Embed Size (px)
Citation preview

Deep Ocean EcosystemsFrancesco Canganella, University of Tuscia, Italy
Chiaki Kato, Japan Marine Science and Technology Centre, Japan
The term ‘deep ocean’ typically describes any marine ecosystem located at depths greater
than 500 metres. This environment is characterized by an elevated hydrostatic pressure, an
average temperature of 2–48C, the absence of sunlight, and the scarce availability of
organic food. Specific organisms are associated with the deepest areas, and pressure-
adapted animals as well as microorganisms inhabit these peculiar ecosystems.
The Deep Ocean Ecosystem
It is well known that the Earth’s biosphere is mainlycomposed of water; nevertheless, despite the efforts ofresearchers and improvements in scientific methods, only asmall portion of the oceans is easily accessible and so themajority of the marine world is still unknown. In profile,the ocean can be divided into three layers: an upper, wind-mixed zone; a multilayer zone, and a uniform, deep-waterregion.
It will be useful to give an idea of the extent of deepoceans as derived from statistical interpretation: (1) theirmaximum depth is almost 11 km and about 75% of thetotal area has a depth ranging from 3 to 6 km; (2) the meanheight of land above sea level is about 800 m, whereas themean depth of the seas is 3.8 km; (3) the total weight of seawater is estimated at 1.5� 1018 tonnes. The distribution ofdeep sea regions throughout the world is shown in Figure 1.
The deep ocean ecosystem, as well as the oceans ingeneral, can be described in terms of physico-chemicalparameters. Although the deep ocean structure is not asvariable as that of shallow waters, many changes andsignificant motions can still occur.
The temperature structure of the oceans is the best-known parameter: all waters warmer than 58C are confinedto the upper layers (depth5 1000 m); the rest of the oceanis cold, even within the tropics and at the equator. At theshallowest depths, vertical motions that can producemixing and temperature changing often occur, exceptwhen a thermocline acts as a barrier: in that case anytransfer through the water layers is very difficult. At greaterdepths there is no seasonal variation and the distribution isthe same all year round. The density of the water isdetermined mainly by its temperature. Both parameters arequite stable in deep oceans, where the heaviest water is atthe bottom. The more stable this layering, the more energyis required to rearrange density and temperature distribu-tions. That is why thermoclines are more stable as depthincreases (and therefore density also increases).
It is not possible to describe the salinity of the oceans inthe same way as can be done for temperature, becausesalinity affects the density to a lesser extent than does
temperature. In the deep, both temperature and salinity canonly be altered by mixing with other water masses. The ratioof these two parameters is usually characteristic for aparticular water mass and, once it is acquired at the surface,no significant changes occur in distribution during sinking.
The concentration of dissolved oxygen in the deep oceanis much lower than on the surface: between 500 and 1000 mit can be measured at a minimal concentration of0.5 mL L2 1. Higher amounts are typically found at theabyssal floor owing to the scarcity of living organisms andto their low metabolic rates.
Whereas the planetary wind system drives the surfacecirculation of the ocean, the circulation in the deep seadepends on density differences, largely determined bywater temperature and salinity. Studies of thermohalinecirculation in the deep ocean concentrate on identifiablewater masses classified by their source region and depth ofoccurrence. The mixing of North Atlantic deep water withAntarctic intermediate water and Antarctic bottom waterproduces the largest water mass in the deep sea, with anaverage temperature and salinity of 1.58C and 34.7%,respectively.
Devices for the Investigation of DeepOceans
The development of devices for the oceanographic researchof deep sea ecosystems made astonishing advances duringthe twentieth century and new insights into the deep oceanswere achieved. Most of the progress in the design of marineinstruments and vehicles is related to problems such as theinstallation of large decks, the development of underwaterinstrumentation connected to sonar, and the use ofcameras suspended from a ship by wire and independentvideo systems.
Biological samples are important devices for zoological,botanical and microbial investigations of the deep oceans.The main purpose of the investigator after sampling isusually to get some idea of the total population or
Article Contents
Secondary article
. The Deep Ocean Ecosystem
. Devices for the Investigation of Deep Oceans
. Representative Deep Sea Organisms
. Present and Future Perspectives
1ENCYCLOPEDIA OF LIFE SCIENCES © 2002, John Wiley & Sons, Ltd. www.els.net

community at the position, depth and time at which thesample was taken.
A variety of vessels have been used by oceanographers;particularly significant achievements were obtained by theuse of manned submersibles. Perhaps the most famousscientific submersible is the bathyscaphe Trieste, operatedby the US Navy, with which Swiss scientist AugustePiccard reached the deepest point of the oceans, theMariana Trench at 10 910 m.
Since 1964 the deep submergence vehicle (DSV) Alvinhas been operated by the Woods Hole OceanographicInstitution of Cape Cod (Massachusetts, USA). Thisvehicle (Figure 2) was a pioneer of modern deep sea researchand fundamental discoveries were made following itslaunch. A typical eight-hour dive takes two scientists and apilot as deep as 4500 m. At this depth it takes two hours forthe submarine to reach the bottom and another two toreturn to the surface. The vessel is equipped with threevideos, two cameras, twelve lights and two robotic arms for
sampling. A sample basket is mounted on the front of thesubmarine to carry a variety of instruments as well as to
Figure 1 The distribution of deep sea regions with depths greater than 3000–4000 m throughout the world. Modified after Herring and Clarke (1971).
