Embed Size (px)
Citation preview

Plant Physiol. (1970) 46, 650-654
Enzymic Mechanism of Starch Breakdown in GerminatingRice SeedsIII. a-AMYLASE ISOZYMES'
Received for publication May 7, 1970
YOSHIMASA TANAKA, ToMOKO ITO, AND TAKASHI AKAZAWASeikagaku Seigyo Kenkyu Shisetsu, Nagoya University, School of Agriculture, Chikusa, Nagoya, Japan
ABSTRACT
The formation of amylase isozymes in germinating rice(Oryza sativa) seeds was studied by isoelectric focusing onpolyacrylamide gel disc electrophoresis. Time sequencecomparisons of the amylase zynmogram were made betweenextracts from gibberellic acid-treated embryoless and em-bryo-attached 1half-endosperm of rice seeds. In both cases,4 major and 9 to 10 minor isozyme bands were detectable atthe maximal stage of the enzyme induction. However, in theembryo-attached half-seeds, bands started to diminishafter the 5th day of incubation, in agreement with the re-sults of time sequence analyses of enzyme activities. Nearlyidentical patterns of amylase isozyme bands on a polv-acrylamide gel disc electrophoresis in combination withisoelectric focusing indicate the intrinsic role of gibberellicacid in the starch breakdown in germinating rice seeds.We tentatively assign the newly synthesized enzymes to bea-amylases based on experimental results concerning thelability of the preparation on a prolonged treatment at pH3.3 and the stability on heat treatment for 13 minuttes at70 C.
Our previous experimental results on both chemical analysesand determination of enzyme activities have shown that theamylolytic pathway is the major mechanism for the breakdownof reserve starch in endosperm tissues of germinating rice seeds(17, 18). This finding is in good agreement with results reportedby several workers on the formation of a-amylase in germinatingcereal seeds, i.e., barley and rice, induced by gibberellic acid(2, 16, 19, 20, 22-24, 29-32, 35). A series of extensive studiescarried out by Varner and his associates (3, 4, 6, 30-33) havewell established the synthesis de novo of a-amylase in germinatingbarley seeds. The pattern of the enhancement of a-amylaseactivities in embryoless half-seeds in response to GA treatmentwas found to be analogous to that occurring in the intact germi-nating seeds. They have proposed that GA moves from embryoto aleurone cells, where it promotes the synthesis of a-amylase.The production of GA in embryos of developing barley seedshas been demonstrated by Radley (26, 27). However, there isno direct proof that the biochemical events occurring in intactseeds and that occurring in the GA-treated half-seeds are identi-cal. This question must be critically examined, because isozymesof a-amylase were detected in the GA-treated embryoless half-
1 This investigation was supported in part by the United StatesDepartment of Agriculture under the PL 480 Program (FG-JA-126).
seeds as well as in germinating intact seeds of barley (7, 14, 15,21, 33). In an experiment reported in this paper, we have studiedthe time sequence change of amylase isozymes in GA-treatedembryoless and in intact endosperm of rice seeds, to gain deeperinsight into the enzymic mechanism of the hydrolytic breakdownof reserve starch. The isoelectric focusing technique (12) on apolyacrylamide gel disc electrophoresis proved to be advan-tageous for detection of many isozymes (13, 34).
