Embed Size (px)
Citation preview

Appraisal and reappraisal of cardiac therapy Edited by Arthur C. DeGraff and Julian Frieden
Electrophysiology and pharmacology of cardiac
arrhythmias. VIII. Cardiac effects of
diphenylhydantoin. 6
Andrew L. Wit, Ph.D.* Michael R. Rosen, M.D.* Brian F. Hoffman, M.D. New York. N. Y.
III. Cellular electrophysiological effects of DPH and possible mechanisms for its antiarrhythmic actions
The therapeutic actions of DPH probably are due to its effects on conduction, refractoriness, and automaticity of cardiac fibers. Such effects may be mediated both by direct actions of the drug on the heart and by effects exerted through the cardiac autonomic nerves.
Effects of DPH on conduction of the cardiac impulse; effects on resting membrane potential, action potential amplitude, phase 0 upstroke velocity (‘li,,,,,). DPH may exert several different actions on resting membrane potential, action potential . amplitude, and phase 0 upstroke velocity (V,,,) of both atria1 and Purkinje fibers. These actions seem to depend on (1) the concen- tration of DPH, (2) the [K] in the extracellular environment, and (3) the electrophysiological condition of the fibers; that is, whether resting membrane potential, action potential amplitude, and phase 0 upstroke velocity are abnormally low or within normal limits. The extracellular [K] has pronounced influences on the electrophysiologic actions of DPH. When [K] is low ((3 mM), low concentrations of DPH (1 to 3 pg/ml.) may increase the resting membrane potential, \jmar of phase 0, and action potential amplitude of both atriaP9+’ and Purkinje fibem5*. 53 The membrane
From the Department of Pharmacology, Columbia University College of Physicians and Surgeons, New York, N. Y.
Received for publication April 8, 1975.
Reprint requests to: Michael R. Roeen, M.D., Department of Pharma- cology, Columbia University, College of Physicians and Surgeons, 630 West 168th St., New York, N. Y. 1032.
*Dm. Wit and Rosen are Senior Investigator of the New York Heart Association.
September, 1975, Vol. 90, No. 3, pp. 397-404
responsiveness curve also may be shifted to the left and upward, indicating an increase in q,,,,, at all levels of membrane potential at which an action potential can be elicited. Conduction velocity also may be increased by DPH. All these effects are particularly pronounced if these parameters initially are reduced as by exposure to cold, mechanical trauma, or toxic concentrations of digitalis (Fig. 6). When resting membrane potential, Vi,,, of phase 0, and action potential amplitude are normal, a low concentration of DPH does not have any effect. When [K] is low, higher concentrations of DPH (5 to 20 pg/ml.) do not signifi.cantly alter the resting membrane potential, V,,, of phase 0, action potential ampli- tude, or the membrane responsiveness curve and therefore do not influence conduction of t,he action potential.
When the [K] concentration of the superfusate is at normal plasma values (4 to 5 mM), or slightly elevated, DPH has more of a depressant effect on the action potential of both atria1 and Purkinje fibers. Low concentrations (1 to. 3 pg/ ml.) do not increase the resting potential, V,,, of phase 0, or action potential amplitude as greatly as in low [K], although some improvement may still occur.50 Higher concentrations of DPH (5 to 20 pg/ml.) depress these parameters and also may slow conduction. The membrane responsiveness curve of Purkinje fibers is also shifted to the right, indicating a depression of Qm,, of phase 0 at all levels of membrane potential at which an action potential can be elicited .54 This depressant effect is not as great as that of quinidine or procaine amide.