Figure 2 The mannedsubmersible Alvin built in 1964and operatedby theWoods Hole Oceanographic Institution.
Deep Ocean Ecosystems
2

store sediment cores and probes. Another high-techresearch submersible, the Nautile, is operated by Frenchscientists and largely exploited to investigate the oceans,particularly hydrothermal vent areas.
Further improvements in technical devices and roboticvehicles have been made by scientists at the Japan MarineScience and Technology Centre (JAMSTEC). Newmanned submersibles such as SHINKAI 6500 and newremotely operated vehicles such as KAIKO (Figure 3)enabled sampling at the Mariana Trench and investigationof the deep ocean as never before. JAMSTEC developedunique facilities for the study of barophilic microorgan-isms. The Deep-Bath System (Moriya et al., 1995) iscomposed of three large units by which samples are firstcollected in closed vessels, then diluted and finallycultivated under the same conditions as those occurringat the site of collection. The cultivation unit is shown inFigure 4.
Representative Deep Sea Organisms
In the open ocean there are two self-contained ecosystems:neustonic and epipelagic. It has been reported that veryclose to the sea surface there is a flora of small flagellatesmixed with bacteria. Neustonic herbivores, such ascopepods and larvaceans, presumably graze on this film.Below the epipelagic zone, herbivorous forms – copepods,euphausiidis and so forth – may be engulfed by flying-fishes and migrating mesopelagic fishes, such as lantern-fishes. Planktonic species typical of deep waters such asGaetaenus pileatus, Pontoptilus muticus, Eucopia unguicu-lata and Eukrohnia fowleri certainly support the food webin these areas.
Below the epipelagic zone there are no primaryproducers. To exist and grow below the photosyntheticzone, plants must turn to organic food and animals oftenmigrate to surface waters during the night, resting by day.
At the level of the bathypelagic zone there is no evidenceof daily migration. The total biomass of bathypelagic life at
Figure 3 The remotely operated vehicle KAIKO operated by JAMSTEC,with which it was possible to sample the bottom of the Mariana Trench for
the first time.
Figure 4 The Deep-Bath System by which microorganisms can be
cultivated at up to 65 MPa and at temperatures over 1008C.
Deep Ocean Ecosystems
3

2000 m has been estimated to be on the order of 1/50 of theamount in the surface waters. In descending frommesopelagic to bathypelagic levels, animals become lessdiverse, much less abundant and more carnivorous. Onereason for the decreased diversity of animals withincreasing depths is a larger competition for food coupledwith lower basal metabolism.
An interesting finding is that a higher diversity of deepsea animals has been observed under warm water of theocean, with a significant fall passing poleward throughtemperate regions. The main factor behind this greaterdiversity is the production of plant food, which is more orless constant over the year in subtropical and tropicalwaters, but markedly seasonal in temperate and polar seas.
In a descent from shelf to slope, a marked increase in thediversity of the in-fauna and epibenthic fauna occurs atdepths between 100 and 300 m. Below this there is little orno change in temperature throughout the year, whereas at100 m the annual change amounts to 10.58C.
Physical conditions are sufficiently constant at any levelin the deep ocean that one should expect no barriers to thespread of animals. Some deep sea forms certainly live inmore than one ocean but, while copepods and fishes arequite cosmopolitan, few of the larger benthic invertebratesseem to be widely distributed.
In a general overview of deep ocean ecosystems it is notpossible to describe most of organisms that inhabit theseareas. However, some representative species are describedhere.
Protozoans
The most important protozoans in the ocean are thepredominantly bottom-living foraminiferans and theplanktonic radiolarians (Cavalier-Smith, 1993). The Glo-bigerina species have formed great deposits, the Globiger-ina oozes, over a third of the deep ocean floor where theyare found at depths of 2500–4500 m in large densities.Symbiotic dinoflagellates have been found in Foraminiferaand Radiolaria, and they have been phylogeneticallystudied by small-subunit rDNA analysis (Gast and Caron,1996).
Ciliates and flagellates are usually abundant in sedi-ments, but no significant observations have been reportedon their presence in the deep sea.
Sponges
Sponges are more complex than Protozoa in that they areanimals composed of many cells of more than one type,arranged roughly in layers. Water is drawn through tinypores as a result of the beating of flagella located on specialcells. The meshwork supporting the body is made ofskeletal spicules, on the basis of which the sponges areclassified.
Among the sponges inhabiting the deep ocean, mostrepresentative are the hexactinellids (Hyalospongiae) withsix-rayed silica spicules, and the Demospongiae with eitherdifferent-shaped silica spicules or the protein spongin.
Coelenterates
The coelenterates are radially symmetrical animals built oftissue-forming cells in two distinct layers, or epithelia,separated by a noncellular gelatinous layer, the mesogloea.The group is divided into Hydrozoa, Scyphozoa andAnthozoa.
Most hydrozoans, like Velella and Porpita, are floatingforms but the former has a deep-water larval stage. AmongScyphozoa most deep-water species are very large owing toexpansion of the gelatinous mesogloea. These medusae areusually luminescent and colourful, like Tesserantha con-nectens. Moreover, they can be highly poisonous, like thecommon Mediterranean cubomedusans Carybdea marsu-pialis and Carybdea rastoni (Peca et al., 1997; Nagai et al.,2000).
