Embed Size (px)
Citation preview

ENERGY TRANSFORMATION IN PHOTOSYNTHESIS AND THE RELATION OF PHOTOSYNTHESIS TO RESPIRATION
BY C. P. WHITTINGHAM Botany School, Cambridge
(Received 7 January 1954)
CONTENTS
I. Introduction . . . . . . . . . . . . . 11. The absorption of radiant energy and its transformation into chemical energy .
111. The chloroplast reaction . . . . . . . . . . . IV. Known types of energy transfer . . . . . . . . . V. Possible role of high-energy phosphate compounds . . . . . .
VI. Interdependence of respiration and photosynthesis . . . . . . ( I ) Respiration during a steady state of photosynthesis . . . . (2) Respiration during a non-steady state of photosynthesis . . . .
VII. The fixation m d reduction of carbon dioxide . . . . . . . VIII. Summary . . . . . . . . . . . . .
IX. References . . . . . . . . . . . . . X. Addendum . . . . . . . . . . . . .
PAGE
' 40 ' + I
' 46
' 47 ' 48
' 50
' 54 . j8 . 60 . 62 . 64
. j0
I . INTRODUCTION
The essential metabolic activities of the green plant, photosynthesis and respiration, differ in at least one major feature : the former is confined to those parts of the plant which contain the pigment chlorophyll, whereas the latter is a process common to all parts of the plant and indeed to most living organisms. Thus until recently photosynthesis was regarded as a problem peculiar to the study of plant physiology, whereas respiration was most intensively studied in animals and in micro-organisms such as yeast, and the conclusions reached applied by the plant physiologist in his interpretation of the less detailed studies of higher plant metabolism.
Owing in part to the greater variety of cell types and the consequent greater degree of localization of metabolic activities in animal tissues, the isolation and purification of the enzymes concerned in respiration has met with considerable success. This has made possible the study of the partial processes or reactions catalysed by the isolated enzymes in vitro and led to an understanding as to the manner in which the individual reactions are linked together in vivo. I n this way the relationship between oxidative processes and the formation of phosphorylated esters was demonstrated. So far no enzymes known to be specifically concerned with photosynthesis have been isolated. The simplest unit with which it has been possible to demonstrate photochemical activity, which may be related to photo- synthesis, is the fragmented chloroplast, probably at a level of organization com- parable to that of the mitochondria. Thus, inevitably, models of photosynthesis

Photosynthesis and respiration 41 have been expressed primarily in kinetic terms, in contrast to those of respiration which have been mainly in terms of enzymes.
The study of the kinetics of photosynthesis, that is, of the relationship between the rate of oxygen production or of carbon dioxide consumption and different external factors such as concentration of carbon dioxide or light intensity, led to the formulation of a mechanism of photosynthesis consisting of a chain of reactions, some photochemical and some thermochemical. In addition to carbon dioxide, water and chlorophyll, other compounds were postulated as intermediates in the reaction, and these were defined solely in terms of their kinetic characteristics.
Suggestions as to the nature of both the photochemical and the dark reactions involved in photosynthesis have come from studies in comparative biochemistry. As a result of work with photosynthetic bacteria, van Niel(1941) suggested that the photochemical reaction in photosynthesis may be interpreted as an oxidation- reduction reaction photosensitized by chlorophyll, in which light energy produces from water a hydrogen donor able to reduce carbon dioxide. Some experimental support for this view was obtained when it was shown that chloroplasts isolated from the plant were able to sensitize oxidation-reduction reactions between water and a number of hydrogen acceptors. According to this view, the dark reactions of photosynthesis are considered to be similar to those involved in respiration. The unique feature of photosynthesis is the ability to convert energy absorbed by the photosynthetic pigments into a form with which the plant is able to reverse, at least in part, the respiratory reactions, thus resulting in synthesis of compounds at the level of reduction of carbohydrate. The use of radioactive carbon dioxide has indeed shown that during illumination many of the compounds labelled in the green plant are those already known to be concerned in respiration. The primary problem in photosynthesis then becomes the elucidation of the photochemical processes, which may be considered to terminate with the formation of a hydrogen donor.
11. THE ABSORPTION OF RADIANT ENERGY AND ITS TRANSFORMA- TION INTO CHEMICAL ENERGY
In addition to the two primary excited states which are formed when the chlorophyll molecule absorbs blue or red light and which are short lived, there is considerable evidence for the existence of a metastable (or ‘triplet’) state which is longer lived. The photochemical mechanism by means of which the energy of excitation is transferred to this metastable state and then subsequently to a stable chemical form is still unknown. However, it is probable that among all the photosynthetic pig- ments chlorophyll a has a unique relationship to the process of energy transforma- tion. The fluorescence spectrum of a cell suspension, when it is illuminated by light of a given wave-length, may be compared with the fluorescence spectrum of the isolated pigments responsible for the absorption of light at this wave-length. The excitation of the pigment in the cell may result only in fluorescence characteristic of that pigment, but if energy is transferred from the absorbing pigment to a second

42 C. P. WHITTINGHAM pigment, the latter may also contribute to the fluorescent light. Experiments show that such transfer of energy between photosynthetic pigments does occur in the cell. For example, in a suspension of diatoms illuminated with light which is pre- dominantly absorbed by the carotenoid fucoxanthol, the fluorescence is similar in quality and intensity to that emitted when the exciting light is absorbed only by chlorophyll. Hence energy has been transferred in the cell from the fucoxanthol, which absorbed the light, to chlorophyll. Similarly, energy transfer has been shown from chlorophyll b to chlorophyll a in Chlorella (Duysens, 1952); from the caro- tenoid fucoxanthol to chlorophyll b in diatoms (Dutton, Manning & Duggar, 1943 ; Wassink & Kersten, 1946); from phycocyanin to chlorophyll a in the blue-green alga Oscillatoria (Duysens, I~SZ), and from phycoerythrin through phycocyanin to chlorophyll a in the red alga Porphyridium (Duysens, 1952). Chlorophyll a is the only pigment common to all classes of photosynthetic plant, with the exception of the bacteria in which bacterio-chlorophyll probably plays a similar role. Excitation of any of the photosynthetic pigments ultimately results in excitation of chlorophyll a.
Prior to studies of fluorescence, the same conclusion had been reached as a result of measurements of the quantum efficiency of photosynthesis for light absorbed in different regions of the spectrum. In addition, these observations provide measure- ments of the efficiency with which light is transferred. Thus, it had ben shown that the photosynthetic efficiency of light absorbed by carotenoids in Chlorella was about half that of light absorbed by chlorophyll (Emerson & Lewis, 1943); light absorbed by fucoxanthol in the diatoms was about as efficient as that absorbed by chlorophyll (Dutton & Manning, 1943; Tanada, 1951); and light absorbed by phycocyanin in the blue-green alga Chroococcus about as efficient as that absorbed by chlorophyll (Emerson & Lewis, 1942). In the red algae it was found that the efficiency of light absorbed by phycoerythrin was greater than that for light absorbed by chlorophyll (Haxo & Blinks, 1950); this was later also found to be true for the excitation of fluorescence (French & Young, 1952). This has been attributed to the existence in these plants of a form of chlorophyll, chlorophyll d, which is inactive both with respect to fluorescence and to photosynthesis. The experimental observations suggest that phycocyanin and phycoerythrin transfer excitation energy preferentially to chlorophyll a.
The nature of the reaction in which excited chlorophyll a subsequently par- ticipates is not known. Information might be obtained from two types of studies: (I) the study of spectra, and (2) the study of quantum efficiency. No change in absorption spectrum of the plant has been observed during or consequent upon photosynthesis. Thus we are left with the second possibility. The quantum efficiency depends both on the nature of the photochemical reactions and the subsequent dark reactions. The photosynthetic bacteria differ from the green plant in the necessity for some external hydrogen donor other than water which becomes oxidized during photosynthesis. Comparison of the photosynthetic efficiency in these organisms with that in the green plant will indicate whether the photochemical mechanism in these two groups is the same.

Photosynthesis and respiration 43 Larsen, Yocum & van Niel (1952) made investigations with green sulphur
bacteria capable of three types of photosynthesis, the reduction of carbon dioxide with concomitant oxidation of hydrogen, of thiosulphate, or of tetrathionate. As shown in the following equations, the free energy of reduction per molecule of carbon dioxide varies widely. Nevertheless, it was found that the quantum requirement was in all cases the same, namely, about ~ o q u a n t a per molecule of carbon dioxide, a value in agreement with that found by Wassink, Katz & Dorrestein (1942) for the reduction of carbon dioxide by hydrogen in the purple sulphur bacteria :
C0,+2H,-(CH20) +H,O+ 1.7 kcal., (1)
CO, + & S,O,” + 8 H,O 2 (CH,O) + SO4” + H+ + 29’2 kcal., (2)
CO2++ S4O6’’+Y H,O-(CH,O)++ SO,‘’+? H++30*6 kcal. (3) If these efficiencies represent the attainment of a minimum quantum requirement, it may be argued that all three processes involve a common primary reaction.
There has been considerable discussion as to the minimum quantum requirement for photosynthesis in the green plant under steady-state conditions. In the steady state there is good evidence that in many plants the main product has an average level of reduction equivalent to that of carbohydrate. Thus, the energy requirement in the standard state is of the order of 115,000 cal. per mole of carbon dioxide reduced; it will differ only very little from this for the concentrations existing in the cell :
If we consider the energy available from the activated state of chlorophyll to be approximately the same as that of the maximum for the red absorption band (660 mp, 43,000 cal.), the minimum quantum requirement is 2.7. Franck (1953) has given thermodynamic reasons why a quantum requirement of less than 5 is improbable.
Recent measurements of photosynthetic efficiency in the steady state in the green plant have been largely confined to suspensions of the unicellular alga Chlorella. Whether the suspension used has absorbed only a fraction of the incident light or almost all of it, most observers have obtained quantum requirements of 8-10 quanta per molecule of oxygen produced. (These results have been reviewed (Whittingham, 1952 a).) However, Warburg and his colleagues have repeatedly claimed efficiencies approaching 3 quanta per molecule of oxygen. Warburg, Geleick & Briese (195 I a) attribute this difference to the use by all other workers of too low concentrations of carbon dioxide. They claim that maximum efficiencies can only be observed in the presence of concentrations of carbon dioxide as high as 5 % . This is in marked contrast to the ‘saturation ’ of photosynthesis at high intensities of illumination with very much lower concentrations of carbon dioxide, when that for half maximal rate is of the order of 0*002+3*02% (Warburg, 1919; Whittingham, 1952~). From kinetic principles it follows that at low light intensities, such as used for the deter- mination of efficiency, a yet smaller concentration should be required for ‘satura-
CO, + 2H20 - & C,H,,O, + H20 + 0, + I I 5,000 cal. (4)