The action of low concentrations of DPH (1 to 3 ,ug/ml.) to improve resting membrane potential,
American Head cJournal 397
Wit, Rosen, and Ho&an
0
- 70 -65
VM
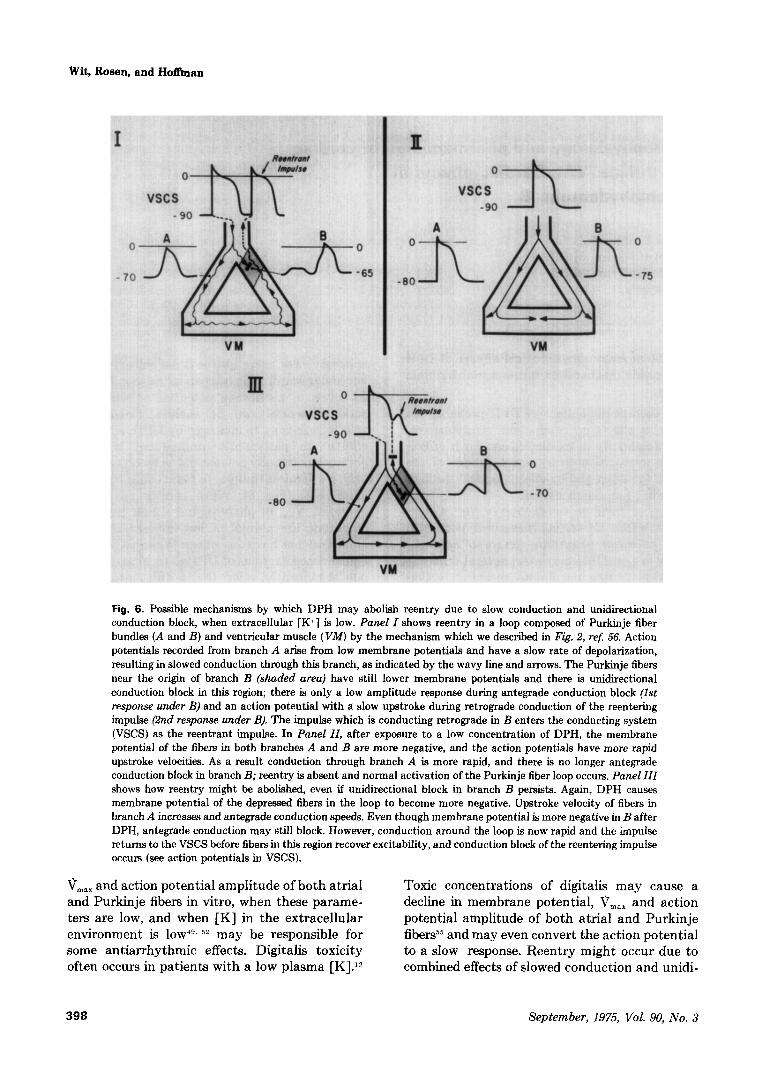
Fig. 6. Possible mechanisms by which DPH may abolish reentry due to slow conduction and unidirectional conduction block, when extracellular [K+ J is low. Panel I shows reentry in a loop composed of Purkinje fiber bundles (A and B) and ventricular muscle (Vhf) by the mechanism which we described in Fig. 2, ref. 56. Action potentials recorded from branch A arise from low membrane potentials and have a slow rate of depolarization, resulting in slowed conduction through this branch, as indicated by the wavy line and arrows. The Purkinje fibers near the origin of branch B (shaded area) have still lower membrane potentials and there is unidirectional conduction block in this region; there is only a low amplitude response during antegrade conduction block (1st response under B) and an action potential with a slow upstroke during retrograde conduction of the reentering impulse (2nd response under B). The impulse which is conducting retrograde in B enters the conducting system (VSCS) as the reentrant impulse. In Panel II, after exposure to a low concentration of DPH, the membrane potential of the fibers in both branches A and B are more negative, and the action potentials have more rapid upstroke velocities. As a result conduction through branch A is more rapid, and there is no longer antegrade conduction block in branch B; reentry is absent and normal activation of the Purkinje fiber loop occurs. Panel III shows how reentry might be abolished, even if unidirectional block in branch B persists. Again, DPH causes membrane potential of the depressed fibers in the loop to become more negative. Upstroke velocity of fibers in branch A increases and antegrade conduction speeds. Even though membrane potential is more negative in B after DPH, antegrade conduction may still block. However, conduction around the loop is now rapid and the impulse returns to the VSCS before fibers in this region recover excitability, and conduction block of the reentering impulse occurs (see action potentials in VSCS).
‘j,,, and action potential amplitude of both atria1 Toxic concentrations of digitalis may cause a
and Purkinje fibers in vitro, when these parame- decline in membrane potential, V,,, and action ters are low, and when [K] in the extracellular potential amplitude of both atria1 and Purkinje environment is 10~~“. 52 may be responsible for fibers”” and may even convert the action potential some antiarrhythmic effects. Digitalis toxicity to a slow response. Reentry might occur due to often occurs in patients with a low plasma [K].” combined effects of slowed conduction and unidi-
398 September, 1975, Vol. 90, No. 3

Electrophysiology and pharmacology of cnrdinc arrhythmias
rectional conduction block consequent to these changes by any of the mechanisms we have described previously (see Figs. 2 A and B, and Fig. 3 of ref. 56). Improvement of resting membrane potential or V,,, by DPH might abolish the unidirectional block, restoring a normal activa- tion sequence in the reentrant pathway, or speed conduction of the impulse in the reentrant pathway to such an extent. that it would return to its point of origin before the cardiac fibers at this locus had recovered excitability (Fig. 6). There- fore the reentering impulse would block before it could conduct back into the ventricles, and premature beats due to reentry would be abol- ished. Such an effect of DPH on reentry is strictly hypothetical and has not been demonstrated directly. The effect of DPH on conduction of slow responses resulting from toxic concentrations of digitalis has not been investigated. Since these actions of DPH to improve conduction may not occur when [K] is normal or elevated, under these conditions antiarrhythmic effects may not be exerted by this mechanism. Although DPH does slightly depress conduction in Purkinje fibers when [K] is in the normal range, this effect is not as great as the action of quinidine or procaine amide, which can block conduction in reentrant pathways. Therefore it is uncertain whether DPH can exert any antiarrhythmic effects on reentry by the same mechanism as quinidine and procaine amide. On the other hand, we do not know the effects of DPH on diseased cardiac fibers with low membrane potentials, or on cardiac fibers with slow response action potentials. Selective drug depression of such cardiac fibers, without signifi- cant depression of normal fibers, remains a possi- bility. Antiarrhythmic actions of DPH at normal plasma [K] may be exerted by other effects rather than on conduction of the primary impulse, such as effects on delayed afterdepolarizations which may cause digitalis arrhythmias,55 effects on conduction of premature impulses or effects on spontaneous diastolic depolarization (see below).
