Embed Size (px)
Citation preview

THE JOURNAL OB B~o~oo~cnr, CHEMISTRY
Vol. 245, No. 7, Issue of April 10, pp. 1790-1799, 1970
Printed in U.S.A.
Enzymic Synthesis of O-Alkyl Bonds in Glycerolipids* (Received for publication, November 3, 1969)
FRED SNYDER, BOYD MALONE, AND M. L. BLANK
From the Medical Divi.sion,$ Oak Ridge Associated Universities, Oak Ridge, Tennessee 37830
SUMMARY
We have found that 0-alkyl bonds in glycerolipids are synthesized by microsomal enzymes of preputial gland tumors when diiydroxyacetone-P or glyceraldehyde-3-P and 1 J4C long chain fatty alcohols are used as substrates; ATP, coenzyme A, and Mg++ are essential cofactors. The 0-alkyl moieties formed from the 1-“C- and 9,10-3H-labeled long chain fatty alcohols show that the fatty alcohol chains are directly incorporated. Dihydroxyacetone-P is an obligate source of the glycerol portion of glyceryl ethers, since only the dihydroxyacetone-P served as a precursor when l- hydroxy-3-chloro-2-propanone phosphate was used as a specific inhibitor of triose-P isomerase, which is present in the microsomal preparation. Kinetic data on the following parameters were obtained: time, CoA, Mg++, ATP, protein, pH, and inhibitors (phosphate and I-hydroxy-3-chloro-Z- propanone phosphate). We documented the chromato- graphic behavior of the intermediates (O-alkyldihydroxy- acetone and 0-alkyldihydroxyacetone-P) and identified the 0-alkyl linkage by thin layer and gas-liquid chromatography of the isopropylidene derivatives of the 0-alkylglycerols formed by LiAlH4 reduction. Only the 1 isomeric O- alkylglycerols could be isolated after LiAlH4 reduction.
Although other nucleotides (ADP and ITP) substituted for ATP in this system, glutathione or cysteine could not replace the CoA requirement. Metals other than Mg++ did not provide optimal conditions, and copper ions inhibited the biosynthesis of 0-alkyl bonds. To some extent, dihy- droxyacetone was able to replace the dihydroxyacetone-P requirement, but it is possible that phosphorylation occurred in the system.
The enzymic synthesis of the 0-alkyl linkage is localized in microsomes and appears to decrease as the tumor ages. The preputial gland tumors contain significant quantities of O- alkyl- and 0-alk-1-enyl-linked lipid classes, but the enzyme system described was unable to make 0-alk-1-enyl bonds under our experimental conditions. However, the similar composition of the 0-alkyl and 0-alk-1-enyl moieties (16:0, 18:0, and 18:l) suggests that interconversions between these two lipid classes or their precursors may occur.
Ether-linked glycerolipids containing 0-alkyl bonds exist in polar and nonpolar classes. Their biochemistry (l), their syn-
* This research was supported in part by American Cancer Society Grant E-596A.
$ Under contract with the United States Atomic Energy Com- mission.
thesis (Z), their chemical-physical properties (3), and their analysis (Z-5) have recently been reviewed. Tsujimoto and Toyama (6) discovered the ether bond in glycerolipids nearly 50 years ago, yet the biosynthesis of the ether linkage in alkyl glyceryl ethers has remained a mystery until recently (7-9). Incorporation of fatty alcohols in wivo (16-15) and structural studies (16) of double bond locations in the alkyl chains of glyc- eryl ethers and in fatty alcohols suggested that fatty alcohols could be direct precursors of the alkyl chain. Furthermore, long chain fatty alcohols having chain lengths similar to the alkyl chains in glyceryl ethers have also been found in normal and neoplastic cells of mammals (17).
We isolated a cell-free system that could incorporate long chain fatty alcohols into alkyl glyceryl ethers and their acylated counterparts (7). In the absence of an added triose-P source, none of the organelles or the supernatant fractions isolated from homogenates of preputial gland tumors could synthesize the ether bond exoept when the microsomal and supernatant frac- tions were combined. We soon learned that the supernatant fraction was serving as a source of the glycerol moiety, and that glyceraldehyde-3-P was a substrate (8). Friedberg (18) also obtained evidence in tivo with 3H, l4C-glycerol that suggested a triose-P was involved in glyceryl ether synthesis. Our further investigation revealed that a triose-P isomerase was present in the microsomes (19, 20) and this made it difficult to distinguish between glyceraldehyde-3-P and DHAPl as the true precursor. The presence of triose-P isomerase in microsomes was apparently not caused by contamination from the soluble protein fraction, since glycerophosphate dehydrogenase was absent from the microsomes (21). Our use (21) of a specific inhibitor of triose-P isomerase, l-hydroxy-3-chloro-2-propanone phosphate (22)) finally revealed that DHAP was the obligate triose-P precursor that participates with long chain fatty alcohols in the formation of 0-alkyl linkages. The biosynthetic pathway for the formation of 0-alkyl bonds occurs in microsomes isolated from normal (19, 20) and neoplastic cells (7, 8, 23). A detailed description of the biosynthetic reaction, including kinetic and structural studies, is provided in this paper and a companion paper (24).
EXPERIMENTAL METHODS
Source of Microsomes-The enzyme fractions used in these experiments were obtained from preputial gland tumors (ESR- 586) grown in C57BL/6 mice. The source of the tumor, the transplantation technique, the excision of the tumor, and the
r The abbreviation used is: DHAP, dihydroxyacetone-P. 2 Recent studies on the synthesis of the 1-hydroxy-3-chloro-2-
propanone phosphate (HARTMAN, F. C., Biochemistry, in press) and its use in characterizing the active sites of triose-P isomerase (HARTMAN, F. C., J. Amer. Chem. Sot., in press) have been carried out by Dr. Hartman (personal communication).
