sentido de “equilibrio
estático” (Christensen et al. 2000). Si la exigencia total de energía de un grupo particular
excediera su propia producción (incluyendo la energía necesaria para la respiración),
entonces el grupo se encuentra fuera de “balance”. Este grado de desbalance en los flujos, es
exteriorizado para cada grupo por el valor de Eficiencia Ecotrófica (EEi) calculado
(Christensen et al., 2000). Los valores de EEi son estimados generalmente por el modelo (a
partir de la ecuación 3) y su valor varía dentro del intervalo de 0 ≤ EEi ≤ 1.
𝐸𝐸𝑖 = 𝑌𝑖 + 𝐸𝑖 + 𝐵𝑎𝑖 + 𝑀2𝑖 ∗𝐵𝑖
𝑃𝑖⁄
En los casos donde todos los parámetros de entrada han sido estimados para todas las
presas de un grupo de predadores Ecopath puede estimar la biomasa y la tasa de
consumo/biomasa para el grupo de predadores. Similarmente, donde se conoce la P/B, B y
EEi de todas las presas de un predador j, además de la depredación total sobre estas presas, j
entonces es posible estimar directamente la B o Q/B para el predador.
Un valor de EEi mayor a la unidad, indica que la demanda total de energía en un grupo
funcional excede la producción total y el mantenimiento de ese grupo, lo cual no es posible.
El modelo fue balanceado revisando los valores de la EEi. Cuando se encontraron valores
inconsistentes de EEi entonces se realizaron los ajustes en los datos de entrada siguiendo los
criterios sugueridos por Christensen et al. (2000) hasta obtener salidas congruentes.
Generalmente los valores con mayor incertidumbre en los datos de entrada al modelo son las
dietas, por ello, el proceso de balanceo del conjunto de ecuaciones inicia ajustando la matriz
depredar-presa (Christensen et al., 2000).
Ecuación 11

19
7.2.7 Matriz de predador/presa
La construcción de la matriz predador/presa fue hecha basándose en los estudios de los
contenidos estomacales realizados en la región para las diferentes especies. En casos donde
no existía información publicada, se utilizó la dieta de la misma especie en un ecosistema
similar, o bien de otra especie similar, suponiendo que no existen cambios substanciales
(Tabla 4).

20
Tabla 4. Referencias usadas para obtener las dietas de las especies representantes de los grupos funcionales
incluidos en el modelo.
Grupo Referencias
Tiburón Cortés, 1999; Bethea et al. (2007)
Mero Brulé & Rodríguez-Canché (1993); Brulé &
Déniel (1994); Giménez-Hurtado et al. (2001)
Cazón Compagno (1984); Hoffmayer et al. (2006)
Bonito Duarte & Garcia (2004)
Peto Saloman & Naughton (1983a(
Huachinango Bradley & Bryan (1975); Camber (1955)
Pargo Riviera Arriaga (1990)
Sierra Saloman & Naughton (1983b)
Róbalo Andrade et al. (2013)
Pámpano Finucane (1969)
Ronco Arreguín-Sánchez et al. (1993)
Atún Olson & Boggs (1986)
Rubia Rincón-Sandoval et al. (2009)
Corvina García et al. (1988)
Raya Cu-Salazar et al. (2010)
Mojarra Aguirre-León & Yáñez-Arancibia (1986)
Lisa Sánchez Rueda (1995)
Trucha de mar García et al. (1988)
Jurel Saloman & Naughton (1984)
Anchoveta Vega-Cendejas (1998)
Sardina Chumba-Segura (1995); Smith (1997). Vega-
Cendejas et al. (1997)
Delfín Cabrera-Neri (2005)
Aves marinas Zetina-Rejón & Arreguín-Sánchez (2003)
Tortugas marinas Dean Hartog (1980); Zetina-Rejón & Arreguín-
Sánchez (2003)
Pulpo Solís-Ramírez (1967). Boucaud-Camou &
Boucher-Rodoni (1983). Baeza-Rojano (2013)
Calamar Boucaud-Camou & Boucher-Rodoni (1983);
Vecchione & Hand (1989)

21
Camarón rojo Reyes-Yedra & Raz-Guzmán (2012); Stoner &
Zimmerman (1988)
Langosta Colinas-Sáncehez & Briones-Fourzán (1990);
Cruz et al. (1987)
Jaiba Laughlin (1982). Raz-Guzmán & De la Lanza,
(1993)
Zooplancton Arreguín-Sánchez et al. (1993); Zetina-Rejón &
F. Arreguín-Sánchez (2003)
Microcrustaceos Arreguín-Sánchez et al. (1993)
Poliquetos Arreguín-Sánchez et al. (1993)
Equinodermos Conand (2005); Scheibling (1982); Menge (1982).

22
7.3 Incertidumbre del modelo
Mediante la rutina “Pedigrí” de Ecopath, se evaluó la calidad del modelo en función del
origen de los datos de entrada y su confiabilidad. Para cada uno de los parámetros de entrada
de cada grupo funcional se indicó su origen y confiabilidad de acuerdo a un proceso de
categorización, utilizando los valores sugeridos por el Ecopath. Con base a las opciones
seleccionadas en cada parámetro de cada grupo, se calculó un índice de pedigrí (Pe), como
ponderación de todos los índices específicos al pedigrí del parámetro. Estos valores son
usados para calcular el Pe global del modelo. Este índice varía desde 0 para aquellos modelos
de baja calidad, es decir, cuyos datos de entrada son estimados por Ecopath o asumidos de
otros modelos, hasta 1 para aquellos modelos de gran calidad, o lo que es lo mismo, cuyos
datos de entrada son muestreados adecuadamente, en la zona de estudio, referentes a las
mismas especies consideradas dentro de cada grupo funcional y por tanto, presentan una alta
precisión en sus estimaciones (Christensen et al., 2008). El índice se calcula de la siguiente
manera:
𝑃𝑒 = ∑𝐼𝑖𝑗
𝑛
𝑛
𝑖=1
donde Iij corresponde al valor del índice para cada dato de entrada y n es el número de datos
de entrada al modelo.
7.4 Atributos tróficos de los grupos funcionales
7.4.1 Nivel trófico
El nivel trófico muestra la posición en la que se encuentra ubicado un componente biótico
dentro de la red trófica de un ecosistema, Ecopath supone un nivel trófico de 1 para los
productores primarios y el detritus, mientras que para el resto de los grupos funcionales se
calcula a partir de la relación (Odum & Held, 1975) establecieron la siguiente relación:
𝑁𝑇𝑗 = 1 + ∑ 𝐷𝐶𝑖𝑗𝑛𝑗=1 ∗ 𝑁𝑇𝑗
donde NT es el nivel trófico del grupo; ∑ 𝐷𝐶𝑖𝑗𝑛𝑗=𝑖 representa el consumo promedio de las
presas en el grupo funcional i; y NTj representa el nivel trófico de la presa.
Ecuación 12
Ecuación 13

23
7.4.2 Índice de omnivoría
Para conocer en términos cuantitativos el comportamiento alimentario de los consumidores,
se utilizó el índice de omnívora (Pauly et al., 1987), este índice ofrece información sobre el
grado de especialización trófica y es calculado como la varianza de los niveles tróficos de las
presas de un predador (Christensen et al., 2005) y se expresa como:
𝐼𝑂𝑖 = ∑ (𝑁𝑇𝑗 − (𝑁𝑇𝑖 − 1))𝑛𝑗=1 ∧2∗ 𝐷𝐶𝑖𝑗
donde n es el número de grupos funcionales en el sistema, NTi es el nivel trófico; i y j de la
representan la presa y el predador, respectivamente, y DCij es la porción de las presas i en
la dieta del predador j.
Un valor de cero indica que un predador solo actúa en un nivel trófico, por lo que se le puede
considerar como especialista, mientras que un valor mayor a cero indica que el predador se
alimenta en diferentes niveles tróficos.
7.5 Atributos del ecosistema
7.5.1 Flujos del sistema
Ecopath estima los flujos totales de energía del ecosistema y cuantifica los flujos totales de
biomasa del ecosistema, los cuales sirven como indicadores de la medida del tamaño del
ecosistema (Ulanowicz 1986). La estimación de los flujos totales se realiza sumando los
flujos de los siguientes componentes: a) consumo total, b) exportación total (pesca), c)
respiración total y d) flujo total de detritus. En estas estimaciones también se incluyen los
cambios existentes entre las presas y los predadores.
Ecuación 14

24
7.5.2 Índice de conectividad
Es la proporción de los flujos existentes respecto a los flujos posibles que puedan existir en
el ecosistema. El número de posibles conexiones en un modelo de Ecopath puede ser
estimado como (N – 1)2, donde N es el número de grupos vivos en el ecosistema. El número
actual de conexiones posibles en una red alimenticia es altamente proporcional al número de
grupos en el sistema.
7.5.3 Impactos tróficos
Los impactos tróficos directos e indirectos que ejerce cada grupo sobre los demás (Ulanowicz
& Puccia, 1990) puede ser calculado a través de la matriz de impactos tróficos mixtos que
representa las interacciones directas, como son la competencia o la depredación, e indirectas,
como son las posibles cascadas tróficas y la depredación beneficiosa entre grupos funcionales
que impactan (i) y grupos impactados (j) (Paine, 1966; Menge, 1995; Pace et al., 1999). La
última situación implicaría una cascada trófica en la que el impacto directo negativo que un
predador tiene sobre una presa se ve compensado por el impacto indirecto que este predador
le causa, por ejemplo, consumiendo también otros predadores de esta presa o competidores
en el ecosistema (Christensen et al., 2008). Los valores de los elementos de la matriz van de
-1 a 1 y representan los cambios proporcionales de cada grupo funcional ante un eventual
incremento del grupo que impacta; y son proporcionales unos con respecto a otros. Los
valores de la matriz se calculan de la siguiente manera:
𝑀𝑇𝐼𝑖𝑗 = 𝐷𝐶𝑖𝑗 − 𝐹𝐶𝑖𝑗
donde MTIij es el impacto trófico mixto, DCij es la matriz de dieta, y FCij es la proporción de
las pérdidas dadas por la depredación sobre j debido al predador.
7.5.4 Indicadores de crecimiento y desarrollo
Ulanowicz (1986) propone los conceptos de Ascendencia (A), Capacidad de desarrollo (C)
y el Overhead (O), para describir atributos de organización del ecosistema. De acuerdo a a
este autor, estas son las características que definen el desarrollo y crecimiento de un sistema,
que a su vez están en íntima relación con los atributos propuestos por Odum (1969).
Ecuación 15

25
La Ascendencia es un atributo del sistema que es utilizado como un indicador para
medir el crecimiento y desarrollo del ecosistema en términos de flujos de energía (Ulanowicz,
1986). Además representa tanto el tamaño como la organización de un ecosistema, ya que
refleja la magnitud de la energía que está fluyendo dentro del ecosistema y a su vez representa
la densidad de enlaces que implica una capacidad de auto-organización para encaminarse a
una etapa de madurez y de pleno desarrollo. Se calcula de la siguiente manera:
𝐴 = ∑ 𝑇𝑖𝑗𝑙𝑜𝑔𝑖,𝑗 (𝑇𝑖𝑗𝑇∘∘
𝑇𝑖,𝑇,𝑗)
donde Tij es el flujo de energía del grupo funcional i (presa)a otro grupo funcional j
(predador) y T○○ representa los flujos totales de energía el cual se expresa de la siguiente
forma:
𝑇∘∘ = ∑ 𝑇𝑖𝑗𝑖,𝑗
La capacidad de desarrollo es el límite máximo teórico de la Ascendencia. Este fue calculado
de la siguiente forma:
𝐶 = ∑ 𝑇𝑖𝑗𝑙𝑜𝑔 (𝑇𝑖𝑗
𝑇∘∘)𝑖,𝑗
El ovehead es un indicador que cuantifica el potencial de reserva que tiene el sistema para
responder en caso de perturbaciones externas (Ulanowicz, 1986) y es la diferencia entre A y
C, y se representa a partir de la siguiente ecuación:
𝑂 = − ∑ 𝑇𝑖𝑗
𝑖,𝑗
𝑙𝑜𝑔 (𝑇𝑖𝑗
2
𝑇𝑖⋅𝑇∙𝑗)
Ecuación 16
Ecuación 17
Ecuación 18
Ecuación 19

26
7.6 Indicadores estructurales
Se usaron índices de centralidad con base en la topología de redes, para complementar los
indicadores de nivel de grupo funcional propuestos por el modelo Ecopath. Con base en
rutinas construida en el paquete igraph para R® versión 3.2.2 se estimaron los índice de grado
(Di), cercanía (Cci) e intermediación (BCi) para cada grupo funcional, a partir de la matriz de
consumo obtenida del modelo Ecopath.
El Di es la medida más simple de centralidad, dada por el número total de conexiones
(Izquierdo & Hanneman, 2006), tanto como predador o como presa para cada uno de los
grupos funcionales incluidos en el modelo. El índice de grado que se utilizó en el presente
trabajo fue ponderado por la intensidad de los flujos, y se calculó como:
𝐷𝑖 = 𝐷𝑖𝑛 𝑖 ∗ 𝑤𝑖 + 𝐷𝑜𝑢𝑡 𝑖 ∗ 𝑤𝑖
donde Din i y Dout i son los flujos tróficos de entrada y salida del grupo i, respectivamente; y
wi es la magnitud de las conexiones en términos de los flujos tróficos.
El Cci, se define como la capacidad de un vértice o nodo para transmitir su efecto a
todos los elementos de la red. El supuesto de este índice es que, los miembros más centrales
de la red son aquellos que se encuentran a una menor distancia del resto. Los valores más
altos de este índice indican que existe una mayor facilidad de acceso al resto de los miembros
de la red (Borgatti et al., 2002). El índice se calcula de la siguiente manera:
𝐶𝑐𝑖 = 𝑁−1
∑ 𝑑𝑖𝑗𝑁𝑗=1
donde N corresponde al número de nodos (grupos funcionales) en la red; ∑ =𝑁𝑗=1 𝑑𝑖𝑗 es la
suma de las distancias geodésicas entre nodos en la red.
El índice de intermediación mide el número de veces que aparece un nodo (i) en las
rutas cortas entre cada par de nodos j y k (Molina et al. 2006). Es una medida que da una
aproximación de la importancia de un grupo funcional como conector dentro de la red y su
eliminación pudiera causar que muchos pares de nodos se desconectaran o que al menos la
conexión entre ellos fuera más lejana. Los nodos con un valor alto de intermediación son
importantes y claves por ser los más centrales de la red o aquellos que vinculan subgrupos o
bloques diferentes (Izquierdo & Hanneman, 2006). Se calcula como:
Ecuación 20
Ecuación 21
Ecuación 22

27
𝐵𝐶𝑖 =
∑ 𝑔𝑖𝑘(𝑖)𝑗<𝑘𝑔𝑖𝑘
⁄
[(𝑁 − 1)(𝑁 − 2)
2⁄ ]
donde: gjk es el número de rutas cortas en la red; gjk (i) es el número de rutas cortas en las
cuales i aparece; N es el número de nodos totales.
7.7 Simulación temporal
El análisis de los cambios de biomasa a través del tiempo se realizó a través de la simulación
dinámica medio del modelo Ecosim (Walters & Pauly 1997), donde los cambios de la
biomasa se expresan a través de una serie de ecuaciones diferenciales acopladas. En la
Ecosim la ecuación 3 es transformada a su forma dinámica y expresada como sigue:
𝑑𝐵𝑖
𝑑𝑡= 𝑓(𝐵) − 𝑀0 − 𝐹𝑖𝐵𝑖 − ∑ 𝐶𝑖𝑗 (𝐵𝑖>𝐵𝑗)
𝑛
𝑗=1
donde i son los grupos funcionales, dB/dt representa la tasa de cambio de biomasa en el
tiempo; f(B) es la tasa de producción, y es una función de Bi cuando la especie i es un
productor primario, y para consumidores 𝑓(𝐵) = 𝑔𝑖 ∑ 𝐶𝑗𝑖𝑛𝑗=1 (𝐵𝑖>𝐵𝑗), donde gi = Bi (Pi/Bi) que
representa la eficiencia neta de crecimiento, M0 representa la mortalidad distinta de pesca y
depredación; Fi representa la mortalidad por pesca, y Cij (Bi, Bj) es la función usada para
predecir del consumo y representa la probabilidad de encuentros entre presa y predador, así
como el comportamiento fisiológico (tal como la saciedad).
La ecuación básica de Ecosim (ecuación 23) permite, entre otros, incorporar factores
forzantes que afectan la producción del grupo especificado, permitiendo la propagación de
la señal a través de la red trófica.
De acuerdo con Arreguín-Sánchez (2012) y Arreguín-Sánchez et al. (2015) los
efectos del cambios en temperatura a través de la Oscilación Multidecadal del Atlántico
(AMO por sus siglas en inglés), han influido sustancialmente en la dinámica del Banco de
Campeche. Con el objetivo de investigar el papel del mero en el ecosistema, incluida las
diferentes condiciones del ecosistema derivadas de los efectos ambientales, se utilizó el AMO
como factor forzante de la producción primaria, registrando los cambios en el tiempo de las
Ecuación 23