Effects on actionpotential duration, refractor-i- mss and conduction ofpremature impul..ses. The initiation of reentry and sustained reentrant arrhythmias may result from slow conduction and unidirectional block of premature impulses (Fig. 7).” Antiarrhythmic drugs may exert thera- peutic effects by modifying conduction of prema- ture impulses.
In both atria1 and Purkinje fibers, when the
American Heart Journal
membrane responsiveness curve is shifted to the left (low DPH concentration and low [K],) the effect of the drug is to speed conduction of premature impulses. This action might abolish slowed conduction and unidirectional block of premature impulses which result in reentry, although this has not yet been demonstrated.
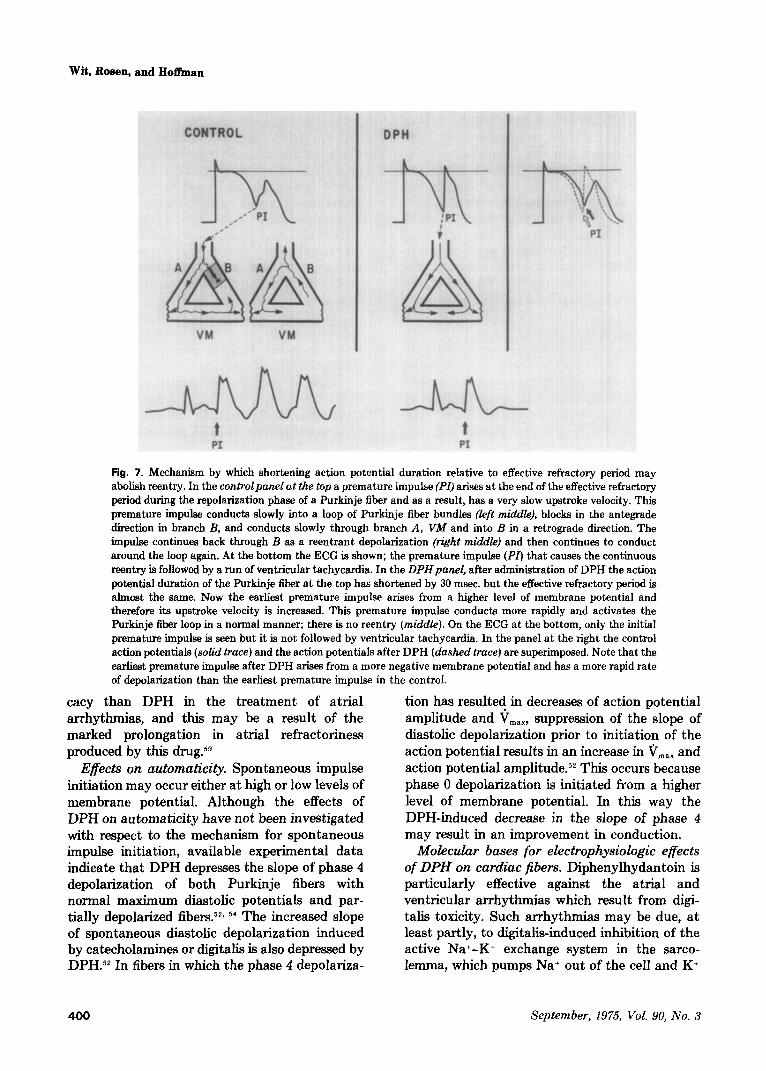
DPH also may improve conduction of prema- ture impulses in Purkinje fibers, even when the membrane response curve is not shifted, by signif- icantly shortening the action potential duration, an action similar to that of lidocaine. The effec- tive refractory period is also decreased although the reduction is not as great as t,he reduction in action potential duration (Fig. 7).“’ When this occurs, the earliest premature impulses which can be initiated during repolarization arise from higher membrane potentials and therefore have a greater V,,, and amplitude than do the earliest premature impulses initiated prior to drug admin- istration. As a result, premature impulses should conduct more rapidly after DPH, and both reentry due to slow conduction and unidirec- tional block of premature impulses would be abolished”” (Fig. 7). This effect occurs indepen- dently of [K] in the range of 3 to 5 mM and at a wide range of DPH concentrations (3 to 20 pg/ ml.). Shortening of the action potential duration and effective refractory period of Purkinje fibers might also have an antiarrhythmic effect by another mechanism. We have previously de- scribed mechanisms whereby reentry might result from inhomogenous conduction of premature impulses through regions where Purkinje fiber action potential durations and effective refrac- tory periods have been altered (see Fig. 8 and 9 in ref. 57). Our discussion of t.he possible antiar- rhythmic actions of lidocaine in these situations may also apply to DPH (see Figs. t: and 7 in ref. 58). Again, experimental data demonstrating the ability of DPH to abolish reent,ry t)y shortening action potential duration have nl)t been pub- lished.