Among the over 6000 species of Anthozoa, many deep-water forms are known. The gorgonians, sea fans and seafeathers, have a skeleton of either calcareous spicules orprotein gorgonin. They are found at depths up to 4000 m,and are often oriented according to the water currents. Theblack corals are also deep-water species and, owing to theirslender black branch-like skeleton, they are rather similarto the gorgonians.
The phylogenetic relationships within the Anthozoahave been recently determined by nuclear 18 rDNAsequences (Berntson et al., 1999).
Crustaceans
Crustaceans may be considered the insects of the seabecause of their successful colonization of the marineenvironment.
Ostracods are almost entirely enclosed in a bivalvedcarapace or shell, they are usually very small and they canbe found in the bottom sediments even in the deep sea.Whereas pelagic species are mostly carnivores or scaven-gers, bottom species are detritus feeders. The shells ofbenthic species are often thickened and sculpted, and theseempty shells are used as geological markers in looking foroil deposits or classifying sediments.
Copepods can account for 70–90% of the biomass of azooplankton sample. They are usually quite small, buttheir size increases to reach a maximum of 17 mm at 500–2000 m depth; many of these deep-water species are alsoluminescent. Copepods supply the main food source ofmany fishes, particularly sardines and herrings, but alsodeep sea species.
Barnacles (or cirripedes) are characterized by their fixedlifestyle due to the development of a calcified protective
Deep Ocean Ecosystems
4

plate. They are the only crustaceans highly adapted to asessile life and species are found on a variety of bases, fromfuel oil to whale jaws and turtles.
Amphipods are largely represented by euphausiids,which feed on either live prey or detritus. At the surface,huge swarms of individuals (krill), such as Euphausiasuperba, are eaten by whales in the southern hemisphere.Genetic subdivision of Antarctic krill (Euphasia superbaDana) populations has been determined (Zane et al., 1998).Many deep sea species of euphausiids can reach large sizesof up to 150 mm in length.
Decapods have a carapace fused with the thoracicsegments and covering the gills. They are able to live indeep waters, but usually no species are observed deeperthan 6000 m. Benthic forms are usually larger than pelagicspecies and most are basically carnivorous. Deep-watershrimps, prawns and crabs are commonly deep red, orangeor brown, often with very complex light organs beneath thecuticle. A remarkable exception is represented by shrimpsand crabs inhabiting deep sea hydrothermal vent areas.Here huge populations of individuals belonging to highlyselected species surround the emissions of hot water andthrive on bacteria and trace minerals (Figure 5).
Molluscs
Over 31 000 species of molluscs live in the sea, but most ofthem are found along the continental shelf or shore.Among the forms living in deep waters, the ‘living fossil’Neopilina galathea, was isolated for the first time in 1952 at3570 m depth. This organism is less than 4 cm long andshows a pronounced and distinctive body segmentation.This observation led to speculation on the development ofmolluscs from segmented animals such as the annelids.
Other molluscs are commonly found on the deep bottom ofthe oceans and represent a significant part of theecosystem.
Aplacophora (worm-like molluscs), Polyplacophora(chitons) and Scaphopoda (elephant tusk shells) live atdepths ranging from 4000 to 7000 m, but these groups arenot important in deep waters. Gastropods (snails), on theother hand, are quantitatively representative of the deepsea floor biota. Prosobranchs (with gills at the front) liveparticularly at the bottom of the oceans; Janthina,Atlanta,Carinaria and Pterotrachea are some of the genera thatbelong to this group and they are characterized by a largevariability of shell size and morphology.
Lamellibranchs or pelecypods (bivalves) are the mol-luscs living at the greatest depths (over 10 000 m). In deep-adapted species likeAbra spp. the gill becomes smaller andacts to transport incoming sediment from the siphon to thepalps. Deeper forms usually have larger palps that are usedto increase the absorption of food from the oligotrophicenvironment.
Species like Calyptogena magnifica (Figure 6) that live inH2S-enriched ecosystems are of interest because of theirsymbiosis with specific psychrophilic microorganisms, andbecause of a peculiar anatomical structure. The relation-ships of these molluscs with bacterial endosymbionts aswell as their physiological traits have been extensivelyinvestigated (Distel et al., 1988; Blum and Fridovich,1984).
Within the group of molluscs, cephalopods are veryimportant organisms that live in large groups on thesurface as well as in deep waters. The ‘living fossil’Nautilusis a survivor of thousands of relatives that existed duringthe Palaeozoic era. Spirula, which lives exclusively at greatdepths, is the only living cephalopod with a coiled, internalshell. Both organisms achieve buoyancy by sucking waterout of the cavities within their shells.
The most important groups of cephalopods in the deepsea are without doubt squid and octopuses, particularly the
Figure 5 Shrimps and crabs observed around hydrothermal vent
emissions at 1100 m depth.Figure 6 Samples of Calyptogena magnifica collected at the Okinawa
Trough (Japan).
Deep Ocean Ecosystems
5

former, which can be found in large numbers. Singleindividuals have been measured up to 20 m long. Squidgenerally have ten tentacles that bear hundreds of suckers;they usually swim extremely rapidly by means of a jet ofwater ejected from a small tube under the head. Mitochon-drial DNA sequences of cephalopods have been used todetermine their molecular phylogeny (Bonnaud et al., 1997).