MATERIALS AND METHODS
Half-endosperm Method of Germinating Rice Seed. Seeds ofrice plants, Oryza sativa L. var. Fujiminori harvested in Septem-ber 1969, were used throughout this study. Seeds were dehulledby hand, and the whole seeds were cut into two parts with arazor blade. Both embryoless and embryo-attached half-seedswere sterilized by soaking for 1 min in absolute ethanol and thendipping for 20 min in an aqueous solution of 10%c (w v) bleachingpowder (NaOCI) (14). After thoroughly rinsing with sterilizeddistilled H20, 10 each of embryoless and embryo-attached half-seeds were placed aseptically in 10-ml flasks containing 3 ml ofeither 2 X 10-2,ug GA ml or distilled H20, previously sterilizedby autoclaving at 120 C for 15 min. Then, the contents of theflasks were aseptically incubated at 30 C. No discernible changesoccurred in embryoless half-seeds, without the addition of GA.However, the dissolution of starchy reserve occurred in the(+GA) system after 5 days of incubation. The development ofboth shoot and root in embryo-attached half-seeds was essen-tially the same as that occurring in normally germinating seeds.Enzyme Assays. At each germination stage, the seed pieces
were removed from the incubation medium and were ground ina chilled mortar with 3.0 ml of 0.01 M tris-HCl buffer (pH 8.5).After centrifugation of the homogenate at 15,000 rpm for 15min, an aliquot of the supernatant solution was used for theassay of a-amylase activities by the method of Shuster and Gifford(28). An aliquot of each incubation medium was assayed forextracellular enzyme activity. As a quantitative measure of therelative enzyme activities, unit activities (mg of starch brokendown per seed) were calculated from the data for 5 min of incuba-tion at 30 C (cf 17).Measurements of enzyme activities were also carried out by
determining the increase in the amount of reducing sugars formedby the method of Bernfeld (1) with maltose as reference. Ab-sorbancy at 540 nm was measured by a Hitachi model 356 twowave length spectrophotometer.
In order to detect the possible presence of ,B-amylase in thecrude extract, both heat and low pH treatments were carried outfollowing the method of Paleg (23). To an aliquot (1.0 ml) ofthe crude extract (I) was added 1.0 ml of 0.01 M Ca-acetate, andthe mixture incubated at 70 C for 15 min. Then the mixture wasplaced in an ice bath until used for measurements of enzyme
650
STARCH BREAKDOWN IN RICE SEEDS. III
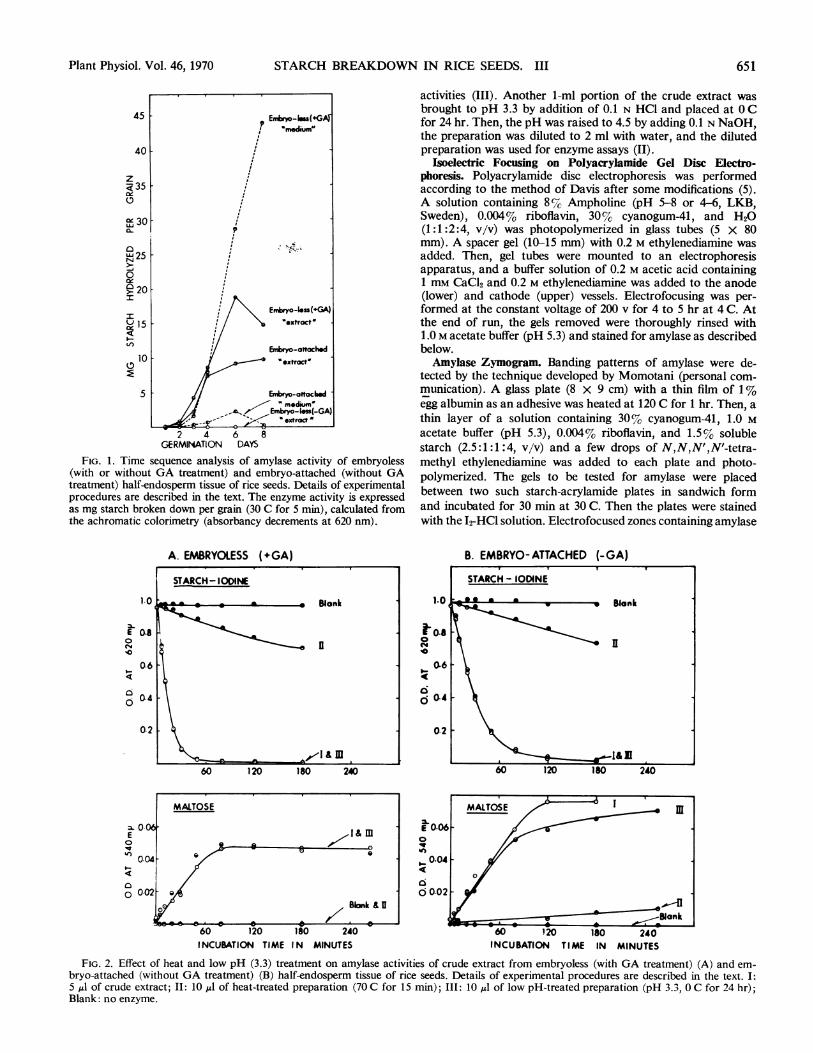
GERMINATION DAYSFIG. 1. Time sequence analysis of amylase activity of embryoless
(with or without GA treatment) and embryo-attached (without GAtreatment) half-endosperm tissue of rice seeds. Details of experimentalprocedures are described in the text. The enzyme activity is expressedas mg starch broken down per grain (30 C for 5 min), calculated fromthe achromatic colorimetry (absorbancy decrements at 620 nm).