DPH does not change the actton potential duration and effective refractory period of atria1 fibers as much as in ventricular fibers. Rather, atria1 fibers show minimal changes in these parameters after DPH administration. This lack of a prominent effect on the effective refractory period of atria1 fibers may be a cause of the ineffectiveness of DPH against most atria1 arrhythmias. Quinidine has a much greater effi-
399
Wit, Rosen, and Hoffman
PI
QPW
m ‘PI :
t PI
Fig. 7. Mechanism by which shortening action potential duration relative to effective refractory period may abolish reentry. In the controlpanel at the top a premature impulse (Pr) arises at the end of the effective refractory period during the repolarization phase of a Purkinje fiber and as a result, has a very slow upstroke velocity. This premature impulse conducts slowly into a loop of Purkinje fiber bundles (left middle), blocks in the antegrade direction in branch B, and conducts slowly through branch A, VM and into B in a retrograde direction. The impulse continues back through B as a reentrant depolarization (right middle) and then continues to conduct around the loop again. At the bottom the ECG is shown; the premature impulse (PZ) that causes the continuous reentry is followed by a run of ventricular tachycardia. In the DPHpanel, after administration of DPH the action potential duration of the Purkinje fiber at the top has shortened by 30 msec. but the effective refractory period is almost the same. Now the earliest premature impulse arises from a higher level of membrane potential and therefore its upstroke velocity is increased. This premature impulse conducts more rapidly and activates the Purkinje fiber loop in a normal manner; there is no reentry (middle). On the ECG at the bottom, only the initial premature impulse is seen but it is not followed by ventricular tachycardia. In the panel at the right the control action potentials (soZid truce) and the action potentials after DPH (dashed truce) are superimposed. Note that the earliest premature impulse after DPH arisea from a more negative membrane potential and has a more rapid rate of depolarization than the earliest premature impulse in the control.
cacy than DPH in the treatment of atria1 arrhythmias, and this may be a result of the marked prolongation in atria1 refractoriness produced by this drug.$”
Effects on automaticity. Spontaneous impulse initiation may occur either at high or low levels of membrane potential. Although the effects of DPH on automaticity have not been investigated with respect to the mechanism for spontaneous impulse initiation, available experimental data indicate that DPH depresses the slope of phase 4 depolarization of both Purkinje fibers with normal maximum diastolic potentials and par- tially depolarized fibers.“‘, 54 The increased slope of spontaneous diastolic depolarization induced by catecholamines or digitalis is also depressed by DPH.52 In fibers in which the phase 4 depolariza-
tion has resulted in decreases of action potential amplitude and V,,,, suppression of the slope of diastolic depolarization prior to initiation of the action potential results in an increase in V,,, and action potential amplitude.52 This occurs because phase 0 depolarization is initiated from a higher level of membrane potential. In this way the DPH-induced decrease in the slope of phase 4 may result in an improvement in conduction.
Molecular bases for electrophysiologic effects of DPH on cardiac fibers. Diphenylhydantoin is particularly effective against the atria1 and ventricular arrhythmias which result from digi- talis toxicity. Such arrhythmias may be due, at least partly, to digitalis-induced inhibition of the active Na+-K+ exchange system in the sarco- lemma, which pumps Na+ out of the cell and K+
400 September, 1975, Vol. 90, No. 3

Electrophysiology and pharmacology of cardiac arrhythmias
back into the cell.“” Such inhibition would result in an increase in intracellular [Na] and a decline in intracellular [K]. An increase in intracellular [Na] might result in a decrease in the,electro- chemical gradient for Na and therefore V,,, and action potential amplitude would decline. A decrease in [Kli would result in a decline in resting membrane potential and this would partially or completely inactivate the fast inward Na+ current. These effects may result in conduc- tion disturbances, reentry, or abnormal automa- ticity.