Echinoderms
These organisms, which are very important for the ecologyof the deep ocean, show no true segmentation, are usuallyradially symmetrical and have rows of hydrostatic tube feetand calcareous skeletons.
Feather stars (class Crinoidea) exist as either stalked orstalkless forms and have been observed at depths up to1500 m. Sea stars (Class Asteroidea), brittle stars (ClassOphiuroidea), sea urchins (Class Echinoidea) and seacucumbers (Class Holothuroidea) are common inhabi-tants of deep waters and, although they have differentfeeding behaviour, these groups contain littoral speciesthat extend into abyssal regions as well as exclusivelyabyssal species. Organisms such as Ophiomusium lymaniand Onoirophanta mutabilis live typically at depthsbetween 1000 and 5000 m, often in very large aggregations.
Asteroidea and Ophiuroidea are among the fastestorganisms living in the deep sea. Individuals ofHymenastermembranaceus, Bathybiaster vexillifer and Ophiomyxatumida, observed between 1000 and 3000 m depth, arecapable of moving at speeds from 1.7 to 6 cm min2 1 (Gage,1991).
Protochordates
The main characteristics of this group of animals are thepresence of gill clefts and a skeletal stiffening rod. They aredivided into Hemichordata (acorn worm and tube-livingforms) and primitive chordata (tunicates). Deep sea speciesare common, either as colonial aggregates or as solitaryforms; the blood of some species contains high concentra-tions of various metals, perhaps important for specificbiosyntheses.
Fish
Over 25 000 species of living fish are known and most ofthem live on the continental shelf. Among the fish that canbe typically observed in deep waters or near the bottom arethe shark Deania calcea, the macrurid Macrurus guntheri,the cod Mora mediterranea, the electric ray Torpedonobiliana, the eel Synaphobranchus pinnatus, the gulpereel Eurypharynx pelecanoides, the brotulids (Bassogigas),the chimaeras (Chimera), the rat-tails (Macrouridae), andthe angler fish Melanocoetus johnsoni. Some of theseorganisms have unique morphologies, often with large
jaws, light organs, large teeth and long appendages. Deepsea fishes typically move very slowly and behave asaggressive carnivores; so far they have not been observedat depths greater that 9000 metres.
Deep sea fish such as species belonging to the genusCoryphaenoides have been investigated by mitochondrialDNA sequences in order to determine their molecularphylogeny (Morita, 1999).
Microorganisms
The microorganisms living in the deep sea deserve morespecific description because they have been extensivelyinvestigated during recent decades either for their physio-logical properties or for their potential applications(Canganella, 1996). As a result of numerous samplingexpeditions, many new species have been described,including hyperthermophilic and barophilic strains.
ZoBell and Johnson (1949) initiated work on the effect ofhydrostatic pressure on microbial activities. The term‘barophilic’ was first used then, defined today in terms ofoptimal growth at pressure higher than 0.1 MPa or arequirement of increased pressure for growth. Manymicroorganisms in the deep sea are extremophiles such ashalophiles, thermophiles, psychrophiles, and barophiles orpiezophiles (the term ‘piezophile’ has been proposed as areplacement for barophile based on the translations of theGreek prefixes baro indicating weight and piezo indicatingpressure Yayanos (1995)). Some of these microorganismscannot survive in so-called ‘moderate’ environments.
Isolation and growth properties
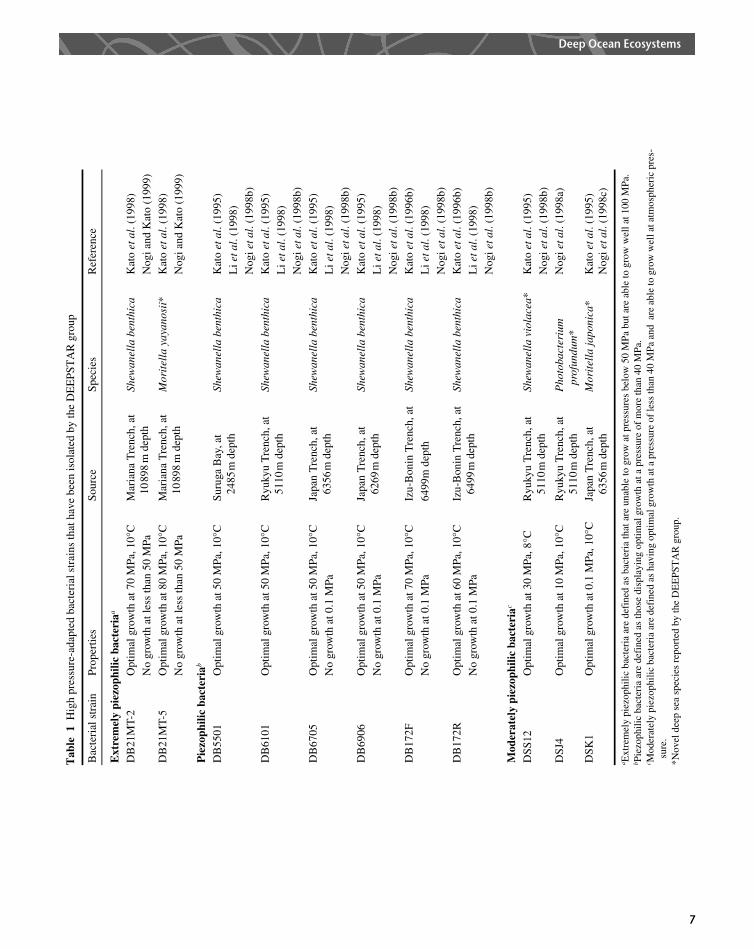
Bacteria living in the deep sea display several unusualfeatures that allow them to thrive in this extremeenvironment. The first piezophilic bacterium isolated as apure culture was reported in 1979 (Yayanos et al., 1979).The spirillum-like bacterial strain CNPT-3 had a rapiddoubling rate at 50 MPa, but did not grow into colonies forseveral weeks under atmospheric pressure. This bacteriumdid not lose any of its piezophilic characteristics after atleast ten transfers, but they were retained between transfersif pressure was kept at 58 MPa. Numerous piezophilic andpiezotolerant bacteria have been isolated and character-ized by the DEEPSTAR group at JAMSTEC; theorganisms were collected from deep sea sediments atdepths ranging from 2500 m to 11 000 m by sterilized mudsamplers (Kato et al., 1995, 1996a, 1996b, 1998). Some ofthese strains are listed in Table1. Most of the isolated strainsare not only piezophilic or piezotolerant but alsopsychrophilic, and cannot be cultured at temperaturesabove 208C.