A. EMBRYOLESS (+GA)
1.
E0
410
0
M. o o
~-0.04E0
IV4
O 0.02
MALTOSE
I& m
=Bonk & u
60 120 180 240INCUBATION TIME IN MINUTES
activities (III). Another 1-ml portion of the crude extract wasbrought to pH 3.3 by addition of 0.1 N HCI and placed at 0 Cfor 24 hr. Then, the pH was raised to 4.5 by adding 0.1 N NaOH,the preparation was diluted to 2 ml with water, and the dilutedpreparation was used for enzyme assays (II).
Isoelectric Focusing on Polyacrylamide Gel Disc Electro-phoresis. Polyacrylamide disc electrophoresis was performedaccording to the method of Davis after some modifications (5).A solution containing 8% Ampholine (pH 5-8 or 4-6, LKB,Sweden), 0.004% riboflavin, 30% cyanogum-41, and H20(1:1:2:4, v/v) was photopolymerized in glass tubes (5 X 80mm). A spacer gel (10-15 mm) with 0.2 M ethylenediamine wasadded. Then, gel tubes were mounted to an electrophoresisapparatus, and a buffer solution of 0.2 M acetic acid containing1 mM CaCl2 and 0.2 M ethylenediamine was added to the anode(lower) and cathode (upper) vessels. Electrofocusing was per-formed at the constant voltage of 200 v for 4 to 5 hr at 4 C. Atthe end of run, the gels removed were thoroughly rinsed with1.0 M acetate buffer (pH 5.3) and stained for amylase as describedbelow.Amylase Zymogram. Banding patterns of amylase were de-
tected by the technique developed by Momotani (personal com-munication). A glass plate (8 X 9 cm) with a thin film of 1%egg albumin as an adhesive was heated at 120 C for 1 hr. Then, athin layer of a solution containing 30% cyanogum-41, 1.0 Macetate buffer (pH 5.3), 0.004% riboflavin, and 1.5% solublestarch (2.5:1:1:4, v/v) and a few drops of N,N,N',N'-tetra-methyl ethylenediamine was added to each plate and photo-polymerized. The gels to be tested for amylase were placedbetween two such starch-acrylamide plates in sandwich formand incubated for 30 min at 30 C. Then the plates were stainedwith the I2-HC1 solution. Electrofocused zones containing amylase
B. EMBRYO-ATTACHED (-GA)
60 120 180 240INCUBATION TIME IN MINUTES
FIG. 2. Effect of heat and low pH (3.3) treatment on amylase activities of crude extract from embryoless (with GA treatment) (A) and em-bryo-attached (without GA treatment) (B) half-endosperm tissue of rice seeds. Details of experimental procedures are described in the text. I:5 ,Al of crude extract; II: 10 ,IA of heat-treated preparation (70 C for 15 min); III: 10 ,dl of low pH-treated preparation (pH 3.3, 0 C for 24 hr);Blank: no enzyme.
Plant Physiol. Vol. 46, 1970 651
TANAKA, ITO, AND AKAZAWA
activities hydrolyzed the starch film and appeared as decolorizedbands on a blue background.