28
biomasas de los grupos funcionales y de los atributos del ecosistema. Para ello la información
sobre el AMO fue obtenida de la NOAA (National Oceanic Atmospheric Administration)
(http://www.esrl.noaa.gov/psd/data/correlation/amo.data) para el periodo 1961 a 2011.
7.8 Interpretación del papel del mero
Con la finalidad de identificar el papel del mero en el ecosistema, las salidas del modelo
dinámico (periodo 1961 a 2011) se emplearon relacionar la razón A/C (ascendencia sobre la
capacidad, como medida de orden del ecosistema) y la biomasa de los grupos funcionales.
Se identificaron grupos con los diferentes patrones (tendencias) obtenidos, los cuales se
relacionaron con diferentes atributos de los grupos funcionales tales como: nivel trófico,
consumo biomasa (Q/B). De igual manera con índices de flujo de la red trófica como son:
ascendencia, capacidad de desarrollo del sistema y overhead; a partir de los cuales se
construyeron indicadores a nivel grupo funcional relativos al orden y resiliencia. Con esta
información se realizaron el análisis estadístico de componentes principales con el objetivo
de identificar la asociación entre los atributos de cada grupo y del ecosistema, y en particular
los relativos al mero, con el fin de identificar a través de ellos su papel en el ecosistema.
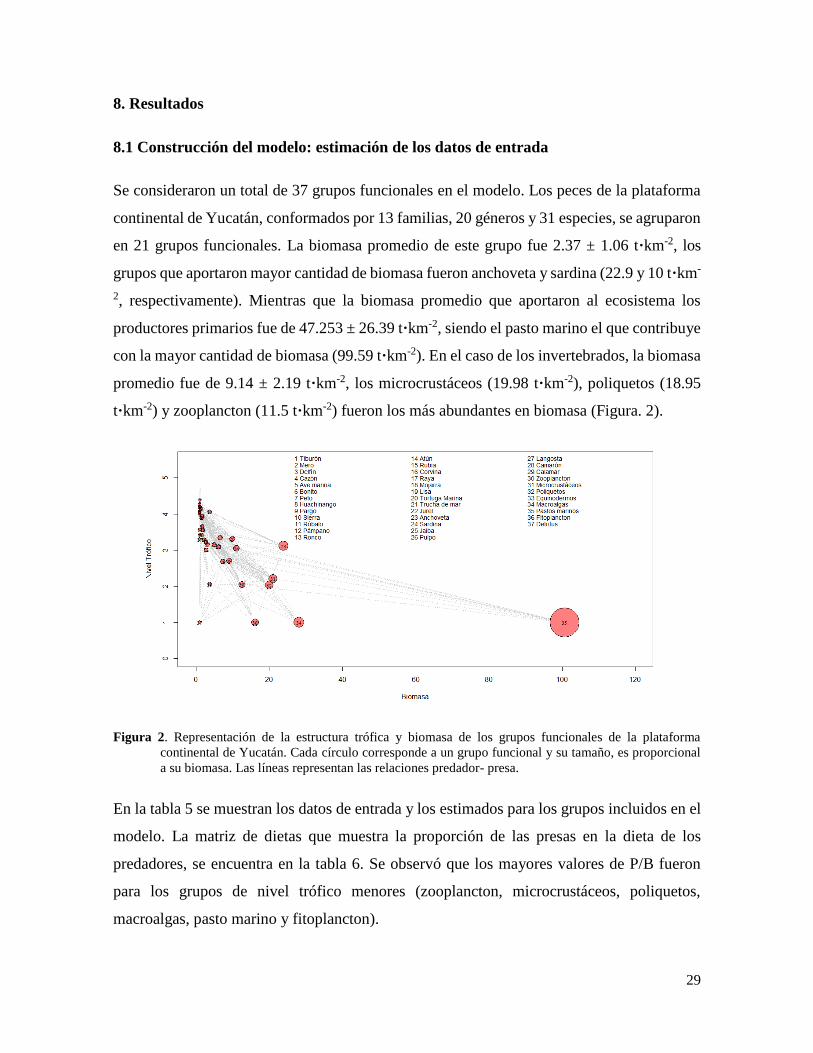
29
8. Resultados
8.1 Construcción del modelo: estimación de los datos de entrada
Se consideraron un total de 37 grupos funcionales en el modelo. Los peces de la plataforma
continental de Yucatán, conformados por 13 familias, 20 géneros y 31 especies, se agruparon
en 21 grupos funcionales. La biomasa promedio de este grupo fue 2.37 ± 1.06 t·km-2, los
grupos que aportaron mayor cantidad de biomasa fueron anchoveta y sardina (22.9 y 10 t·km-
2, respectivamente). Mientras que la biomasa promedio que aportaron al ecosistema los
productores primarios fue de 47.253 ± 26.39 t·km-2, siendo el pasto marino el que contribuye
con la mayor cantidad de biomasa (99.59 t·km-2). En el caso de los invertebrados, la biomasa
promedio fue de 9.14 ± 2.19 t·km-2, los microcrustáceos (19.98 t·km-2), poliquetos (18.95
t·km-2) y zooplancton (11.5 t·km-2) fueron los más abundantes en biomasa (Figura. 2).
Figura 2. Representación de la estructura trófica y biomasa de los grupos funcionales de la plataforma
continental de Yucatán. Cada círculo corresponde a un grupo funcional y su tamaño, es proporcional
a su biomasa. Las líneas representan las relaciones predador- presa.
En la tabla 5 se muestran los datos de entrada y los estimados para los grupos incluidos en el
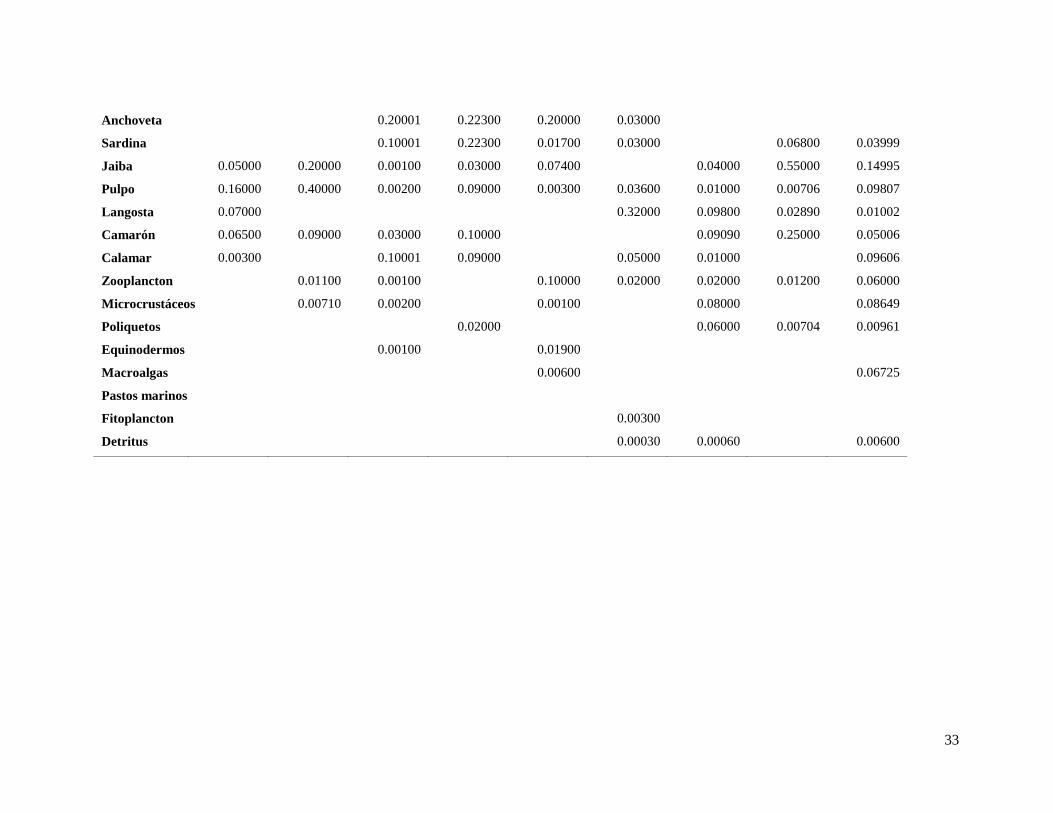
modelo. La matriz de dietas que muestra la proporción de las presas en la dieta de los
predadores, se encuentra en la tabla 6. Se observó que los mayores valores de P/B fueron
para los grupos de nivel trófico menores (zooplancton, microcrustáceos, poliquetos,
macroalgas, pasto marino y fitoplancton).
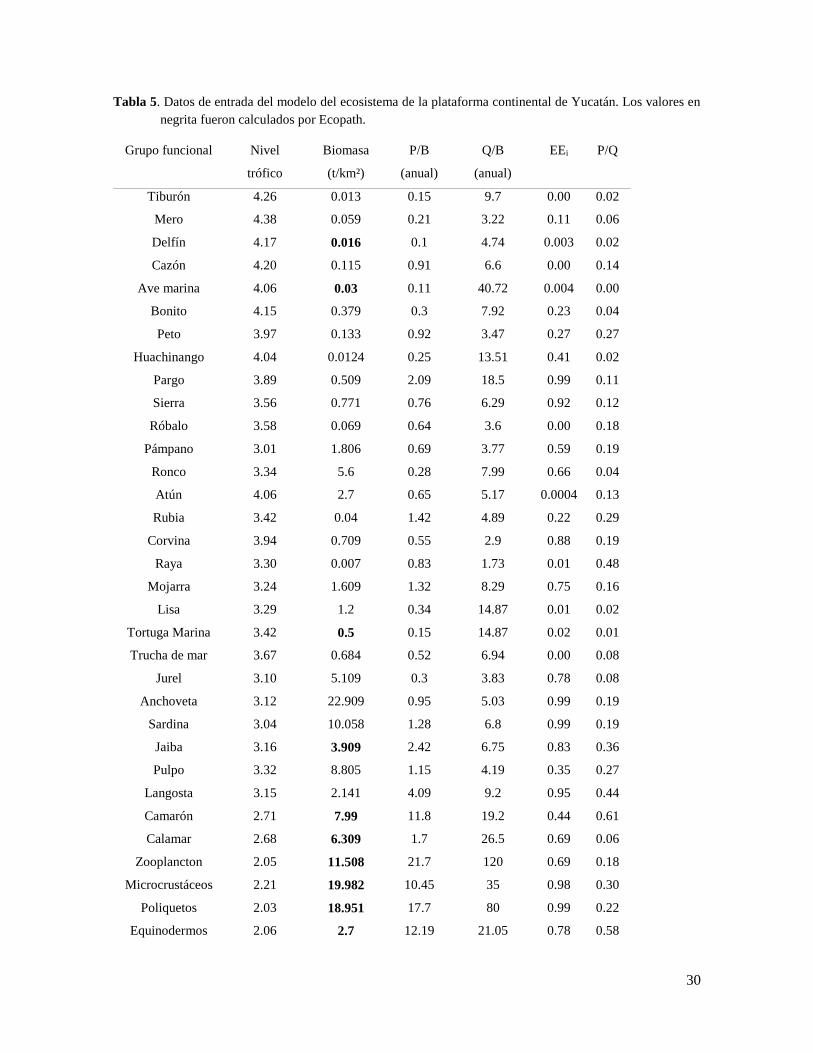
30
Tabla 5. Datos de entrada del modelo del ecosistema de la plataforma continental de Yucatán. Los valores en
negrita fueron calculados por Ecopath.
Grupo funcional Nivel
trófico
Biomasa
(t/km²)
P/B
(anual)
Q/B
(anual)
EEi P/Q
Tiburón 4.26 0.013 0.15 9.7 0.00 0.02
Mero 4.38 0.059 0.21 3.22 0.11 0.06
Delfín 4.17 0.016 0.1 4.74 0.003 0.02
Cazón 4.20 0.115 0.91 6.6 0.00 0.14
Ave marina 4.06 0.03 0.11 40.72 0.004 0.00
Bonito 4.15 0.379 0.3 7.92 0.23 0.04
Peto 3.97 0.133 0.92 3.47 0.27 0.27
Huachinango 4.04 0.0124 0.25 13.51 0.41 0.02
Pargo 3.89 0.509 2.09 18.5 0.99 0.11
Sierra 3.56 0.771 0.76 6.29 0.92 0.12
Róbalo 3.58 0.069 0.64 3.6 0.00 0.18
Pámpano 3.01 1.806 0.69 3.77 0.59 0.19
Ronco 3.34 5.6 0.28 7.99 0.66 0.04
Atún 4.06 2.7 0.65 5.17 0.0004 0.13
Rubia 3.42 0.04 1.42 4.89 0.22 0.29
Corvina 3.94 0.709 0.55 2.9 0.88 0.19
Raya 3.30 0.007 0.83 1.73 0.01 0.48
Mojarra 3.24 1.609 1.32 8.29 0.75 0.16
Lisa 3.29 1.2 0.34 14.87 0.01 0.02
Tortuga Marina 3.42 0.5 0.15 14.87 0.02 0.01
Trucha de mar 3.67 0.684 0.52 6.94 0.00 0.08
Jurel 3.10 5.109 0.3 3.83 0.78 0.08
Anchoveta 3.12 22.909 0.95 5.03 0.99 0.19
Sardina 3.04 10.058 1.28 6.8 0.99 0.19
Jaiba 3.16 3.909 2.42 6.75 0.83 0.36
Pulpo 3.32 8.805 1.15 4.19 0.35 0.27
Langosta 3.15 2.141 4.09 9.2 0.95 0.44
Camarón 2.71 7.99 11.8 19.2 0.44 0.61
Calamar 2.68 6.309 1.7 26.5 0.69 0.06
Zooplancton 2.05 11.508 21.7 120 0.69 0.18
Microcrustáceos 2.21 19.982 10.45 35 0.98 0.30
Poliquetos 2.03 18.951 17.7 80 0.99 0.22
Equinodermos 2.06 2.7 12.19 21.05 0.78 0.58
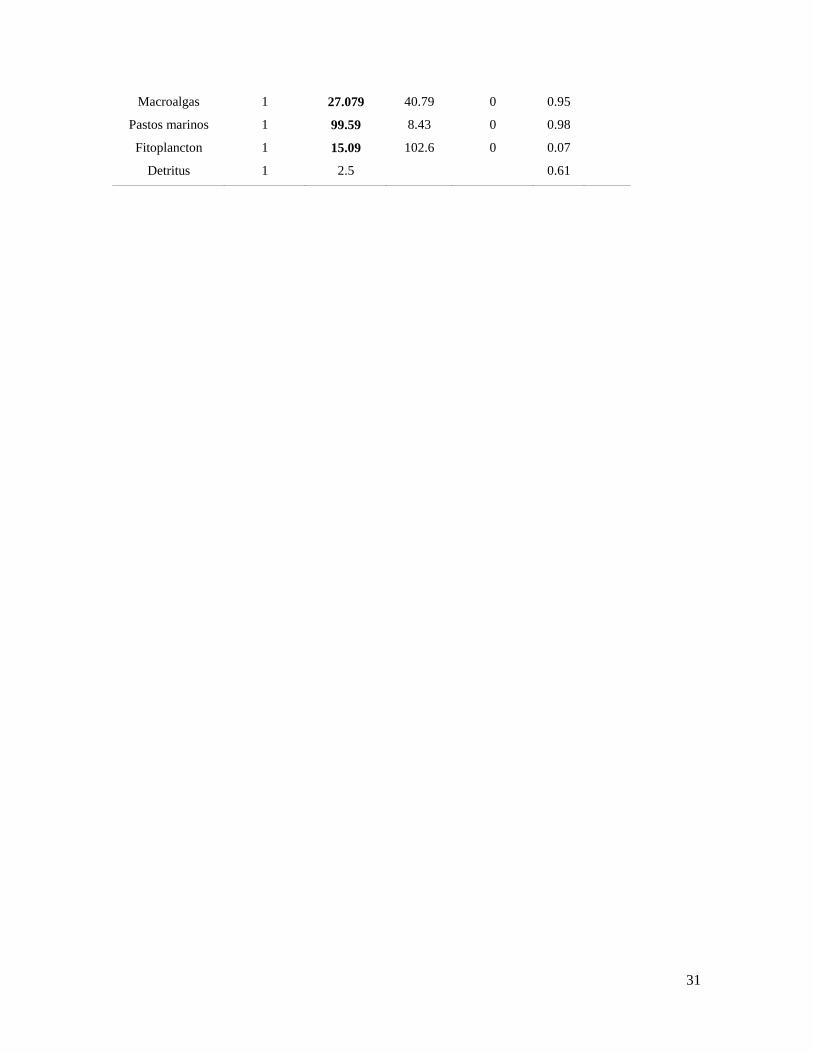
31
Macroalgas 1 27.079 40.79 0 0.95
Pastos marinos 1 99.59 8.43 0 0.98
Fitoplancton 1 15.09 102.6 0 0.07
Detritus 1 2.5
0.61
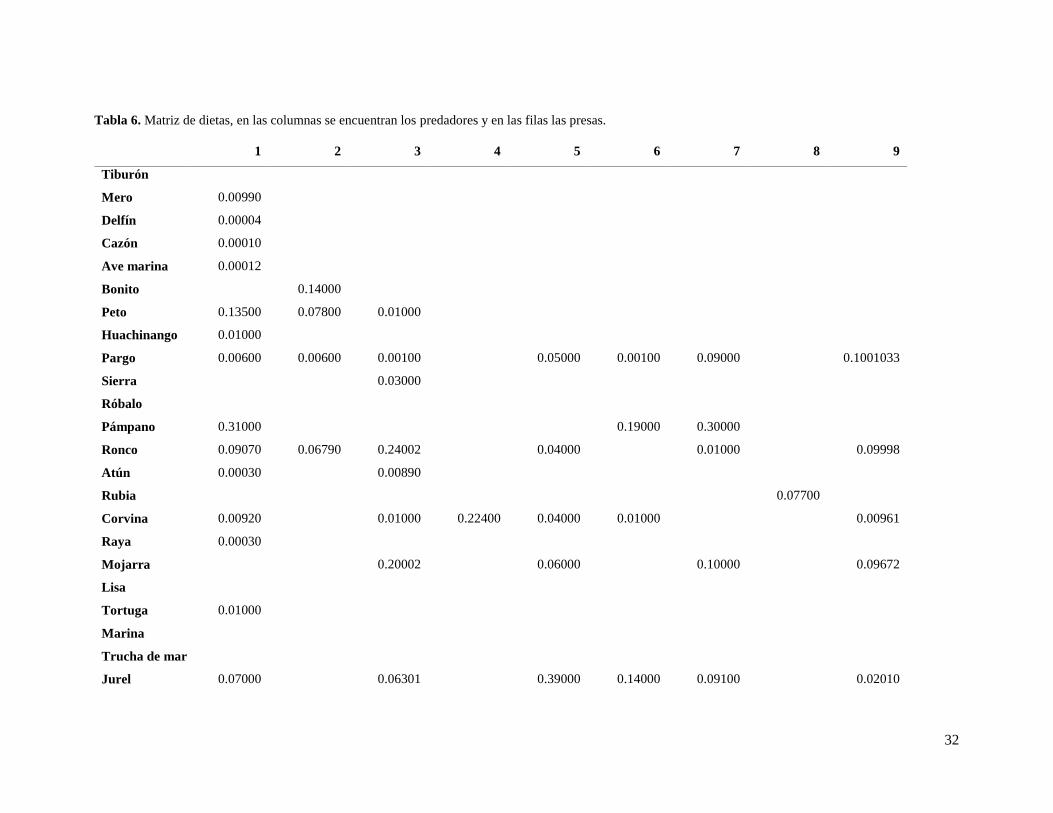
32
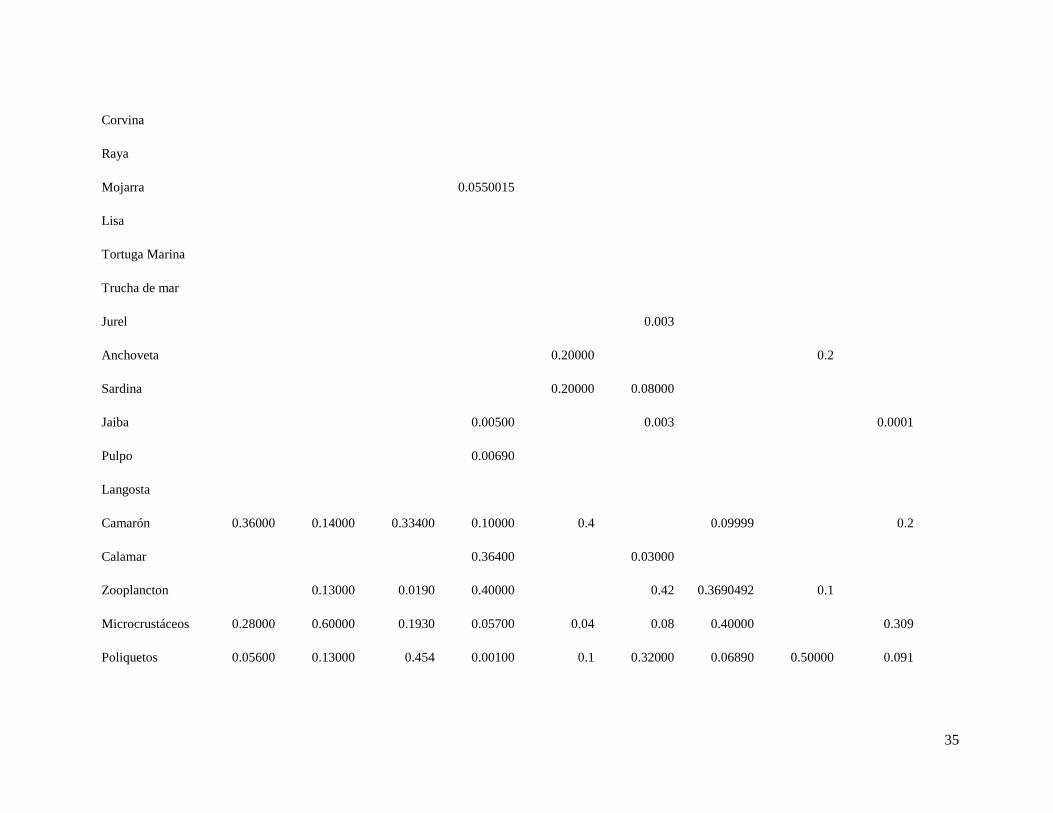
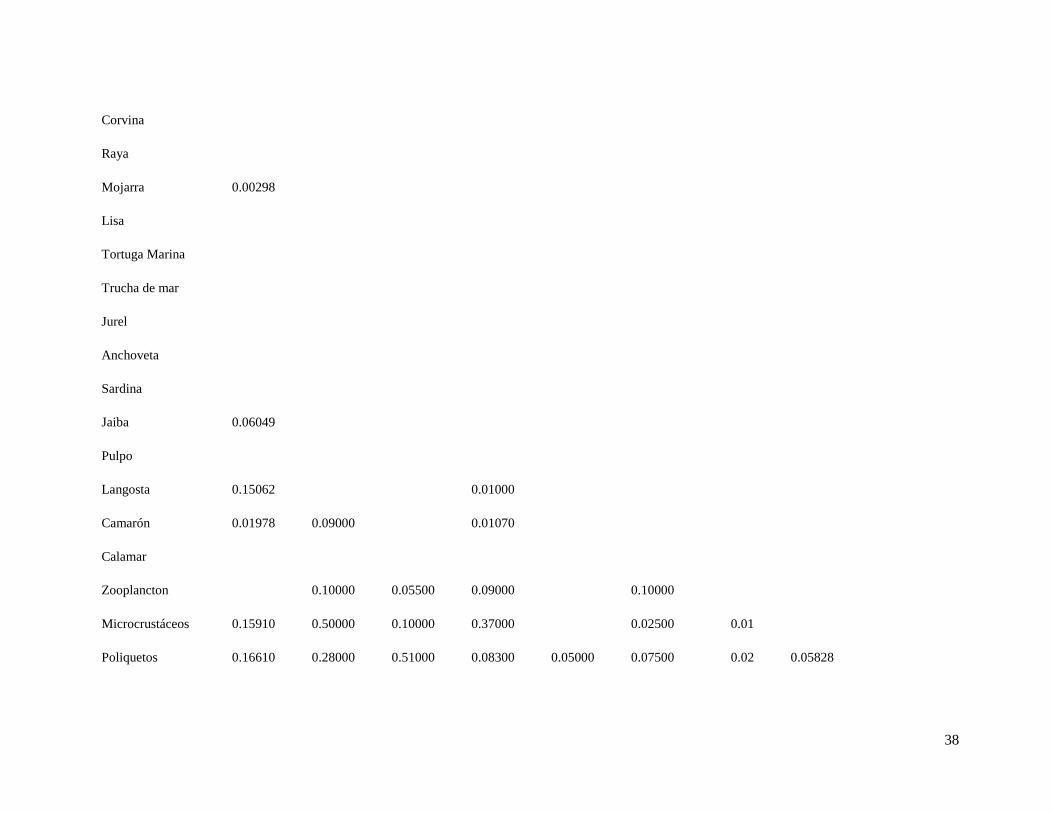
Tabla 6. Matriz de dietas, en las columnas se encuentran los predadores y en las filas las presas.
1 2 3 4 5 6 7 8 9
Tiburón
Mero 0.00990
Delfín 0.00004
Cazón 0.00010
Ave marina 0.00012
Bonito
0.14000
Peto 0.13500 0.07800 0.01000
Huachinango 0.01000
Pargo 0.00600 0.00600 0.00100
0.05000 0.00100 0.09000
0.1001033
Sierra
0.03000
Róbalo
Pámpano 0.31000
0.19000 0.30000
Ronco 0.09070 0.06790 0.24002
0.04000
0.01000
0.09998
Atún 0.00030
0.00890
Rubia
0.07700
Corvina 0.00920
0.01000 0.22400 0.04000 0.01000
0.00961
Raya 0.00030
Mojarra
0.20002
0.06000
0.10000
0.09672
Lisa
Tortuga
Marina
0.01000
Trucha de mar
Jurel 0.07000
0.06301
0.39000 0.14000 0.09100
0.02010
33
Anchoveta
0.20001 0.22300 0.20000 0.03000
Sardina
0.10001 0.22300 0.01700 0.03000
0.06800 0.03999
Jaiba 0.05000 0.20000 0.00100 0.03000 0.07400
0.04000 0.55000 0.14995
Pulpo 0.16000 0.40000 0.00200 0.09000 0.00300 0.03600 0.01000 0.00706 0.09807
Langosta 0.07000
0.32000 0.09800 0.02890 0.01002
Camarón 0.06500 0.09000 0.03000 0.10000
0.09090 0.25000 0.05006
Calamar 0.00300
0.10001 0.09000
0.05000 0.01000
0.09606
Zooplancton 0.01100 0.00100
0.10000 0.02000 0.02000 0.01200 0.06000
Microcrustáceos 0.00710 0.00200
0.00100
0.08000
0.08649
Poliquetos
0.02000
0.06000 0.00704 0.00961
Equinodermos
0.00100
0.01900
Macroalgas
0.00600
0.06725
Pastos marinos
Fitoplancton
0.00300
Detritus
0.00030 0.00060
0.00600