Taxonomy
Many deep sea piezophilic bacteria have been shown tobelong to the gamma-Proteobacteria group through
Deep Ocean Ecosystems
6
Tab
le1
Hig
h pr
essu
re-a
dapt
ed b
acte
rial
str
ains
that
hav
e be
en is
olat
ed b
y th
e D
EE
PST
AR
gro
up
a Ext
rem
ely
piez
ophi
lic b
acte
ria
are
defi
ned
as b
acte
ria
that
are
una
ble
to g
row
at p
ress
ures
bel
ow 5
0 M
Pa b
ut a
re a
ble
to g
row
wel
l at 1
00 M
Pa.
b Pie
zoph
ilic
bact
eria
are
def
ined
as
thos
e di
spla
ying
opt
imal
gro
wth
at a
pre
ssur
e of
mor
e th
an 4
0 M
Pa.
c Mod
erat
ely
piez
ophi
lic b
acte
ria
are
defi
ned
as h
avin
g op
timal
gro
wth
at a
pre
ssur
e of
less
than
40
MPa
and
are
abl
e to
gro
w w
ell a
t atm
osph
eric
pre
s-su
re.
*Nov
el d
eep
sea
spec
ies
repo
rted
by
the
DE
EPS
TA
R g
roup
.
Bac
teri
al s
trai
nP
rope
rtie
sSo
urce
Spec
ies
Ref
eren
ce
Ext
rem
ely
piez
ophi
lic b
acte
riaa
DB
21M
T-2
Opt
imal
gro
wth
at 7
0 M
Pa, 1
0°C
Mar
iana
Tre
nch,
at
1089
8m
dep
thSh
ewan
ella
ben
thic
aK
ato
et a
l. (1
998)
No
grow
th a
t les
s th
an 5
0 M
PaN
ogi a
nd K
ato
(199
9)
DB
21M
T-5
Opt
imal
gro
wth
at 8
0 M
Pa, 1
0°C
Mar
iana
Tre
nch,
at
1089
8m
dep
thM
orit
ella
yay
anos
ii*
Kat
o et
al.
(199
8)
No
grow
th a
t les
s th
an 5
0 M
PaN
ogi a
nd K
ato
(199
9)
Pie
zoph
ilic
bact
eria
b
DB
5501
Opt
imal
gro
wth
at 5
0 M
Pa, 1
0°C
Suru
ga B
ay, a
t24
85m
dep
thSh
ewan
ella
ben
thic
aK
ato
et a
l. (1
995)
Li e
t al.
(199
8)
Nog
i et a
l. (1
998b
)
DB
6101
Opt
imal
gro
wth
at 5
0 M
Pa, 1
0°C
Ryu
kyu
Tre
nch,
at
5110
m d
epth
Shew
anel
la b
enth
ica
Kat
o et
al.
(199
5)
Li e
t al.
(199
8)
Nog
i et a
l. (1
998b
)
DB
6705
Opt
imal
gro
wth
at 5
0 M
Pa, 1
0°C
Japa
n T
renc
h, a
t63
56m
dep
thSh
ewan
ella
ben
thic
aK
ato
et a
l. (1
995)
No
grow
th a
t 0.1
MPa
Li e
t al.
(199
8)
Nog
i et a
l. (1
998b
)
DB
6906
Opt
imal
gro
wth
at 5
0 M
Pa, 1
0°C
Japa
n T
renc
h, a
t62
69m
dep
thSh
ewan
ella
ben
thic
aK
ato
et a
l. (1
995)
No
grow
th a
t 0.1
MPa
Li e
t al.