RESULTS
Results presented in Figure 1 show the time sequence analysesof amylase activity of variously treated endosperm samples,employing the I2-starch decoloration method. In both embryoless(+GA) and embryo-attached (-GA) half-seed endosperms, amarked increase in enzyme activity occurred after a lag periodof about 1 day. However, there was a notable difference betweenthe two systems. Amylase activity in the medium of GA-treated,embryoless endosperm tissue showed a nearly linear increasebetween 3 and 8 days, while that from the seed extract increasedslightly between 3 and 6 days and declined thereafter. After 8days essentially no starch-degrading activity was detectable inthe absence of GA. In the embryo-attached endosperm, on theother hand, the enzyme activity began to level off after about 4to 5 days; the pattern was similar to the one observed in intactgerminating seeds (cJf Fig. 3 of Ref. 17). Practically no enzymeactivity was measurable in the medium containing the embryo-attached endosperm tissue.That the enzyme examined is an a-amylase is suggested by the
results shown in Figure 2. The breakdown of soluble starch bythe crude extract from both GA-treated embryoless and embryo-attached half-seeds as measured by the 12-starch reaction (ab-sorbancy at 620 nm) was hardly affected by heating at 70 C for15 min. But lowering the pH of the preparation to 3.3 resultedin a drastic decline in enzyme activity. An identical trend wasobserved by analyzing the maltose formed (1). It is thus impli-
DAYS AFTER
cated that j3-amylase may play a minor role in hydrolyzing thereserve starch in germinating rice seed endosperm, unlike thecase with barley seeds (23). By comparing the results of the twomethods of detecting starch hydrolysis as presented in Figure 2,it will be noted that the amount of low molecular weightreducing sugars formed accounts for only a part of the total starchbroken down in seeds.Zymograms illustrated in Figures 3 and 4 are the results of
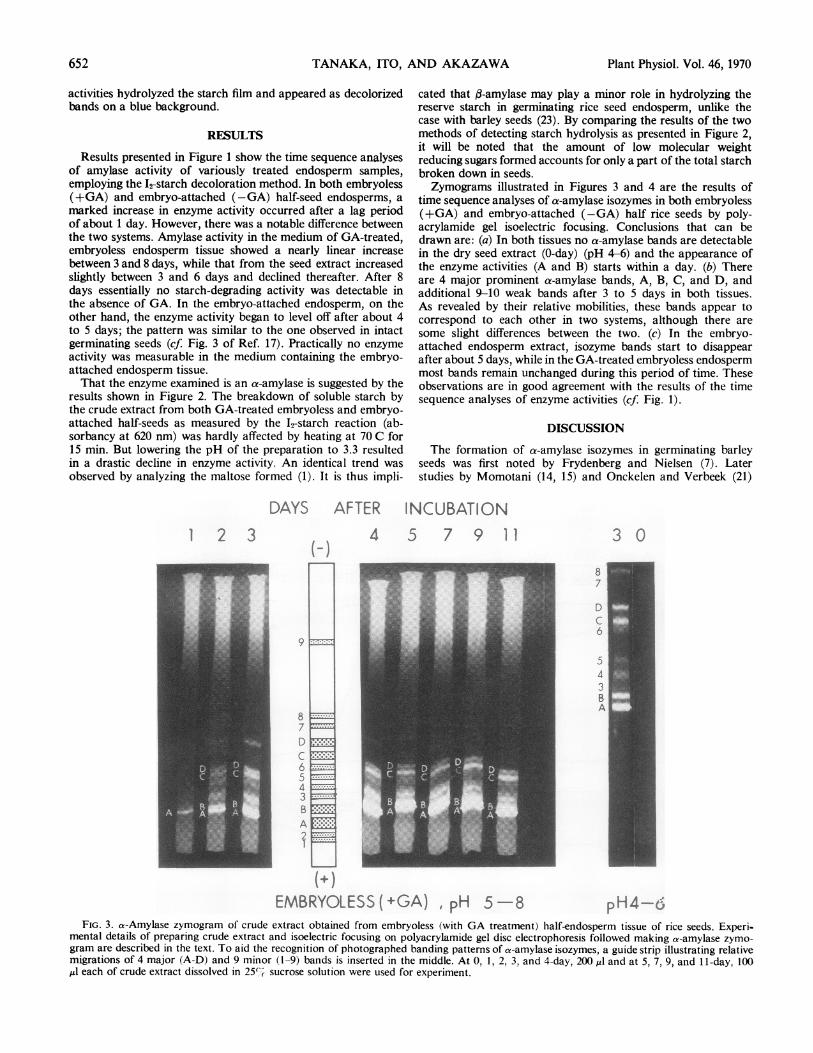
time sequence analyses of a-amylase isozymes in both embryoless(+GA) and embryo-attached (-GA) half rice seeds by poly-acrylamide gel isoelectric focusing. Conclusions that can bedrawn are: (a) In both tissues no a-amylase bands are detectablein the dry seed extract (0-day) (pH 4-6) and the appearance ofthe enzyme activities (A and B) starts within a day. (b) Thereare 4 major prominent a-amylase bands, A, B, C, and D, andadditional 9-10 weak bands after 3 to 5 days in both tissues.As revealed by their relative mobilities, these bands appear tocorrespond to each other in two systems, although there aresome slight differences between the two. (c) In the embryo-attached endosperm extract, isozyme bands start to disappearafter about 5 days, while in the GA-treated embryoless endospermmost bands remain unchanged during this period of time. Theseobservations are in good agreement with the results of the timesequence analyses of enzyme activities (cf. Fig. 1).
DISCUSSION
The formation of a-amylase isozymes in germinating barleyseeds was first noted by Frydenberg and Nielsen (7). Laterstudies by Momotani (14, 15) and Onckelen and Verbeek (21)
INCUBATION
. .,. ._
F...-.-....
_ ...
4 D 7 9 11
EMBRYOLESS(+GA) , pH 5 -8FIG. 3. a-Amylase zymogram of crude extract obtained from embryoless (with GA treatment) half-endosperm tissue of rice seeds. Experi-