34
Tabla 6. Continuación
17 18 19 20 21 22 23 24 25
Tiburón
Mero
Delfín
Cazón
Ave marina
Bonito
Peto
Huachinango
Pargo
Sierra
Róbalo
Pámpano
Ronco
Atún
Rubia
35
Corvina
Raya
Mojarra
0.0550015
Lisa
Tortuga Marina
Trucha de mar
Jurel
0.003
Anchoveta
0.20000
0.2
Sardina
0.20000 0.08000
Jaiba
0.00500
0.003
0.0001
Pulpo
0.00690
Langosta
Camarón 0.36000 0.14000 0.33400 0.10000 0.4
0.09999
0.2
Calamar
0.36400
0.03000
Zooplancton 0.13000 0.0190 0.40000
0.42 0.3690492 0.1
Microcrustáceos 0.28000 0.60000 0.1930 0.05700 0.04 0.08 0.40000
0.309
Poliquetos 0.05600 0.13000 0.454 0.00100 0.1 0.32000 0.06890 0.50000 0.091

36
Equinodermos 0.27000
0.0001
0.33
Macroalgas
0.01 0.06
0.039
Pastos marinos
0.00100
0.02 0.07
Fitoplancton
0.06100 0.011 0.001
Detritus 0.034
0.00300 0.0508792 0.14000

37
Tabla 6. Continuación
26 27 28 29 30 31 32 33
Tiburón
Mero
Delfín
Cazón
Ave marina
Bonito
Peto
Huachinango
Pargo
Sierra
Róbalo
Pámpano
Ronco
Atún
Rubia
38
Corvina
Raya
Mojarra 0.00298
Lisa
Tortuga Marina
Trucha de mar
Jurel
Anchoveta
Sardina
Jaiba 0.06049
Pulpo
Langosta 0.15062
0.01000
Camarón 0.01978 0.09000
0.01070
Calamar
Zooplancton 0.10000 0.05500 0.09000
0.10000
Microcrustáceos 0.15910 0.50000 0.10000 0.37000
0.02500 0.01
Poliquetos 0.16610 0.28000 0.51000 0.08300 0.05000 0.07500 0.02 0.05828

39
Equinodermos 0.43093
0.00650
Macroalgas
0.14650
0.47300 0.10000 0.20000 0.00030
Pastos marinos 0.02800 0.15040
0.47700 0.20000
Fitoplancton 0.00200 0.03800 0.42988
0.49985
Detritus 0.01000
0.50000 0.77000 0.44187
40
8.2 Balanceo del modelo
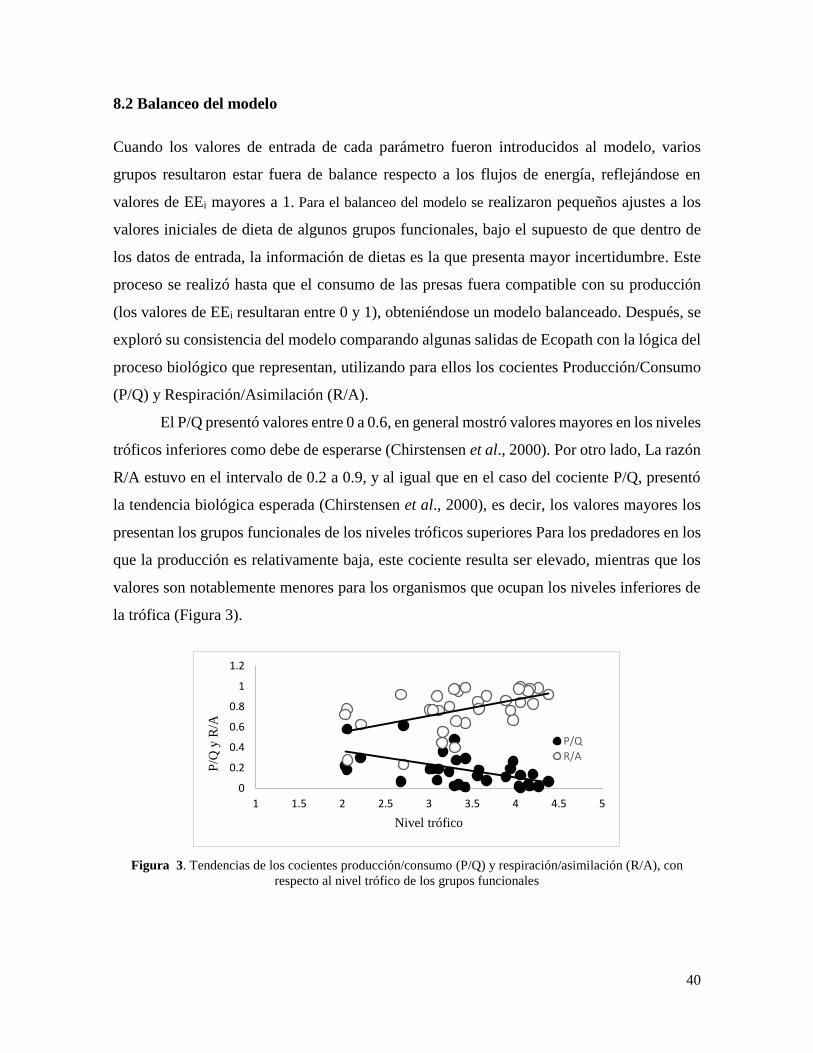
Cuando los valores de entrada de cada parámetro fueron introducidos al modelo, varios
grupos resultaron estar fuera de balance respecto a los flujos de energía, reflejándose en
valores de EEi mayores a 1. Para el balanceo del modelo se realizaron pequeños ajustes a los
valores iniciales de dieta de algunos grupos funcionales, bajo el supuesto de que dentro de
los datos de entrada, la información de dietas es la que presenta mayor incertidumbre. Este
proceso se realizó hasta que el consumo de las presas fuera compatible con su producción
(los valores de EEi resultaran entre 0 y 1), obteniéndose un modelo balanceado. Después, se
exploró su consistencia del modelo comparando algunas salidas de Ecopath con la lógica del
proceso biológico que representan, utilizando para ellos los cocientes Producción/Consumo
(P/Q) y Respiración/Asimilación (R/A).
El P/Q presentó valores entre 0 a 0.6, en general mostró valores mayores en los niveles
tróficos inferiores como debe de esperarse (Chirstensen et al., 2000). Por otro lado, La razón
R/A estuvo en el intervalo de 0.2 a 0.9, y al igual que en el caso del cociente P/Q, presentó
la tendencia biológica esperada (Chirstensen et al., 2000), es decir, los valores mayores los
presentan los grupos funcionales de los niveles tróficos superiores Para los predadores en los
que la producción es relativamente baja, este cociente resulta ser elevado, mientras que los
valores son notablemente menores para los organismos que ocupan los niveles inferiores de
la trófica (Figura 3).
Figura 3. Tendencias de los cocientes producción/consumo (P/Q) y respiración/asimilación (R/A), con
respecto al nivel trófico de los grupos funcionales
0
0.2
0.4
0.6
0.8
1
1.2
1 1.5 2 2.5 3 3.5 4 4.5 5
P/Q
y R
/A
Nivel trófico
P/Q
R/A

41
El estimador del Pedigrí (Pe) fue de 0.503, el cual nos indica que el modelo presenta
una calidad media con respecto al origen de los datos de entrada al modelo.
8.3 Atributos de los grupos funcionales
Se obtuvieron cuatro niveles tróficos de acuerdo a lo estimado por Ecopath, los predadores
tope del ecosistema fuero el mero rojo y tiburón, con NT de 4.38 y 4.26, respectivamente. En
general los peces ocuparon los niveles cuatro y tres, el grupo de aves marina y delfín ocupan
el nivel cuatro (4.17 y 4.06, respectivamente). El pulpo (3.32), jaiba (3.16) y langosta (3.15)
se ubicaron en el nivel trófico tres, mientras que el resto de los grupos estuvieron ubicados
en el nivel dos.
El índice de omnívoría calculado, sugiere que los grupos son en mayor o menor
consumidores especialistas; los grupos rubia (0.54), pargo (0.53) y sardina (0.44) fueron los
que presentaron los valores más altos, por lo cual se les puede considerar a estos grupos como
consumidores generalistas.
Los grupos de nivel tróficos menores (principalmente zooplancton y fitoplancton)
estuvieron dirigidos mayormente hacia detritus. Los grupos camarón, equinodermos, raya y
langosta fueron los que presentaron la mayor eficiencia neta del ecosistema (0.76, 0.72, 0.59
y 0.55, respectivamente) (Tabla 7).
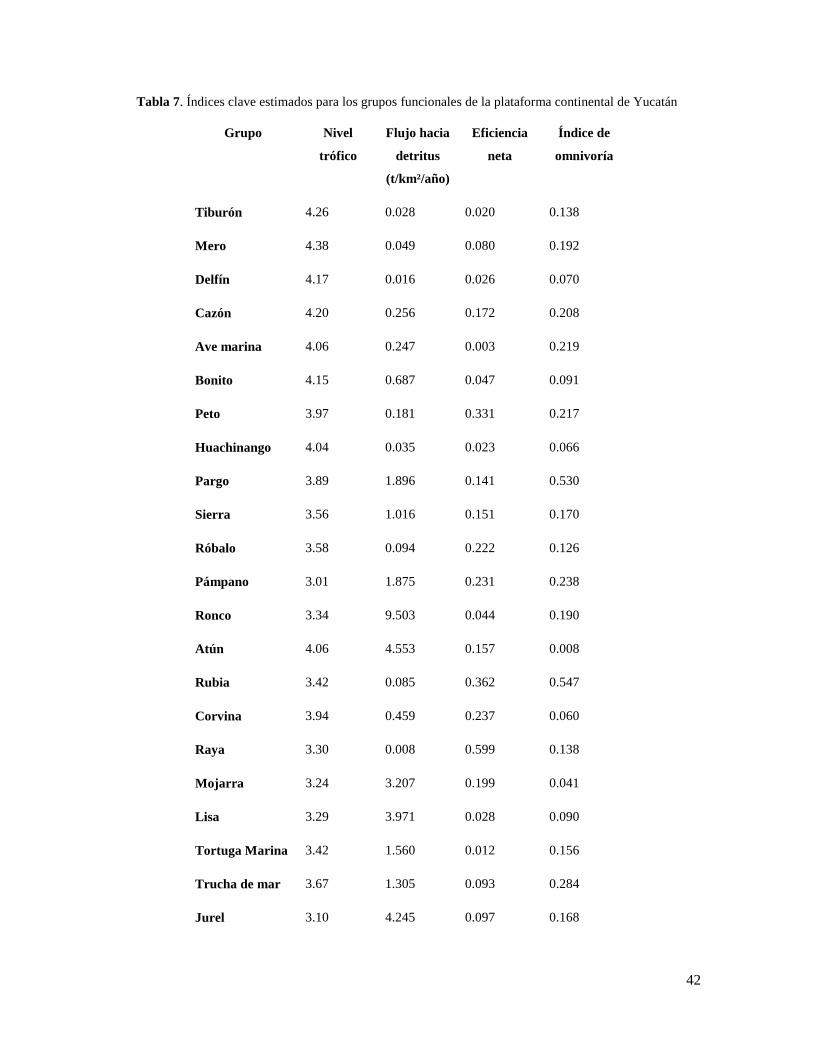
42
Tabla 7. Índices clave estimados para los grupos funcionales de la plataforma continental de Yucatán
Grupo Nivel
trófico
Flujo hacia
detritus
(t/km²/año)
Eficiencia
neta
Índice de
omnivoría
Tiburón 4.26 0.028 0.020 0.138
Mero 4.38 0.049 0.080 0.192
Delfín 4.17 0.016 0.026 0.070
Cazón 4.20 0.256 0.172 0.208
Ave marina 4.06 0.247 0.003 0.219
Bonito 4.15 0.687 0.047 0.091
Peto 3.97 0.181 0.331 0.217
Huachinango 4.04 0.035 0.023 0.066
Pargo 3.89 1.896 0.141 0.530
Sierra 3.56 1.016 0.151 0.170
Róbalo 3.58 0.094 0.222 0.126
Pámpano 3.01 1.875 0.231 0.238
Ronco 3.34 9.503 0.044 0.190
Atún 4.06 4.553 0.157 0.008
Rubia 3.42 0.085 0.362 0.547
Corvina 3.94 0.459 0.237 0.060
Raya 3.30 0.008 0.599 0.138
Mojarra 3.24 3.207 0.199 0.041
Lisa 3.29 3.971 0.028 0.090
Tortuga Marina 3.42 1.560 0.012 0.156
Trucha de mar 3.67 1.305 0.093 0.284
Jurel 3.10 4.245 0.097 0.168
43
Anchoveta 3.12 23.293 0.236 0.117
Sardina 3.04 13.849 0.235 0.447
Jaiba 3.16 6.864 0.448 0.159
Pulpo 3.32 14.007 0.343 0.215
Langosta 3.15 4.349 0.555 0.074
Camarón 2.71 83.329 0.768 0.253
Calamar 2.68 36.784 0.080 0.360
Zooplancton 2.05 353.441 0.226 0.050
Microcrustáceos 2.21 143.039 0.373 0.182
Poliquetos 2.03 306.644 0.276 0.034
Equinodermos 2.06 18.532 0.723 0.058
Macroalgas 1 52.040 - 0
Pastos marinos 1 12.969 - 0
Fitoplancton 1 1439.522 - 0
Detritus 1 0 0 0.445
8.4 Atributos del ecosistema
8.4.1 Flujos del sistema
En la tabla 8 se muestran el resumen de los principales atributos básicos del ecosistema. Se
puede observar que la producción total en términos de flujos fue 10428.09 t/km2/año, del cual
el 42.11 % fue consumida dentro del ecosistema, 9.19 % fue exportada (incluyendo la pesca),
el 24.04 % fue utilizada en los procesos respiratorios y el resto (23.39 %) fue enviado a
detritus. Por otra parte la producción primaria neta fue 985.39 t/km2/año, y la relación
Producción primaria total/biomasa total 12.51.
El cociente producción primaria total/respiración total (PPT/RT) expresa la porción
de energía que puede ser utilizada (producción primaria) en el ecosistema en relación al gasto

44
metabólico que realizan los componentes vivientes (respiración) (Williams & Follows,
2003). De acuerdo a Odum (1969) esté cociente resulta útil para establecer la medida de
madurez en un ecosistema. Al disminuir el valor de esta relación el sistema se vuelve más
maduro (PPT/RT < 1). Para el ecosistema de la plataforma continental de Yucatán PPT/RT
fue de 1.39, indicando que el ecosistema se encontró en estado de inmadurez, debido a que
este índice fue mayor a uno (PPT/RT > 1). La Producción Primaria Neta del Sistema (PPNS)
resulto ser un poco menor a la Producción Primaria Neta Total (PPNT).
El índice de omnivoría del sistema fue calculado como un valor promedio del índice
de omnivoría de todos los grupos funcionales, el cual fue de 0.17, este valor fue relativamente
bajo, lo cual podría indicar que la red trófica del ecosistema de la plataforma continental de
Yucatán es lineal, es decir, los grupos que conforman el ecosistema tiene preferencia por
algunos grupos en particular (Tabla 8).

45
Tabla 8. Resumen de estadísticos del ecosistema de la plataforma continental de Yucatán
Parámetro Valor Unidades
Suma de los flujos de consumo 4391.799 t/km2/año
Suma de los flujos de exportación 985.3907 t/km2/año
Suma de los flujos de respiración 2506.94 t/km2/año
Suma de los flujos al detritus 2543.958 t/km2/año
Flujo total del sistema 10428.09 t/km2/año
Suma total de la producción 4498.83 t/km2/año
Nivel trófico medio de la captura 2.512347
Eficiencia gruesa del sistema 1.01E-07
Producción primaria neta calculada 3492.33 t/km2/año
Producción primaria total/respiración total 1.393065
Producción neta del sistema 985.3904 t/km2/año
Producción primaria total/biomasa total 12.51299
Biomasa total/flujos totales 0.0267639 t/km2/año
Biomasa total (excluyendo al detritus) 279.0963 t/km2
Capturas totales 0.000352401 t/km2/año
Índice de conectividad 0.2030888
Índice de Omnívoría del sistema 0.1762386
8.4.2 Índice de conectividad
El índice de conectividad para la trama trófica del ecosistema de la plataforma continental de
Yucatán, fue de 0.20, lo cual corresponde a un 20 % de uniones tróficas del total posibles
entre los grupos funcionales.