44 C. P. WHITTINGHAM tion’. Emerson & Whittingham (unpublished) in independent work have been unable to repeat the observations of Warburg et al. ; they find in the steady state no effect of carbon dioxide concentration (in the range o.z-5-0%) on rate of photo- synthesis either at high or low light intensities. In later work Warburg et aZ. (1953) used ‘new’ cultures which apparently did not show the same effect of concentration of carbon dioxide as the old cells. A further paper elucidating these facts is promised by Warburg.
The difference between the values for photosynthetic efficiency obtained by different workers was at first assigned to errors of technique. There is, however, a more fundamental difficulty than merely that of minimizing errors of measurement. With low light intensities the estimated photosynthesis is dependent on the value assumed for respiration in the light. Brackett, Olson & Crickard (1953) observed with ChZoreZZa variations in rate of respiration during periods of alternate light and dark such as have been used in determinations of efficiency. However, they claimed that provided the value for respiration is interpolated from dark periods between the light periods there is no dependence of photosynthetic efficiency on intensity of illumination as previously claimed by Kok (1948). The effect of carbon dioxide concentration on efficiency claimed by Warburg et aZ. has been interpreted by Steemann Nielsen (1953) as due to an effect on respiration. In his experiments in which the concentration of carbon dioxide was varied between 0-02 and 5yo, no effect of concentration on rate of photosynthesis was observed by Steemann Nielsen at high light intensities, except for an inhibitory effect at the highest concentrations (between I and 5 % ) . At lower light intensities there was little effect on rate of photosynthesis for concentrations between 0.05 and 0.5 yo but increase from 0.5 to 5 yo resulted in a 5 0 % increase in rate. The rates of oxygen exchange were deter- mined using the Winkler method necessitating the measurement of an average rate of photosynthesis during a period of one to many hours. Hence no evidence could be obtained as to possible variation of rate with time. On the basis of this experiment Steemann Nielsen suggested that high concentrations of carbon dioxide inhibit respiration in the light; in the dark no effect of concentration of carbon dioxide on respiration occurs. Thus if the rate of respiration in the light is assumed to be the same as in the dark the photosynthesis is calculated to be greater than it actually is; this error will have the greatest effect at relatively low light intensities when the rate of photosynthesis is of the same order as the rate of respiration.
The most recent experiments of Warburg (1953), using very thin suspensions, demonstrated, duringperiods of illumination lasting several hours, a quantum require- ment for photosynthesis of + I quantalmolecule oxygen even when no correction was made for respiration. The respiration in the steady dark state was about one-twentieth the observed rate of photosynthesis. Correction for respiration gave a value for the quantum requirement of 3.88. The cells used had a very high maximum rate of photosynthesis (60 volumes of oxygen per volume of cell per hour), and it is stated that quantum requirements of 4 can be observed with these cells when photo- synthesizing at a rate two-thirds that of their maximum. It thus appears improbable

Photosynthesis and respiration 45 that the difference between the low and high determinations for quantum require- ment can be attributed solely to a difference in rate of respiration in light and dark. A fully satisfactory explanation as to the causes of the difference between the determinations of photosynthetic efficiency by different workers has not yet been given.
Because the quantum requirement for photosynthesis is of the same order in the green plant and the bacteria, van Niel(1941) has suggested that the photochemical reaction in photosynthesis is the same in these organisms. Under most conditions this reaction is characterized by a quantum requirement for the over-all reduction of carbon dioxide of 10 quanta per molecule. The common photochemical step in all photosyntheses is suggested to be one in which activated chlorophyll (or bacterio- chlorophyll) directly reacts with some molecule (A) which becomes thereby poten- tially capable of accepting hydrogen from water; the water is oxidized to oxygen in the green plant or oxidizes an externally supplied hydrogen donor (e.g. hydrogen sulphide) in the bacteria, the reduced product, AH,, subsequently reducing carbon dioxide, probably in a combined form:
Common primary photochemical reaction: ( 5 ) Common reduction of carbon dioxide: (6) Reaction of oxidized radical: green plant 4(OH)-zH,O + O,,
4H,O + zA -2AH2 + 4(OH); CO, + zAH,-(CH,O) + H,O + zA
1 (7) bacteria 4(OH) + zH,S-4H2O + 2s. The reactions subsequent to the photochemical step were presumed to occur with the loss of free energy ; the energy requirement for the whole sequence is determined by that of the first step alone. Calvin & Barltrop (1952) have recently made the suggestion that the substance represented by A in these equations may be a com- pound containing S-S linkages which become reduced to the SH form during the photochemical reaction.
According to this mechanism, hydrogen transfer reactions play an important part in photosynthesis. The transfer of hydrogen to substance A might be formulated either involving transfer of two hydrogen atoms at a time as represented in equation (8) or involving transfer of one hydrogen atom at a time as in (9):
OT
These equations assume that each of the four hydrogen atoms necessary for the reduction of a molecule of carbon dioxide to carbohydrate originates in an identical manner. For the reduction of carbon dioxide under standard conditions this would require that substance B in equation (9) should have an oxidation-reduction poten- tial lower than that of the standard hydrogen electrode ( -0q4V. at pH7.0; 28,750 cal. free energy per H transfer).

46 C. P. WHITTINGHAM
III. THE CHLOROPLAST REACTION
Chloroplasts isolated from the living plant are able to catalyse the reduction of a variety of substances in light, e.g. ferric oxalate, ferricyanide, quinone. Amongst naturally occurring compounds, cytochrome and coenzymes I and 2 (DPN+; TPN+) (Hill & Scarisbrick, 1940; Warburg & Luttgens, 1946; Vishniac & Ochoa, 1952) can be shown to react with the chloroplast system only by indirect methods. The substances of greatest reducing power, TPN+ and DPN+, are reduced to only a small extent even when oxygen is almost completely removed from the system. The standard oxidation-reduction potential of the coenzymes is -0.30 V. (pH 7.0); according to the mechanism just considered in order to achieve reduction of carbon dioxide under realizable concentrations and in the presence of oxygen at atmospheric pressures, it is necessary to postulate a substance B with a standard potential not less than -0.44 V. Thus, if the chloroplast reaction is considered to be the same as the photochemical reaction occurring in photosynthesis, it is apparent that when isolated the system does not reduce substances to an extent which the simple mechanism of photosynthesis so far considered would lead one to expect.
It is, therefore, of importance to question whether the chloroplast reaction is analogous rather than identical with the photochemical reaction in photosynthesis in the living plant. At low light intensities, using either ferric oxalate (French & Rabideau, 1945), quinone, ferric oxalate plus ferricyanide (Ehrmantraut & Rabino- witch, 1952) or methaemoglobin (Hill, Davenport & Whittingham, unpublished) as hydrogen acceptors, the quantum requirement is about 10-12 quanta per molecule of oxygen, that is, of the same order as for photosynthesis. However, it appears that at higher light intensities the rate of the chloroplast reaction increases less rapidly with increase in light intensity than does photosynthesis (Clendenning & Ehrman- traut, 1950; also Hill, Davenport & Whittingham, unpublished); nevertheless, the maximal rates appear to be of the same order. One explanation for these two facts taken together has been given by Tolmach (1951) in terms of Frank’s general theory of photosynthesis. It is suggested that the products of the primary photo- chemical reaction break down in back reactions unless stabilized by a catalyst; it is postulated that the products formed in the chloroplast reaction are less rapidly stabilized than are those involved in photosynthesis.
The chloroplast system may not reduce TPN+ directly; the latter may merely represent an ultimate stable hydrogen acceptor. Tolmach (195 I) showed that TPN+ would act catalytically with an unwashed chloroplast preparation under conditions of low partial pressures of oxygen ; the nature of the ultimate hydrogen acceptor in this preparation was not ascertained. Washing the chloroplasts destroyed the effect possibly by removal of some natural hydrogen acceptor.
The simple mechanism described by equations (s), (6) and (7) may need modifi- cation. For example, a number of hydrogen transfers may occur before a reductant is produced which can reduce carbon dioxide. These may result by a process of ‘energetic dismutation’ in the formation of a hydrogen donor of a more reducing