mental details of preparing crude extract and isoelectric focusing on polyacrylamide gel disc electrophoresis followed making a-amylase zymo-
gram are described in the text. To aid the recognition of photographed banding patterns of a-amylase isozymes, a guide strip illustrating relativemigrations of 4 major (A-D) and 9 minor (1-9) bands is inserted in the middle. At 0, 1, 2, 3, and 4-day, 200 IA and at 5, 7, 9, and 11-day, 100
,ul each of crude extract dissolved in 25'% sucrose solution were used for experiment.
3)01 2 3
9
87
6543BA2
87
DC6
J.4
BA
pH4-c
652 Plant Physiol. Vol. 46, 1970

Plant Physiol. Vol. 46, 1970 653STARCH BREAKDOWN IN RICE SEEDS. Il[
DAYS AFTER INCUBATION
1 2 3
9
87DC6543BA21
4 5 7 9 11
H(I
....
,. ...
.........--. 4 0
DC6
543BA1,2
EMBRYO-ATTACHED (-GA), pH 5-8 pH4-6FIG. 4. a-Amylase zymogram of crude extract obtained from embryo-attached (without GA treatment) half-endosperm tissue of rice seeds
Experimental procedures employed were essentially identical with those explained for Figure 3.
involved the use of GA-treated half-seeds. The latter workershave also described the presence of five isozymes in crude extractsfrom developing seeds. However, details on the correspondenceof isozyme bands to those in the GA-treated half-seeds aremissing. Based on technical problems inherent in the presentexperiments, it is difficult to establish that the a-amylase isozymesfrom embryoless (+GA) half-seeds are truly identical to thosefrom embryo-attached half-seeds. In fact, it appears that thereare some details which differ slightly, but we are inclined tospeculate that the patterns of a-amylase isozymes from the twosystems are basically similar, if not identical.The inducible formation of a-amylase in barley half-seeds
due to the exogenous addition of GA has been taken as circum-stantial evidence for the promoting effect of GA on the enzymesynthesis in embryo. Two lines of experimental results appearto substantiate this thesis; (a) production ofGA in the scutellumof barley embryos during the onset of germination (22, 26, 27),and (b) the ultrastructural studies on the aleurone cells of GA-treated barley half-seeds in comparison with that of normal seeds(9-11, 25). Therefore, while there remains much to be studiedto correlate the biochemical events occurring in the germinatingcereal seeds and the accompanying morphological changes, weconsider that our present finding provides additional experi-mental evidence for the intrinsic role ofGA in the process of thebreakdown of reserve starch in germinating cereal grains.The nature and the physiological function of multiple forms
of a-amylase that occur in germinating cereal seeds cannot, atpresent, be assessed. Some of the isozymes detected might haveresulted from the degradation with or without modification ofthe enzyme molecules during the preparative steps. Among somepossibilities, the proteolytic breakdown of the newly synthesizeda-amylase by a similarly GA-induced protease (8) might have
caused the formation of multiple forms of the enzyme that arereadily detectable by gel electrophoresis. As yet we presumethat some of these isozymes are functioning intracellularly.Our recent experiments (29) have demonstrated that the a-amylase isozymes formed in the GA-treated barley endospermare different in their action patterns as well as their kineticproperties. It is thus probable that various types of a-amylaseare functioning cooperatively in the complete breakdown ofreserve starch in the germinating rice seeds.