1790
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from
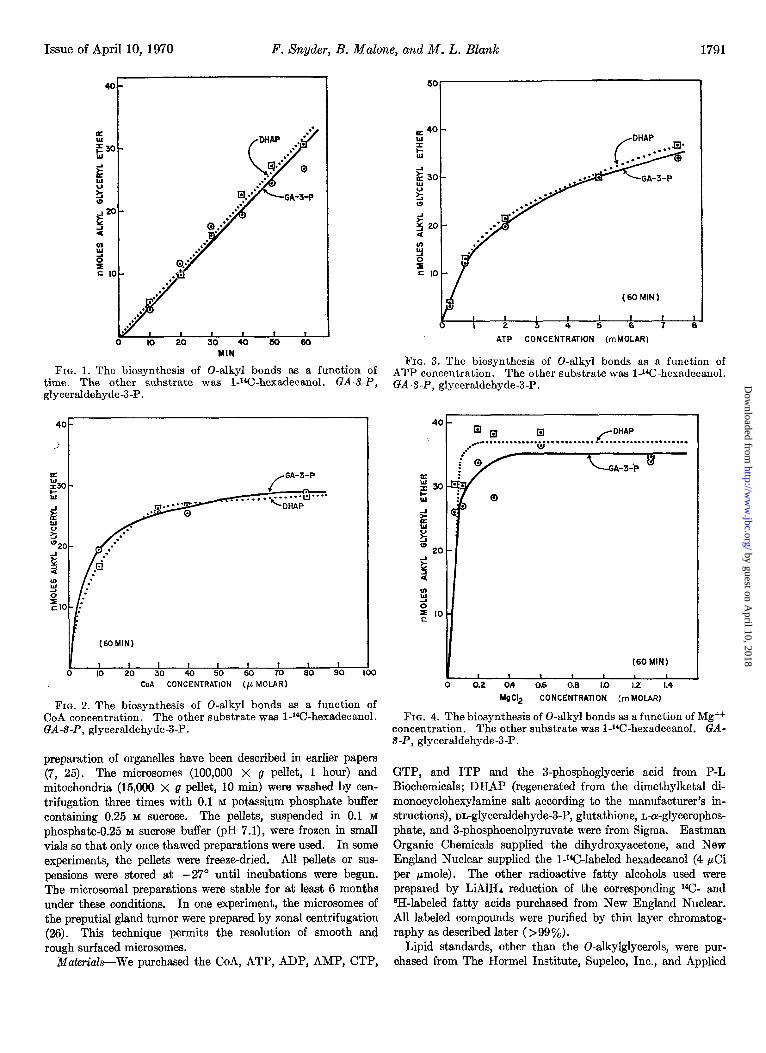
Issue of April 10, 1970 F. Snyder, B. Malone, and M. L. Blank 1791
FIG. 1. The biosynthesis of 0-alkyl bonds as a function of time. The other substrate was l-14C-hexadecanol. GA-S-P, glyceraldehyde-3-P.
t 1 I I 1 1 1 I I I 0 IO 20 30 40 50 60 70 60 90 100
Cop. CONCENTRATION (,u MOLAR)
FIG. 2. The biosynthesis of 0-alkyl bonds as a function of CoA concentration. The other substrate was l-l*C-hexadecanol. GA-S-P, glyceraldehyde-3-P.
preparation of organelles have been described in earlier papers (7, 25). The microsomes (100,000 X g pellet, 1 hour) and mitochondria (15,000 X g pellet, 10 min) were washed by cen- trifugation three times with 0.1 M potassium phosphate buffer containing 0.25 M sucrose. The pellets, suspended in 0.1 M
phosphate-O.25 M sucrose buffer (pH 7.1), were frozen in small vials so that only once thawed preparations were used. In some experiments, the pellets were freeze-dried. All pellets or sus- pensions were stored at -27” until incubations were begun. The microsomal preparations were stable for at least 6 months under these conditions. In one experiment, the microsomes of the preputial gland tumor were prepared by zonal centrifugation (26). This technique permits the resolution of smooth and rough surfaced microsomes.
Mate&&-We purchased the CoA, ATP, ADP, AMP, CTP,
I 1 1 1 I 1 t I I 2 3 4 5 6 ? 6
ATP CONCENTRATION (mMOLAR)
FIG. 3. The biosynthesis of 0-alkyl bonds as a function of ATP concentration. The other substrate was 1JGhexadecanol. GAS-P, glyceraldehyde-3-P.
q J DHAP
&” ..I.......... @.... . . . . . . . . . . . . . . . . . . . . . . . . . . . *. .*
(60 MIN)
I t 1 I I I 0 0.2 0.4 OS
MgC~ CONCEON;8RTl~~ (m fk?) “4
FIG. 4. The biosynthesis of 0-alkyl bonds as a function of Mg++ concentration. The other substrate was 1-W-hexadecanol. GA- S-P, glyceraldehyde-3-P.
GTP, and ITP and the 3-phosphoglyceric acid from P-L Biochemicals; DHAP (regenerated from the dimethylketal di- monocyclohexylamine salt according to the manufacturer’s in- structions), nn-glyceraldehyde-3-P, glutathione, n-a-glycerophos- phate, and 3-phosphoenolpyruvate were from Sigma. Eastman Organic Chemicals supplied the dihydroxyacetone, and New England Nuclear supplied the 1-I%-labeled hexadecanol (4 PCi per pmole). The other radioactive fatty alcohols used were prepared by LiAIHa reduction of the corresponding 14C- and *H-labeled fatty acids purchased from New England Nuclear. All labeled compounds were purified by thin layer chromatog- raphy as described later ( >99%).