Photosynthesis and respiration 47 potential, the necessary energy being derived from the oxidation of some of the product of the initial reaction. For example, BH, the product of reaction (9), the primary photochemical step, may reduce a substance X which then in turn reduces carbon dioxide. If this be the case it is not necessary that TPN+/TPNH should itself be a carrier in the sequence of hydrogen transfers.
n--2 n-2 nBH + ~ [OJ + X 2 XH, + nB + ~ H,O. 4 2
Such coupled reactions have been frequently considered with reference to mechanisms of respiration.
IV. KNOWN TYPES OF MECHANISM OF ENERGY TRANSFER Before continuing the discussion of coupled reactions in photosynthesis, we shall consider mechanisms of reaction coupling which have become clear from studies of respiration. The essential feature of energetic coupling or linkage is the presence of an intermediate common to two systems which may be considered separately as representing one, a system of negative free-energy change coupled with another of smaller positive free-energy change. The known mechanisms of coupling are of two types, oxidative-reductive coupling and oxido-phosphorylative coupling. An example of the former is the dehydrogenation of two substances, both of which, in the presence of their respective enzymes, transfer hydrogen to the same acceptor. Thus, the oxidation of glucose-6-phosphate (G6P) to 6-phosphogluconic acid in the presence of glucose-6-phosphate dehydrogenase results in the reduction of coenzyme 2 (TPN+); the reduced TPNH so formed can be utilized in the presence of ‘malic
i (11) glucose-6-phosphate + TPN++6 phosphogluconic + TPNH + H+, pyruvic acid + TPNH + CO, + H+,---malic acid + TPN+, M n + +
enzyme’ and manganous ions to fix carbon dioxide in malic acid. (Vishniac& Ochoa, 195-2). With low pressures of carbon dioxide the equilibrium of the second step is far over to the side of decarboxylation; a position of apparent equilibrium, favouring the formation of malic acid, can be attained only if a large fraction of the coenzyme is maintained in the reduced form. This can be achieved by continuously supplying glucose-6-phosphate. The apparent equilibrium can be shifted still further in the direction of carboxylation by addition of fumarase which catalyses the removal by dehydration of some of the malate formed in the reaction.
Oxido-phosphorylative coupling occurs in the linkage of the oxidation of phospho- glyceraldehyde in the presence of triose phosphosphate dehydrogenase and TPN+ with the formation of adenosine triphosphate (ATP) from adenosine diphosphate (ADP) and free phosphate. The standard free-energy change of the over-all reaction is small, showing that the energy of oxidation is largely conserved in the phosphate ester, each ‘energy-rich’ ester storing of the order of 10,000 cal. CH,OH,PO,* CHOH * CHO + H3P04 + DPN+ +ADP+
CH20H2P03.CHOH*COOH+ATP+DPNH+H+. (12) * [O,] represents oxygen either free or in a combined form.

48 C. P. WHITTINGHAM Subsequently hydrolysis of the ‘energy-rich’ ester is coupled to an ‘energy- requiring’ reaction, thus making possible the transfer of energy from the oxidation of phosphoglyceraldehyde to mechanical work in the muscle or to synthetic reactions such as the formation of starch in the plant.
V. POSSIBLE ROLE OF HIGH-ENERGY PHOSPHATE COMPOUNDS
We now consider whether mechanisms of energy transfer of the phosphorylative type may not play a role in photosynthesis.
As we have discussed, chloroplast suspensions illuminated in the presence of TPN+ but in the almost complete absence of oxygen will reduce TPN+ at least to a small extent. This reaction could then be coupled to an oxidation-reduction reaction in which TPNf is oxidized and carbon dioxide reduced. Thus in the example of coupled oxidation-reduction represented by equation (I I), light energy via chloroplasts could replace the energy otherwise supplied from the oxidation of G6P. Ochoa and others, using illuminated chloroplasts, ‘malic enzyme’ and TPN+, have demonstrated carboxylation of pyruvic acid by coupling with the chloroplast reaction (Vishniac & Ochoa, 1952; Tolmach, 1951; Arnon, 1951).
It is, however, most improbable that in the cell, the ratio of TPNH/TPN+ could be maintained sufficiently high so that the reduction of carbon dioxide to carbo- hydrate could be appreciably effected in a coupled reaction even supposing the concentration of product maintained low. For example, if the ratio TPNH/TPN+ were maintained at IOOO/I the ratio of CO,/CH,OH in equilibrium with this would be IO,OOO/I. If, however, we combine with an oxidative-reductive coupling a phosphorylative oxidation, then the energy requirement for the reduction of carbon dioxide can be obtained, with the possibility of accumulating reduced products. By combining the standard energy of hydrolysis of a high-energy phosphate ester (10,000 cal.) with that for the reduction of TPN+ (25,400 cal.), we have sufficient energy per hydrogen atom for the reduction of carbon dioxide to carbohydrate (28,800 cal.). Alternatively, we may postulate an oxidative-reductive coupling as represented by equation (10).
The theoretical possibility that phosphorylation is involved in photosynthesis has been frequently suggested (Ruben, 1943; Umbreit, 1947; Kandler, 1950; Rabinowitch, I ~ S I ) , but experimental evidence in support of this view has been lacking until recently. Emerson, Stauffer & Umbreit (1944) investigated the changes in inorganic and organic phosphates during photosynthesis in Chlorella, but could find only small effects. Wassink, Tjia tz Wintermans (1949) demonstrated small changes in inorganic phosphate during photosynthesis in the purple sulphur bacteria Chromatium ; also it was shown that illumination of Chlorella was accom- panied by an increase in phosphate insoluble in trichloracetic acid, this effect being most pronounced in absence of carbon dioxide (Wassink, Wintermans & Tjia, 1951). In the latter case, however, the effect of carbon dioxide could be alternatively obtained by addition of glucose.
Great interest was stimulated by the findings of Vogler & Umbreit (1942) with

Photosynthesis and respiration 49 the chemosynthetic bacterium Thiobacillus thiooxidans. During a period of oxida- tion of sulphur in the absence of carbon dioxide it was found that inorganic phos- phate was taken up by the cells; then, subsequently, under anaerobic conditions in the presence of carbon dioxide but in the absence of sulphur, carbon dioxide was taken up and inorganic phosphate liberated into the medium. Considerable doubt was raised as to whether the carbon dioxide taken up was in fact reduced to carbo- hydrate, for had this been the case it would have required an estimated maximal thermodynamic efficiency of about 90 % (Rabinowitch, 1945). Further, as Baalsrud & Baalsrud (1952) point out, even if only one molecule of ATP were required for the fixation of each molecule of carbon dioxide, it would be necessary to suppose that all the nitrogen in the cell was present as ATP. Baalsrud & Baalsrud repeated experiments of the type reported by Vogler & Umbreit and were able to show that in presence of a hydrogen donor, carbon dioxide was consumed and converted to a compound as reduced as carbohydrate. Evidence for this was obtained by deter- mining the difference in oxygen consumption when the bacteria were oxidizing thiosulphate in the presence and absence of carbon dioxide; this difference was equal to the amount of carbon dioxide assimilated. These workers, unlike Vogler & Umbreit, were unable to find any uptake of carbon dioxide in the absence of thiosulphate subsequent to a period of oxidation of thiosulphate in the absence of carbon dioxide. Small changes in inorganic phosphate were found in the presence of added thiosulphate, but these were only slightly more in the absence of carbon dioxide than in its presence. Thus the recent work of Baalsrud & Baalsrud has made the significance of the earlier work of Vogler & Umbreit doubtful.
Using a new method of analysis for ATP, dependent on the stimulation of luminescence in a firefly extract, Strehler (1953) has been able to demonstrate changes in ATP content of Chlorella during illumination using an extract of the cells in boiling water. After a period of anaerobiosis, addition of oxygen resulted in an increase in ATP concentration rising to a steady value in about I min. ; illumina- tion resulted in a rise in concentration to a maximum value, attained in about I min., followed by a subsequent fall to a steady value within 2-3 min. The steady-state level of ATP was greater at lower light intensities whether in the presence or absence of carbon dioxide. At 25' C. the steady level was higher after addition of oxygen than after illumination; at 4' C. the reverse was true. It was suggested that at low light intensities the rate of production of ATP exceeded the rate of consump- tion, whereas at high light intensities the rate of consumption dominates owing to synthetic reactions which do not appear to be entirely dependent on the presence of carbon dioxide. A ratio of I mole ATP per 6 moles oxygen produced was observed. Kandler (1950) previously had demonstrated changes in orthophosphate in Chlo- rella, and these are consistent with the changes in phosphate ester found by Strehler.
Changes in ATP, or indeed in general distribution of phosphate, consequent upon illumination in the green plant cell do not necessarily imply that phosphoryla- tion results directly from the photochemical reaction of photosynthesis. Lehninger
4 Biol. Rev. 30