The loss of myocardial K+ during digitalis toxicity is reflected by a marked increase in coronary sinus venous [K].“’ Helfant and asso- ciate@* have reported that DPH reverses this loss of K+ from the heart at the time it is abolishing the digitalis-induced arrhythmias.62 DPH preven- tion of K+ loss and Na+ accumulation by guinea pig myocardial cells after digitalis intoxication also has been reported.63 Such effects may result from antagonism by DPH of the digitalis-induced inhibition of the Nat-K+ pump, and restoration of more Na+-K+ exchange and gradients. The resultant decrease in [Na], and increase in [K], would restore resting membrane potential to normal, increasing the availability of the Na+ carrier system. Unfortunately other studies have failed to demonstrate any ability of DPH to prevent K6 efflux during digitalis intoxication, and this question must be clarified.64-67 If DPH can stimulate Nat-K+ exchange pumping, this effect might also explain the action of DPH to retard the decline of maximum diastolic potential in Purkinje fibers exposed to hypoxia. Hypoxia
depresses Na+-K+ pumping, and causes an increase in [Na], and a decrease in [K]i.6R
In addition to possible effects on active ion transport, DPH might affect membrane ionic conductances, although as yet there are no studies to demonstrate this. A possible increase in Na+ conductance has been postulated to mediate the increase in V,,, seen in depressed cardiac fibers after exposure to DPH,“’ and an increase in K+ conductance has been suggested to mediate its depressant effect on spontaneous diastolic depo- larization and the acceleration of repolarization in Purkinje fibers.@ As we have mentioned above, DPH can decrease Vma, under certain conditions and this effect could result from a depressant effect of the drug on Na+ conductance and the fast inward current.“’ The depressant effect of
American Heart Journal
DPH on V,,, of atria1 fibers can be reversed by increasing [Na] in the perfusate, suggesting that this depressant effect is due to a drug-induced decline in Na+ conductance.“”
IV. Effects of DPH on the nervous system as a
mechanism for its antiarrhythmic actions
Cardiac arrhythmias associated with patholog- ical conditions such as myocardial infarction, as well as digitalis-induced arrhythmias, may result in part from altered nervous system activity”, 7E and the effects of the nervous system on electro- physiological events in the heart. After myocar- dial infarction, or during digitalis toxicity, there is an increase in activity in the cardiac sympathetic nerves.T’, 72 This may enhance spontaneous dias- tolic depolarization in latent pacemaker fibers, alter refractoriness in cardiac fibers and result in reentry, or enhance the slow inward current in areas where cells have low membrane potentials and thus facilitate reentry. DPH decreases the efferent sympathetic activity in the cardiac nerves which may be responsible for arrhythmias accompanying digitalis toxicity.‘” This decrease in sympathetic activity results from a depressant effect of DPH on the sympathetic centers in the central nervous system?” and may be an impor- tant antiarrhythmic effect of the drug.
V. Effects of DPH on the electrophysiology of the
In Situ Heart
Effects on the SA node. When administered to experimental animals or humans DPH has a variable effect on sinus rate which may increase, remain unaltered, or decrease.“. ii jr. These effects may be mediated through the autonomic nervous systemT6 and may depend on whether the sinus node is normal or diseased. That direct effects of DPH on the sinus node action potential probably are not important has been shown in studies on isolated rabbit atria, in which the action potential and slope of phase 4 depolarization of sinus node cells were unaltered by a wide range of DPH concentrations.a’g
The acceleratory effect of DPH on sinus impulse initiation, which has been observed in some instances, may be provoked reflexly through alterations in autonomic tone resulting from slight hypotension, or pain at the injection site,‘” or may be exerted through effects of DPH on the central or autonomic nervous systems which have not yet been described.76 It has been suggested
401

Wit, Rosen, and Hoi&an
that DPH has vagolytic effects, but this sugges- tion has been disputed/g
High plasma levels of DPH which may produce undesirable neurological symptoms usually do not depress the sinus node in patients without sinus node disease. However, there have been reports of severe sinus bradycardia or sinus arrest as a toxic response to DPH, although these actions appear to be unusual.77 Preexisting sinus node disease may be responsible for the drug- induced depression of sinus node function. This is suggested since experimental procedures such as stretch or mechanical trauma, which cause sinus node depression in the isolated rabbit heart, render the sinus node more easily depressed by high DPH concentrations.4g An additional factor to be considered is the commercial solvent for DPH, a solution of propylene glycol and ethyl alcohol with a pH of 11 to 12, which also may depress sinus node function.7s
Effects on atrium. It is difficult to determine which of the several electrophysiological effects of DPH on atria1 transmembrane potentials demon- strated in vitro are important for the in situ heart. DPH does not markedly slow atria1 conduction velocity,79 therefore it probably does not signifi- cantly decrease V,,, of normal atria1 fibers in situ. The effect of DPH on atria1 refractoriness is uncertain; both shortening of atria1 refractori- nesssO and no changeT5* 76 have been reported. Since the commercial diluent for DPH may prolong the effective refractory period, this may obscure any accelerating effect of the drug on repolarization. DPH has also been reported to decrease atria1 diastolic thresholds0 and to increase itTg, 76 Again, the effects of the drug must be separated from the combined effects of drug and commercial diluent.