Lipid standards, other than the 0-alkylglycerols, were pur- chased from The Hormel Institute, Supelco, Inc., and Applied
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from
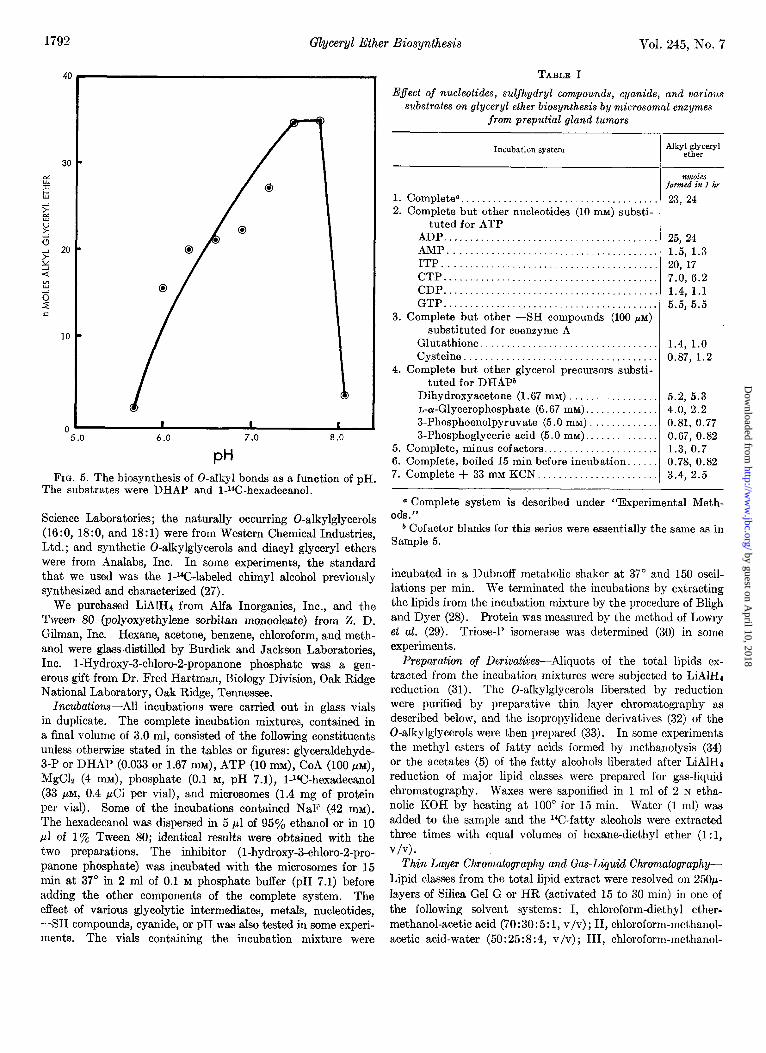
1792 Glyceryl Ether Biosynthesis Vol. 245, No. 7
0 5 .o 6.0 7.0 8.0
PH
TABLE I
Effect of nucleotides, sulfhydryl compounds, cyanide, and various substrates on ylyceryl ether biosynthesis by microsomal enzymes
FIG. 5. The biosynthesis of 0-alkyl bonds as a function of pH. The substrates were DHAP and l-14C-hexadecanol.
Science Laboratories; the naturally occurring 0-alkylglycerols (16 :O, 18 :O, and 18: 1) were from Western Chemical Industries, Ltd.; and synthetic 0-alkylglycerols and diacyl glyceryl ethers were from Analabs, Inc. In some experiments, the standard that we used was the 1-14C-labeled chimyl alcohol previously synthesized and characterized (27).
We purchased LiAlHd from Alfa Inorganics, Inc., and the Tween 80 (polyoxyethylene sorbitan monooleate) from Z. D. Gilman, Inc. Hexane, acetone, benzene, chloroform, and meth- anol were glass-distilled by Burdick and Jackson Laboratories, Inc. l-Hydroxy-3-chloro-2propanone phosphate was a gen- erous gift from Dr. Fred Hartman, Biology Division, Oak Ridge National Laboratory, Oak Ridge, Tennessee.
IncuZ&ions-All incubations were carried out in glass vials in duplicate. The complete incubation mixtures, contained in a final volume of 3.0 ml, consisted of the following constituents unless otherwise stated in the tables or figures: glyceraldehyde- 3-P or DHAP (0.033 or 1.67 mu), ATP (10 mu), CoA (100 HM), MgC12 (4 mM), phosphate (0.1 M, pH 7.1), l-14C-hexadecanol (33 PM, 0.4 PCi per vial), and microsomes (1.4 mg of protein per vial). Some of the incubations contained NaF (42 mM).
The hexadecanol was dispersed in 5 ~1 of 95% ethanol or in 10 ~1 of 1% Tween 80; identical results were obtained with the two preparations. The inhibitor (l-hydroxy-3-chloro-2-pro- panone phosphate) was incubated with the microsomes for 15 min at 37” in 2 ml of 0.1 M phosphate buffer (pH 7.1) before adding the other components of the complete system. The effect of various glycolytic intermediates, metals, nucleotides, -SH compounds, cyanide, or pH was also tested in some experi- ments. The vials containing the incubation mixture were
from preputial gland tumors
Incubation system.
1. Complete”.................................... 2. Complete but other nucleotides (10 mM) substi-
tuted for ATP
%??WleS jomed in1 hr
23, 24
ADP. . . . . . . I . . . . . . AMP . . . . ITP . . . . CTP . . . CDP. . . . . . GTP . . . . .
3. Complete but other -SH compounds (100 PM)
substituted for coenzyme A
25, 24 1.5, 1.3 20, 17 7.0, 6.2 1.4, 1.1 5.5, 5.5
Glutathione................................. Cysteine....................................
4. Complete but other glycerol precursors substi- tuted for DHAPb
1.4, 1.0 0.87, 1.2
Dihydroxyacetone (1.67 mM) 5.2, 5.3 L-oc-Glycerophosphate (6.67 mM). . 4.0, 2.2 3-Phosphoenolpyruvate (5.0 mM) 0.81, 0.77 3-Phosphoglyceric acid (5.0 mM) 0.67, 0.82
5. Complete, minus cofactors.. 1.3, 0.7 6. Complete, boiled 15 min before incubation. 0.78, 0.82 7. Complete + 33 mM KCN. . . 3.4, 2.5
o Complete system is described under “Experimental Meth- ods.”
b Cofactor blanks for this series were essentially the same as in Sample 5.
incubated in a Dubnoff metabolic shaker at 37” and 150 oscil- lations per min. We terminated the incubations by extracting the lipids from the incubation mixture by the procedure of Bligh and Dyer (28). Protein was measured by the method of Lowry et al. (29). Triose-P isomerase was determined (30) in some experiments.