46
8.4.3 Impactos tróficos mixtos
En general, el análisis muestra que la mayoría de los grupos funcionales presentaron un
impacto negativo sobre sí mismos, interpretado como efecto densodependiente, por lo cual
cada grupo depende directamente de su biomasa. Los grupos de detritus, productores
primarios, microcrustáceos, zooplancton y camarón ejercen impactos positivos en la mayoría
de los grupos del ecosistema, principalmente en los consumidores de niveles tróficos de
segundo y tercer nivel como anchovetas, jaiba, mojarra y raya. Mientras que los grupos como
róbalo, huachinango, tiburón ejercen un alto impacto negativo sobre los grupos de lisa, rubia
y huachinango (Figura 4).
Por otro lado, los grupos funcionales que presentaron los mayores impactos negativos
fueron las mojarra, corvina, poliquetos, anchoveta, jurel, ronco camarón y macroalgas.
Mientras que los grupos con menos impactos negativos fueron el mero rojo, delfín sierra,
huachinango, tortuga marina, pastos marinos y fitoplancton.
Con respecto al mero rojo, los grupos que estuvieron impactados de manera negativa
en mayor proporción con respecto a otros son: bonito, peto, huachinango y raya, mientras
que los grupos con impactos positivos fueron: tiburón, sierra, pámpano, y jurel (Fig. 4).
También fue posible observar la importancia que tienen las flotas atuneras y flota
mayor y menor de mero rojo, sobre el grupo de delfín y tiburón, respectivamente (Fig. 4.)
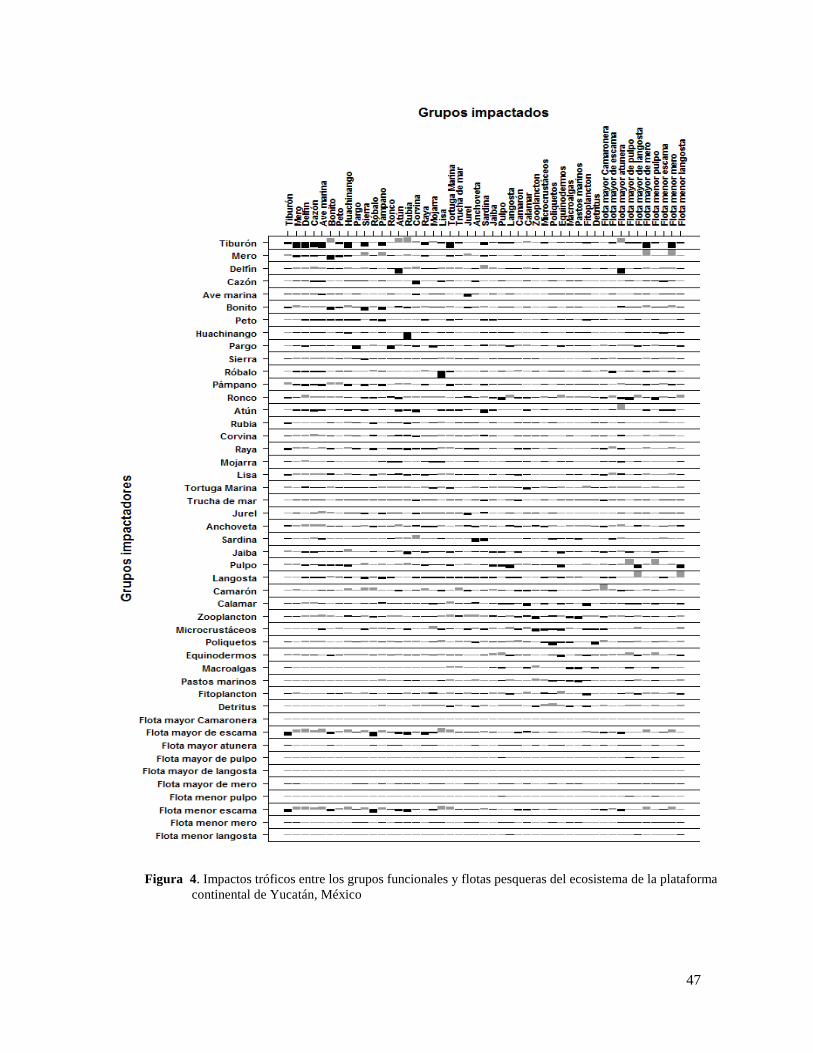
47
Figura 4. Impactos tróficos entre los grupos funcionales y flotas pesqueras del ecosistema de la plataforma
continental de Yucatán, México
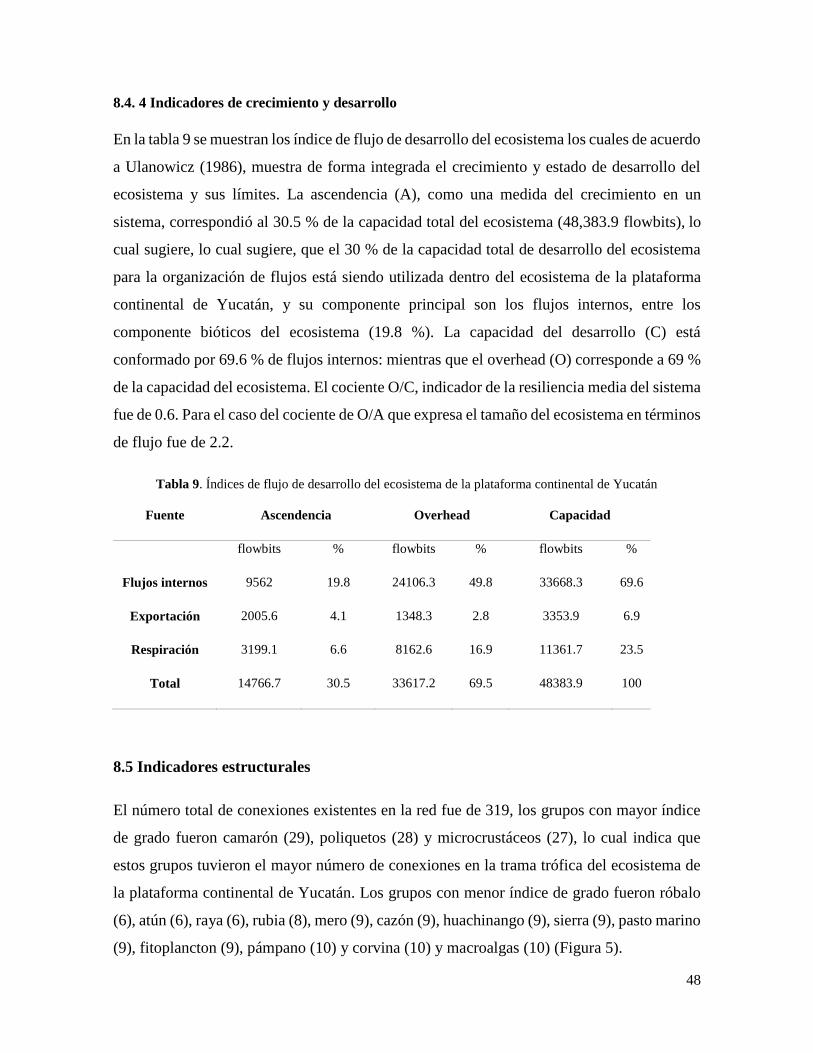
48
8.4. 4 Indicadores de crecimiento y desarrollo
En la tabla 9 se muestran los índice de flujo de desarrollo del ecosistema los cuales de acuerdo
a Ulanowicz (1986), muestra de forma integrada el crecimiento y estado de desarrollo del
ecosistema y sus límites. La ascendencia (A), como una medida del crecimiento en un
sistema, correspondió al 30.5 % de la capacidad total del ecosistema (48,383.9 flowbits), lo
cual sugiere, lo cual sugiere, que el 30 % de la capacidad total de desarrollo del ecosistema
para la organización de flujos está siendo utilizada dentro del ecosistema de la plataforma
continental de Yucatán, y su componente principal son los flujos internos, entre los
componente bióticos del ecosistema (19.8 %). La capacidad del desarrollo (C) está
conformado por 69.6 % de flujos internos: mientras que el overhead (O) corresponde a 69 %
de la capacidad del ecosistema. El cociente O/C, indicador de la resiliencia media del sistema
fue de 0.6. Para el caso del cociente de O/A que expresa el tamaño del ecosistema en términos
de flujo fue de 2.2.
Tabla 9. Índices de flujo de desarrollo del ecosistema de la plataforma continental de Yucatán
Fuente Ascendencia Overhead Capacidad
flowbits % flowbits % flowbits %
Flujos internos 9562 19.8 24106.3 49.8 33668.3 69.6
Exportación 2005.6 4.1 1348.3 2.8 3353.9 6.9
Respiración 3199.1 6.6 8162.6 16.9 11361.7 23.5
Total 14766.7 30.5 33617.2 69.5 48383.9 100
8.5 Indicadores estructurales
El número total de conexiones existentes en la red fue de 319, los grupos con mayor índice
de grado fueron camarón (29), poliquetos (28) y microcrustáceos (27), lo cual indica que
estos grupos tuvieron el mayor número de conexiones en la trama trófica del ecosistema de
la plataforma continental de Yucatán. Los grupos con menor índice de grado fueron róbalo
(6), atún (6), raya (6), rubia (8), mero (9), cazón (9), huachinango (9), sierra (9), pasto marino
(9), fitoplancton (9), pámpano (10) y corvina (10) y macroalgas (10) (Figura 5).
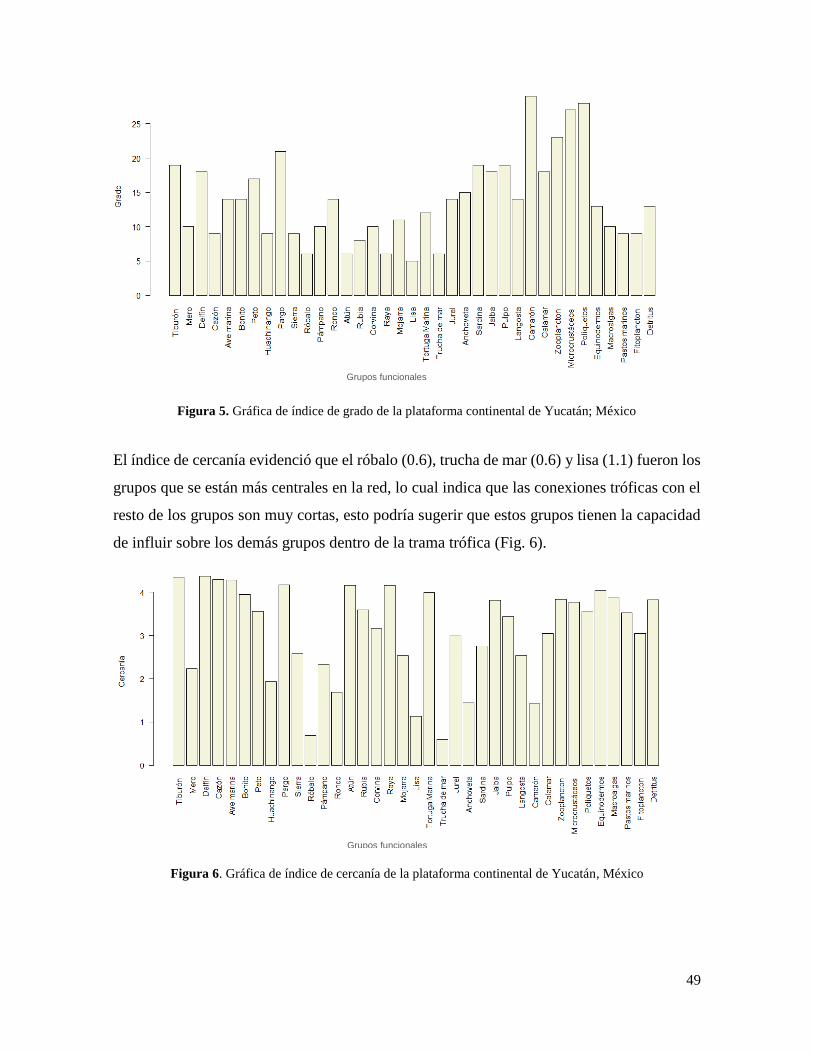
49
Figura 5. Gráfica de índice de grado de la plataforma continental de Yucatán; México
El índice de cercanía evidenció que el róbalo (0.6), trucha de mar (0.6) y lisa (1.1) fueron los
grupos que se están más centrales en la red, lo cual indica que las conexiones tróficas con el
resto de los grupos son muy cortas, esto podría sugerir que estos grupos tienen la capacidad
de influir sobre los demás grupos dentro de la trama trófica (Fig. 6).
Figura 6. Gráfica de índice de cercanía de la plataforma continental de Yucatán, México
Grupos funcionales
Grupos funcionales
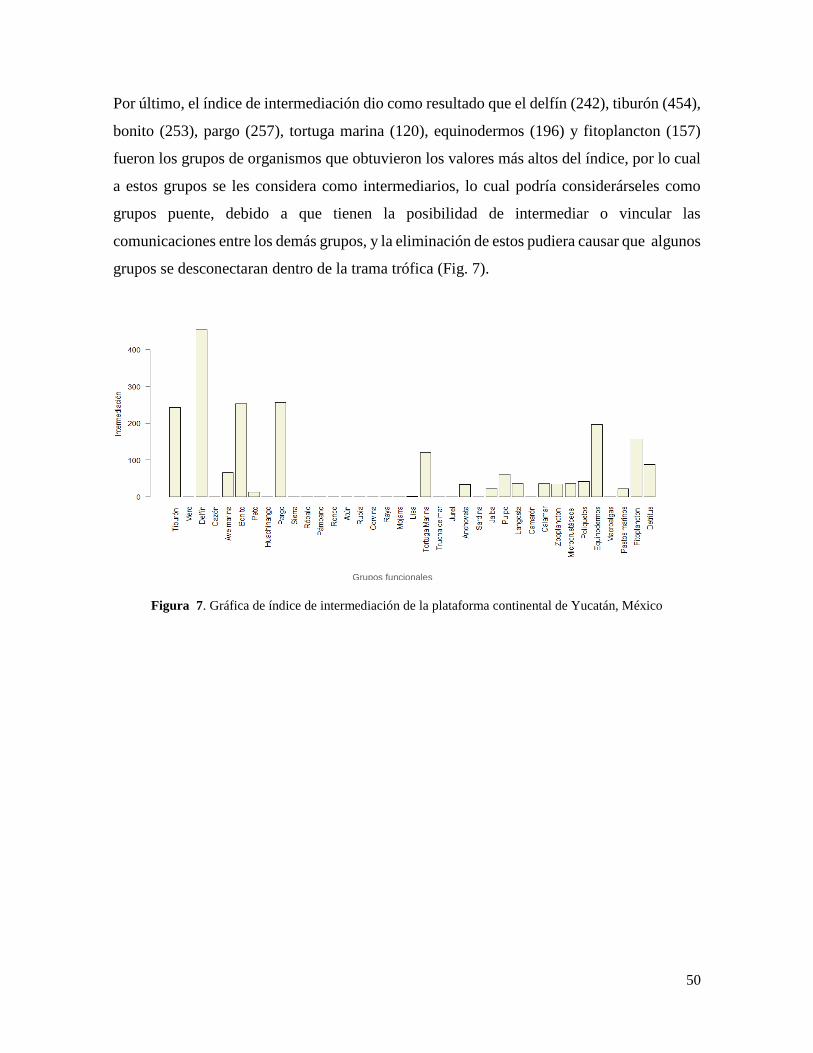
50
Por último, el índice de intermediación dio como resultado que el delfín (242), tiburón (454),
bonito (253), pargo (257), tortuga marina (120), equinodermos (196) y fitoplancton (157)
fueron los grupos de organismos que obtuvieron los valores más altos del índice, por lo cual
a estos grupos se les considera como intermediarios, lo cual podría considerárseles como
grupos puente, debido a que tienen la posibilidad de intermediar o vincular las
comunicaciones entre los demás grupos, y la eliminación de estos pudiera causar que algunos
grupos se desconectaran dentro de la trama trófica (Fig. 7).
Figura 7. Gráfica de índice de intermediación de la plataforma continental de Yucatán, México
Grupos funcionales
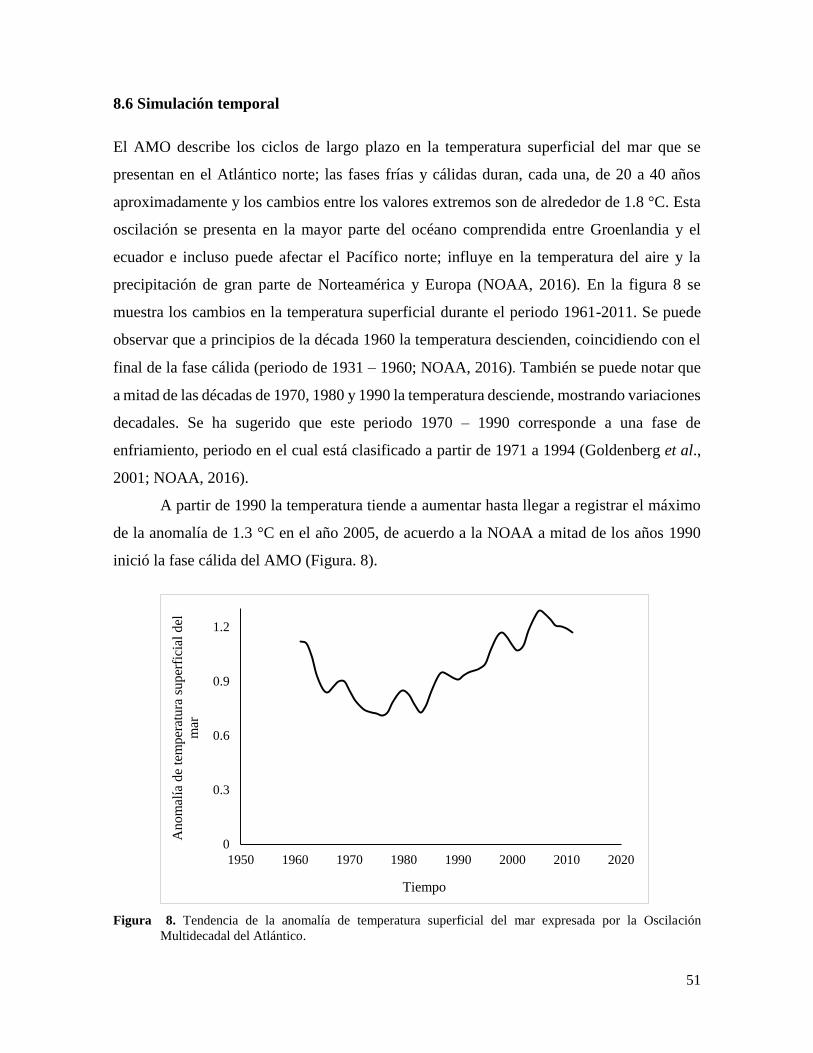
51
8.6 Simulación temporal
El AMO describe los ciclos de largo plazo en la temperatura superficial del mar que se
presentan en el Atlántico norte; las fases frías y cálidas duran, cada una, de 20 a 40 años
aproximadamente y los cambios entre los valores extremos son de alrededor de 1.8 °C. Esta
oscilación se presenta en la mayor parte del océano comprendida entre Groenlandia y el
ecuador e incluso puede afectar el Pacífico norte; influye en la temperatura del aire y la
precipitación de gran parte de Norteamérica y Europa (NOAA, 2016). En la figura 8 se
muestra los cambios en la temperatura superficial durante el periodo 1961-2011. Se puede
observar que a principios de la década 1960 la temperatura descienden, coincidiendo con el
final de la fase cálida (periodo de 1931 – 1960; NOAA, 2016). También se puede notar que
a mitad de las décadas de 1970, 1980 y 1990 la temperatura desciende, mostrando variaciones
decadales. Se ha sugerido que este periodo 1970 – 1990 corresponde a una fase de
enfriamiento, periodo en el cual está clasificado a partir de 1971 a 1994 (Goldenberg et al.,
2001; NOAA, 2016).
A partir de 1990 la temperatura tiende a aumentar hasta llegar a registrar el máximo
de la anomalía de 1.3 °C en el año 2005, de acuerdo a la NOAA a mitad de los años 1990
inició la fase cálida del AMO (Figura. 8).
Figura 8. Tendencia de la anomalía de temperatura superficial del mar expresada por la Oscilación
Multidecadal del Atlántico.
0
0.3
0.6
0.9
1.2
1950 1960 1970 1980 1990 2000 2010 2020
Anom
alía
de
tem
per
atura
super
fici
al d
el
mar
Tiempo

52
8.7 Interpretación del papel del mero
De las tendencias entre el orden del ecosistema (A/C) y biomasa de los grupos funcionales
resultantes de las simulaciones temporales con Ecosim se observó, en general un patrón en
el cual al decrecer la biomasa decrece el orden del sistema (Figura 9). También se pudo
observar que existe un mínimo de biomasa a partir del cual, con poco cambio en la biomasa
el orden incrementa para posteriormente alcanzar un máximo a partir del cual la biomasa
tiende a decrecer. De este patrón generalizado, puede inferirse que es un patrón de carácter
cíclico, no obstante presenta las variantes mostradas en la figura 9 para los 37 grupos
funcionales, los cuales se ilustran con los grupos funcionales de mero, sierra, corvina y pastos
marinos.
En el caso del mero, el patrón tiende a ser mejor definido con un descenso gradual de la
biomasa y del orden del ecosistema, por casi dos décadas (1960 - 1980). Posteriormente, la
biomasa permanece baja y el orden aumenta, también por casi dos décadas (1960 - 1980) y
finalmente, la siguiente década (1990) el orden se mantiene estable con un aumento de
biomasa. Para posteriormente seguir aumentando la biomasa con un ligero decremento del
orden en el sistema (Fig. 9 a). Este patrón corresponde con cambios graduales en la biomasa
que siguen la tendencia de factor forzante.
Para los patrones representados por sierra (Fig. 9 b) y corvina (Fig. 9 c), se observa mayor
variabilidad en la tendencia del cambio de la biomasa, en relación al nivel trófico del grupo,
con la acotación de que en cada caso el patrón de la trayectoria de la relación orden versus
biomasa se va observando menos definido. Finalmente, para los pastos marinos, el patrón
general se presenta pero de manera más estrecha que en los casos anteriores (Fig. 9 d).
Se puede observar que en el primer patrón, a medida que la biomasa disminuye a través del
tiempo el orden también sigue esta tendencia; una vez que la biomasa alcanza un valor
mínimo, el orden del sistema aumenta, sin cambio en la biomasa, por cerca de dos décadas
(1960 - 1980). Eventualmente la biomasa aumenta gradualmente y el orden del sistema se
estabiliza, posteriormente tiende a decrecer de manera gradual. De manera particular, con
respecto al patrón de la figura 9 a, b, correspondiente al mero, en este se encuentran
principalmente los grupos de predadores como son de tiburón, mero, delfín, aves marinas,
bonito, peto, huachinango, sierra, róbalo, ronco, raya, y jurel, los grupos trucha de mar, lisa
y tortuga marina también estuvieron caracterizados dentro de este patrón. En el segundo caso