Effects on the AV node. Antiarrhythmic concentrations of DPH have a variable effect on conduction and refractoriness of the normal AV node in both experimental animals and in humans. DPH often accelerates AV nodal conduction slightly.8* ~3 ‘es 81. 82 However, in some instances no effect or a slight slowing is observed.8s 75 Acceleration of AV nodal conduc- tion is not dependent on changes in heart rate since it also occurs when atria1 rate is maintained constant.75, 82 Concomitantly, the AV nodal effec- tive and functional refractory periods may be reduced but this effect, too, is not invariable.75 Even when large doses of DPH are administered,
significant depression of AV nodal conduction usually does not occur and therefore this is not a commonly encountered toxic effect of the drug.
The acceleratory effects of DPH on AV nodal conduction are more marked in the canine heart after AV nodal conduction time has been pro- longed by toxic concentrations of digitalis8’ DPH concentrations which abolish the digitalis- induced ventricular arrhythmias consistently reduce AV nodal conduction time under this experimental condition. DPH also accelerates AV nodal conduction in the canine heart after it has been slowed by toxic concentrations of procaine amide.83 The lack of deleterious effects of DPH on AV nodal conduction in the setting of digitalis toxicity, and the ability to overcome the digitalis- induced impairment of AV nodal conduction in the experimental animal suggest an advantage in utilizing DPH in humans to treat digitalis- induced arrhythmias. There is no published data to indicate that DPH similarly can overcome digitalis-induced AV nodal impairment in hu- mans but DPH, unlike procaine amide, does not seem to exacerbate AV nodal conduction impair- ment during treatment of digitalis-toxic arrhyth- mias.
The observation that DPH may improve depressed AV nodal conduction in animals does not indicate that DPH should be used as a therapeutic intervention for AV nodal conduction disturbances due to digitalis or other etiologies in humans. The effects of DPH, when AV nodal conduction is depressed by disease, is not known. Certainly, caution should be utilized in the administration of DPH to patients with diseased AV nodes and depressed conduction since several clinical reports indicate a worsening of AV nodal conduction which may progress to complete AV block in this circumstance.R’
The possibility of an acceleratory effect of DPH on AV nodal conduction suggests that caution should be used if the drug is administered for ventricular arrhythmias in the presence of atria1 flutter or fibrillation. One would predict that DPH could increase the number of impulses successfully traversing the AV node and therefore increase ventricular rate. Clinical experience has actually demonstrated a decrease in the ventric- ular response to atria1 fibrillations in some patients.” If DPH, in fact, accelerates AV nodal conduction, increased concealment of atria1 impulses in the AV node may be the mechanism
402 September, 1975, Vol. 90, No. 3

Electrophysiology and pharmacology of cardiac arrhythmias
for the slowed ventricular response during atria1 fibrillation.
The mechanism by which DPH sometimes accelerates AV nodal conduction in the experi- mental animal and in humans is uncertain. Improvement in AV nodal conduction is not seen in the denervated canine heartT6 or in the isolated blood perfused canine heart,S4 suggesting that the acceleratory effect is exerted via the nervous system, possibly a decrease in vagal activity, and not through a direct effect of the drug on the AV node. Further clarification of the exact mecha- nism is required.
Effects on the ventricular specialized con- ducting system and ventricular muscle. In the intact or denervated canine heart, and in the human heart, as well, therapeutic and toxic levels of DPH do not significantly slow conduction in the His-Purkinje system (do not alter the H-V intervals in the His bundle electrogram) nor do they slow conduction in ventricular muscle; there is no significant effect of the drug on the electro- cardiographic QRS complex.“. x 81. 92 This is consistent with the observations that DPH does not markedly depress v,,,,, of phase 0 of the Purkinje fiber or ventricular muscle action poten- tial. The effect of DPH on the diseased conducting system is uncertain and caution should be taken when administering the drug to patients with intraventricular conduction defects or bundle branch block because of the possibility of further conduction depression.
Even though DPH may sometimes improve v,,X and conduction velocity in isolated Purkinje fiber bundles, as described above, only one study has shown DPH to speed His-Purkinje conduc- tion in the intact heart; this occurred in the canine heart after conduction had been depressed by toxic concentrations of procaine amide.83
DPH shortens the relative and effective refrac- tory period and improves conduction of prema- ture impulses in the ventricular conducting system and ventricular muscle in both canine and human hearts.:“. x0. k:> This effect is probably due to the shortening of the action potential duration which was described above.