Preparation of Derivatives-Aliquots of the total lipids ex- tracted from the incubation mixtures were subjected to LiAIH, reduction (31). The 0-alkylglycerols liberated by reduction were purified by preparative thin layer chromatography as described below, and the isopropylidene derivatives (32) of the 0-alkylglycerols were then prepared (33). In some experiments the methyl esters of fatty acids formed by methanolysis (34) or the acetates (5) of the fatty alcohols liberated after LiAlHI reduction of major lipid classes were prepared for gas-liquid chromatography. Waxes were saponified in 1 ml of 2 N etha- nolic KOH by heating at 100” for 15 min. Water (1 ml) was added to the sample and the i4C-fatty alcohols were extracted three times with equal volumes of hexane-diethyl ether (1: 1,
v/v). Thin Layer Chromatography and Gas-Liquid Chromatography-
Lipid classes from the total lipid extract were resolved on 250~- layers of Silica Gel G or HR (activated 15 to 30 min) in one of the following solvent systems: I, chloroform-diethyl ether- methanol-acetic acid (70:30:5: 1, v/v) ; II, chloroform-methanol- acetic acid-water (50:25:8:4, v/v) ; III, chloroform-methanol-
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from
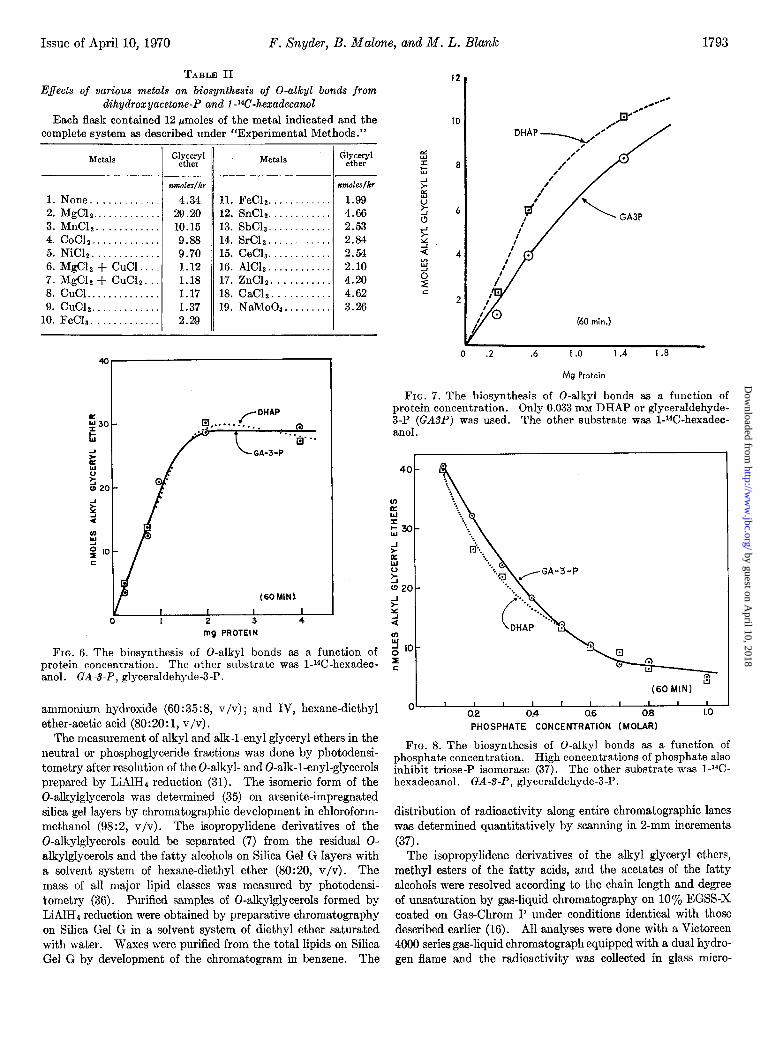
Issue of April 10, 1970 F. Snyder, B. Malone, and M. L. Blank 1793
TABLE II
Effects of various metals on biosynthesis of 0-alkyl bonds from dihydrox yacetone-P and 1 -W-hexadecanol
Each flask contained 12 pmoles of the metal indicated and the complete system as described under “Experimental Methods.”
Metals
1. None. ............ 2. MgCls ............. 3. MnC& ............ 4. CoClz ............. 5. NiC12. ............ 6. MgClz + C&l .... 7. MgClz + CuClz. .. 8. CuCl.. ............ 9. CuCl*. ............
10. FeCL. ............
-
- nmoles/hr
4.34 29.20 10.15 9.88 9.70 1.12 1.18 1.17 1.37 2.29
Metals
11. FeC12 ............ 12. SnC12. ........... 13. SbCla ............ 14. SrClz. ........... 15. CeC13. ........... 16. AK&. ........... 17. ZnClz ........... 18. CaC12. .......... 19. NaMoOl .........
zmoles/hr
1.99 4.66 2.53 2.84 2.54 2.10 4.20 4.62 3.26
4C-
30 -
20 -
IO -
1 0
1 1 I I 1 2 3 4
mg PROTEIN
FIG. 6. The biosynthesis of 0-alkyl bonds as a function of protein concentration. The other substrate was 1-W-hexadec- anol. GA-S-P, glyceraldehyde-3-P.
ammonium hydroxide (60 : 35 : 8, v/v) ; and IV, hexane-diethyl ether-acetic acid (80:20:1, v/v).