Acknowledgments-The authors wish to record their most sincere gratitude to Dr. T.Kosuge, University of California, Davis, for critically reading the manuscript andsuggesting some changes. Technical advice provided by Dr. Y. Momotani, OsakaPrefectural University, Osaka, for isoelectric focusing polyacrylamide gel disc electro-phoresis is gratefully acknowledged.
LITERATURE CITED
1. BERNFELD, P. 1955. Amylase, a and jS. In: S. P. Colowick and N. 0. Kaplan, eds.,Methods in Enzymology, Vol. 1. Academic Press, New York. pp. 149-58.
2. BRIGGS, D. E. 1963. Biochemistry of barley germination: Action of gibberellic acidon barley endosperm. J. Inst. Brew. 69: 13-19.
3. CHRISPEELS, M. J. AND J. E. VARNER. 1967. Gibberellic acid-enhanced synthesisand release of a-amylase and ribonuclease by isolated barley aleurone layers.Plant Physiol. 42: 398-406.
4. CHRISPEELS, M. J. AND J. E. VARNER. 1967. Hormonal control of enzyme synthesis:On the mode of action of gibberellic acid and abscisin in aleurone layers ofbarley. Plant Physiol., 42: 1008-1016.
5. DAVIS, B. J. 1964. Disc electrophoresis. II: Method and application to human serumproteins. Ann. N. Y. Acad. Sci. 121: 404-427.
6. FILNER, P. AND J. E. VARNER. 1967. A test for de novo synthesis of enzymes:Density labeling with H2018 of barley a-amylase induced by gibbereUic acid.Proc. Nat. Acad. Sci. U.S.A. 58: 1520-1526.
7. FRYDENBERG, 0. AND G. NIELSEN. 1965. Amylase isozymes in germinating barleyseeds. Hereditas 54: 123-139.
8. JACOBSEN, J. V. AND J. E. VARNER. 1967. Gibberellic acid-induced synthesis byprotease by isolated aleurone layers of barley. Plant Physiol. 42: 1596-1600.

-TANAKA, ITO, AND AKAZAWA
9. JONES, R. L. 1969. Gibberellic acid and the fine structure of barley aleurone cells.I. Changes during the lag-phase of a-amylase synthesis. Planta 87: 119-133.
10. JONEs, R. L. 1969. Gibberellic acid and the fine structure of barley aleurone cells.II. Changes during the synthesis and secretion of a-amylase. Planta 88: 73-86.
11. JONES, R. L. 1969. The fine structure of barley aleurone cells. Planta 85: 319-375.12. JONSSON, M. AND E. PETTERSSON. 1968. Isoelectric focusing of plant pigments.
Sci. Tools 15: 2-6.13. MALACINSKI, G. M. AND W. J. RUTrER. 1969. Multiple molecular forms ofa-amy-
late from the rabbit. Biochemistry 8: 4382-4390.14. MOMOTANI, Y. AND J. KATO. 1966. Isozymes of a-amylase induced by gibberellic
acid in embryoless grains of barley. Plant Physiol. 41: 1395-1396.15. MOMOTANI, Y. AND J. KATO. 1967. Hormonal regulation on the induction ot
a-amylase isozymes in the embryoless endosperm of barley. Plant Cell Physiol.8: 439-445.