50 C. P. WHITTINGHAM (195 I), using rat-liver particles, has demonstrated that the oxidation of DPNH may result in phosphorylations, and Vishniac & Ochoa (1952) have shown, by mixing illuminated chloroplasts with mung bean mitochondria in the presence of added DPN and ADP, that illumination in chloroplasts can result in phosphorylation. Phosphorylation was dependent on the presence of oxygen and is presumably associated with the oxidation of some reduced product of the chloroplast reaction. In the living cell this product might be an intermediate less reduced than carbo- hydrate or it might be carbohydrate itself.
Such a mechanism would make photosynthesis dependent on a concomitant oxidative process (not necessarily involving free oxygen) which resulted in phos- phorylation. The over-all efficiency of photosynthesis would be related to the efficiency of the phosphorylative reaction. With cyclophorase preparations from rabbit kidney a production of three phosphate ester molecules per atom of oxygen has been demonstrated; hence the complete oxidation of one molecule of carbo- hydrate to carbon dioxide and water could supply 36 phosphate ester molecules. Supposing the transfer of each of the four atoms of hydrogen for the reduction of a molecule of carbon dioxide to carbohydrate to require one molecule of T P N H and one of ATP, then the over-all efficiency of photosynthesis would approach that of the photochemical step. If, however, the phosphorylative process were relatively inefficient the efficiency of photosynthesis would be much lower than that of the photochemical process.
VI. INTERDEPENDENCE OF RESPIRATION AND PHOTOSYNTHESIS
The rate of respiration of the cell must be limited either by the electron transporting system or by the phosphorylative system coupled to the oxidative reactions. (A discussion of this subject has been given in a recent review by Turner (I~sI).) If in the green plant light results in the production of reduced substances and possibly of phosphorylated esters, it might be expected that the rate of respiration would be different in the light from that in the dark. The direction of change will depend on the balance between the rate of production and the rate of consumption of both hydrogen donor and phosphorylated esters. Hence studies of the effect of light on respiration are of significance for the development of mechanisms of photosynthesis.
( I ) Respiration during a steady state of photosynthesis Measurements of the rate of respiration in the dark immediately before and after
illumination have tended to suggest that apart from general drifts in rate with time there is little effect of light on respiration. Recently, Brown (1953) has made use of mass isotopes for determining the rate of gas exchange due to respiration during illumination. In these experiments tracer oxygen (lsO1sO) was added to the gas phase in a known molar ratio to non-tracer oxygen (laO1eO). With oxygen tracer there is little exchange with the water or Chlorella cells present. Thus the uptake of tracer is proportional to the consumption of oxygen in respiration. Any oxygen
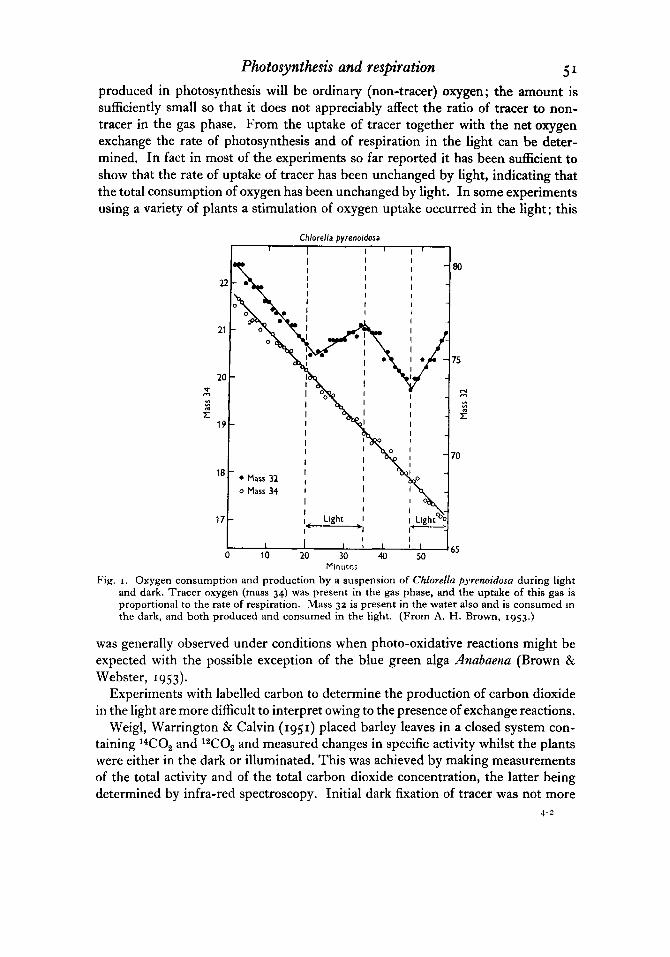
Photosynthesis and respiration 5 1
produced in photosynthesis will be ordinary (non-tracer) oxygen; the amount is sufficiently small so that it does not appreciably affect the ratio of tracer to non- tracer in the gas phase. From the uptake of tracer together with the net oxygen exchange the rate of photosynthesis and of respiration in the light can be deter- mined. In fact in most of the experiments so far reported it has been sufficient to show that the rate of uptake of tracer has been unchanged by light, indicating that the total consumption of oxygen has been unchanged by light. In some experiments using a variety of plants a stimulation of oxygen uptake occurred in the light; this
Fig. I . Oxygen consumption and production by a suspension of Chlorella pyrenoidom during light and dark. Tracer oxygen (mass 34) was present in the gas phase, and the uptake of this gas is proportional to the rate of respiration. Mass 32 is present in the water also and is consumed in the dark, and both produced and consumed in the light. (From A. H. Brown, 1953.)
was generally observed under conditions when photo-oxidative reactions might be expected with the possible exception of the blue green alga Anabaena (Brown & Webster, 1953).
Experiments with labelled carbon to determine the production of carbon dioxide in the light are more difficult to interpret owing to the presence of exchange reactions.
Weigl, Warrington & Calvin (1951) placed barley leaves in a closed system con- taining 14C02 and 12C02 and measured changes in specific activity whilst the plants were either in the dark or illuminated. This was achieved by making measurements of the total activity and of the total carbon dioxide concentration, the latter being determined by infra-red spectroscopy. Initial dark fixation of tracer was not more

52 C. P. WHITTINGHAM than 5 yo of the total uptake in the light, so that changes in the light were largely due to photochemical reactions. In the light the specific activity rose initially for about 10 min. ; this was attributed to isotope discrimination in photosynthesis. After this time the concentration of carbon dioxide was reduced almost to zero and there was practically no net photosynthesis; a rapid fall to a final steady value for specific activity occurred. The light was then turned off and the specific activity rose attaining a steady value in 5 min. whose magnitude was inversely proportional to the total preceding light period. This rise subsequent to the attainment of a constant specific activity in the light can be interpreted if we assume that in the light the respiration of assimilatory products containing 14C is less than in the dark. The authors proposed that in the light the main source of carbon dioxide production was from endogenous reserves; only a small amount of carbon dioxide came from recently assimilated carbon. In the dark the recently formed assimilates are oxidized in a constant proportion to the endogenous reserves.
Van Norman & Brown (1952) investigated the gas exchange of Chlorella in a closed system containing 12C02, 13C02 and 14C02. A recording mass spectrometer was used to determine separately each species of carbon dioxide molecule. Their results were similar to those of Weigl et al., with the exception that in the primary phase in the light the ratio of I4C/l2C in the gas phase does not rise but decreases slowly; this is presumably related to the fact that in the published experiment there is a consumption of 14C02 in the dark prior to illumination which continued un- changed up to the time when illumination commenced. In the experiments of Weigl et al. the rate of incorporation of 14C02 in the dark prior to illumination was small.
Steeman Nielsen (1953) reported measurements of the rate of production of 14C0, in the presence of 12C02 for Chlorella which had previously photosynthesized in the presence of 14C02 and thus contained labelled carbon compounds. The rate of production of 14C02 was considerably less in the light than in the dark. This was interpreted as due to an inhibition by light of the respiration of substances recently formed in photosynthesis; but in addition to possible exchange reactions, which may be initiated by light, there is also in these experiments the possibility of reassimila- tion in the light of carbon dioxide produced in respiration.
The difficulty in interpreting experiments of this type is that three different processes are occurring, photosynthesis, respiration and exchange. The two latter may proceed at a different rate in the light and dark. Further, there is the effect of mass discrimination in each of these processes. Weigl et al. assumed the respiratory production of carbon dioxide in the light to be one-half that in the dark; this as- sumption led to a factor for discrimination in the light for 14C02/13C02 of 0.83. On the other hand, van Norman & Brown assumed that the rate of carbon dioxide production was unchanged in the light; nevertheless, the resulting factor calculated for discrimination was almost the same, namely, 0.86 for 14C/12C. For 13C/12C they derived a value of 0.99. It appears that no precise estimate as to the effect of light on the rate of production of carbon dioxide can be deduced from these data; only