53
(Fig. 9 c, d), se encuentran grupos de nivel trófico intermedio y primario, como son cazón,
pargo, sierra, pámpano, atún, rubia, mojarra, anchoveta, sardina, jaiba, pulpo, langosta,
camarón, calamar, zooplancton, microcrustáceos, poliquetos, equinodermos, macroalgas,
fitoplancton y detritus. El tercer patrón (Fig. 9 e, f) únicamente es representado por la corvina.
En este sentido es fácil observar, por la tendencia de la biomasa en el tiempo, que dicho
patrón está fuertemente asociado a la variabilidad intrínseca del grupo reflejada por el
modelo. Por último, el cuarto patrón de variación es representado únicamente por pastos
marinos.
Para el análisis de componentes principales se obtuvo una clasificación de 68. 17 %,
usando dos componentes principales (Tabla 10), en el componente principal 1 estuvieron
clasificados los atributos Ascendencia, Capacidad y Orden (Tabla 11), lo cual resulta
congruente dada la relación entre estos atributos (ver ecuación 16 y 17). El componte
principal 2 estuvo conformado por los atributos biomasa y nivel trófico (Tabla 11). Este
análisis demostró que los atributos Ascendencia, Capacidad y Orden son lo que tienen una
mayor correlación dentro del ecosistema. De igual manera los atributos Ascendencia y
Capacidad fueron los que obtuvieron mayor correlación con la mayoría de los grupos
funcionales, incluyendo al mero rojo; cabe destacar que para este grupo los atributos nivel
trófico, Q/B y A/C también tuvieron una correlación (Tabla. 12), esta correlación podría
deberse a las características biológicas del grupo (como: depredador tope, tipo de hábito
alimenticio generalista, responde rápido ante cualquier perturbación en el ecosistema).
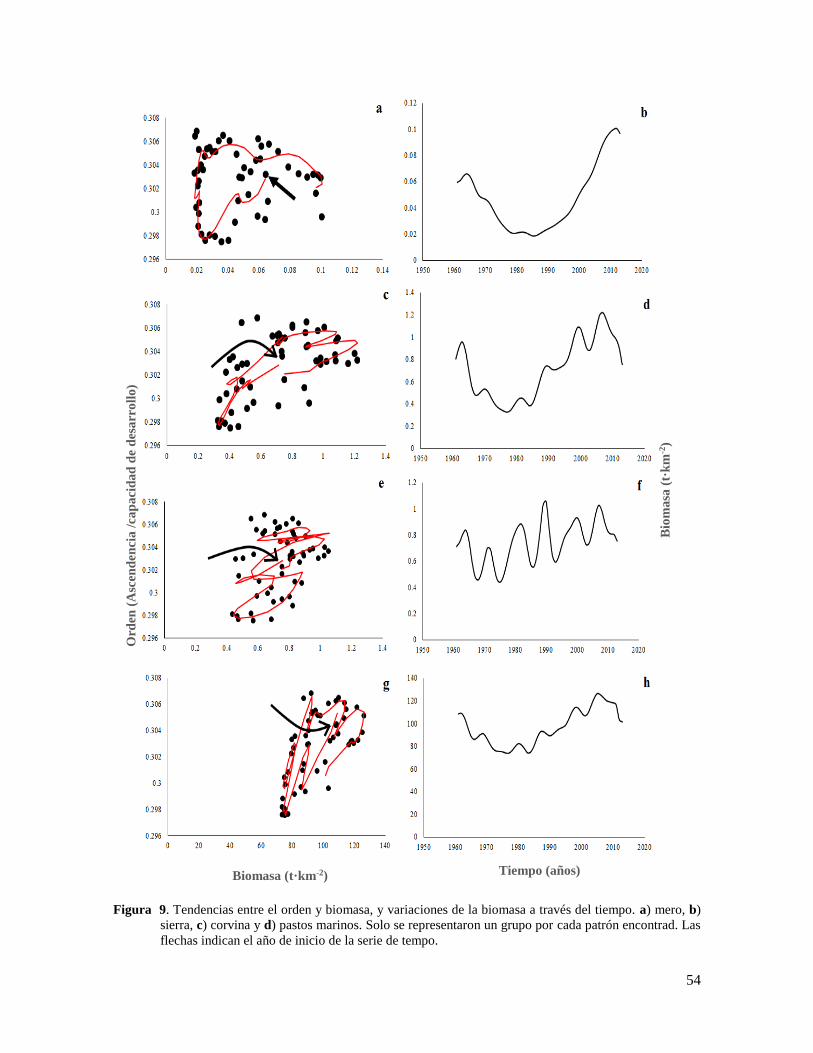
54
Figura 9. Tendencias entre el orden y biomasa, y variaciones de la biomasa a través del tiempo. a) mero, b)
sierra, c) corvina y d) pastos marinos. Solo se representaron un grupo por cada patrón encontrad. Las
flechas indican el año de inicio de la serie de tempo.
Bio
ma
sa (
t·k
m-2
)
Ord
en (
Asc
end
en
cia
/ca
pa
cid
ad
de
des
arr
oll
o)
Tiempo (años) Biomasa (t·km-2)
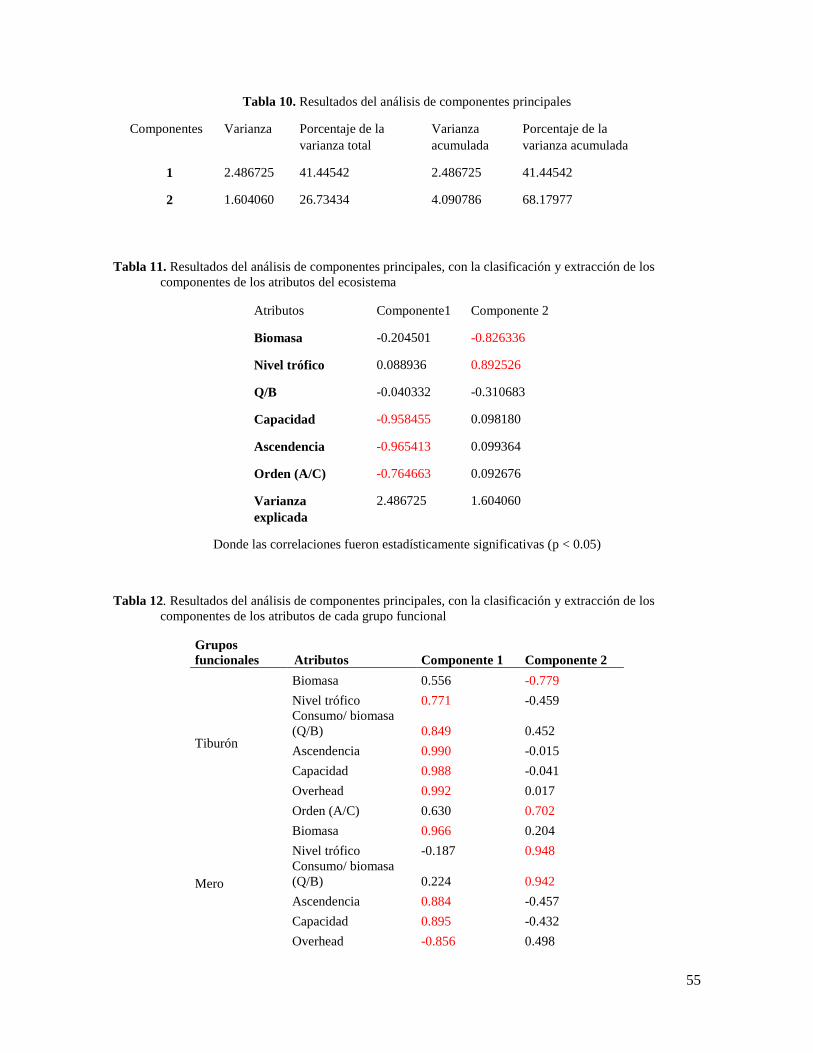
55
Tabla 10. Resultados del análisis de componentes principales
Componentes Varianza Porcentaje de la
varianza total
Varianza
acumulada
Porcentaje de la
varianza acumulada
1 2.486725 41.44542 2.486725 41.44542
2 1.604060 26.73434 4.090786 68.17977
Tabla 11. Resultados del análisis de componentes principales, con la clasificación y extracción de los
componentes de los atributos del ecosistema
Atributos Componente1 Componente 2
Biomasa -0.204501 -0.826336
Nivel trófico 0.088936 0.892526
Q/B -0.040332 -0.310683
Capacidad -0.958455 0.098180
Ascendencia -0.965413 0.099364
Orden (A/C) -0.764663 0.092676
Varianza
explicada
2.486725 1.604060
Donde las correlaciones fueron estadísticamente significativas (p < 0.05)
Tabla 12. Resultados del análisis de componentes principales, con la clasificación y extracción de los
componentes de los atributos de cada grupo funcional
Grupos
funcionales Atributos Componente 1 Componente 2
Tiburón
Biomasa 0.556 -0.779
Nivel trófico 0.771 -0.459
Consumo/ biomasa
(Q/B) 0.849 0.452
Ascendencia 0.990 -0.015
Capacidad 0.988 -0.041
Overhead 0.992 0.017
Orden (A/C) 0.630 0.702
Mero
Biomasa 0.966 0.204
Nivel trófico -0.187 0.948
Consumo/ biomasa
(Q/B) 0.224 0.942
Ascendencia 0.884 -0.457
Capacidad 0.895 -0.432
Overhead -0.856 0.498
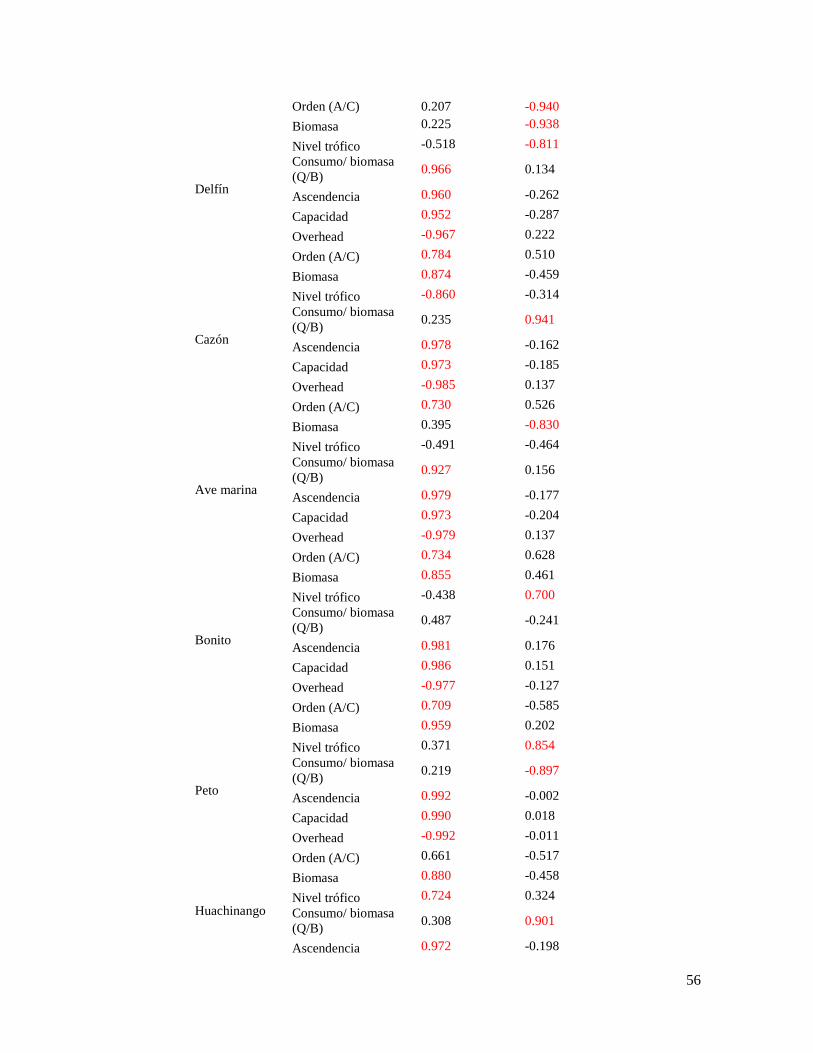
56
Orden (A/C) 0.207 -0.940
Delfín
Biomasa 0.225 -0.938
Nivel trófico -0.518 -0.811
Consumo/ biomasa
(Q/B) 0.966 0.134
Ascendencia 0.960 -0.262
Capacidad 0.952 -0.287
Overhead -0.967 0.222
Orden (A/C) 0.784 0.510
Cazón
Biomasa 0.874 -0.459
Nivel trófico -0.860 -0.314
Consumo/ biomasa
(Q/B) 0.235 0.941
Ascendencia 0.978 -0.162
Capacidad 0.973 -0.185
Overhead -0.985 0.137
Orden (A/C) 0.730 0.526
Ave marina
Biomasa 0.395 -0.830
Nivel trófico -0.491 -0.464
Consumo/ biomasa
(Q/B) 0.927 0.156
Ascendencia 0.979 -0.177
Capacidad 0.973 -0.204
Overhead -0.979 0.137
Orden (A/C) 0.734 0.628
Bonito
Biomasa 0.855 0.461
Nivel trófico -0.438 0.700
Consumo/ biomasa
(Q/B) 0.487 -0.241
Ascendencia 0.981 0.176
Capacidad 0.986 0.151
Overhead -0.977 -0.127
Orden (A/C) 0.709 -0.585
Peto
Biomasa 0.959 0.202
Nivel trófico 0.371 0.854
Consumo/ biomasa
(Q/B) 0.219 -0.897
Ascendencia 0.992 -0.002
Capacidad 0.990 0.018
Overhead -0.992 -0.011
Orden (A/C) 0.661 -0.517
Huachinango
Biomasa 0.880 -0.458
Nivel trófico 0.724 0.324
Consumo/ biomasa
(Q/B) 0.308 0.901
Ascendencia 0.972 -0.198
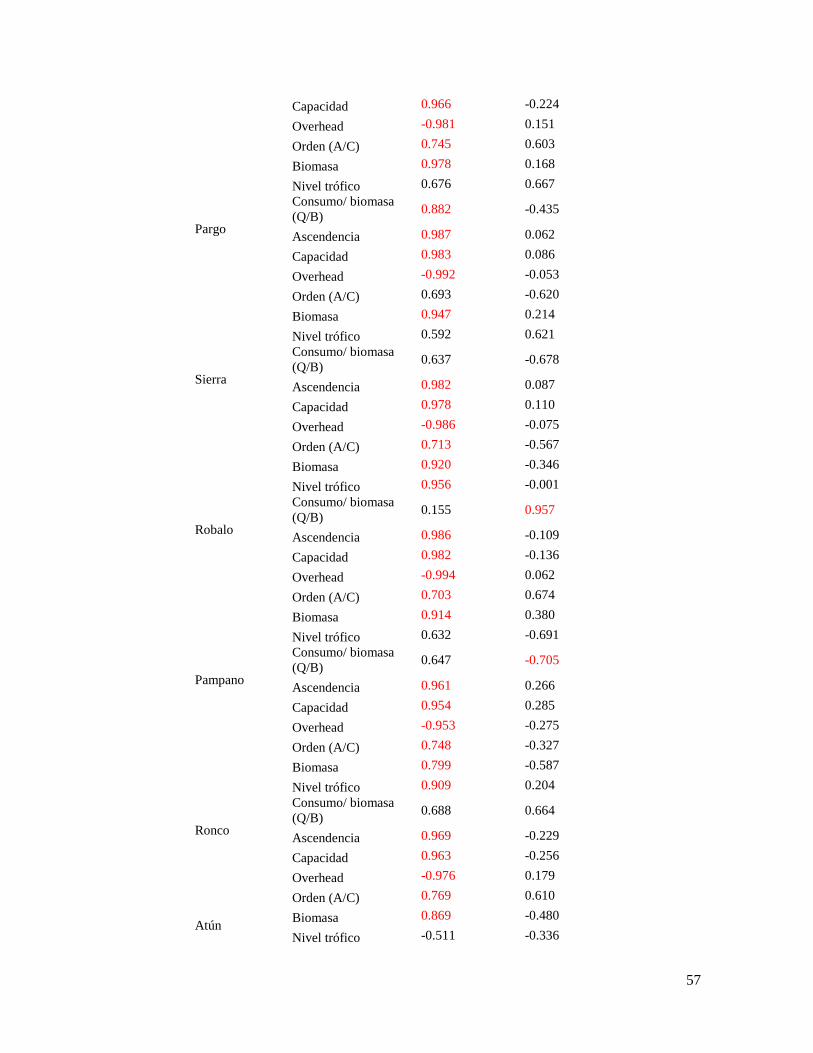
57
Capacidad 0.966 -0.224
Overhead -0.981 0.151
Orden (A/C) 0.745 0.603
Pargo
Biomasa 0.978 0.168
Nivel trófico 0.676 0.667
Consumo/ biomasa
(Q/B) 0.882 -0.435
Ascendencia 0.987 0.062
Capacidad 0.983 0.086
Overhead -0.992 -0.053
Orden (A/C) 0.693 -0.620
Sierra
Biomasa 0.947 0.214
Nivel trófico 0.592 0.621
Consumo/ biomasa
(Q/B) 0.637 -0.678
Ascendencia 0.982 0.087
Capacidad 0.978 0.110
Overhead -0.986 -0.075
Orden (A/C) 0.713 -0.567
Robalo
Biomasa 0.920 -0.346
Nivel trófico 0.956 -0.001
Consumo/ biomasa
(Q/B) 0.155 0.957
Ascendencia 0.986 -0.109
Capacidad 0.982 -0.136
Overhead -0.994 0.062
Orden (A/C) 0.703 0.674
Pampano
Biomasa 0.914 0.380
Nivel trófico 0.632 -0.691
Consumo/ biomasa
(Q/B) 0.647 -0.705
Ascendencia 0.961 0.266
Capacidad 0.954 0.285
Overhead -0.953 -0.275
Orden (A/C) 0.748 -0.327
Ronco
Biomasa 0.799 -0.587
Nivel trófico 0.909 0.204
Consumo/ biomasa
(Q/B) 0.688 0.664
Ascendencia 0.969 -0.229
Capacidad 0.963 -0.256
Overhead -0.976 0.179
Orden (A/C) 0.769 0.610
Atún Biomasa 0.869 -0.480
Nivel trófico -0.511 -0.336
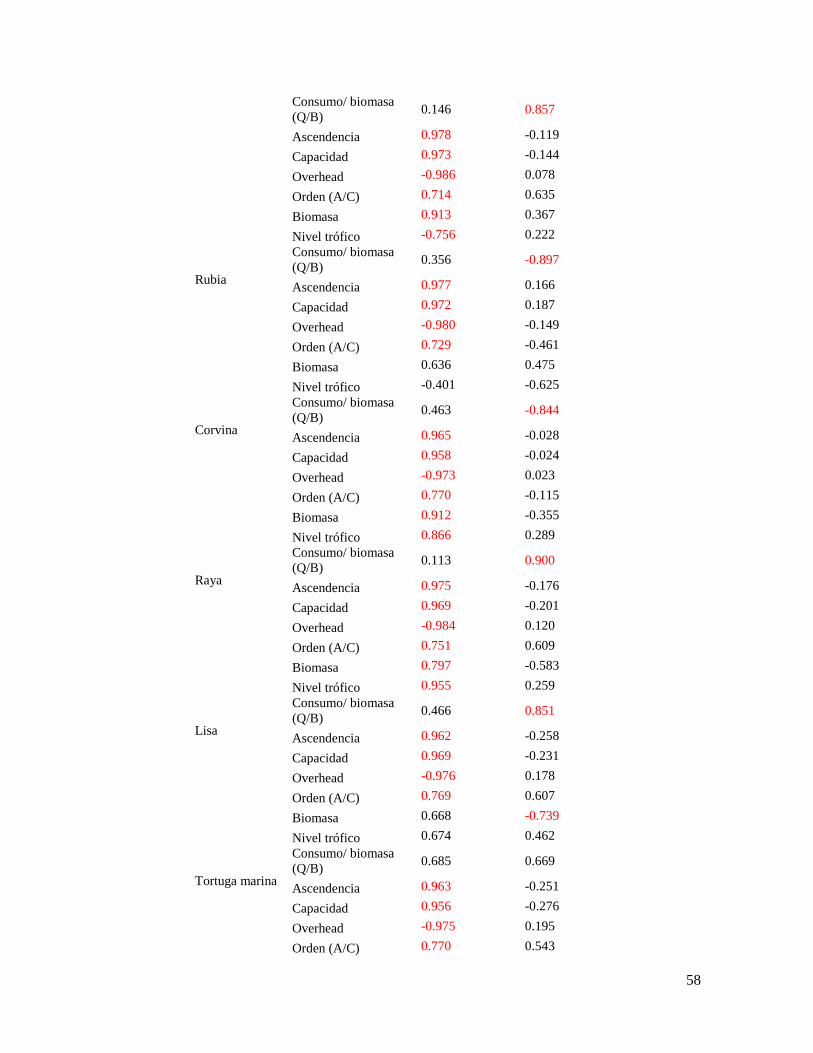
58
Consumo/ biomasa
(Q/B) 0.146 0.857
Ascendencia 0.978 -0.119
Capacidad 0.973 -0.144
Overhead -0.986 0.078
Orden (A/C) 0.714 0.635
Rubia
Biomasa 0.913 0.367
Nivel trófico -0.756 0.222
Consumo/ biomasa
(Q/B) 0.356 -0.897
Ascendencia 0.977 0.166
Capacidad 0.972 0.187
Overhead -0.980 -0.149
Orden (A/C) 0.729 -0.461
Corvina
Biomasa 0.636 0.475
Nivel trófico -0.401 -0.625
Consumo/ biomasa
(Q/B) 0.463 -0.844
Ascendencia 0.965 -0.028
Capacidad 0.958 -0.024
Overhead -0.973 0.023
Orden (A/C) 0.770 -0.115
Raya
Biomasa 0.912 -0.355
Nivel trófico 0.866 0.289
Consumo/ biomasa
(Q/B) 0.113 0.900
Ascendencia 0.975 -0.176
Capacidad 0.969 -0.201
Overhead -0.984 0.120
Orden (A/C) 0.751 0.609
Lisa
Biomasa 0.797 -0.583
Nivel trófico 0.955 0.259
Consumo/ biomasa
(Q/B) 0.466 0.851
Ascendencia 0.962 -0.258
Capacidad 0.969 -0.231
Overhead -0.976 0.178
Orden (A/C) 0.769 0.607
Tortuga marina
Biomasa 0.668 -0.739
Nivel trófico 0.674 0.462
Consumo/ biomasa
(Q/B) 0.685 0.669
Ascendencia 0.963 -0.251
Capacidad 0.956 -0.276
Overhead -0.975 0.195
Orden (A/C) 0.770 0.543
59
Trucha de mar
Biomasa 0.909 -0.379
Nivel trófico 0.236 -0.404
Consumo/ biomasa
(Q/B) 0.236 0.917
Ascendencia 0.990 -0.086
Capacidad 0.987 -0.113
Overhead -0.992 0.050
Orden (A/C) 0.683 0.689
Jurel
Biomasa 0.908 -0.406
Nivel trófico -0.711 0.110
Consumo/ biomasa
(Q/B) 0.558 0.796
Ascendencia 0.974 -0.160
Capacidad 0.969 -0.186
Overhead -0.973 0.123
Orden (A/C) 0.728 0.632
Anchoveta
Biomasa 0.933 0.304
Nivel trófico 0.470 -0.794
Consumo/ biomasa
(Q/B) 0.764 -0.516
Ascendencia 0.964 0.250
Capacidad 0.957 0.270
Overhead -0.957 -0.260
Orden (A/C) 0.753 -0.350
Sardina
Biomasa 0.886 0.229
Nivel trófico -0.367 0.879
Consumo/ biomasa
(Q/B) 0.774 -0.135
Ascendencia 0.976 0.204
Capacidad 0.970 0.224
Overhead -0.979 -0.153
Orden (A/C) 0.741 -0.462
Pulpo
Biomasa 0.971 0.091
Nivel trófico -0.874 -0.116
Consumo/ biomasa
(Q/B) 0.076 -0.959
Ascendencia 0.988 0.034
Capacidad 0.984 0.048
Overhead -0.997 -0.039
Orden (A/C) 0.677 -0.350
Mojarra
Biomasa 0.969 Nivel trófico 0.947 Consumo/ biomasa
(Q/B) 0.596
Ascendencia 0.969 Capacidad 0.962