(199
8)
Nog
i et a
l. (1
998b
)
DB
172F
Opt
imal
gro
wth
at 7
0 M
Pa, 1
0°C
Izu-
Bon
in T
renc
h, a
tSh
ewan
ella
ben
thic
aK
ato
et a
l. (1
996b
)
No
grow
th a
t 0.1
MPa
6499
m d
epth
Li e
t al.
(199
8)
Nog
i et a
l. (1
998b
)
DB
172R
Opt
imal
gro
wth
at 6
0 M
Pa, 1
0°C
Izu-
Bon
in T
renc
h, a
t 64
99m
dep
thSh
ewan
ella
ben
thic
aK
ato
et a
l. (1
996b
)
No
grow
th a
t 0.1
MPa
Li e
t al.
(199
8)
Nog
i et a
l. (1
998b
)
Mod
erat
ely
piez
ophi
lic b
acte
riac
DSS
12O
ptim
al g
row
th a
t 30
MPa
, 8°C
Ryu
kyu
Tre
nch,
at
5110
m d
epth
Shew
anel
la v
iola
cea*
Kat
o et
al.
(199
5)
Nog
i et a
l. (1
998b
)
DSJ
4O
ptim
al g
row
th a
t 10
MPa
, 10°
CR
yuky
u T
renc
h, a
t 51
10m
dep
thP
hoto
bact
eriu
mpr
ofun
dum
*N
ogi e
t al.
(199
8a)
DSK
1O
ptim
al g
row
th a
t 0.1
MPa
, 10°
CJa
pan
Tre
nch,
at
6356
m d
epth
Mor
itel
la ja
poni
ca*
Kat
o et
al.
(199
5)N
ogi e
t al.
(199
8c)
Deep Ocean Ecosystems
7

comparison of 5S and 16S rDNA sequences. As a result of ataxonomic study based on its 5S rDNA sequences, theobligate piezophilic bacterium Colwellia hadaliensis wasassigned to the Proteobacteria gamma-subgroup (Deminget al., 1988).
DeLong et al. (1997) reported that 11 cultivatedpsychrophilic and piezophilic deep sea bacteria wereaffiliated with one of the genera within the gamma-subgroup: Shewanella, Photobacterium, Colwellia, Mor-itella, and an unidentified genus. The only deep seapiezophilic species among these genera were reported tobeS. benthica in the genusShewanella (Deming et al., 1984;MacDonell and Colwell, 1985) and C. hadaliensis in thegenus Colwellia (Deming et al., 1988) prior to the reportsfrom the DEEPSTAR group. Since then, four newpiezophilic species within those genera have been describedbased on chromosomal DNA–DNA hybridization studiesand several other taxonomic properties.
New species of bacteria have been identified amongpiezophilic bacterial isolates.Photobacteriumprofundum, anew species, was identified and described as the moderatelypiezophilic strains DSJ4 and SS9 (Nogi et al., 1998a). P.profundum strain SS9 has been studied extensively withregard to the molecular mechanisms for pressure regula-tion (Bartlett et al., 1989, 1996; Welch and Bartlett, 1998).
The moderately piezophilic strain DSS12 isolated fromthe Ryukyu Trench at a depth of 5110 m was identified asShewanella violacea (Nogi et al., 1998b), a novel piezophilicspecies belonging to the Shewanella piezophile branch (Liet al., 1998).
Strain DSK1, which is a moderately piezophilic bacter-ium isolated from the Japan Trench, was identified asMoritella japonica. This is the first piezophilic speciesidentified in the genus Moritella (Nogi et al., 1998c).The type strain of the genus Moritella is M. marina,previously known asVibriomarinus (Urakawa et al., 1998),which is one of the most common psychrophilic organismsisolated from marine environments (Colwell andMorita, 1964).
Present and Future Perspectives
The deep sea is a truly dark world where sunlight neverpenetrates; and it is a cold world at 2–48C. Because of this,people have long imagined that the deep sea is a dead worldwith no living organisms. However, as described above,scientists have found that a number of organisms exist inthe deep sea; even in the Mariana Trench, the deepest seatrench on earth, many microorganisms have been isolated.The deep sea is a vital world where many living organismsare active and highly adapted.
Deep sea research has historically addressed two mainissues: (1) understanding such an extreme environment,observing new creatures and phenomena; and (2) investi-
gating the presence of valuable resources that may beexploited in different ways. To achieve the first aim, oneshould first investigate the diversity and the novelty of deepsea organisms. Then it will be necessary to elucidate themechanisms of adaptation to such environments charac-terized by high hydrostatic pressure, low (or sometimeshigh) temperatures, darkness, oligotrophy, etc. After thesesteps it may be reasonable to search for useful organismssuitable for potential industrial applications and offeringnew opportunities for improvement of the quality of life.
There are various questions and/or expectations aboutliving organisms in the deep-sea: How can they adapt tosuch high hydrostatic pressure? Are there any specialorganisms unique to environments with such extremeconditions? Are such special organisms useful to us? Thesequestions and expectations are still unanswered andremain issues for future research.
The exploration and study of the ocean floor duringrecent decades has allowed geophysicists to reveal theexpansion of the ocean floor and to quantify its growthrates. Physico-chemical conditions in deep sea environ-ments have also been studied, particularly the relationshipsbetween sedimentary basins, fluids, gas hydrates andvolcanic phenomena.