Photosynthesis and respiration 53 when the discrimination factor has been established from other evidence, and more information obtained about possible photo-induced exchange reactions, can it be hoped that work with carbon isotopes will provide unequivocal data as to the effect of light on respiration.
Benson & Calvin (1950) used a different approach and compared the distribution of 14C in Chlorella cells which had been illuminated in presence of l*CO, and were then placed in the absence of carbon dioxide in helium (absence of oxygen) either in light or dark. Glutamic and isocitric acids were labelled only when a period of dark followed the period of photosynthesis. This was interpreted as showing that the Krebs cycle was operative in the dark but was suppressed in the light. The published chromatograms do not show an accumulation of phosphopyruvic or of malic acid in the light- as compared to the dark-treated cells; this might have been expected if the Krebs cycle were not functioning and pyruvic acid (or near deriva- tives of it) were accumulating. Alanine (the amino-acid corresponding to pyruvic acid) appears to be labelled to a greater extent in the dark than in the light.
Calvin & Massini (1952) have proposed that in the light the oxidative decarboxy- lation of pyruvic acid is inhibited, thus suppressing the Krebs cycle. They suggest that this is due to the reduction in the light of lipoic acid, a co-factor for the forma- tion of active acetyl groups from pyruvic acid; the co-factor is known to be effective only in the oxidized form. Gibbs (1953) has found a change in distribution of label as between alanine and malic acid at high and low light intensities, and regards this as support for the hypothesis that the Krebs cycle is suppressed in light.
If the only effect of light on the respiratory system was the reduction of lipoic acid, there would result an inhibition of the oxidative decarboxylation of pyruvic acid and a tendency for phosphoglyceric acid to rise above its dark concentration level. Phosphoglyceric acid may also arise in another way in the light, namely, by carboxylation of a two-carbon acceptor. The resultant increase in concentration of PGA would tend to cause a decrease in concentration of ATP which would be dependent on the presence of carbon dioxide. Since Strehler has shown that whilst the steady-state level of ATP decreases in the light whether carbon dioxide is present or not, yet there is an initial rise at the onset of illumination, this probably indicates that ATP is utilized in the light and also produced as a result of a light- initiated process.
Owing to the interreaction of three different processes in the light, which has been discussed previously, it is difficult to obtain reliable measurements of the rate of production of carbon dioxide together with the consumption of oxygen due to respiration in the light. The possibility exists that in the light hydrogen transfer occurs directly or indirectly from the product of the photochemical step to oxygen, probably accompanied by concomitant phosphorylation. Such a process would compete with other oxygen-consuming processes, such as the oxidative decarboxy- lations of the Krebs cycle. If, as the experiments of Brown indicate, the over-all rate of oxygen consumption is unaltered by light, this constancy may be not simply fortuitous but rather that the over-all rate of oxygen uptake is limited by some other

54 C. P. W H I T T I N G H A M controlling factor such as a terminal oxidase or a control of some of the hydrogen transferring reactions by the ratio of ATP/ADP. Little is known about respiratory mechanisms in Chlorella. Holzer & Holzer (1952) have demonstrated that an acetone powder of Chlorella is able to catalyse the formation of phosphoglyceric acid and triose phosphate from hexose diphosphate. Detailed discussion of possible respiratory intermediates which might react with products of the photochemical primary reaction must await further elucidation of respiratory pathways in Chlorella.
According to these suggestions, addition of an alternative oxidative pathway, as, for example, the addition of nitrate to the photosynthesizing cell, might permit the photochemical step to proceed more rapidly. With high light intensities Warburg & Negelein (1920) demonstrated an increase in rate of production of oxygen when nitrate was added to illuminated Chlorella. If in these experiments nitrate is sub- stituting for oxygen as hydrogen acceptor for the reduced product of the photo- chemical step, the increased rate of production of oxygen will not be accompanied by a corresponding increase in the rate of reduction of carbon dioxide. This rate will remain the same at high light intensities but will be decreased at low light intensities. This has been shown to be the case in the experiments of Cramer & Myers (1948) and those of van Niel, Allen & Wright (1953), in which the presence of nitrate at low light intensities resulted in a decrease in consumption of carbon dioxide as would be expected according to the suggested mechanism. Evans & Nason (1953) have purified from a number of higher plants an enzyme which catalyses the reduc- tion of nitrate to nitrite, utilizing TPNH or DPNH as hydrogen donors. Illuminated grana isolated from soya-bean leaves can catalyse reduction of nitrate to nitrite in the presence of T P N ; no further reduction of nitrite was observed in agreement with the observations of Kessler (1953) using chloroplasts isolated from spinach. If the photochemical reaction of photosynthesis produces T P N H in vivo, then in the presence of nitrate and the nitrate-reducing enzyme the T P N H would be re- oxidized without effecting any reduction of carbon compounds. As discussed earlier, chloroplasts can catalyse the reduction of coenzymes I and 2 to only a limited extent even under anaerobic conditions; it is therefore of interest that Evans & Nason found reduction of nitrate by grana from soya-bean leaves in the presence of added T P N even under aerobic conditions. Other substances in addi- tion to TPNH may be able to act as hydrogen donor to nitrate.
.
( 2 ) Respiration during a non-steady state of photosynthesis In some recent investigations rapid methods of determining gas exchange have
been used to measure respiration during the first minute of dark following short periods of illumination. Immediately following the cessation of a period of illumina- tion of I min. or shorter a much enhanced uptake of oxygen has been observed by Burk & Warburg (1951) using a manometric method. Results obtained with the manometric method when used to determine rapidly changing rates during non- steady conditions may be quantitatively and possibly qualitatively misleading.
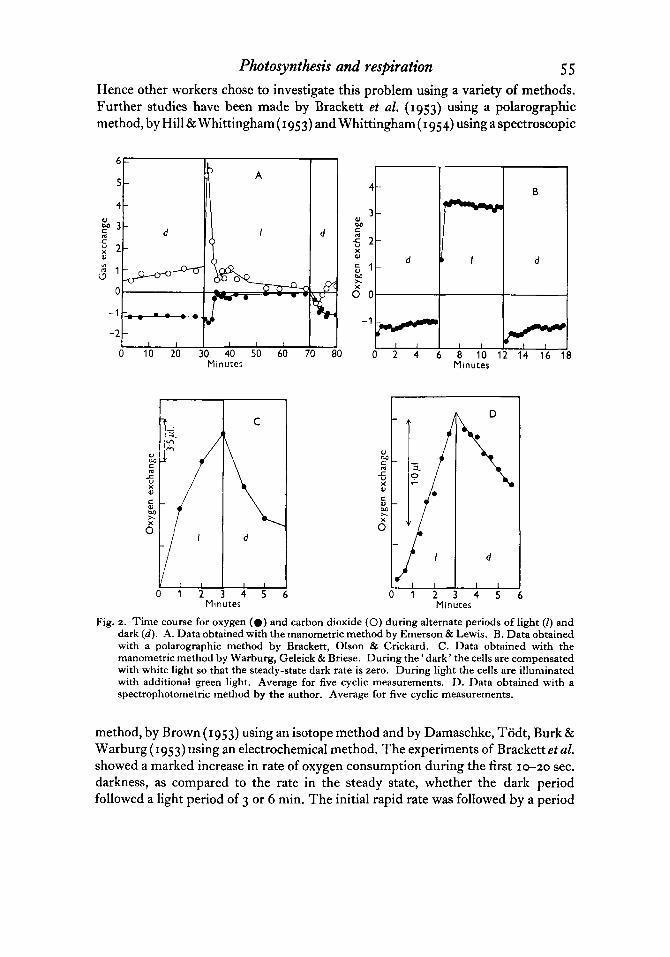
Photosynthesis and respiration 5 5 Hence other workers chose to investigate this problem using a variety of methods. Further studies have been made by Brackett et al. (1953) using a polarographic method, by Hill & Whittingham( 1953) and Whittingham( 1954) using a spectroscopic
d
z h
I
4 5l
- 2 c I- I I 1 I I I I 1 1 0 10 20 30 40 50 60 70 80
Minutes
0 uo C m r
X
C
M x
6
B 4-
3 - M
-g 2 -
5 0
X d B I d
E 1 - M x
I I I 1
0 2 4 6 8 1 0 1 2 1 4 1 6 ' Mi n u tes
I
Fig. 2. Time course for oxygen (0 ) and carbon dioxide (0) during alternate periods of light (I) and dark (d). A. Data obtained with the manometric method by Emerson & Lewis. B. Data obtained with a polarographic method by Brackett, Olson & Crickard. C. Data obtained with the manometric method by Warburg, Geleick & Briese. During the ' dark' the cells are compensated with white light so that the steady-state dark rate is zero. During light the cells are illuminated with additional green light. Average for five cyclic measurements. D. Data obtained with a spectrophotometric method by the author. Average for five cyclic measurements.
method, by Brown (1953) using an isotope method and by Damaschke, Todt, Burk & Warburg (1953) using an electrochemical method. The experiments of Brackett etal. showed a marked increase in rate of oxygen consumption during the first 10-20 sec. darkness, as compared to the rate in the steady state, whether the dark period followed a light period of 3 or 6 min. The initial rapid rate was followed by a period