60
Overhead -0.97 Orden (A/C) 0.761
Jaiba
Biomasa 0.973 Nivel trófico 0.929 Consumo/ biomasa
(Q/B) 0.657
Ascendencia 0.97 Capacidad 0.963 Overhead -0.976 Orden (A/C) 0.763
Langosta
Biomasa 0.791 Nivel trófico 0.81 Consumo/ biomasa
(Q/B) 0.952
Ascendencia 0.956 Capacidad 0.948 Overhead -0.947 Orden (A/C) 0.771
Camarón
Biomasa 0.992 Nivel trófico 0.946 Consumo/ biomasa
(Q/B) 0.969
Ascendencia 0.979 Capacidad 0.974 Overhead -0.98 Orden (A/C) 0.721
Calamar
Biomasa 0.975 Nivel trófico -0.968 Consumo/ biomasa
(Q/B) 0.388
Ascendencia 0.983 Capacidad 0.978 Overhead -0.991 Orden (A/C) 0.723
Zooplancton
Biomasa 0.991 Nivel trófico 0.98 Consumo/ biomasa
(Q/B) 0.973
Ascendencia 0.985 Capacidad 0.98 Overhead -0.986 Orden (A/C) 0.715
Microcrustáceos
Biomasa 0.964 Nivel trófico -0.698 Consumo/ biomasa
(Q/B) 0.957

61
Ascendencia 0.985 Capacidad 0.981 Overhead -0.991 Orden (A/C) 0.704
Poliquetos
Biomasa 0.979 Nivel trófico -0.821 Consumo/ biomasa
(Q/B) 0.947
Ascendencia 0.975 Capacidad 0.969 Overhead -0.985 Orden (A/C) 0.749
Equinodermos
Biomasa 0.966 Nivel trófico -0.965 Consumo/ biomasa
(Q/B) 0.948
Ascendencia 0.974 Capacidad 0.968 Overhead -0.979 Orden (A/C) 0.753
Macroalgas
Biomasa 0.99 Ascendencia 0.987 Capacidad 0.982 Overhead -0.987 A/C 0.715
Pastos marinos
Biomasa 0.99 Ascendencia 0.988 Capacidad 0.983 Overhead -0.988 A/C 0.711
Fitoplancton
Biomasa 0.993 Ascendencia 0.986 Capacidad 0.981 Overhead -0.987 A/C 0.719
Detritus
Biomasa 0.995 Ascendencia 0.986 Capacidad 0.981 Overhead -0.988 A/C 0.719

62
9. Discusión
9. 1 Construcción del modelo: estimación de los datos de entrada
Cuando se construyen modelos tróficos cuya característica es el balance energético de los
flujos de biomasa, para una misma área y en tiempos distintos, estos modelos pueden permitir
evaluar importantes cambios en la estructura, organización y función del ecosistema respecto
al tiempo (Arreguín-Sánchez et al., 1993).
Uno de los parámetros más importantes en los modelos tróficos de ecosistema tipo Ecopath
es la biomasa (Christensen & Walkers, 2004). Se considera que este parámetro sea uno de
los últimos que el modelo Ecopath calcule, debido a que la biomasa que estima no es la
biomasa que necesariamente tendría cierto grupo funcional en el ecosistema, sino la biomasa
necesaria para lograr un balance de masas dentro del modelo. En este sentido, el modelo de
la plataforma continental de Yucatán se construyó con datos de biomasa calculados a partir
de datos de avisos de arribo de las pesquerías que se desarrollan en la zona para los peces, y
para el resto de los grupos se dejó que el modelo estimara su biomasa.
Los grupos delfín, ave marina, tortuga marina, jaiba, camarón, calamar, zooplancton,
microcrustáceos, poliquetos, equinodermos, macroalgas, pastos marinos y fitoplancton en los
cuales la biomasa fue estimada por Ecopath son los que presentan una mayor certidumbre
con respecto a la biomasa debido a que fue calculada con información de la plataforma
continental de Yucatán. Los grupos de productores primarios fueron los que aportaron mayor
biomasa al ecosistema de la plataforma continental de Yucatán (141.75 t·km-2), esta biomasa
es mayor a la reportada por Arreguín-Sánchez y colaboradores (1993) para el mismo
ecosistema la cual fue 0.000014 t·km-2 y a pesar de que en 2000 se ajustó el modelo por
Arreguín-Sánchez y colaboradores, la cantidad de biomasa reportada sigue siendo menor
(0.000061) a lo que se reporta en el presente trabajo, sin embargo, en los anteriores trabajos
se consideran solo dos grupos de productores primarios: fitoplancton y productores
bentónicos, del cual el último grupo considera a una sola especie como productores
bentónicos (Rhizophora mangle), por tal motivo se infiere que la biomasa representada para
estos dos modelos sea menor que la que se reportó en el presente trabajo. Y se considera que
este valor se tome en cuenta solo como referencia preliminar, puesto que al ser estimado por
el propio programa tiene algunas implicaciones que hacen suponer una sub-estimación de

63
estos valores. Sin embrago, no fue el mismo caso para los datos de biomasa de algunos grupos
de invertebrados, debido a que la importancia de estas comunidades en el flujo de materia y
energía entre el sedimento y los consumidores secundarios, han influido para ser objeto de
una evaluación detallada en cuanto a diversidad y abundancia.
Por otra parte el aporte del mero rojo en biomasa al ecosistema es relativamente bajo (0.05
t·km-2). A pesar de ello, se ha reportado que las especies claves son aquellas que presentan
una biomasa relativamente baja, ya que tienen un papel importante sobre la composición y
el funcionamiento de las comunidades del ecosistema (Power et al., 1996), y como se pueden
constatar con la matriz de dieta el mero rojo influye potencialmente en la abundancia de los
grupos tróficos (bonito, peto, pargo, ronco, jaiba, pulpo, camarón, langosta) y por lo tanto en
la dinámica del ecosistema. Este resultado hace constar con lo reportado por el trabajo de
Arreguín-Sánchez et al., (1993 y 2000).
A pesar de que el Golfo de México y particularmente la plataforma de Yucatán son
zonas de gran interés de estudio, debido a la alta productividad biológica, existe poca
información para algunos grupos funcionales en la zona de estudio, como falta de datos
biológicos, ecológicos y pesqueros requeridos para el cálculo de P/B y Q/B para la mayoría
de los de los invertebrados, quienes juegan un papel ecológico importante debido a su gran
diversidad y alta biomasa de algunos grupos, así como para los vertebrados superiores
(mamíferos, aves y tortugas marinas) existen pocos estudios en el área de estudio sobre la
alimentación, abundancia, crecimiento y reproducción. A pesar de esto, fue posible obtener
los datos de entrada suficientes para construir el modelo que representan la estructura del
ecosistema y de sus grupos funcionales. Aunque las estimaciones de P/B son pocas, la
resultaron ser consistentes respecto a los parámetros estimados para modelos de la zona
(Arreguín-Sánchez et al., 1993 y 2000) y cercanos (como la sonda de Campeche, Zetina-
Rejón, 2004, y la Bahía Ascención, Quintana Roo, Vidal, 2003).
9.2 Balanceo del modelo
Uno de los principales supuestos del enfoque Ecopath es el balance de masas (Polovina 1984,
Christensen & Pauly 1992), lo que significa que las entradas (materia o energía) y salidas
deben de ser iguales, dándose un balance entre los componentes del ecosistema. El
cumplimiento del supuesto implica realizar ajustes graduales de acuerdo a criterios

64
biológicos de cada grupo, a gran parte de la información original, principalmente en la matriz
de dietas ya que son los que mayor incertidumbre contienen (Christensen & Walters 2004,
Christensen et al. 2005, Galván-Piña 2005). Por lo cual se estableció que la EEi fuera menor
de 1, esto se obtuvo, en los grupos ecológicos donde fue necesario, modificar ligeramente (±
5 %) los consumos en la dieta inicial de predador para cada presa, evitando el añadir y
eliminar presas. En relación a los valores de EEi (Tabla 4), algunos peces registraron valores
altos a 0.90 o cercanos, como es el caso de: pargos, sierra, corvina, anchoveta, sardina. Estos
resultados indican que la producción de cada grupo es altamente utilizada en términos de
depredación natural dentro del ecosistema o de explotación por parte de las flotas
comerciales. Por el contrario, los valores más bajos de EEi fueron presentados por los
predadores apicales, es decir, por el tiburón, mero rojo, delfín, cazón, ave marina, bonito y
atún, este resultado podría indicar que estos grupos son escasamente consumidos de forma
natural (Bayle-Sempere et al., 2013). Mientras que algunos invertebrados como: la langosta,
microcrustáceos, poliquetos, fueron los que obtuvieron los valores bastantes elevados de EEi
(> 0.95), sugiriendo que son altamente consumidos en el ecosistema tal y como lo sugiere el
índice de grado para estos grupos. Para el caso de los productores primarios el pasto marino
seguido de las macroalgas fueron los que obtuvieron valores superiores a 0.90, esto podía
sugerir que el ecosistema de la plataforma continental de Yucatán provee una alta
productividad la cual en parte se encuentra modulada por los incrementos de temperatura en
la zona de estudio (Arreguín-Sánchez et al., 2015).
El modelo construido fue consistente con los criterios propuestos por Christensen et
al., (2005), es decir los indicadores utilizados para verificar el equilibrio termodinámico del
modelo tuvieron la tendencia general esperada a lo largo de la trama trófica. El cociente P/Q
fue mayor para los niveles tróficos inferiores y disminuye conforme se incrementa el nivel
trófico, el coeficiente R/A fue mayor en grupos con mayor actividad y muy cercanas a
predadores tope, ya que la respiración no puede exceder a la proporción de alimento que es
asimilada. Las tendencias de estos indicadores encontradas en este estudio son similares a las
reportadas por otros autores en distintos ecosistema (Salcido-Guevara & Arreguín-Sánchez,
2007; Zetina-Rejón et al., 2016; Heymans et al., 2016). Lo anterior sugiere que nuestro
modelo trófico tipo Ecopath es consistente ecológicamente

65
9.3 Atributos de los grupos funcionales
La información sobre los aspectos tróficos de las especies permite identificar la posición que
ocupan los grupos funcionales dentro de la trama trófica, y por tanto evalúa su nivel trófico
dentro de la comunidad, este dato es importante cuando se pretende cuantificar el impacto de
la actividad pesquera en ecosistemas explotados (Pauly et al., 1998a; Stergiou & Karpouzi,
2002). De este modo, la caracterización de la estructura trófica en un ecosistema marino
puede servir de base para el mantenimiento del balance del nivel trófico de la comunidad y
en consecuencia, frenar el descenso progresivo del mismo, inducido por la pesca y enmarcado
dentro del enfoque de la gestión pesquera basado en el ecosistema (Pauly et al., 1998a;
Branch et al., 2010).
En análisis general de la dieta, reveló que los predadores basan su alimentación en
grupos de invertebrados, siendo el camarón, los microcrustáceos y pulpo las presas más
importantes (ver Tabla 5), y contrastando con la matriz de dieta de trabajos en la zona
(Arreguín-Sánchez et al., 1993; Arreguín-Sánchez et al., 2000) y cerca de la zona (Zetina-
Rejón 2004; Cabrera 2005) coincide la dieta de los predadores tope, siendo los
microcrustáceos y camarón los más abundantes como ítem presa. Y en caso particular la dieta
del mero rojo, dio la información sobre la posición que ocupa dentro de la red trófica, siendo
este un predador tope, mostrando una mayor diversidad trófica y por tanto, un
comportamiento más generalista dentro del ecosistema.
9.4 Atributos del ecosistema
De acuerdo a Odum (1969) existen atributos que describen la madurez de los ecosistemas
(entiéndase como la madurez del ecosistema), y uno de estos atributos es la energía de la
comunidad, el cual considera los flujos que se dan en el ecosistema, entre ellos destacan el
cociente de producción primaria total/respiración total (PPT/RT), Producción Neta del
Sistema (PNS), Producción Primaria Neta Total (PPNT) y la relación Producción primaria
total/biomasa total (PPT/BT). Conforme los sistemas maduran la eficiencia de transferencia
de energía incrementa, esto provoca que lo que se consume tienda a ser similar a los que se
produce, por lo cual se considera que las pérdidas de energía vía respiración aproximan a las
producciones, esto se manifiesta en que la relación PPT/RT tienda a uno. En el caso del

66
ecosistema de la plataforma de Yucatán resulto ser un ecosistema inmaduro y en desarrollo,
lo cual indica que la producción es mayor a la respiración (PPT/RT > 1), y esto se constata
con que el valor de la Producción Neta del Sistema (PNS) es menor a la Producción Primaria
Neta Total (PPNT), lo cual puede deberse a que la biomasa de los productores primarios es
relativamente grande en el ecosistema, y por tanto la respiración total no influye mucho en
la PPNT, por lo mismo el cociente PPT/RT fue mayor a uno.
Una de las posibles razones por la cual el ecosistema se encuentre en estado inmaduro se
debe a que esta relación tiende a ser subestimada, debido a que no se incluyen dentro del
modelo a grupos cuya función es la descomposición de materia orgánica, como las bacterias
(Christensen & Pauly, 1993). No obstante, Arreguín-Sánchez et al., (1993) reportaron que el
ecosistema de la plataforma continental del norte de Yucatán se encuentra en un estado
maduro (PPT/RT = 0.75), quizás el resultado de ambos análisis varié por el periodo en que
se realizó cada modelo, además cabe resaltar, que en el trabajo de Arreguín-Sánchez et al.,
(1993) los datos de entrada al modelo de los grupos funcionales se extrajeron de literatura
reportada en diferentes periodos. En este sentido, el modelo construido en este estudio debido
a la procedencia de la información de entrada tiene una mayor incertidumbre para definir el
estado de madurez de la plataforma continental de Yucatán.
Por otra parte, la teoría de desarrollo de Odum (1969), propone que mientras el cociente
PPT/BT se aproxime a uno, el ecosistema puede ser considerado en un estado de mayor
madurez. El valor obtenido para el modelo de la plataforma continental de Yucatán fue 12.51,
y este valor es mayor de lo reportado por Arreguín-Sánchez et al., (1993) (PPT/BT = 6.97),
lo cual confirma que el ecosistema analizado en este estudio pudiera encontrarse en un estado
inmaduro. Este estado es muy frecuente en ecosistemas explotados y con recursos sobre-
explotados (donde el esfuerzo es dirigido mayormente a la captura de especies objetivos), ya
que los impactos que genera esta actividad, produce que el esfuerzo de captura se vea
mermado y, por consiguiente se re-oriente hacia otros recursos no objetivo, lo cual genera
cambios en la abundancia y diversidad de las especies, y por consiguiente se tendría efectos
indirectos sobre la red trófica por pérdida de predadores de nivel trófico alto e intermedio, de
tal modo que el ecosistema se mantienen en un constante estado sucesión que puede ser
interpretado como inmadurez (Gislason & Sinclair, 2000; Trenkel & Rochet, 2003; Odum,
1969).

67
Particularmente centrándonos en el caso de la pesquería del mero rojo, las capturas en
Yucatán han registrado variaciones a lo largo del tiempo (DOF 2104), y de igual manera, se
ha observado en los últimos 15 años la presencia de especies asociadas a la pesca del mero
ha aumentado. Y esto se hace notar con el caso de la pesca del pulpo, ya que los cambios
observados en las capturas desembarcadas de mero, han sido explicados como un efecto la
adición del pulpo como especie objetivo por parte de las mismas embarcaciones, cuando
antes éstas solo asignaban su esfuerzo a pesca de escama. Puede observarse que las capturas
de mero rojo muestran una tendencia a disminuir cuando aumentan las de pulpo y viceversa.
Por otro lado el deterioro de las poblaciones de mero rojo también se ha visto reflejado con
una tendencia a la baja en las capturas, especialmente si se les compara con los niveles
alcanzados en la década de 1990, pasando así del primer lugar al segundo, después del pulpo
(Anexo 4 y 5).
Con respecto al índice de omnivoría fue para la mayoría de los grupos relativamente
bajo considerándolos como grupos con hábitos especialistas, sin embargo, se puede observar
que los grupo tienen un cierto hábito generalista, y se puede inferir que el cálculo del índice
le da mayor peso al ítem de presas que se consume más en la dieta de los grupos, por ende
considera en general a los grupos como especialistas, a pesar de esto, se puede constatar con
la matriz de dietas (Tabla 5) que en general los grupos son en menor proporción generalistas,
tal es el caso de grupos como el peto, pargo, tiburón, mero rojo, cazón, ave marina, trucha de
mar, sardina y calamar que presenta un elevado número de ítems de presas en su dieta.
Por otro lado el índice de conectividad indica que sólo existe un 20 % de uniones
tróficas posibles entre los grupos funcionales. Este índice depende del tamaño del sistema,
ya que es utilizado para medir la complejidad de la trama trófica, por lo cual este índice es
susceptible a la información que se utilice en la asignación de las dietas, de tal manera este
índice no puede ser comparable entre modelos (Galván-Piña 2005; Lercari et al., 2010).
Los valores obtenidos por el análisis de impactos tróficos, sugiere que el mero rojo y
otros grupos funcionales (delfín, sierra, huachinango, pastos marinos y fitoplancton) fueron
importantes para el ecosistema de la plataforma de Yucatán, debido a que estos grupos

68
ocasionaron efectos positivos sobre la mayoría de los grupos funcionales incluidos en el
modelo.
9.5 Indicadores de crecimiento y desarrollo
De acuerdo a Christensen (1992) la madurez también está relacionada con la estabilidad y a
su vez con el overhead o resiliencia del ecosistema (Christensen & Pauly 1993). El overhead
para el ecosistema de la plataforma continental de Yucatán fue 69.5 % y la ascendencia 30.5
%, lo cual podría indicar que cerca del 30 % de la capacidad total de desarrollo del ecosistema
para la organización de flujos está siendo utilizada. Para otros ecosistemas cercanos al área
de estudio ha sido relativamente menor el overhead, tal es el caso de la Sonda de Campeche
(Zetina-Rejón & Arreguín-Sánchez, 2003) con 55 %, y para la plataforma continental de
Tabasco fue de 47. 8 % (Cabrera-Neri, 2005). Esto refleja que no todos los ecosistemas se
ubican en el mismo estado de desarrollo debido a la dinámica propia de los flujos de energía
las relaciones tróficas entre los componentes del ecosistema.
A pesar de que el ecosistema de la plataforma continental de Yucatán muestre un nivel de
crecimiento característico de sistemas inmaduros o en desarrollo, la red trófica funciona de
manera eficiente, y su capacidad de respuesta ante perturbaciones externas es lo
suficientemente alta.
9.6 Indicadores estructurales
Con respecto a los índices de centralidad, los valores de grado del nodo reflejaron que el
camarón, poliquetos y microcrustáceos, son los nodos con mayor número de conexiones
tróficas en el ecosistema de la plataforma continental de Yucatán, siendo estos grupos los
que se encuentran más comunicados que el resto en términos de flujo de energía. Por otra
parte, al hacer un análisis de frecuencias observamos que se presentan valores bajos de
interacción siendo entre 5 y 10 el grado de nodo más frecuente, es decir, solamente pocos
componentes del sistema presenta un alto número de conexiones, mientras que la mayoría se
encuentra interactuando en la red mediante un reducido número de conexiones tróficas
(Anexo 1). Por lo tanto se observa que el ecosistema de la plataforma continental de Yucatán
presenta la mayor parte de sus componentes con un número de conexiones tróficas