DPH will also suppress automaticity of ventric- ular pacemakers in the intact canine heart. Such automaticity can be measured indirectly by determining the idioventricular rate after vagal stimulation has arrested the atria, or in animals with complete A-V block. DPH has minimal
American Heart Journal
depressant effects on ventricular rate unless it has been enhanced by digitalis.“”
Summary
DPH is highly effective against both atria1 and ventricular arrhythmias resulting from digitalis toxicity and should be considered one of the primary antiarrhythmic drugs in this clinical situation. Clinical studies have indicated that DPH, for the most part, is ineffective against atria1 arrhythmias and not markedly effective against ventricular arrhythmias associated with acute or chronic cardiac disease. For the latter ventricular arrhythmias, DPH is usually consid- ered only after other antiarrhythmic drug therapy has failed. Studies on the mechanisms responsible for the antiarrhythmic effects of DPH are, as yet, inconclusive. There is good evidence that its actions differ from those of the commonly used drugs with local anesthetic effects. The extent to which it exerts direct effects on the heart which are therapeutically relevant remains to be demonstrated; further, there is solid evidence that its effects on the central nervous system may be of prime importance in its antiar- rhythmic efficacy.
REFERENCES
49.
50.
51.
52.
53.
54.
55.
56.
57.
Strauss, H. C., Bigger, J. T., Jr., Bassett, A. L., and Hoffman, B. F.: Actions of diphenylhydantoin on the electrical properties of isolated rabbit and canine atria, Circ. Rea. 23:463, 1968. Jensen, R. A., and Katzung, B. G.: Electrophysiological actions of diphenylhydantoin on rabbit atria: Depen- dence on stimulation frequency, potassium and sodium, Circ. Res. 26:17, 1970. Singh, B. N., and Vaughan Williams, E. M.: Effect of altering potassium concentration on the action of lido- Caine and diphenylhydantoin on rabbit atria1 and ventricular muscle, Circ. Rea. 29:286, 1971. Bigger, J. T., Jr., Bassett, A. L., and Hoffman, B. F.: Electrophysiological effects of diphenylhydantoin on canine Purkinje fibers, Circ. Res. 22:221, 1968. Bassett, A. L., Bigger, J. T., Jr., and Hoffman, B. F.: “Protective” action of diphenylhydantoin on canine Purkinje fibers during hypoxia, J. Pharmacol. Exp. Ther. 173:336, 1970. Rosen, M. R., Danilo, P., Alonso, M., and Pippenger, C.: Effect,s of diphenylhydantoin on the electrophysiologic properties of blood-superfused Purkinje fibers, Fed. Proc. 34:775, 1975. Rosen, M. R., Wit, A. L., and Hoffman, B. F.: Cardiac antiarrhythmic and toxic effects of digitalis, AM. HEART 3. 89:391, 1975. Wit, A. L., Rosen, M. R., and Hoffman, B. F.: Electro- physiology and pharmacology of cardiac arrhythmias. II. Relationship of normal and abnormal electrical activity of cardiac fibers to the gene& of arrhythmias. B. Reen- try, Section I, AM. HEARTJ. 88:664,1974. Wit, A. L., Rosen, M. R., and Hoffman, R. F.: Electro-
403

Wit, Rosen, and Hofhan
56.
59.
60.
61.
62.
63.
64.
65.
66.
67.
66.
69.
70.
71.
physiology and pharmacology of cardiac arrhythmias II. Relationship of normal and abnormal electrical activity of cardiac fibers to the genesis of arrhythmias. B. Reen- try, Section II, AM. HEART J. 88:798, 1974. Rosen, M. R., Wit, A. L., and Hoffman, B. F.: Cardiac antiarrhythmic etfects of lidocaine, AM. HEART J. 89:526, 1975. Hoffman, B. F., Rosen, M. R., and Wit, A. L.: Cardiac effects of quinidine and procaine amide, AM. HEART J. 89:804, 1975 Part A, and 90:117, 1975 Part B. Lee, K. S., and Klaus, W.: Subcellular bases for the mechanism of inotropic action of cardiac glycosides, Pharmacol. Rev. 23:193, 1971. Regan, T. J., Tahners, F. N., and Hellerns, H. K.: Myocardial transfer of sodium and potassium: Effect of acetylstrophanthadin in normal dogs, J. Clin. Invest. 35:1220, 1956. Helfant, R. H., Ricciutti, M. A., Scherlag, B. J., and Damato, A. N.: Effect of diphenylhydantoin sodium (Dilantin) on myocardial A-V potassium difference, Am. J. Physiol. 