The measurement of alkyl and alk-1-enyl glyceryl ethers in the neutral or phosphoglyceride fractions was done by photodensi- tometry after resolution of the 0-alkyl- and 0-alk-1-enyl-glycerols prepared by LiAlHd reduction (31). The isomeric form of the 0-alkylglycerols was determined (35) on arsenite-impregnated silica gel layers by chromatographic development in chloroform- methanol (98:2, v/v). The isopropylidene derivatives of the 0-alkylglycerols could be separated (7) from the residual O- alkylglycerols and the fatty alcohols on Silica Gel G layers with a solvent system of hexane-diethyl ether (80:20, v/v). The mass of all major lipid classes was measured by photodensi- tometry (36). Purified samples of 0-alkylglycerols formed by LiAIHt reduction were obtained by preparative chromatography on Silica Gel G in a solvent system of diethyl ether saturated with water. Waxes were purified from the total lipids on Silica
Ii 0 .2 .6 1 .o 1 .4 1 .8
Mg Protein
FIG. 7. The biosynthesis of 0-alkyl bonds as a function of protein concentration. Only 0.033 mM DHAP or glyceraldehyde- 3-P (GASP) was used. The other substrate was 1-W-hexadec- an01 .
01 I I I I I I t I I I 0.2 0.4 0.6 0.6 1.0
PHOSPHATE CONCENTRATION (MOLAR)
FIG. 8. The biosynthesis of 0-alkyl bonds as a function of phosphate concentration. High concentrations of phosphate also inhibit triose-P isomerase (37). The other substrate was 1-W- hexadecanol. GA-S-P, glyceraldehyde-3-P.
distribution of radioactivity along entire chromatographic lanes was determined quantitatively by scanning in 2-mm increments (37).
The isopropylidene derivatives of the alkyl glyceryl ethers, methyl esters of the fatty acids, and the acetates of the fatty alcohols were resolved according to the chain length and degree of unsaturation by gas-liquid chromatography on 10% EGSS-X coated on Gas-Chrom P under conditions identical with those described earlier (16). All analyses were done with a Victoreen 4000 series gas-liquid chromatograph equipped with a dual hydro-
Gel G by development of the chromatogram in benzene. The gen flame and the radioactivity was collected in glass micro-
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from
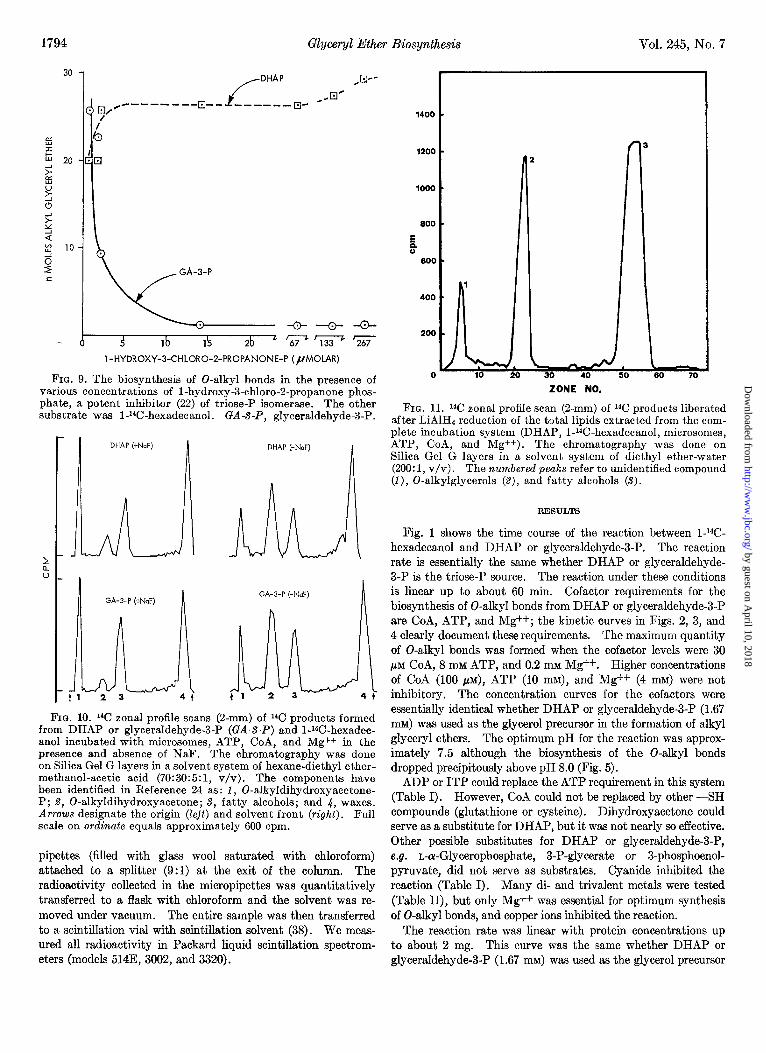
Glyceryl Ether Biosynthesis Vol. 245, No. 7
,/DHAP ,El--
* 0,
:
,I3
i
,e-m-------m-- k El’
--------me c-
2 lb 115 2b ‘167C’EF l-HYDROXY-3-CHLORO-2-PROPANONE-P (j!MOLAR)
FIG. 9. The biosynthesis of 0-alkyl bonds in the presence of various concentrations of I-hydroxy-3-chloro-2-propanone phos- phate, a potent inhibitor (22) of triose-P isomerase. The other substrate was l-l%-hexadecanol. GA-S-P, glyceraldehyde-3-P.
DHAP (tNoF)
l!.AA- DHAP (-NaF)
FIG. 10. 1% zonal profile scans (2-mm) of 14C products formed from DHAP or glyceraldehyde-3-P (GA-S-P) and 1-14C-hexadec- anol incubated with microsomes, ATP, CoA, and Mg++ in the presence and absence of NaF. The chromatography was done on Silica Gel G layers in a solvent system of hexane-diethyl ether- methanol-acetic acid (70: 30: 5: 1, v/v). The components have been identified in Reference 24 as: 1, O-alkyldihydroxyacetone- P; 2, 0-alkyldihydroxyacetone; 8, fatty alcohols; and 4, waxes. Arrows designate the origin (Zejt) and solvent front (right). Full scale on ordinate equals approximately 600 cpm.
pipettes (filled with glass wool saturated with chloroform) attached to a splitter (9: 1) at the exit of the column. The radioactivity collected in the micropipettes was quantitatively transferred to a flask with chloroform and the solvent was re- moved under vacuum. The entire sample was then transferred to a scintillation vial with scintillation solvent (38). We meas- ured all radioactivity in Packard liquid scintillation spectrom- eters (models 514E, 3002, and 3320).