69
relativamente bajo, con respecto al total de conexiones posibles, y de acuerdo a autores como
Estrada (2007); Benedeck et al. (2008) el bajo número de relaciones indirectas entre todos
los componentes del sistema sugiere que un efecto sobre alguno de ellos traería como
consecuencia una restructuración de la red trófica sin llegar al colapso de la misma.
Por su parte el índice de cercanía mostró que los grupos róbalo, trucha de mar y la lisa se
encontraron interactuando en la red mediante rutas más cortas entre los grupos.
Finalmente el índice de intermediación dio como resultado que los grupos el delfín, tiburón,
bonito, pargo, tortuga marina, equinodermos y fitoplancton son grupos puentes en el
ecosistema, lo cual indica que estos grupos son los que presentan la mayor capacidad de tener
efectos tróficos sobre el resto de la red, ya que funcionan como intermediarios de información
en la red trófica. Es decir si estos grupos son removidos pudiesen tener efectos sobre la
mayoría del resto de los nodos considerados, en un periodo corto. El histograma de
frecuencias de intermediación (Anexo 3) mostró fue similar al observado en las frecuencias
del índice de grado, encontrando una distribución recargada hacia los valores bajos (entre
cero y 100), por lo que el análisis nos indica que la mayoría de los organismos comparte
pocas relaciones con los demás.
9.7 Simulación temporal
Existe evidencia que las variaciones climáticas afectan diferentes niveles de organización,
incluyendo poblaciones, comunidades y ecosistemas. Creciente evidencia científica muestra
que la fluctuaciones climáticas afectan la abundancia y biogeografía de los organismo
(Stenseth et al., 2002; Cheung et al., 2012). Contraponiendo los resultados que se obtuvieron
de los patrones de biomasa versus A/C con el AMO. Se puede observar que durante la etapa
de transición de fase fría a cálida (a mitad de la década de los 1990), los patrones (ver figura.
9) se encuentran en un periodo de compensación, es decir, las biomasas y orden registran
valores bajos conforme el tiempo avanza, después se estabilizan, y posteriormente la biomasa
tiende a aumentar, y el orden se mantiene estable. Teóricamente se conoce que un sistema
puede mantener orden si tiene energía en reserva suficiente para hacer frente a perturbaciones
(Ulanowicz, 1986). Por consiguiente, si tomáramos el AMO como un factor de perturbación
en el ecosistema, podríamos deducir este factor climático influye en la abundancia de la
biomasa de cada grupo funcional y por consiguiente en el ecosistema, ya que con cada

70
cambio de temperatura a través del tiempo, el ecosistema tiende a organizarse, es decir el
ecosistema tiene la capacidad de responder eficientemente ante dicha perturbación. Además,
se puede sugerir que durante la fase fría del AMO (periodo de 1970 – 1900) tuvo un efecto
positivo en la abundancia de biomasa.
Cabe mencionar, que cada patrón presentan diferencias a lo largo del tiempo, por
ejemplo se puede observar que en el primer patrón se presenta una sola fluctuación en la
tendencia de biomasa a principios del periodo considerado (Fig. 9b), declina y aumenta a
finales de los 90’s, esto podría deberse en gran parte a que las especies (a excepción de los
grupos como delfín, ave marina y tortuga marina) que conforman este patrón están
respondiendo no solo a las variaciones climáticas sino también al factor pesca. Ante tal
precepto, Ulanowicz et al., (2009) considera que la organización es un mecanismo de
respuesta propia del ecosistema en cuestión que responde a perturbaciones, ya sea de tipo
natural o antrópicas. Por ende, se puede inferir que esos grupos tienden a estar más
presionados en el ecosistema, y su capacidad de respuesta (resiliencia) ante estos eventos es
más prolongada con respecto a las demás especies.
En los demás patrones (Fig, 9 d, f y h), particularmente en el tercero (Fig. 9 f) se
puede observar que existen más cambios en la biomasa a través del tiempo y esto
posiblemente se deba a el cambio de flujos de energía (biomasa) desde el ápice de la cadena
trófica, y por consiguiente los grupos funcionales inferiores tienden a compensar dichos
cambios, puesto que una cantidad de energía cada vez mayor se dedica al mantenimiento del
ecosistema. Este rasgo podría estar ligado a un suceso de succión. Lo cual se constata con lo
dicho en el apartado de atributos del ecosistema, en el presente estudio, se encontró al
ecosistema en estado inmaduro, indicativo de un ecosistema en proceso de desarrollo de
acuerdo a las características propuestas por Odum (1963).
9.8 Interpretación del papel del mero
El estudio ecológico de las especies (con énfasis en conocer el papel que desarrollan en el
ecosistema) es útil para conocer la funcionalidad y estabilidad de un ecosistema, ya que
ninguna especie se encuentra aislada, y las diferentes relaciones bióticas y abióticas que
existen entre ellas suelen ser diferentes. En el caso particular del mero rojo, se pudo

71
caracterizar el papel que ocupa dentro del ecosistema de la plataforma continental de
Yucatán. En primer plano se determinó la estructura trófica del grupo, está mostró un
comportamiento generalista dentro del ecosistema, debido a la amplia gama de ítems presas,
dentro de los cuales destacan grupos como: jaiba, pulpo, camarón, zooplancton, ronco y
pámpano. Las interacciones directas del mero sobre sus presas revelaron que la especie
influye de manera positiva y negativa. El grupo de pulpo fue el único que mostro un impacto
positivo, mientras que el impacto negativo abarco a la mayoría de su ítems presa (peto,
zooplancton, jaiba, camarón y microcrustáceos), esto pudiera deberse principalmente a la
preferencia del mero por estos organismos, sin embargo, también se puede atribuir que este
impacto negativo correspondan a la presión que tienen estos ítems presas por otros grupos,
por lo cual sugeriría que el mero rojo se encuentra compitiendo por estos recursos (ver
sección 8.4.3 Impactos tróficos mixtos). En este trabajo, se hace énfasis en que las
interacciones ecológicas del mero (depredación y competencia) bridan estabilidad ecológica
al ecosistema de la plataforma continental de Yucatán. Sin embargo, existen trabajos en los
cuales se ha discutido por saber cuál es papel de las presa, de los depredadores y de los
competidores en la regulación de las comunidades. En el trabajo de Watt (1964) infiere que
la estabilidad está dada por la cantidad de depredadores y competidores que pueden mantener
en sus niveles de abundancia a una presa determinada. Por otro lado MacArthur (1955)
propone que la estabilidad está en función de la cantidad de fuentes alternativas de alimento
para una especie. Pimm (1991) considera que cuando una presa tiene varios depredadores es
más fácil que estos mantengan a las poblaciones de la presa en un nivel bajo, ya que en caso
de que alguno de los depredadores disminuya, los otros podrán ejercer el control. No
obstante, al analizar los indicadores estructurales se pudo observar que el mero rojo no cuenta
con muchas conexiones dentro del ecosistema, ni tampoco mostró ser un grupo con la
capacidad de influir de manera importante dentro del ecosistema y de igual manera no se
caracterizó por ser un grupo puente con otros grupos dentro del ecosistema. Sin embargo, los
análisis destacan la importancia de las presas del mero, ya que el camarón y microcrustáceos
fueron los grupos con mayor número de conexiones dentro del ecosistema. Este análisis
pudiera plantear que las presas son las que regulan el funcionamiento del ecosistema. De
igual manera los análisis revelan que los competidores potenciales del mero, como el delfín
y tiburón son especie puente dentro del ecosistema, en el supuesto de que elimináramos estos

72
competidores posiblemente tendría en impacto positivo en el incremento de biomasa del
mero, sin embargo, existen un cuestionamiento sobre si el eliminar un competidor se
incrementa la biomasa del otro, ya que se ha comprobado en trabajos (Connel, 1983;
Schoener, 1983) que la eliminación de un competidor no siempre generan un incremento en
la biomasa del otro. Como puede verse, las interacciones ecológicas de los grupos también
brindan estabilidad al ecosistema, sin embargo, se debe de caracterizas cuales tienen mayor
importancia dentro de un grupo y del ecosistema.
En segundo plano, se identificó cuáles son los atributos del mero rojo que pueden
tener mayor influencia en el ecosistema, y por ende en el funcionamiento del grupo. El papel
del mero rojo destaca por ser una especie importante en el ecosistema, no solo como
regulador de la abundancia de sus presas, sino también en el desarrollo del ecosistema. De
manera particular destaca el papel diferencial del mero y tiburones, los cuales juegan un papel
determinante en términos del mantenimiento del orden y organización del ecosistema. Aun
cuando el análisis realizado no exhaustivo, el comportamiento del mero en relación con la
respuesta del ecosistema en los experimento de simulación y del análisis de componentes
principales, refieren que el papel de predador tope es relevante respecto al orden del
ecosistema.

73
10. Conclusión
Con el modelo Ecopath se puedo construir un modelo trófico de flujos de biomasa útil para
conocer la estructura y funcionamiento del ecosistema de la plataforma de Yucatán.
En análisis general de la dieta, reveló que los predadores basan su alimentación en grupos de
invertebrados, siendo el camarón, los microcrustáceos y pulpo las presas más importantes
A pesar de que el ecosistema muestre un nivel de crecimiento característico de sistemas
inmaduros, la red trófica funciona de manera eficiente, y su capacidad de respuesta ante
perturbaciones externas es lo suficientemente alta.
En lo que respecto mero se rojo se determinó como una especie clave, ya que la abundancia
de este grupo tiende a estar altamente relacionada con la estructura y funcionamiento del
ecosistema de la plataforma de Yucatán.
Los indicadores de conectividad no reflejan ningún comportamiento diferencial del mero rojo
con respecto a otros grupos del ecosistema, sin embargo, los indicadores de organización
sugieren un papel diferencial. No obstante, el papel del mero rojo destaca como regulador de
la abundancia de sus ítems presas. De manera particular destaca el papel diferencial del mero,
el cual juega un papel determinante en términos del mantenimiento del orden y organización
del ecosistema.

74
11. Refencias
Abarca-Arenas y Valero-Pacheco. 1993. Toward a Trophic Model of Tamiahua, a Costal
Lagoon in México. Pp: 181- 185. En: Christensen, V. & Pauly, D (Eds.). Trophic
models of aquatics ecosystems. ICLARM, Manila, Philippines. 390.
Aguirre-León, A. & A. Yáñez-Arancibia.1986. Las mojarras de la Laguna de Términos:
Taxonomía, biología, ecología y dinámica trófica (Pisces: Gerreidae). Anales del
Instituto de Ciencias del Mar y Limnología 13 (1): 369 - 443.
Allen K. R. 1971. Relation between production and biomass. Journal of the Fisheries
Research Board of Canada. 28: 1573 - 1581.
Andrade, H., J. Santos & R. Taylor. 2013. Life-history traits of the common snook
Centropomus undecimalis in a Caribbean estuary and large-scale biogeographic
patterns relevant to management. J. Fish Biol. 82: 1951- 1974.
Arias-González, J.E., E. Nuñez-Lara, C. González-Salas & R. Galzin. 2004. Trophic models
for investigation of fishing effect on coral reef ecosystems. Ecological Modelling.
172 (2004):197- 212.
Arreguín-Sánchez, F. 2000. Octopus–red grouper interaction in the exploited ecosystem of
the northern continental shelf of Yucatan, Mexico. Ecol. Model., 129 (2): 119 - 129.
Arreguín-Sánchez, F. 2014. Adaptabilidad y sostenibilidad: un enfoque holístico hacia el
manejo de ecosistemas acuáticos explotados. Ciencia Pesquera. 22(1): 89 - 101
Arreguín-Sánchez, F. & E. Chávez. 1995. An investigation of the trophic role of three pelagic
fishes in a tropical coastal ecosystem of the western Gulf of Mexico, using the
ECOPAT II model. Scien. Mar. 4: 307 – 315.
Arreguín-Sánchez, F., J. C. Seijo & E. Valero-Pacheco. 1993a. An application of Ecopath II
to the North Continental shelf ecosystem of Yucatan, Mexico, Pp: 269 - 278. En V.
Christensen & D. Pauly (eds). Trophic models of aquatic ecosystems. ICLARM.
Conf. Proc. 26, Ciudad de Makati, Filipina.

75
Arreguín-Sánchez, F., J. L. Munro., M. C. Balgos & D. Pauly. 1993b. Biology, fisheries and
culture of tropical groupers and snappers, p. 19-27. ICLARM conf. Proc. 48, Ciudad
de Makati, Filipinas
Arreguín-Sánchez, F., M. Zetina-Rejón., S. Manickchand-Heileman., M. Ramírez-Rodríguez
& L. Vidal. 2004. Simulated response to harvesting strategies in an exploited
ecosystem in the southwestern Gulf of México. Ecological Modelling 172: 421-432.
Arreguín-Sánchez, F., P. Del Monte-Luna & M. Zetina-Rejón. 2015. Climate Change Effects
on Aquatic Ecosystems and the Challenge for Fishery Management: Pink Shrimp of
the Southern Gulf of Mexico. Fisheries, 40 (1): 15 – 19.
Baeza-Rojano, E., P. Domínguez., J. M. Guerra-García., S. Capella., E. Noreña-Barroso., C.
Caamal-Monsrealcb & C. Rosas, 2013b. Marine gammarids (Crustacea:
Amphipoda): a new live prey to culture Octopus maya hatchlings. Aquaculture
Research, 44 (10): 1602 - 1612.
Baum, J. K. & B. Worm. 2009. Cascading top-down effects of changing oceanic predator
abundances. J. Anim. Ecol., 78: 699 - 714.
Bayle-Sempere, J. T., F. Arreguín-Sanchez., P. Sánchez-Jerez., D. Fernández-Jover., P.
Arechavala-López & D Izquierdo Gómez. 2013. Análisis y evaluación ecosistémicos
de la piscicultura marina con "Ecopath with Ecosim" (EwE). Pp: 39- 65. En: Blanco
J. A. (Edt.). Aplicaciones de modelos ecológicos a la gestión de recursos naturales.
OmniaScience, España. 215.
Benedek, Z., F. Jordán & A. Báldi. 2007. Topological keystone species complexes in
ecological interaction networks. Community Ecology 8(1), 1 - 7.
Bradley, E. & C. E. Bryan. 1975. Life history and fishery of the red snapper (Lutjanus
campechanus) in the Northwestern Gulf of Mexico. Proceedings of the Gulf and
Caribbean Fisheries Institute 27:77 - 106.
Branch, T. A., R. Watson., E. A. Fulton., S. Jennings., C. R. McGilliard., G. T. Pablico. & S.
R. Tracey. 2010. The trophic fingerprint of marine fisheries. Nature, 468 (7322): 431-
435.

76
Brulé, T. & L. Rodríguez-Canché. 1993. Food habits of juvenile of red groupers Epinephelus
morio (Valenciennes, 1828), from Campeche Bank, Yucatán, México. Bulletin of
Marine Science 52: 772 - 779.
Browder, J. A. 1993. A pilot model of the Gulf of Mexico continental shelf. Pp: 279 - 284.
En: Christensen, V. & Pauly, D (eds.). Trophic models of aquatics ecosystems.
ICLARM, Manila, Philippines. 390.
Borgatti, S. P., M. G. Everett y L.C. Freeman. 2002. Ucinet 6 for Windows: Software for
Social Network Analysis Harvard: Analytic Technologies.
Boucaud-Camou E & R. Boucher-Rodoni. 1983. Feeding and digestion in cephalopods. En:
Saleuddin ASM, Wilbur KM (eds.) The Mollusca, Physiology, Academic Press, New
York 15 (2): 149 - 187.
Burgos-Rosas, R & M. Pérez-Pérez 2006. Mero. Pp: 505-551. En: Arreguín-Sánchez, F., L.
Beléndez-Moreno., I. M. Gómez-Humarán., R. Solana-Sansore & C. Rangel-Dávalos
(eds.), Sustentabilidad y Pesca Responsable de México: Evaluación y Manejo. 544.
Cabrera-Neri, E. 2005. “Estructura trófica y flujos de energía en el ecosistema de la
plataforma continental de Tabasco, México”. Tesis de Maestría. Centro
Interdisciplinario de Ciencias Marinas Instituto Politécnico Nacional, La Paz Baja
California sur, México. 109.
Camber, C. I. 1955. A survey of the red snapper fishery of the Gulf of México, with special
reference to the Campeche Banks. Florida Board of Conservation, Technical Series
12: 63.
Chavéz, E., M. Garduño & F. Arreguín-Sánchez. 1993. Trophic Dynamic Structure of
Celestun Lagoon, Sourthern Gulf of México. Pp:186- 192. En: Christensen, V. &
Pauly, D (eds.). Trophic models of aquatics ecosystems. ICLARM, Manila,
Philippines. 390.
Christensen, V. 1992. Network analysis of trophic interactions in aquatic ecosystems. Tesis
de Doctorado. Royal Danish School of Pharmacy, Copenhagen. 55.

77
Christensen, V. 1995. Ecosystem maturity –towards quantification. Ecological Modelling.
77: 3 - 32.
Christensen, V., Pauly, D. 1992. ECOPATH II a software for balancing steady-state
ecosystem models and calculating network characteristics. Ecological Modelling, 61:
169 - 185.
Christensen V. & D. Paluy. 1993. Trophic models of aquatics ecosystems. Philippines,
ICLARM Conf. Proc. 26. 390.
Christensen V., C. J. Walters & D. Pauly. 2000. Ecopath with Ecosim Version 4, Help System
Univ. of British Columbia, Fisheries Center, Vancouver, Canada e ICLARM, Penang,
Malasia.
Christensen, V & C. J. Walters. 2004. Ecopath with Ecosim: methods, capabilities and
limitations. Ecological Modelling 172:109 - 139.
Christensen, V., C. J. Walter & D. Pauly. 2005. Ecopath with Ecosim: a User’s Guide.
Fisheries Centre, University of British Columbia, Vancouver., 154.
Christensen, V., Walters, C. J., Pauly, D., Forrest, R. 2008. Ecopath with Ecosim 6 User
Guide. Fisheries Centre, University of British Columbia, Vancouver., 235.
Chumba Segura L. 1995. Interacción trófica entre dos especies de sardina Opisthonema
oglinum (Le Seur 1818) y Harengula jaguana Poy 1865 (Osteichthyes: Cluoeida) que
coexisten en las costas de Celestun, Yucatán, México. Tesis de Maestría, Centro
Interdiciplinario de Ciencias Merinas, La Paz, Baja California Sur, México. 86.
Colinas-Sánchez, F & P. Briones-Fourzán. 1990. Alimentación de las Langostas Panulirus
guttatus y P. argus (Latreille 1804) en el caribe mexicano. Anales del Instituto de
Ciencia del Mar y Limnología. Universidad. Nacional. Autónoma. México. 17 (1):
89 - 106.
Conand, C. 2005. Sea cucumber biology, taxonomy, distribution, biology, conservation
status. En: Bruckner, A. (Ed) The Proceedings of the Technical wor-kshop on the
conservation of sea cucum-bers in the families Holothuridae and Stichopodidae.
NOAA Technical Memo-randum NMFSOPR 44, Silver Spring, MD 239.

78
CONAPESCA. Comisión Nacional de Acuacultura y Pesca. Anuarios Estadísticos de
Acuacultura y Pesca. 2014. Consultado 13 de febrero 2014. En línea:
https://www.gob.mx/conapesca/documentos/anuario-estadistico-de-acuacultura-y-
pesca
Connell, J. H. 1983. On the prevalence and relative importance of interspecific competition:
evidence from field experiments. American Naturalist 122:661-669.
Cruz, R., J. A. Baisre., E. Díaz., R. Brito., C. García., W. Blanco & C. Carrodegas. 1987.
Atlas Biológico-Pesquero de la Langosta en el Archipiélago Cubano. Rev. Cub.
Invest. Pesq. 125.
Cu-Salazar, G., I. Méndez-Loeza & J. C Pérez-Jiménez. 2010. Hábitos alimentarios de la
raya Dasyatis americana en el suroeste del Banco de Campeche. En Díaz-Sánchez A.
W., América Wendolyne., C. G. Aguilar., O. U. Mendoza-Vargas (Eds). Simposium
Nacional de Tiburones y Rayas. Acuario de Veracruz, Veracruz, 23-28 Dean Hartog,
J. C. 1980. Notes on the food of sea turtle: Eretmochelys imbricata (Linnaes) and
assimilation of nutrients in young green turtle Chelonia mydas L. Hepetol. J 3: 26 -
31
De la Cruz-Agüero, G. 1993. A preliminar Model of Mandinga Lagoon, Veracruz, México.
Pp: 193- 196. En: Christensen, V. & Pauly, D (eds.). Trophic models of aquatics
ecosystems. ICLARM, Manila, Philippines. 390.
DOF. Diario Oficial de la Federación. 2014. Consultado en línea el día 19 de mayo de 2014.
En:http://www.inapesca.gob.mx/portal/documentos/publicaciones/CARTA%20NA
CIONAL%20PESQUERA/24082012%20SAGARPA.pdf
Duarte, L. O & C. B. García. 2004. Trophic role of small pelagic fishes in a tropical upwelling
ecosystem. Ecol. Model. 172: 323 - 338.
Finucane, J. H. 1969. Ecology of the pampano (Trachinotus carolinus) and the permit (T.
falcatus) in Florida. Trans. Amer. Fish. Soc. 98 (3): 478 - 486.
Estrada, E. 2007. Characterization of topological keystone species local, global and “meso-
scale” centralities in food webs. Ecological Complexity 4, 48 - 57 pp. Frimodt, C.,

79
1995. Multilingual illustrated guide to the world's commercial warm water fish.
Fishing News Books, Osney Mead, Oxford, England. 215.
Froese. R. & D. Pauly. Editors. 1995. FishBase. World Wide Web electronic publication.
Galván-Piña, V. H. 2005. Impacto de la pesca en la estructura, función y productividad del
ecosistema de la Plataforma Continental de las costas de Jalisco y Colima, México.
Tesis de doctorado. Instituto Politécnico Nacional, Centro interdisciplinario de
Ciencias Marinas. La Paz Baja California sur, México. 106.
García, M. T., A. Yáñez-Arancibia., P. Sánchez-Gil & M. D. L. C. García-Abad. 1988.
Biología y ecología de Cynoscion arenarius Ginsburg, en las comunidades
demersales de la platafomia continental del sur del Golfo de México (Pisces:
Sciaenidae). Rev. Biol. Trop., 36 (1): 1 - 27
García, M. T., A. Yáñez-Arancibia., P. Sánchez-Gil & M. D. L. C. García-Abad. 1988.
Biología y ecología de Cynoscion nothus, en las comunidades demersales de la
plataforma continental del sur del Golfo de México (Pisces: Sciaenidae). Rev. Biol.
Trop, 36 (1): 29 - 54.
Giménez-Hurtado, E., B. Anderes., V. Moreno & R. Burgos. 2001. Aspectos de la conducta
alimentaria del mero (Epinephelus morio) del Banco de Campeche. INP. SAGARPA.
México. Ciencia Pesquera 15: 165-170.
Giménez-Hurtado. 2005. Análisis de la pesca del mero Epinephelus morio (Serranidae:
Pisces, Valenciennes 1928) en el Banco de Campeche. Tesis de doctorado. Centro de
Investigaciones Biológicas del Noroeste. La Paz Baja California sur, México. 93.
Gislason, H & M. Sinclair. 2000. Ecosystem Effects of Fishing. ICES Journal of Marine
Science, 57: 466 – 467.
Goldenberg, S., C. Landsea., A. Nuñez., & W. Gray. 2001. The recent increase in Atlantic
hurricane activity: Causes and implications. Science, 293: 474 – 479.
González, P., S. Zupanovic & H. Ramis. 1974a. Biología pesquera de la Cherna americana
del Banco de Campeche. INP/CIP, Cuba; Res. Invest., (1) pp. 107-111.