214:880,1968. Bashkin, S. I., Dotta, S., and Marks, B. H.: The effects of diphenylhydantoin and potassium on the biological activity of ouabain in the guinea pig heart, Br. J. Pharmacol. 47:85, 1973. Miller, D. T., and Gilmore, J. P.: Influence of diphenyl- hydantoin on the inotropic and potassium losing effects of acetylstrophanthid&AM. HEART J. 80:223, 1970. Goldstein. R. E.. Penzotti. S. C.. Kuehl. K. S.. Prindle. K. H., Hall, C. ‘A., Titus, E. d., and ‘Epstein, S. E.; Correlation of antiarrhythmic effects of diphenylhydan- toin with digoxin-induced changes on myocardial contractility, sodium-potassium adenosine triphosphate activity and potassium efflux, Circ. Res. 33:175, 1973. Godfraind, T., Leane, M., and Pousti, A.: The action of diphenylhydantoin upon drug binding, ionic effects and inotropic action of ouabain, Arch. Int. Pharmacodyn. 191:66, 1971. Spain, R. C., and Chidsey, C. A.: Myocardial Na/K adenosine triphosphate activity during reversal of ouabain toxicity with diphenylhydantoin, J. Pharmacol. Exp. Ther. 179:594, 1971. Hercus, V. M., McDowall, R. J., and Mendel, D.: Sodium exchanges in cardiac muscle, J. Physiol. (Lond.) 129:177, 1955. Bigger, J. T., Jr.: Ant&rhythmic drugs in ischemic heart disease, Hosp. Prac 7:69, 1972. Lipicky, R. J., Gilbert, D. L., and Stillman, I. M.: Diphenylhydantoin inhibition of sodium conductance in squid giant axon, Proc. Nat. Acad. Sci. 69:1758, 1972. Gilis, R. A.: Role of the nervous system in the arrhyth- mias produced by coronary artery occlusion in the cat, AM. HEART J. 81:677, 1971.
72.
73.
74.
75.
76.
77.
78.
79.
60.
81.
82.
6.3.
64.
65.
Gillis, R. A.: Cardiac sympathetic nerve activity: Changes induced by ouabain and propranolol, Science 166:508,1969. Gillis, R. A., McClellan, J. R., Sauer, T. S., and Stan- daert, F. G.: Depression of cardiac sympathetic nerve activity by diphenylhydantoin, J. Pbarmacol. Exp. Ther. 173:599, 1971. Evans, D. E., and Gillis, R. A.: Effect of diphenylhydan- toin and lidocaine on cardiac arrhythmias induced by hypothalamic stimulation, J. Pharmacol. Exp. Ther. 191:506, 1974. Caracta, A. R., Damato, A. N., Josephson, M. E., Ricciutti, M. A., Gallagher, J. J., and Lau, S. H.: Electrophysiologic properties of diphenylhydantoin, Cir- culation 47:1234, 1973. Roaati, R. A., Alexander, J. A., Schaal, S. F., and Wallace, A. G.: Influence of diphenylhydantoin on elec- trophysiological properties of the canine heart, Circ. Res. 21:757, 1967. Unger, A. H., and Sklaroff, H. J.: Fatalities following intravenous use of sodium diphenylhydantoin for cardiac arrhythmias. Report of two cases, J. A. M. A. 200:159, 1967. Louis, S., Kutt, H., and McDowell, F.: Cardiocirculatory changes caused by intravenous dilantin and its solvents, AM. HEART J. 74523, 1967. Russell, J. M., and Harvey, S. C.: Effects of diphenylhy- dantoin on canine atria and A-V conducting system, Arch. Int. Pharmacodyn. 182:219, 1969. Bigger, J. T., Jr., Weinberg, D. I., Kovalik, A. T. W., Harris, P. D., Cranefield, P. F., and Hoffman, B. F.: Effects of diphenylhydantoin on excitability and auto- maticity in the canine heart, Circ. Res. 26:1,1970. Helfant, R. H., Scherlag, B. J., and Daniato, A. N.: The electrophysiological properties of diphenylhydantoin sodium as compared to procaine amide in the normal and digitalis intoxicated heart, Circulation 36:108, 1967. Damato. A. N.. Berkowitz. W. D.. Patton, R. D.. and Lau, S. H.: The’effect of diphenylbydantoin’on atribven- tricular and intraventricular conduction in man, AM. HEART J. 79:51, 1970. Helfant, R. H., Scherlag, B. J., and Damato, A. N.: Use of diphenylhydantoin sodium to dissociate the effects of procainamide on automaticity and conduction in the normal and arrhythmic heart, Am. J. Cardiol. 20:820, 1967. Sasyniuk, B. I., and Dresel, P. E.: The effect of diphenyl- hvdantoin on conduction in isolated, blood-nerfused dog hearts, J. Pharmacol. Exp. Ther. 161:191, i968. - Bissett, J. K., De Soyza, N. D. B., Kane, J. J., and Murphy, M. L.: Improved intraventricular conduction of premature beats after diphenylhydantoin, Am. J. Cardiol. 33:493, 1974.
September, 1975, Vol. 90, No. 3