56 C. P. W H I T T I N G H A M of decreasing rate, a subsequent increase to a new maximum in about I & min. and then a further period of decreasing rate. These authors distinguish the initial phase from the subsequent longer term changes in rate. They showed that the existence of a higher rate during the first few seconds after illumination was unaffected by addition of glucose, and therefore suggested that the immediate increase is not due to an increase in respirable glucose. Chlorella which has been kept for a long time in the dark prior to illumination invariably shows an increased rate of oxygen con- sumption in the following dark period. Such an effect was reported by Emerson & Lewis (1943) and is generally attributed to the formation of respiratory substrate in the light; in these experiments the stimulated rate was found to persist for many minutes. The experiments of Hill et al. and the majority of those of Damaschke et al. were confined to Observations in the presence of an oxygen concentration of between 0.1 and 3.0%. Whittingham (1954) found that after periods of one minute illumination the initial rate of oxygen consumption was sometimes greater than the subsequent steady dark rate; when present the effect could be repeated for a number of cycles of light and dark. The half-time of decay was of the order of 2 min. Subsequent experiments by the author using a recording spectrophotometer confirmed these observations and showed that anomalous rates during transition periods occur only under conditions considered to be abnormal. Damaschke et al. report that they found the period of enhanced oxygen consumption to vary from the order of seconds to that of minutes. Indeed, in their Fig. 2 the enhanced dark rate (which is initially about four times the steady rate) persists for minutes (half-time about 4 min.), and it would be difficult to distinguish this effect from that of the type reported by Emerson & Lewis. On the other hand, Fig. 3 shows an experiment in which after a period of 20 sec. illumination there is a stimulated respiration which disappears within I min. of darkness. No distinction is made in the interpretation of these two experiments in the paper of Damaschke et al. However, it appears reasonable to distinguish between at least two possibilities. First, a stimulation in respiration immediately after illumination owing to an increase in respirable substrate; such an effect would be expected to vary with the pre-treatment of the cells. Secondly, an increase in rate, probably for a much shorter period owing to the persistence after the light is turned off of an intermediate which is rapidly oxidized.
The above observations, made in the presence of relatively high concentrations of carbon doixide, did not, with the exception of those of Burk & Warburg, include measurements of carbon dioxide. A marked stimulation in carbon dioxide produc- tion is observed in the dark immediately after a period of saturating illumination at very low concentrations of carbon dioxide (Weigl et al. 1951; van Norman & Brown, 1952; Whittingham (unpublished)). The half-time is of the order of 2 min. Under conditions of low concentration of carbon dioxide photo-oxidative reactions may complicate the gas exchange, Using high concentrations of carbon dioxide ( 5 % ) in air, Emerson & Lewis (1941) showed with Chlorella that during the first minutes of illumination there was a production or ‘burst’ of carbon dioxide; in the

Photosynthesis and respiration 57 first minutes of dark a consumption or ‘gulp’ was observed. In these experiments the oxygen exchange showed normal inductive effects.
Calvin & Massini (1952) have shown that during the first 2 min. immediately after illumination the amount of labelled phosphoglyceric acid rapidly increases to a value about twice that in the steady state; subsequently it falls back to a low value. They also observed during this period a marked temporary depletion of labelled sucrose. Strehler (1953) has shown that during the same period there is an increase in amount of ATP. These results would be consistent with an accelerated rate of oxidation during the first few seconds of darkness, as compared to the steady dark rate.
Burk & Warburg also observed manometrically the gas exchange of Chlorella during the light period of alternate periods of light and dark of I min. in duration. During the light they find an increased rate of oxygen production in excess of that characteristic of the steady state corresponding to the production of one molecule of oxygen per quantum absorbed. Subsequently this result was confirmed using an electrochemical method (Damaschke et al. 1953). My own experiments with the spectrophotometric method show that only in very few experiments has an increased rate of oxygen production been observed during brief periods of illumination of I min. alternated with equal dark times; even in these cases the increase was not significant. Frequently no effect has been observed on the rate of oxygen produc- tion in the light, and often, though less frequently, there has been no stimulation of the reactions either in the light or dark periods. When these effects wereabsent phenomena described earlier by Hill & Whittingham (1953) as characteristic of an ‘aerobic induction’ phase have been observed. My observations have been made with approximately 5 % carbon dioxide; Burk & Warburg have used a range of concentrations up to 50%. Gaffron (unpublished), determining changes in pH as indicative of changes in concentration of carbon dioxide, has found with Chlorella occasional changes in transient rates both in light and dark using low concentra- tions of carbon dioxide. The transient effects are apparently associated with abnormal treatment of the cells. Burk & Warburg claim that their manometric results show that changes in carbon dioxide exchange occur equal and opposite to those in oxygen. Since the determinations of assimilatory quotient are subject to the same errors as are the rate determinations under non-steady state conditions this point must also await confirmation by a method specific for carbon dioxide.
Measurements in the steady state of photosynthesis fail to indicate any effect of light on oxygen-consuming processes in the green plant cell. Some evidence indicates that during transitory periods marked changes in rate of oxygen consump- tion occur. Burk & Warburg believe that their observations together with the finding of a one-quantum process during brief periods of light alternated with brief dark periods indicate a chemosynthetic mechanism for photosynthesis. Coupled to the primary photochemical reaction, dark reactions occur which supply the further energy necessary for the reduction of carbon dioxide; these reactions can only be observed during transitory periods but continue throughout the steady state. These

58 C . P. W H I T T I N G H A M authors suggest that during the first moments of illumination an intermediate is built up which initiates an oxidative process; the intermediate reacts with carbon dioxide to liberate one molecule of oxygen per quantum absorbed; it is reformed in a respiratory process in which oxygen is consumed. The energy of the oxidation is assumed to be coupled with the reaction in which light energy is utilized. Such a mechanism implies that throughout the steady state of photosynthesis an accelerated oxygen consumption is present but is masked by the greater production of oxygen in the photochemical process. If this is the case it should be possible to observe it using an isotope method of the kind reported by Brown (1953), provided it be granted that the conditions of the two experiments are comparable. One possibility which might under certain conditions mask such an effect is that the oxygen consumed in the oxidative process is preferentially derived from that produced in photosynthesis. Brown did attempt to use a technique of measurement such that any reutilization of oxygen produced photosynthetically was reduced to a minimum. If, on the other hand, we accept the transient effects observed when rapid methods of measurements are used as only transitory phenomena, the results are themselves consistent with the observations of Brown using the isotope method and with earlier circumstantial evidence that during the steady state respiration is not appreciably changed by light.
One might expect, a priori, an interaction between the photochemical hydrogen- donating reacting system and the respiratory system. Physiological studies of the type discussed in this section are one means of analysis for any suchinteraction. Other methods include the analysis of respiratory intermediates in light and dark, as initiated in the work of Strehler; by the use of substances such as 2, 4-dinitro- phenol which inhibit the formation of phosphorylated esters; and by the study of the ‘Pasteur effect’ in ChZoreZZa as a means of determining the controlling effect of the ATP/ADP ratio on the rate of oxygen consumption. How far structural features may be of importance in determining the relationship between respiration and photosynthesis in the living cell will only become apparent when further experimental results have been obtained.
VII. THE FIXATION AND REDUCTION OF CARBON DIOXIDE The preceding sections have been concerned with the nature of the product of the photochemical reaction sequence in photosynthesis. The hypothesis has been examined that this sequence represents a chain of hydrogen transfers resulting with isolated chloroplasts and possibly in the living cell in a reductant as reducing as TPNH. A further problem arises in the discussion of the photosynthetic mechanism, namely, to determine the nature and method of formation of the substance which terminates this chain of hydrogen transfers and forms an intermediate between carbon dioxide and carbohydrate.
This problem has been intensively investigated using radioactive carbon dioxide. This work will not be considered in detail here. It has been recently reviewed by Brown & Frenkel (1953). The first compound in which tracer carbon can be

Photosynthesis and respiration 59 detected during photosynthesis is phosphoglyceric acid (PGA). Labelling is initially confined to the carboxyl group of this compound; later it appears in both the other carbon positions, suggesting the regeneration of PGA by a cyclic process from assimilated carbon. There is no good evidence as to how the carbon dioxide is fixed in PGA ; the evidence, however, clearly shows that it does not enter initially into any of the acids of the Krebs cycle to an appreciable extent.
An alternative approach to the mechanism of carbon dioxide fixation in photo- synthesis may be made via the study of kinetics of the living cell. Measurements of the rate of photosynthesis of Chlorella at different concentrations of carbon dioxide using alkaline bicarbonate carbonate buffer solutions showed that whilst at high concentrations of carbon dioxide the rate of photosynthesis was independent of time of illumination, at low concentrations (0.001 yo) the rate of photosynthesis increased some 500% during the first 2 hr. of illumination. The kinetics of this change in rate were interpreted in terms of a mechanism involving two carboxyla- tion reactions, only one of which produces a photosynthetic reactant (Briggs & Whittingham, 1952). The existence of a further substance was postulated which competes with carbon dioxide for combination with each of the acceptors. Its concentration in the cell was shown to be dependent on the concentration of carbon dioxide during the period of growth immediately prior to measurement. Attempts at specific identification of these hypothetical compounds have not yet been made. In early publications of Calvin and his colleagues (Badin & Calvin, 1950) malic acid was considered to be the product of a second carboxylation reaction; further experi- ments did not substantiate this hypothesis (Bassham, Benson & Calvin, 1950) and the present interpretation of work with tracer carbon indicates the existence of only one primary carboxylation. A second carboxylation could exist only if the product were assumed to be present in such a small amount that labelling in this compound were small compared with that in PGA.
The relationship between rate of photosynthesis and concentration of carbon dioxide has been usually interpreted in terms of the formation of a complex between carbon dioxide and an acceptor. When allowance is made for the unusually long induction phase which occurs at low concentrations of carbon dioxide and the rate of photosynthesis in the steady state alone is considered, half the maximum rate is attained in Chlorella with a concentration of carbon dioxide of 0.9 x 10-6 M (25' C.). This value is the same whether measurements are made in acid or alkaline solution and is thus independent of the presence of bicarbonate ions (Whittingham, 1952~). Assuming reversibility, the free energy of formation of the carboxyl group in vivo will be about - 8000 to - 9000 cal. Bassham et al. (1954) have suggested that PGA is formed by reaction between ribulose diphosphate (RDP) and carbon dioxide; they calculate the free energy of this reaction to be -7000 cal. The continued formation of RDP, presumably from ribulose monophosphate, will require a con- tinued supply of ATP, making carboxylation dependent on phosphorylation.
Inhibition of photosynthesis in Chlorella by cyanide is greater the lower the concentration of carbon dioxide (Whittingham, 1952b). Experiments with tracer