80
Hernández, A., Monroy, C., Moreno, V. y Jiménez, E. 1999. Informe de investigaciones
conjuntas México-Cuba sobre el mero (Epinephelus morio, Valenciennes, 1828) en
el Banco de Campeche. CRIP Yucalpetén, INP-SEPESCA-México, CIP-Cuba
Heymans, J. J., M. Coll., J. S. Link., Mackinson. S., Steenbeek. J., C. Walters & V.
Christensen. 2016. Best practice in Ecopath with Ecosim food-web models for
ecosystem-based management. Ecological Modelling. 33: 173- 184.
Izquierdo, L. R. y R. A. Hanneman. 2006. Introduction to the formal analysis of social
networks using mathematica Version 2. Publicado en línea. En:
http://www.luis.izqui.org/papers/Izquierdo_Hanneman_2006-version2.pdf
Laughlin, R. A. 1982. Feeding habits of the blue crab, Callinectes sapidus Rathbun, in the
Apalachicola Estuary, Florida. Bulletin of Marine Science, 32 (4): 807 - 822.
Lercari, D., L. Bergamino & O. Defeo. 2010. Trophic models in sandy beaches with
contrasting morphodynamics: Comparing ecosystem structure and biomass flow.
Ecological modeling, 221: 2751 - 2759.
Longhurst, A. R. & D. Pauly. 1987. Ecology of tropical oceans. Academic Press, Inc., San
Diego, California, U.S.A., 407.
MacArthur, R. H. 1955. Fluctuations of animal populations, and a measure of community
stability. Ecology 36(3):533-536.
Manickchand-Heileman, S., F. Arreguín-Sánchez, A. Lara-Domínguez & L. A Soto. 1998.
Energy flow and network analisis of Terminos Lagoon SW Gula of Mexico. Journal
of fish Biology. 53: 179 - 197.
Menge, B. A. 1982. Effects of feeding on the environment. En: Jangoux, M. & J. M.
Lawrence (Eds.) Echinoderm nutrition. Balkema, Rotterdam. 521-551.
Menge, B. A. 1995. Indirect effects in marine rocky intertidal interaction webs: patterns and
importance. Ecological Monographs, 65: 21 - 74.
Mexicano-Cíntora G., C. O. Leonce-Valencia., S. Salas & Ma. E. Vega-Cendejas. Recursos
pesqueros de Yucatán: Fichas técnicas y referencias bibliográficas. Centro de
Investigación y Estudios Avanzados (CIVESTAV). Mérida, Yucatán, México. 150.

81
Molina, L. J., A. Quiroga & A. de Federico. 2006. Taller de autoformación con programas
informáticos de análisis de redes sociales. España. 107.
Monroy, C., Giménez, E., Burgos, R. y Moreno, V. 2001. Informe de investigaciones
conjuntas México-Cuba sobre el mero (Epinephelus morio, Valenciennes, 1828) en
el Banco de Campeche. Ministerio de la Industria pesquera, Cuba. 43.
Naeem. S. C., F. S. Chapin III., R. Contanza., P. R. Ehrlich., F. B. Golley., D. U. Hooper., J.
H. Lawton., R. V. O’Neill., H. A. Mooney., O. E. Sala., A. J, Symstad., D. Tilman,
1999. La biodiversidad y elfuncionamiento de los ecosistemas: manteniendo los
procesos natural que sustentan la vida. Ecol. Society of America, Washington. 4: 1-
13
NOAA. National Oceanic and Atmospheric Administration. 2014. Consultado en línea 02 de
diciembre 2014. En: http://www.aoml.noaa.gov/
NOAA. National Oceanic and Atmospheric Administration. 2016. Consultado en línea 22 de
junio 2014. En: http://www.aoml.noaa.gov/phod/faq/amo_faq.php#faq_1
Odum, E. P. 1969. The strategy of ecosystem development. Science, 104: 262 - 270.
Olson, R & C. Boggs. 1986. Apex predation by yellowfin tuna (Thunnus albacares);
independent estimates from gastric evaluation and stomach contents, bionergetics,
and cesium concentrations. Canadian Journal of Fisheries and Aquatic Science 43:
1760 - 1775.
Pace, L .P., Cole, J .J., Carpenter, S. R., Kitchell, J. F. 1999. Trophic cascades revealed in
diverse ecosystems. Trends in Ecology and Evolution, 14 (12): 483 - 488.
Paine, R.T. 1966. Food web complexity and species diversity. American Naturalist, 100: 65-
75.
Pauly, D., Christensen, V., Dalsgaard, J., Froese & R., Torres, F. 1998. Fishing Down Marine
Food Webs. Science, 279: 860 - 863.
Polovina J. J. 1984a. An overview of the ECOPATH model. Fishbyte 2 (2): 5 - 7 pp

82
Power, M. E., Tilman, D., Estes, J. A., Menge, B. A., Bond, W. J., MilIs, L. S., Daily, G.,
Castilla, J. C., Lubchenco, J., Paine, R. T. 1996. Challenges in the Quest for
Keystones. BioScience, 46: 609 - 620.
Prigogine, I. & G. Nicolis. 1977. Self Organization in Non-Equilibrium Systems.
Raz-Guzmán, A & De la Lanza G. 1993. δ 13C del zooplancton, crustáceos decápodos y
anfípodos de laguna de Terminos, Campeche, México con referencias a fuentes de
alimentación y posición trófica. Cien Mar. 19 (2): 245 - 264.
Reyes-Yedra, C & A. Raz-Guzmán 2012. Dieta de las especies macroepibénticas
recolectadas en las bocas de la Laguna de Términos y la zona infralitoral, Campeche.
Ciencias Marinas 16 (46): 3 - 24.
Rincón-Sandoval L. A., T. Brulé,. J. L. Montero-Muñoz & E. Pérez-Díaz. 2009. Dieta de la
Rabirrubia Ocyurus chrysurus (Lutjanidae: Lutjaninae) y su variación temporal en la
costa de Yucatán, México. Pp: 207-218. En: Proceedings of the 62ad Caribbean
Fisheries Institute, Nov 2 - 6. Cumana, Venezuela.
Ritchie, E. G. & C. N. Johnson. 2009. Predator interactions, mesopredator release and
biodiversity conservation. Ecol. Lett., 12: 1 - 18.
Riviera Arriaga, E. 1990. Ecología trófica en dos poblaciones de peces tropicales costeros
(Polydactylus octonemus y Lutjanus synagris) del sur del Golfo de México. Tesis de
Licenciatura de Biólogo. Facultad de Ciencias, Universidad Nacional Autónoma de
México. México, D.F. 59.
Rosado-Solórzano, R. & S. A. Guzmán del Proó. 1998. Peliminary trophic structure model
for Tampamachoco lagoon, Veracruz, México. Ecological Modelling. 109 (1998):
141 - 154.
SAGARPA- Cinvestav. 2008. Informe Final del Plan de Manejo y Operación del comité de
Administración Pesquera de Escama y Pulpo. 2008. Mérida, Yucatán., 137.
Salcido-Guevara, L. A & F. Arreguín-Sánchez. 2007. A benthic ecosystem model of the
Sinaloa continental shelf, México. En: WJF le Quesne, F. Arreguín-Sánchez & S. J.

83
J. Heymans (Eds.) INCOFISH ecosystem models: transiting from Ecopath to
Ecospace. Fisheries Centre Research Report 15 (6): 170 - 188
Saloman, C. H & S. P. Naughton. 1983a. Food of king mackerel, Scomberomorus cavalla,
from the southeastern United States including the Gulf of Mexico. NOAA Tech.
Memo. NMFS-SEFC-126, U.S. Dept. of Commerce, NOAA, NMFS. 25.
Saloman, C. H & S. P. Naughton. 1983b. Food of Spanish mackerel, Scomberomorus
maculatus, from the Gulf of Mexico and southeastern seaboard of the United States.
NOAA Tech. Memo. NMFSSEFC-128, U.S. Dept. of Commerce, NOAA, NMFS.
22.
Saloman, C. H & S. P. Naughton. 1984. Food of crevalle jack (Caranx hippos) from Florida,
Louisiana, and Texas. NOAA Tech Memo. NMFS-SEFC-134, U.S. Dept. of
Commerce, NOAA, NMFS. 34.
Sánchez-Gill, P & A. Yáñez-Arancibia. 1999. Grupos eclógicos funcionales y recursos
pesqueros tropicales. 357- 389 pp. En Flores-Hernández D., P. Sánchez-Gill., J. C.
Seijo & F. Arreguín-Sánchez (Eds.). Análisis y diagnóstico de los recursos pesqueros
críticos del Golfo de México. Universidad Autónoma de Campeche. EPOMEX serie
científica, 7. 496 pp.
Sánchez Rueda, M. P. 1995. Análisis de los hábitos alimenticios de Mugil cephalus (Linneo)
y Mugil curema (Valen Ciennes) con base en la presencia de diatomeas. Tesis de
Maestría. Universidad Nacional Autónoma de México. 55.
Scheibling, R. 1982. Feeding habits of Oreaster reticulatus (Echinodermata: Asteroidea).
Bulletin of Marine Science 32 (2): 504 - 510.Smith, C. L. 1997. National Audubon
Society field guide to tropical marine fishes of the Caribbean, the Gulf of Mexico,
Florida, the Bahamas, and Bermuda. Alfred A. Knopf, Inc., New York. 720.
Schoener, T. W. 1983. Field experiments on interspecific competition. American Naturalist
122:240-285.

84
Shannon, L. J., Moloney, C., Jarre-Teichmann, A. & J. G., Field. 2004. Trophic flows in the
southern Benguela during the 1980’s and 1990’s. Journal of Marine Systems, 39: 83
-116.
SEMARNAT. Secretaría de Medio Ambiente y Recursos Naturales. Consultado 14 de
septiembre 2016. En: línea: www.semarnat.gob.mx
Solís-Ramírez, M. J. 1967. Aspectos biológicos del pulpo, Octopus maya. Instituto Nacional
de Investigaciones Biológico Pesqueras. 18: 1 - 90.
Stergiou, K. I & Karpouzi, V. S. 2002. Feeding habits and trophic levels of Mediterranean
fish. Reviews in Fish Biology and Fisheries, 11: 217 - 254.
Stoner, W. A. & R. J. Zimmerman. 1988. Food pathways associated with penaeid shrimps in
a mangrove fringed estuary. Fishery Bulletin 86 (3): 543 - 551.
Tapia-Garcia, M., A. Yáñez-Arancibia, F. Sánchez-Gil & M. D. L. C. García-Abad. 1988a.
Biología y ecología de Cynoscion arenarius, en las comunidades demersales de la
plataforma continental del sur del Golfo de México (Pisces: Sciaenidae). Rev. Biol.
Trop. 36 (1): 1 - 27.
Taylor, C. C. 1960. Temperature, growth and mortality, of the Pacific cockle. Journal du
Conseil 26: 117-124.
Trenkel, V. M & M. J. Rochet. 2003. Performance of indicators derived from abundance
estimates for detecting the impact of fishing on a fish community. Can. J. Fish. Aquat.
Sci. 60: 67 - 85.
Ulanowicz, R. E. 1986. Growth and Development: Ecosystem Phenomenology. Springer
Verlag (reprinted by Universe, 2000), New York. 203.
Vega-Cendejas, M. E., F. Arreguín-Sánchez & M. Hernández. 1993. Pp: 206 - 213. En:
Christensen, V. & Pauly, D (eds.). Trophic models of aquatics ecosystems. ICLARM,
Manila, Philippines. 390.
Vega-Cendejas, M. E, G. Mexicano-Cintora & A. M. Arce. 1997. Biology of the thread
herring Opisthonema oglinum (Pisces: Clupeidae) from a beach seine fishery of the
Campeche Bank, Mexico. Fish. Res. 30: 117 - 126.

85
Vega-Cendejas, M. E. & F. Arreguín-Sánchez. 2001. Energy fluxes in a mangrove
ecosistema from a coastal lagoon in Yucatan Peninsula, Mexico. Ecological
Modelling, 137 (2001): 119 - 133.
Vecchione M & V. A. Hand. 1989. Digestive-gland histology in paralarval squids
(Cephalopoda, Loliginidae). Fish Bull 87: 995 -100.
Vega-Cendejas, M. E. 1998. Trama trófica de la comunidad nectónica asociada al ecosistema
de manglar en el litoral norte de Yucatán. Tesis Doctorado, Facultad de Ciencias,
Universidad Nacional Autónoma de México, 170.
Vidal, L & M. Basurto. 2003. A preliminary trophic model of Bahía de Ascención, Quina
Roo, México. Fisheries Centre Research Reports. 11 (6): 256 - 264.
Vidal, L. & D. Pauly. 2004. Integration of subsystems models as a tool toward describing
feeding interactions and fisheries impacts in a large marine ecosystem, the Gulf of
Mexico. Ocean & Coastal Management. 47 (2004): 709 - 725.
Walters, C., V. Christensen & D. Pauly. 1997. Structuring dynamic models of exploited
ecosystems from trophic mass balance assessments. Review in Fish Biology and
Fisheries, 7 (2): 139 – 172.
Watt, K. E. F. 1964. Comments on long-term fluctuations of animal populations and
measures of community stability. Canadian Entomologist 96:1434-1442.
Wetzel, R. G. 1995. Death, detritus, and energy flow in aquatic ecosystems.
Williams, R. G. & M. J. Follows. 2003. Physical transport of nutrients and the main tenance
of biological production. Ocean Biogeochemistry, Spinger, 297.
Zetina-Rejón, M. J. & F. Arreguín-Sánchez. 2003. Flujos de energía y estructura trófica de
la Sonda de Campeche, Suroeste del Golfo de México. 55 - 62 pp. En: Wakida
Kusunoki, A., R. Solana Sansores & J. A. Uribe Martínez (Eds.) Memorias del III
Foro de Camarón del Golfo de México y del Mar Caribe. INPSAGARPA y Gob. del
Edo. de Campeche; México. 96.

86
Zetina-Rejón, M. J., N. J. Brown-Peterson., P. Del Monte-Luna., V. H. Cruz-Escalona & M.
S. Peterson. 2016. Trophic circulation in ecosystems. Ecosistemas y Recursos
Agropecuarios. 3 (8): 279- 291.
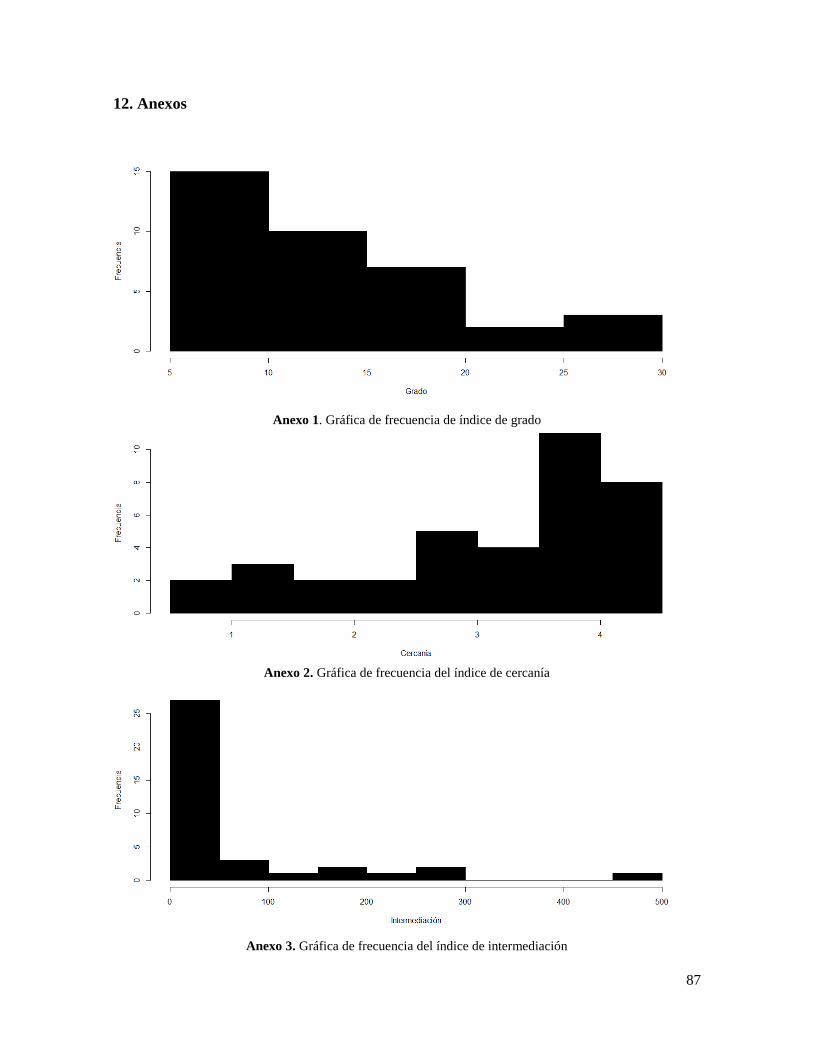
87
12. Anexos
Anexo 1. Gráfica de frecuencia de índice de grado
Anexo 2. Gráfica de frecuencia del índice de cercanía
Anexo 3. Gráfica de frecuencia del índice de intermediación
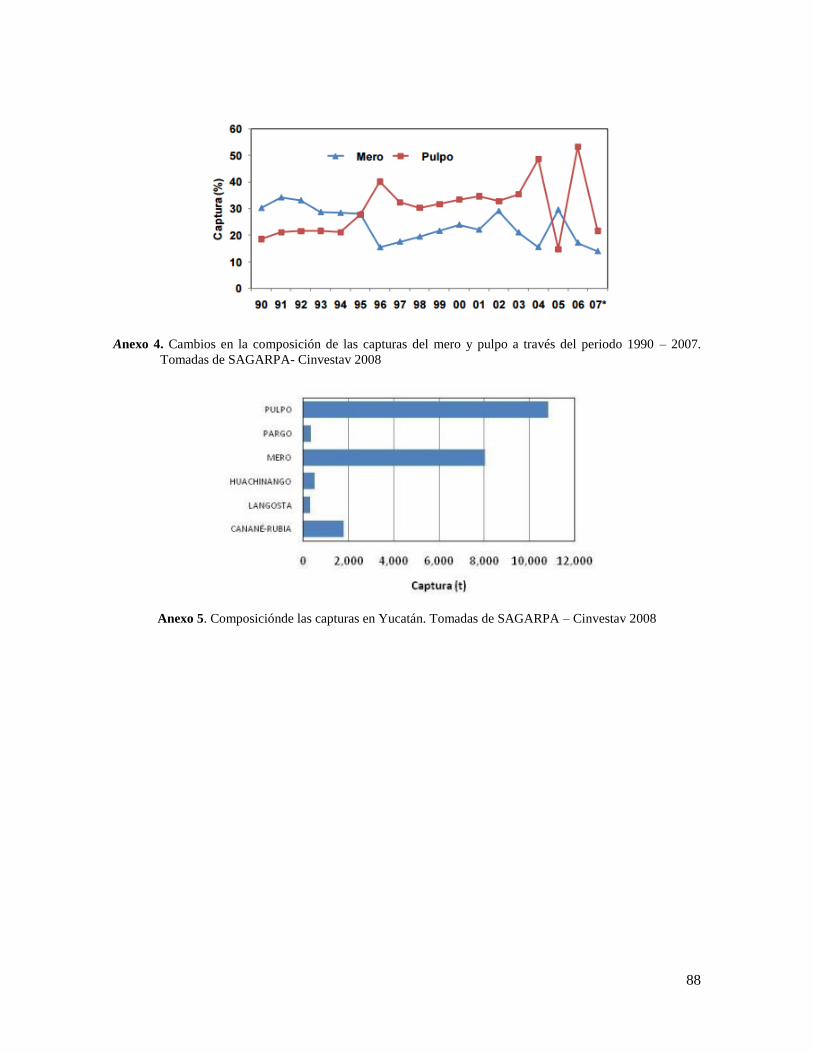
88
Anexo 4. Cambios en la composición de las capturas del mero y pulpo a través del periodo 1990 – 2007.
Tomadas de SAGARPA- Cinvestav 2008
Anexo 5. Composiciónde las capturas en Yucatán. Tomadas de SAGARPA – Cinvestav 2008

89





