Supercritical water, under extreme conditions of highpressure and temperature, is one of the future researchtargets. The solution behaviour of colloid particles andbiomaterials in supercritical water will be investigated asmodels of phenomena occurring above hydrothermalvents and volcanoes.
Other future developments in deep sea research includefurther exploration of oceanic ridges and hydrothermalecosystems; understanding of sedimentary processes andthe flux of organic matter in the deep ocean; and thedevelopment of abyssal observatories.
In terms of microbiology and adaptation to hydrostaticpressure, research will be mainly focused on three topics:isolation of high pressure-loving (piezophilic) microorgan-isms and molecular biological analysis of high-pressuretolerants; research on high-pressure adaptation (piezo-physiology); and investigation of deep sea subsurfacemicroorganisms.
Not much was known about the physiology of piezo-philic bacteria until recently. Now scientists are interestedin retrieving microorganisms from the Earth’s crust, and itis expected that this work will provide valuable insightsconcerning the origin of life and its development.
Bio-informatic studies of the entire genome sequencewill provide valuable information on microbial evolutionand the unique gene set of bacteria. This research is likely tolead to new industrial applications and to methods forthe production of useful enzymes. Research on genomebiology should be expanded to allow new techniques tobe applied to the most interesting deep sea micro-organisms.
Deep Ocean Ecosystems
8

Interesting prospects for the exploitation of the deepocean are the Ocean Drilling Project sponsored by theUnited States and by European countries; the mining ofhydrothermal vent sites that contain significant amounts ofprecious metals; and the development of novel deep seavehicles, towed bathymetric imaging systems and geotech-nical tools.
The results of this research will make a great contribu-tion not only to basic science but also to the development ofindustrial and medical applications, environmental con-servation and other fields. These will include the fields offood processing and chemical production, medicine andbioactive materials production, cleaning of the oceans andof the air, and others. In biomedicine, for instance, crudeextracts of the alga Caulerpa taxifolia have been tested forantiviral activities and in vitro exhibited an inhibitory effectagainst an immunodeficiency virus (Nicoletti et al., 1999).
Frontier research of this kind is drawing attention to anew world of microorganisms and will lead to an expansionof the search for extremophilic organisms, popularly called‘superbugs’, in the Earth’s crust (geobiology) andperhaps even extraterrestrial superbugs beyond the Earth(exobiology).
References
Bartlett DH, Chi E and Welch TJ (1996) High pressure sensing and
adaptation in the deep-sea bacterium Photobacterium species strain
SS9. In: Hayashi R and Balny C (eds) High Pressure Bioscience and
Biotechnology, pp. 29–36. Amsterdam: Elsevier Science.
Bartlett D, Wright M, Yayanos AA and Silverman M (1989) Isolation of
a gene regulated by hydrostatic pressure in a deep-sea bacterium.
Nature 342: 572–574.
Berntson EA, France SC and Mullineaux LS (1999) Phylogenetic
relationships within the class Anthozoa (phylum Cnidaria) based on
nuclear 18S rDNA sequences. Molecular Phylogenetic Evolution 13:
417–433.
Blum J and Fridovich I (1984) Enzymatic defences against oxygen
toxicity in the hydrothermal vent animals Riftia pachyptila and
Calyptogena magnifica. Archives of Biochemistry and Biophysics 228:
617–620.
Bonnaud L, Boucher-Rodoni R and Monnerot M (1997) Phylogeny of
cephalopods inferred from mitochondrial DNA sequences. Molecular
Phylogenetic Evolution 7: 44–54.
Canganella F (1996) Recent insights into the ecology of deep-sea
microorganisms and their potential applications. Annali di Micro-
biologia ed Enzimologia 46: 65–77.
Cavalier-Smith T (1993) Kingdom protozoa and its 18 phyla. Micro-
biological Reviews 57: 953–994.
Colwell RR and Morita RY (1964) Reisolation and emendation of
description of Vibrio marinus (Russell) Ford. Journal of Bacteriology
88: 831–837.
DeLong EF, Franks DG and Yayanos AA (1997) Evolutionary
relationship of cultivated psychrophilic and barophilic deep-sea
bacteria. Applied and Environmental Microbiology 63: 2105–2108.
Deming JW, Hada H, Colwell RR, Luehrsen KR and Fox GE (1984)
The ribonucleotide sequence of 5S rRNA from two strains of deep-sea
barophilic bacteria. Journal of General Microbiology 130: 1911–1920.
Deming JW, Somers LK, Straube WL, Swartz DG and Macdonell MT
(1988) Isolation of an obligately barophilic bacterium and description
of a new genus, Colwellia gen. nov. Systematic and Applied
Microbiology 10: 152–160.
Distel DL, Lane DJ, Olsen GJ et al. (1988) Sulfur-oxidizing bacterial
endosymbionts: analysis of phylogeny and specificity by 16S rRNA
sequences. Journal of Bacteriology 170: 2506–2510.
Gage JD (1991) Biological rates in the deep-sea: a perspective from
studies on processes in the benthic boundary level. Reviews on Aquatic
Sciences 5: 49–64.
Gast RJ and Caron DA (1996) Molecular phylogeny of symbiotic
dinoflagellates from planktonic foraminifera and radiolaria. Mole-
cular Biology and Evolution 13: 1192–1197.
Herring PJ and Clarke MR (eds) (1971) Deep Oceans. London: Barker.