1400
1200
1000
800
E 4
600
400
200
0 i- 60 70
ZONE NO.
FIG. 11. 1% zonal profile scan (2-mm) of 14C products liberated after LiAlH4 reduction of the total lipids extracted from the com- plete incubation system (DHAP, l-14C-hexadecanol, microsomes, ATP, CoA, and Mg++). The chromatography was done on Silica Gel G layers in a solvent system of diethyl ether-water (200: 1, v/v). The numbered peaks refer to unidentified compound (I), 0-alkylglycerols (2), and fatty alcohols (3).
RESULTS
Fig. 1 shows the time course of the reaction between I-14C- hexadecanol and DHAP or glyceraldehyde-3-P. The reaction rate is essentially the same whether DHAP or glyceraldehyde- 3-P is the triose-P source. The reaction under these conditions is linear up to about 60 min. Cofactor requirements for the biosynthesis of O-alkyl bonds from DHAP or glyceraldehyde-3-P are CoA, ATP, and Mg++; the kinetic curves in Figs. 2, 3, and 4 clearly document these requirements. The maximum quantity of 0-alkyl bonds was formed when the cofactor levels were 30 MM CoA, 8 mu ATP, and 0.2 mu Mg++. Higher concentrations of CoA (100 ELM), ATP (10 mM), and Mg++ (4 mM) were not inhibitory. The concentration curves for the cofactors were essentially identical whether DHAP or glyceraldehyde-3-P (1.67 mu) was used as the glycerol precursor in the formation of alkyl glyceryl ethers. The optimum pH for the reaction was approx- imately 7.5 although the biosynthesis of the 0-alkyl bonds dropped precipitously above pH 8.0 (Fig. 5).
ADP or ITP could replace the ATP requirement in this system (Table I). However, CoA could not be replaced by other -SH compounds (glutathione or cysteine). Dihydroxyacetone could serve as a substitute for DHAP, but it was not nearly so effective. Other possible substitutes for DHAP or glyceraldehyde-3-P, e.g. n-a-Glycerophosphate, 3-P-glycerate or 3-phosphoenol- pyruvate, did not serve as substrates. Cyanide inhibited the reaction (Table I). Many di- and trivalent metals were tested (Table II), but only Mg+f was essential for optimum synthesis of 0-alkyl bonds, and copper ions inhibited the reaction.
The reaction rate was linear with protein concentrations up to about 2 mg. This curve was the same whether DHAP or glyceraldehyde-3-P (1.67 mM) was used as the glycerol precursor
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of April 10, 1970 F. Snyder, B. Malone, and M. L. Blank 1795
.:O
II -70:
z
-60%
I;!
-5on
E
-40: u
16:O
G--J-
-30 :
-20
- IO
.O )
13 12 II IO 9 8 7 6 5 4 3 2 I 0 MIN
TUBE #3 56 CPM TUBE #I IICPM 1%
-80
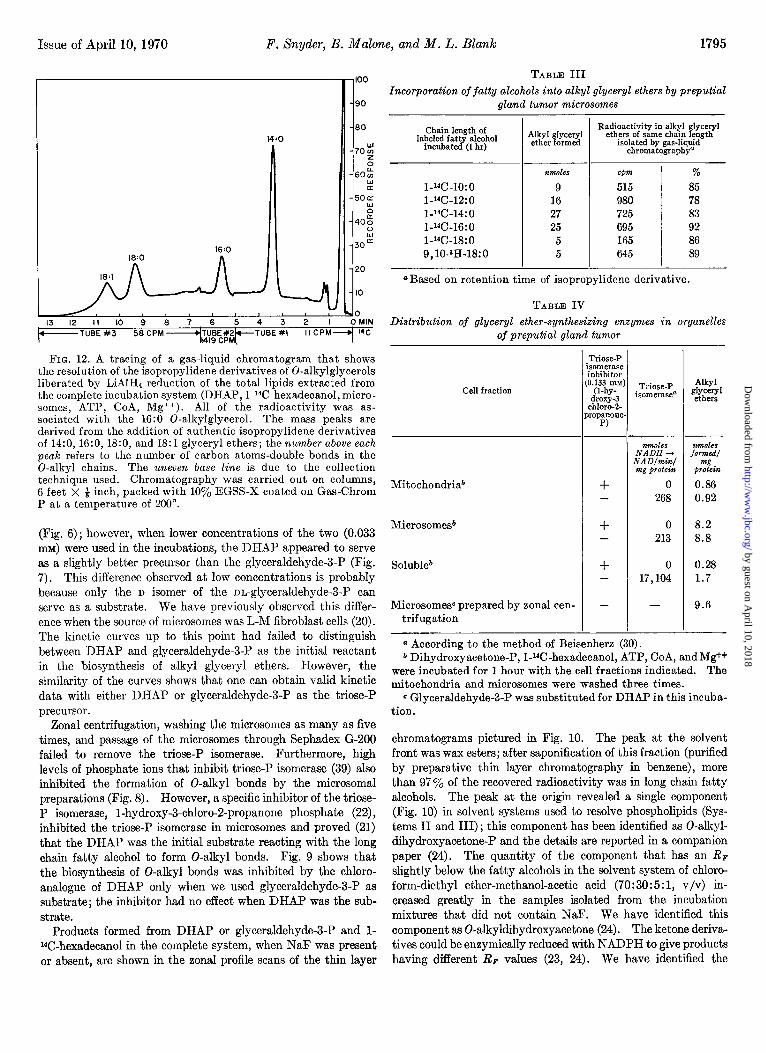
FIG. 12. A tracing of a gas-liquid chromatogram that shows the resolution of the isopropylidene derivatives of 0-alkylglycerols liberated by LiAIH4 reduction of the total lipids extracted from the complete incubation system (DHAP, l-14C-hexadecanol, micro- somes, ATP, CoA, Mg++). All of the radioactivity was as- sociated with the 16:0 0-alkylglycerol. The mass peaks are derived from the addition of authentic isopropylidene derivatives of 14:0, 16:0, 18:0, and 18:l glyceryl ethers; the number above each peak refers to the number of carbon atoms-double bonds in the 0-alkyl chains. The uneven base line is due to the collection technique used. Chromatography was carried out on columns, 6 feet X $ inch, packed with 10% EGSS-X coated on Gas-Chrom P at a temperature of 200”.