16. MURAKAMI, Y. 1966. Bioassay of gibberellins using rice endosperm and some
problems of its application. Bot. Mag. Tokyo 79: 315-397.17. MURATA, T., T. AKAZAWA, AND S. FuKUCHI. 1968. Enzymic mechanism of starch
breakdown in germinating rice seeds. I. An analytical study. Plant Physiol.43: 1899-1905.
18. NOMURA, T., Y. KONO, AND T. AKAZAWA. 1969. Enzymic mechanism of starchbreakdown in germinating rice seeds. II. Scutellum as the site of sucrose syn-
thesis. Plant Physiol. 44: 765-769.19. OGAWA, Y. 1966. Effects of various factors on the increase of a-amylase activity
in rice endosperm induced by gibberellin A3. Plant Cell Physiol. 7: 509-517.20. OGAWA, Y. 1966. On the effects of plant extracts on the a-amylase activity in rice
endosperm. Plant Cell Physiol. 7: 519-525.21. VON ONCKELEN, H. A., AND R. VERBEEK. 1969. Formation of a-amylase isozymes
during germination of barley. Planta 88: 255-260.22. PALEG, L. 1964. Cellular localization of the gibberellin induced response of barley
endosperm. In: Regulateurs Naturels de la Croissance Vegetale, Vol. 88.Centre National de la Recherche Scientifiqcue, Paris. pp. 303-317.
Plant Physiol. Vol. 46, 1970
23. PALEG, L. G. 1960. Physiological effects of gibberellic acid: I. On carbohydratemetabolism and amylase activity of barley endosperm. Plant Physiol. 35: 293-299.
24. PALEG, L. G. 1960. Physiological effects of gibberellic acid: II. On starch hydro-lyzing enzymes of barley endosperm. Plant Physiol. 35: 902-906.
25. PALEG, L. AND B. HYDE. 1964. Physiological effects of gibberellic acid: VII. Elec-tron microscopy of barley aleurone cells. Plant Physiol. 39: 673-680.
26. RADLEY, M. 1967. Site of production of gibberellin-like substances in germinatingbarley embryos. Planta 75: 164-171.
27. RADLEY, M. 1969. The effect of the endosperm on the formation of gibberellin bybarley embryos. Planta 86: 218-223.
28. SHUSTER, L. AND R. H. GIFFORD. 1962. Changes in 3'-nucleotidase during thegermination of wheat embryos. Arch. Biochem. Biophys. 96: 534-540.
29. TANAKA, Y. AND T. AKAZAWA. 1970. a-Amylase isozymes in gibberellic acid-treated barley half-seeds. Plant Physiol. 46: 586-591.
30. VARNER, J. E. 1964. Gibberellic acid-controlled synthesis of a-amylase in barleyendosperm. Plant Physiol. 39: 413-415.
31. VARNER, J. E. AND G. R. CHANDRA. 1964. Hormonal control of enzyme synthesisin barley endosperm. Proc. Nat Acad. Sci. U.S.A. 52: 100-106.
32. VARNER, J. E., G. R. CHANDRA, AND M. J. CHRISPEELS. 1965. Gibberellic acid-controlled synthesis of a-amylase in barley endosperm. J. Cell Comp. Physiol.66: 55-68.
33. VARNER, J. E. AND M. M. JOHRI. 1968. Hormonal control of enzyme synthesis.In: F. Wightman and G. Setterfield, eds., Biochemistry and Physiology ofPlant Growth Substances. Runge Press, Ottawa, Canada. pp. 793-814.
34. WRIGLEY, C. 1968. Gel electrofocusing-A technique for analyzing multipleprotein samples by isoelectric focusing. Sci. Tools 15: 17-23.
35. YoMo, H. 1960. Studies on the amylase activating substances. IV. On the amylaseactivating action of gibberellin. Hakko Kyokaishi 18: 600-602.
654





