Kato C, Sato T and Horikoshi K (1995) Isolation and properties of
barophilic and barotolerant bacteria from deep-sea mud samples.
Biodiversity and Conservation 4: 1–9.
Kato C, Inoue A and Horikoshi K (1996a) Isolating and characterizing
deep-sea marine microorganisms. Trends in Biotechnology 14: 6–12.
Kato C, Masui N and Horikoshi K (1996b) Properties of obligately
barophilic bacteria isolated from a sample of deep-sea sediment from
the Izu-Bonin Trench. Journal of Marine Biotechnology 4: 96–99.
Kato C, Li L, Nakamura Y, Nogi Y, Tamaoka J and Horikoshi K (1998)
Extremely barophilic bacteria isolated from the Mariana Trench,
Challenger Deep, at a depth of 11,000 meters. Applied and Environ-
mental Microbiology 64: 1510–1513.
Li L, Kato C, Nogi Y and Horikoshi K (1998) Distribution of the
pressure-regulated operons in deep-sea bacteria. FEMS Microbiolo-
gical Letters 159: 159–166.
MacDonell MT and Colwell RR (1985) Phylogeny of the Vibrionaceae,
and recommendation for two new genera, Listonella and Shewanella.
Systematic and Applied Microbiology 6: 171–182.
Morita T (1999) Molecular phylogenetic relationships of the deep-sea
fish genus Coryphaenoides (Gadiformes: Macrouridae) based on
mitochondrial DNA. Molecular Phylogenetic Evolution 13: 447–454.
Moriya KT, Inada M, Kyo T and Horikoshi K (1995) Large-scale
fermentation under high hydrostatic pressure using a newly developed
Deep-sea Baro/Thermophile Collection and Cultivation System.
Journal of Marine Biotechnology 2: 175–177.
Nagai H, Takuwa K, Nakao M et al. (2000) Novel proteinaceous toxins
from the box jellyfish (sea wasp) Carybdea rastoni. Biochemistry and
Biophysics Research 275: 582–588.
Nicoletti E, Della Pieta F, Calderone V, Bandecchi P, Pistello M and
Morelli C (1999) Antiviral properties of a crude extract from a green
alga Caulerpa (Vahl) C. Agardh. Phytotherapy Research 13:
245–247.
Nogi Y, Masui N and Kato C (1998a) Photobacterium profundum sp.
nov., a new, moderately barophilic bacterial species isolated from a
deep-sea sediment. Extremophiles 2: 1–7.
Nogi Y, Kato C and Horikoshi K (1998b) Taxonomic studies of deep-sea
barophilic Shewanella species, and Shewanella violacea sp. nov., a new
barophilic bacterial species. Archives of Microbiology 170: 331–338.
Nogi Y, Kato C and Horikoshi K (1998c) Moritella japonica sp. nov., a
novel barophilic bacterium isolated from a Japan Trench sediment.
Journal of General and Applied Microbiology 44: 289–295.
Peca G, Rafanelli S, Galassi G et al. (1997) Contact reactions to the
jellyfish Carybdea marsupialis: observation of 40 cases. Contact
Dermatitis 36: 124–126.
Urakawa H, Kita-Tsukamoto K, Steven SE, Ohwada K and Colwell RR
(1998) A proposal to transfer Vibrio marinus (Russell 1891) to a new
genus Moritella gen. nov. as Moritella marina comb. nov. FEMS
Microbiological Letters 165: 373–378.
Welch TJ and Bartlett DH (1998) Identification of a regulatory protein
required for pressure-responsive gene expression in the deep-sea
Deep Ocean Ecosystems
9

bacteriumPhotobacterium species strain SS9.MolecularMicrobiology
27: 977–985.
Yayanos AA (1995) Microbiology to 10,500 meters in the deep sea.
Annual Review of Microbiology 49: 777–805.
Yayanos AA, Dietz AS and Boxtel RV (1979) Isolation of a deep-sea
barophilic bacterium and some of its growth characteristics. Science
205: 808–810.
Zane L, Ostellari L, Maccatrozzo L, Bargelloni L, Battaglia B and
Patarnello T (1998) Molecular evidence for genetic subdivision of
Antarctic krill (Euphausia superba Dana) populations. Proceedings of
the Royal Society of London and British Biological Sciences 265:
2387–2391.
ZoBell CE and Johnson FH (1949) The influence of hydrostatic pressure
on the growth and viability of terrestrial and marine bacteria. Journal
of Bacteriology 57: 179–189.
Further Reading
Earle S and Giddings A (1980)Exploring theDeep Frontier: TheAdventure
of Man in the Sea. Washington, DC: National Geographic Society.
Ernst WG and Morin JG (1982) The Environment of the Deep-Sea.
Englewood Cliffs, NJ: Prentice-Hall.
Kennish MJ (1994) Practical Handbook ofMarine Science. Boca Raton,
FL: CRC Press.
Prager EJ and Earle SA (2000) The Oceans. New York: McGraw-Hill.
Rowe GT and Pariente V (1992) Deep-sea Food Chains and the Global
Carbon Cycle. Dordrecht: Kluwer Academic.
Deep Ocean Ecosystems
10





