(Fig. 6) ; however, when lower concentrations of the two (0.033 mM) were used in the incubations, the DHAP appeared to serve as a slightly better precursor than the glyceraldehyde-3-P (Fig. 7). This difference observed at low concentrations is probably because only the D isomer of the nL-glyceraldehyde-3-P can serve as a substrate. We have previously observed this differ- ence when the source of microsomes was L-M fibroblast cells (20). The kinetic curves up to this point had failed to distinguish between DHAP and glyceraldehyde-3-P as the initial reactant in the biosynthesis of alkyl glyceryl ethers. However, the similarity of the curves shows that one can obtain valid kinetic data with either DHAP or glyceraldehyde-3-P as the triose-P precursor.
Zonal centrifugation, washing the microsomes as many as five times, and passage of the microsomes through Sephadex G-200 failed to remove the triose-P isomerase. Furthermore, high levels of phosphate ions that inhibit triose-P isomerase (39) also inhibited the formation of 0-alkyl bonds by the microsomal preparations (Fig. 8). However, a specific inhibitor of the triose- P isomerase, 1-hydroxy-3-chloro-2-propanone phosphate (22), inhibited the triose-P isomerase in microsomes and proved (21) that the DHAP was the initial substrate reacting with the long chain fatty alcohol to form 0-alkyl bonds. Fig. 9 shows that the biosynthesis of 0-alkyl bonds was inhibited by the chloro- analogue of DHAP only when we used glyceraldehyde-3-P as substrate; the inhibitor had no effect when DHAP was the sub- strate.
Products formed from DHAP or glyceraldehyde-3-P and l- *4C-hexadecanol in the complete system, when NaF was present or absent, are shown in the zonal profile scans of the thin layer
TABLE III
Incorporation of fatty alcohols into alkyl glyceryl ethers by preputial gland tumor microsomes
Chain length of labeled fatty alcohol
incubated (1 hr)
l-‘CL10: 0 i-"c-12:0 l-‘C-14: 0 l-‘*C-16:0 l-“C-18:0 9,10-ZH-18: 0
Alkyl glyceryl Radioactivity in alkyl glyceryl
ether formed ethers of same chain length
isolated by gas-liquid chromatography5
nmoles CM %
9 515 85 16 980 78 27 725 83 25 695 92 5 165 86 5 645 89
aBased on retention time of isopropylidene derivative.
TABLE IV
Distribution of glyceryl ether-synthesizing enzymes in organelles of preputial gland tumor
Cell fraction
Mitochondriab
Microsomesb
Solubleb
Microsomesc prepared by tonal cen- trifugation
ij
(t
p:
--
Alkyl ) Triose-P isomerase’ “%Z
_- nmoles nmoles
NADR -) jormed/ NAD/min/ mg $w&Gz *Zi:;gein
0 0.86 268 0.92
0 8.2 213 8.8
0 0.28 17,104 1.7
- 9.6
- - a According to the method of Beisenhere (30). b Dihydroxyacetone-P, 1-I%-hexadecanol, ATP, CoA, and Mg++
were incubated for 1 hour with the cell fractions indicated. The mitochondria and microsomes were washed three times.
0 Glyceraldehyde-3-P was substituted for DHAP in this incuba- tion.
chromatograms pictured in Fig. 10. The peak at the solvent front was wax esters; after saponification of this fraction (purified by preparative thin layer chromatography in benzene), more than 97% of the recovered radioactivity was in long chain fatty alcohols. The peak at the origin revealed a single component (Fig. 10) in solvent systems used to resolve phospholipids (Sys- tems II and III) ; this component has been identified as O-alkyl- dihydroxyacetone-P and the details are reported in a companion paper (24). The quantity of the component that has an Rp slightly below the fatty alcohols in the solvent system of chloro- form-diethyl ether-methanol-acetic acid (70:30:5: 1, v/v) in- creased greatly in the samples isolated from the incubation mixtures that did not contain NaF. We have identified this component as 0-alkyldihydroxyacetone (24). The ketone deriva- tives could be enzymically reduced with NADPH to give products having different Rp values (23, 24). We have identified the
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from
1796 Glyceryl Ether Biosynthesis Vol. 245, No. 7
products as 0-alkylglycerols (23, 24) after ensymic NADPH reduction of the 0-alkyldihydroxyacetone and as 0-alkyl acyl glycerophosphate after enzymic NADPH reduction of the O-al- kyldihydroxyacetone-P (23).