60 C. P. WHITTINGHAM carbon in which the uptake of tracer immediately after illumination was determined showed that this uptake was inhibited by addition of cyanide (Gaffron, Fager & Rosenberg, I 95 I). These results suggest that cyanide predominantly inhibits photo- synthesis by affecting the reaction concerning carbon dioxide. Two possibilities exist: either cyanide inhibits the reactions directly concerned in the reduction of carbon dioxide or it inhibits some linked reaction which is energetically coupled with the fixation of carbon dioxide. It is not improbable that this energy-yielding reaction involves an oxidation and that cyanide-a compound whose inhibitory action is frequently associated with oxidase activity-predominantly affects carboxylation by inhibiting a coupled oxidative activity. This may be one reaction in photosynthesis involving phosphorylations, for which evidence has been discussed earlier in this article.
The photochemical reactions of isolated chloroplasts show no induction pheno- mena, the production of oxygen invariably commencing at its full rate; chloroplasts also lack the ability to reduce carbon dioxide. It may be inferred that induction phenomena are related to the utilization of carbon dioxide as the ultimate hydrogen acceptor in photosynthesis. Unfortunately, information is lacking concerning the induction phenomena for both oxygen and carbon dioxide simultaneously. The experiments of Burk & Warburg, based on a manometric measurement of rapidly changing rates, cannot be regarded as reliable. Measurements of the time course for uptake of carbon dioxide during the induction phase of photosynthesis, using a method specific for carbon dioxide and of rapid response, have not been made simultaneously with measurements of oxygen production. Thus it is not possible to say whether during the initial period of illumination there is an excess oxygen pro- duction relative to the deficit of carbon dioxide uptake, as might be expected in terms of the mechanism considered above.
If no anomalies can be observed during the induction phase this would suggest that if an oxidative reaction is coupled to carbon dioxide fixation it cannot involve free oxygen. This conclusion would be consistent with the observations of Brown using tracer oxygen and also with the recent work of Allen (1954) which shows that under certain conditions photosynthesis can proceed at maximal rates even in the presence of very low concentrations of free oxygen.
VIII. SUMMARY Mechanisms of photosynthesis have frequently been formulated solely in kinetic terms; these must be related to present knowledge of the enzyme systems possibly concerned. Knowledge of specific enzyme systems involved in photosynthesis is lacking; only the general hypothesis exists that photosynthesis must involve hydrogen transfer from water to carbon compounds.
In photosynthesis excited chlorophyll a is presumed to produce from water a hydrogen donor in a reaction common to all photosynthetic organisms. Oxidation- reduction reactions of this type are photosensitized by chloroplasts isolated from the plant. With such in vitro chloroplast preparations the most reducing substance

Photosynthesis and respiration 61 yet known capable of reduction is coenzyme 2 (triphosphopyridine nucleotide or TPN+); this system would be unable to reduce carbon dioxide to the extent which occurs in photosynthesis. Thus, if TPN+ is the most reducing substance formed in the photochemical reaction of photosynthesis in vivo, a further ' energy-yielding ' intermediate is also required. By analogy with respiratory reactions such an inter- mediate may originate in an oxidative reaction coupled to the photochemical system. Changes in the concentration of adenosine triphosphate have been observed consequent upon illumination of the green plant cell, and it is suggested that both phosphorylation and hydrogen transfer are probably concerned in the reduction of carbon dioxide.
If this were the case, an interaction between photosynthesis and respiration might be expected; for the rate of respiration in animal mitochondria is apparently deter- mined by the phosphorylative reactions. Experimental evidence shows that the over-all rate of consumption of oxygen in the light in the steady state is unchanged in photosynthesis; clear evidence as to the rate of production of respiratory carbon dioxide in the light is lacking. During brief transient periods when green cells are given intermittent light and dark of I min. duration the rate of consumption of oxygen in the dark may be increased. This might be attributed to an enhanced rate of normal aerobic respiration due to the formation of photosynthetic inter- mediates other than sugars; it might also be attributed to secondary processes involving the uptake of oxygen such as reversible complex formation in the dark and photo-dissociation in the light. If the former interpretation is accepted the accelerated respiration may occur either only during the first moments of light or dark or it may continue throughout the light. Many experimental observations indicate that a stimulation of respiration does not persist throughout the period of illumination.
Further experiments are required to relate the possible enzymic mechanisms to the rate phenomena observed with whole cells. These should include not only further kinetic investigations but also quantitative analysis of respiratory inter- mediates both in light and dark. In the meanwhile, speculation as to a possible enzymic mechanism of photosynthesis may be useful even prior to the isolation of enzymes, first as a means of co-ordinating experimental data, and secondly as a guide as to which steps in photosynthesis represent an enzymic system peculiar to the green plant.
Present evidence is not inconsistent with the view that the photochemical step results in the formation of reduced coenzyme. The reduction of carbon dioxide is made possible either by coupling the oxidation of the coenzyme with phosphoryla- tions and the subsequent reduction of the phosphorylated carboxyl group or by oxido-reductive coupling between the reduced coenzyme and some other un- identified hydrogen donor of higher reducing potential leading to the formation of a hydrogen donor able to reduce the carbon dioxide either as carboxyl or possibly also in this case as the phosphate ester.
I wish to thank Prof. G. E. Briggs and Dr R. Hill for help during the preparation of this manuscript.

62 C. P. WHITTINGHAM
IX. REFERENCES ALLEN, F. (1954). Ph.D. thesis, Chicago. ARNON, D. I. (195 I). Extracellular photosynthetic reactions. Nature, Lond., 167, 1008. BAALSIWD, K. & BAALSRUD, K. S. (1952). The role of phosphate in carbon dioxide assimilation of
Thiobucilli. Phosphorus metabolism, 2, 544. BADIN, E. J. & CALVIN, M. (1950). The path of carbon in photosynthesis. IX. Photosynthesis,
photoreduction and the hydrogen, oxygen carbon dioxide dark reaction. J . Amer. chem. SOC. 72, 5266.
BASSHAM, J. A., BENSON, A. & CALVIN, M. (1950). The path of carbon in photosynthesis. VIII. Thc role of malic acid. J . biol. Chem. 185, 781.
BASSHAM, J. A., BENSON, A., KAY, L., HARRIS, A., WILSON, A. & CALVIN, M. (1954). Path of carbon in photosynthesis. J . Amer. chem. Sac. 76, 1760.
BENSON, A. A. & CALVIN, M. (1950). The path of carbon in photosynthesis. VII. Respiration and photosynthesis. J . exp. Bo t . I, 63.
BRACKETT, F. S., OLSON, R. A. & CRICKARD, R. G. (1953). Respiration and intensity dependence ot photosynthesis in Chlorella. J . gen. Physiol. 36, 529.
BRIGGS, G. E. & WHITTINGHAM, C. P. (1952). Factors affecting the rate of photosynthesis of Chlore l l~ at low concentrations of carbon dioxide. N m Phytol. 51, 236.
BROWN, A. H. (1953). The effects of light on respiration using isotopically enriched oxygen. Amer. J . Bot . 40, 719.
BROWN, A. H. & FRENKEL, A. W. (1953). Photosynthesis. Annu. Rev. P1. Physiol. 4, 22. BROWN, A. H. & WEBSTER, G. C. (1953). The influence of light on the rate of respiration of the bluc
green alga Anabaena. Amer. J . Bot . 40, 753. BURK, D. & WARBURG, 0. (1951). Ein Quanten-Reaktion und Kreisprozess der Energie bei der
Photosynthese. Z . Naturf. 66, 12. CALVIN, M. & BARLTROP, J. A. (1952). A possible primary quantum conversion act of photosynthesis.
J . Amer. chem. Sac. 74, 6153. CALVIN, M. & MASSINI, P. (1952). The path of carbon in photosynthesis. XX. Steady state.
Experientiu, 8, 445. CLENDENNING, K. A. & EHRMANTRAUT, H. (1950). Photosynthesis and Hill reactions by whole
Chlorella cells in continuous and flashing light. Arch. Biochem. 29, 387. CRAMER, M. & MYERS, J. (1948). Nitrate reduction and assimilation in Chlorella. J. gen. Phjtsiol.
DAMASCHKE, K., T ~ T , F., BURK, D. & WARBURG, 0. (1953). An electrochemical demonstration of the energy cycle and maximum quantum yield in photosynthesis. Biochim. biophys. Ac ta , 12,347.
DUTTON, H. J. & MANNING, W. M. (1943). Evidence for carotenoid-sensitized photosynthesis in the diatom Nitzschia Closterium. Amer. J . Bot . 28, 5 16.
DUTTON, H. J., MANNING, W. M. & DUGGAR, B. M. (1943). Chlorophyll fluorescence and energ?- transfer in the diatom Nitzschia closterium. J . phys. Chem. 47, 308.
DUYSENS, L. N. M. (1952). Transfer of excitation energy in photosynthesis. Thesis, Utrecht. EHRMANTRAUT, H. & RABINOWITCH, E. I. (1952). Kinetics of Hill reaction. Arch. Biochem. 38, 67. EMERSON, R. & LEWIS, C. M. (1939). Factors influencing the efficiency of photosynthesis. Amer. J.
Bot. 26, 808. EMERSON, R. & LEWIS, C. M. (1941). Carbon dioxide exchange and the measurement of the quantum
yield of photosynthesis. Amer. J . Bo t . 28, 789. EMERSON, R. & LEWIS, C. M. (1942). The photosynthesis efficiency of phycocyanin in Chroococcus
and the problem of carotenoid participation in photosynthesis. J. gen. Physiol. 25, 579. EMERSON, R. & LEWIS, C. M. (1943). The dependence of the quantum yield of Chlorellm photcl-
synthesis on wave length of light. Amer. J. Bat. 30, 165. EMERSON, R. L., STAUFFER, J. F. & UMBREIT, W. W. (1944). Relationships between phosphoryla-
tion and photosynthesis in Chlorella. Amer. J . Bot. 31, 107. EVANS, H. J. & NASON, A. (1953). Pyridine nucleotide-nitrate reduction from extracts of highcr
plants. Plant Physiol. 28, 233. FRANCK, J. (1953). Participation of respiratory intermediates in photosynthesis. Arch. Biochem. 45.
190. FRENCH, C. S. & RABIDEAU, G. S. (1945). The quantum yield of oxygen production by chloroplasts
suspended in solutions containing ferric oxalate. J. gen. Physiol. 28, 329. FRENCH, C. S. & YOUNG, V. K. (1952). The fluorescence spectra of red algae and the transfcr of
energy from phycoerythrin to phycocyanin and chlorophyll. J . gen. Physiol. 35, 873.
32s 93.

Photosynthesis and respiration 63 GAFFRON, H., FACER, E. W. & ROSENBERG, J. L. (1951). Intermediates in photosynthesis. Synip.
GIBBS, M. (1953). Effect of light intensity on the distribution of C14 in sunflower leafmetabolism
HAXO, F. T. & BLINKS, L. R. (1950). Photosynthetic action spectra of marine algae. J. gen. Physiol.
HILL, R. & SCARISBRICK, R. (1940). The reduction of ferric oxalate by isolated chloroplasts. Proc.
HILL, R. & WHITTINGHAM, C . P. (1953). The induction phase of photosynthesis in Chlorella deter-
HOLZER, M. & HOLZER, E. (1952). Enzyme des Kohlenhydratstoffwechseln in Chlorella. Chem. Ber.
KANDLER, 0. (1950). Uber die Beziehungen zwischen Phosphathaushalt und Photosynthese.
KESSLER, E. (1953). Uber den Mechanismus der Nitratereduction von Griinalgen. Flora, 140, I. KOK, B. (1948). A critical consideration of the quantum yield of Chlorella photosynthesis. Enzy-
mologia, 13, I. LARSEN, H., YOCUM, C. S. 81 VAN NIEL, C. B. (1952). On the energetics of the photosyntheses in
green sulphur bacteria. J . gen. Physiol. 36, 161. LEHNINGER, A. L. (195 I ) . Phosphorylation coupled to oxidation of dihydro-diphosphopyridine
nucleotides. J. biol. Chem. 190, 345. VAN NIEL, C. B. (1941). The bacterial photosyntheses and their importance for the general problem of
photosynthesis. Advanc. Enzymol. I, 263. VAN NIEL, C. B., ALLEN, M. B. & WRIGHT, B. E. (1953). On the photochemical reduction of nitrate
by algae. Biochini. biophys. Acta, 12, 67. VAN NORMAN, R. & BROWN, A. H. (1952). The relative rates of photosynthetic assimilation of iso-
topic forms of carbon dioxide. Plant Physiol. 27, 691. RABINOWITCII, E. I . (1945). Photosynthesis, vol. I. Academic Press. RABINOWITCH, E. I. (1951). Photosynthesis. Annu. Rev. Phys. Chem. 2, 361. RUBEN, S. (1943). Photosynthesis and phosphorylation. J. Amer. chem. SOC. 65, 279. STEEMANN NIELSEN, E. (1953). Carbon dioxide concentration, respiration during photosynthesis
and maximum quantum yield of photosynthesis. Physiol. Plant, 6 , 316. STREHLER, 13. L. (1953). Firefly luminescence in the study of energy transfer mechanisms. 11.
Adenosine triphosphate and photosynthesis. Arch. Biochem. 43, 67. TANADA, T. (195 I ) . The photosynthetic efficiency of carotenoid pigments in Navicula minima. Amer.
J. Bot. 38, 276. TOLMACH, L. J. (195 I). The influence of triphosphopyridine nucleotide and other physiological
substances upon oxygen evolution from illuminated chloroplasts. Arch. Biochem. 33, 120. TURNER, J . S. (1951) . Respiration. The Pasteur effect in plants. Annu. Rev. P1. Physiol. 2,
14.5. UMBREIT, N. (1947). Problems of autotrophy. Bact. Rev. 11, 157. VISHNIAC, W. & OCHOA, S. (1952). Fixation of carbon dioxide coupled to photochemical reduction
of pyridine nucleotides by chloroplast preparations. J. biol. Chem. 95, 75. V~CLER, K. G. & UMBREIT, W. W. (1942). Studies on the metabolism of the autotrophic bacteria.
111. The nature of the energy storage material active in the chemo-synthetic process. J. gen. Physiol. 26, 157.
WARBURG, 0. (1919). Uber die Geschwindigkeit der photochemischen Kohlensaurezersetzung in lebenden Zellen. Biochenr. 2. 100, 230.
WARBURG, 0. (1953). Messung des Quantenbedarfs dcr Photosynthese fiir sehr dunne Zellsuspen- sionen. Biochim. biophys. Acta, 12, 356.
WARBURG, 0. & GELEICK, H. (1951). Uber den Gewinn im Kreisprozess der Photosynthese. 2. Naturf. 6 6 , 134.
WARBURG, O., GELEICK, H. & BRIESE, K. (195 I a). Weitere Steigerung des Energiegewinns im Kreis- prozess der Photosynthese. Z . Nuturf. 6 6 , 285.
WARBURG, O. , GELEICK, H. & BRIESE, K. (19516). uber die Aufspaltung der Photosynthese in Lichtreaktion und Ruckreaktion. 2. Naturf. 6 b , 417.
WARBURG, O., KRIPPAHL, G., BucHIioLz, W. & SCHR~DER, W. (1953). Weiterentwicklung der Methoden zur Messung der Photosynthese. 2. Naturf. 8 b , 675.
WARBURG, 0. & LUTTGENS, W. (1946). Photochemische Reduktion des Chinons in griinen Zellen und Granula. Biochimia, 11, 303.
Soc. exp. Biol. 5 , 262,
during photosynthesis. Arch. Biochem. 45, 156.
33, 389.
Roy. SOC. B, 129, 238.
mined by a spectroscopic method. New Phytol. 52, 133.
85, 655.
2. Naturf. 5 b , 423.

64 C. P. WHITTINGHAM WARBURG, 0. & NEGELEIN (1920). uber die Reduktion der Saltpetersaure in griinen Zellen. Bio-
chem. 2. 11,66. WASSINK, E. C., KATZ, E. & DORRESTEIN, R. (1942). On photosynthesis and fluorescence of bacterio-
chlorophyll in Thiorhodaceae. Enzymologia, 10, 285. WASSINK, E. C. & KERSTEN, J. A. H. (1946). Observations sur le spectre d’absorption et sur le rBle
des carotenoides dans la photosynthese des Diatomtes. Enzymologk, 12, .3. WASSINK, E. C., TJIA, J. E. & WINTERMANS, J. F. G. M. (1949). Phosphate exchanges in purple
sulphur bacteria in connection with photosynthesis. Proc. K . Akad. Wet. Amst. 52, 412. WASSINK, E. C., WINTERMANS, J. F. G. M. & TJIA, J. E. (195 I). Phosphate exchanges in Chlorella in
relation to conditions for photosynthesis. Proc. K . Akad. Wet. Amst. 54, 41. WEIGL, J. W., WARRINGTON, P. M. & CALVIN, M. (1951). The relation of photosynthesis to respira-
tion. J . Amm. chem. SOC. 73, 5 0 5 8 . WHITTINGHAM, C. P. (1952a). The chemical mechanism of photosynthesis. Bot. Rev. 18, 245. WHITTINGHAM, C. P. (1952b). Inhibition of photosynthesis by cyanide. Nature, Lond., 169, 838. WHITTINGHAM, C. P. (1952~). Rate of photosynthesis and concentration of carbon dioxide in
WHITTINGHAM, C. P. (1954). Photosynthesis in Chlorella during intermittent illumination of long Chlorella. Nature, Lond., 170, 1017.
periodicity. Plant Physiol. (in the press).
ADDENDUM
Arnon, Allen & Whatley (1954) have recently reported that chloroplast prepara- tions from spinach are able to catalyse in the light the conversion of AMP to ATP and the fixation of tracer carbon dioxide. They believe their preparations to consist largely of whole chloroplasts and they suggest that fragmentation results in the loss of these activities. Previously most workers have used chloroplast preparations consisting largely of fragments ; such preparations have been referred to in this article as the chloroplast system. The recent work implies that the chloroplast itself contains not only the catalytic system responsible for the chloro- plast reaction exhibited by the fragments but the further systems necessary for phosphorylation and uptake of tracer carbon. Thus the preparations of Arnon et al. behave as if they consisted of a mixture of fragmented chloroplasts together with mitochondria.
REFERENCE ARNON, D. I., ALLEN, M. B. & WHATLEY, F. R. (1954). Photosynthesis by isolated chloroplasts.
Nature, Lond., 174, 394.