Total lipids obtained from the complete system containing DHAP or glyceraldehyde-3-P as the triose-P precursor were reduced with LiAlH4 to produce 0-alkylglycerols. The zonal profile scan of the thin layer chromatogram of LiAlHrreduced lipids obtained from the DHAP-incubated samples is pictured in Fig. 11. A minor component near the origin after LiAlH4 reduc- tion was unidentified. The 0-alkylglycerols were isolated under
FIQ. 13. The biosynthesis of 0-alkyl bonds as a function of the number of days after transplanting the preputial gland tumor. The substrates were 1-W-hexadecanol and glyceraldehyde-3-P. The activities are based on the incubation of the complete system containing 1.4 mg of protein incubated for 1 hour.
identical chromatographic conditions by preparative thin layer chromatography and the isopropylidene derivatives were pre- pared for gas-liquid chromatographic analyses. The isopro- pylidene could be resolved from other lipid classes present by thin layer chromatography (7). The formation of the isopro- pylidene derivative showed that the 0-alkyl bond was in position 1. The fact that only the 1 isomer of the glyceryl ether was labeled with 14C was further substantiated by thin layer chro- matography of the 0-alkylglycerols (liberated by LiAIII4 reduc-
TABLE V
Lipid class composition of preputial gland tumor9 grown in mice
Lipid class
Sterol esters. . _............ _... __ _. Waxes................................... Alk-1-enyl diacyl glyceryl ether. Alkyl diacyl glyceryl ether. . Triglycerides. . . . . Free fatty acids. . . . . . . . . Sterols + diglycerides. . . . . . . Alk-1-enyl ethanolamine-containing phos-
pholipid . . . . . . . . . . . . . . . . . . Alkyl ethanolamine-containing phospho-
lipid................................... Diacyl ethanolamine-containing phospho-
lipid................................... Alk-1-enyl choline-containing phospholipid Alkyl choline-containing phospholipid. . Diacyl choline-containing phospholipid. Other phospholipids. . . .
a Three weeks after transplantation.
- I-
-
Weight
%
28.8b Trace
0.7 2.2
31.1 3.4 7.3
2.26
0.6”
5.0 0.2c O.gc
12.9 5.0
* Gas-liquid chromatography revealed at least four other sterols in addition to cholesterol in this fraction.
c Determined after LiAlH4 reduction of purified fractions of the choline- and ethanolamine-containing phospholipids.
TABLE VI
Distribution (weight percent) of acyl, alkyl, and alk-1 -enyl hydrocarbon moieties in lipid classes of preputial gland tumors in mice
Chain length less than
14:o Trace Trace 14:o 2.5 4.7 14: 1 Trace 3.4 15:o Trace 6.0 16:0 25.0 15.6 16:l 7.0 9.8 17:o 0.4 0.6 18:0 6.0 6.0 18: 1 37.4 26.4 18:2 17.8 6.3 20: 0 0.4 2.3
18: 3 or 20: 1 1.7 2.9 20:3 or 22:0 Trace 0.8 20:4 or 22: 1 1.8 11.4
22:xc Trace 2.4
-
_-
-
Trifyc$ide a
_-
-
Diacyl glyceryl ether -
--
-
WkYU
Trace 3.9 0.7 1.2
22.8 5.0 1.8
12.7 21.2
3.9 1.7 4.5 1.3 7.6
11.7
.-
-
- Diacyl glyceryl
ether (alk- l-enyl HP)
Trace 3.8
3.8 39.0
12.8 28.2
2.7b
6.5
3.2
Ethanolamine- containing
phospholipid (XYl)
0.4 Trace
8.6 3.3
12.2 25.2
5.5 0.7 0.7 1.9
21.2 20.3
-
I
-
~thanolamine- containing
phospholipid (alk-1-enyl)
1.1 Trace
46.5 7.9
20.1 21.8 2.6
Trace
Trace
-
1
--
-
Sthanolamine- containing
phospholipld bkY 1)
Trace Trace
37.4 4.9
25.9 26.2
1.7 1.8
Trace Trace
2.1
-L :holine-containing phospholipid -
_-
-
(Ad
0.6 Trace
28.2 5.3
10.5 27.6 8.3 1.0 0.4 2.5
12.7 2.9
-
_-
-
(Akrl)
2.1 Trace Trace 35.7
3.4
9.0 47.1
2.7 Trace
0 (Ho) indicates that these samples were hydrogenated with Adams catalyst before analyses (16). * Identified as 19:O. c 22:X represents chain lengths with more than one double bond.
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of April 10, 1970 F. Snyder, B. Malone, and M. L. Blank 1797
tion of total lipids) on arsenite-impregnated layers (35). Perio- date oxidation of the ether-linked products also verified that only the 1 isomer was formed (23, 24). The tracing of the gas-liquid chromatogram (Fig. 12) shows the resolution of mass and radio- activity as based on the retention time of known isopropylidene derivatives of 0-alkylglycerols. Table III gives further analysis of the 0-alkylglycerols after the incubation of various chain lengths of fatty alcohols. The chain length of the radioactive 0-alkyl moiety in the isopropylidene derivatives of the O-alkyl- glycerols resolved by gas-liquid chromatography was always identical with the chain length of the l-14C- or 9,10-3H-fatty alcohols used as precursors.
Activity in the formation of 0-alkyl bonds is highest in the microsomes (Table IV) and only DHAP was utilized when 1-hydroxy-3-chloro-2-propanone phosphate was used as an inhibitor of triose-P isomerase in the subcellular fractions. Tri- ose-P isomerase was present in all subcellular fractions tested and it was inhibited by the chloro-analogue of DHAP in each instance (Table IV). The presence of triose-P isomerase in microsomal (21, 40) and mitochondrial (41) fractions has been reported earlier. Microsomes prepared by zonal centrifugation (26) also contained the enzymes that could synthesize 0-alkyl bonds. The zonal centrifugation profiles revealed that the preputial gland tumors, like certain other neoplasms, contained mainly rough endoplasmic reticulum (42).
Formation of 0-alkyl bonds in glycerolipids of the preputial gland tumors is dependent upon the age of the tumor (Fig. 13). The highest activities were observed during the early stages of growth when the tumor first became large enough for excision (2 to 4 g). The ether-synthesizing activity progressively de- creased with time after transplantation of the tumor. On some occasions, the microsomes from older tumor preparations did not synthesize 0-alkyl ethers; this finding may be related to the degree of necrosis.
The lipid composition of whole tumors (3 weeks after trans- plantation) is shown in Table V. Significant quantities of alkyl glyceryl ethers are present, mainly as the diacyl type (-2%). The major lipid classes consist of sterol esters, triglycerides, free sterols, and phosphatidylcholine. The 0-alkyl, 0-alk-1-enyl, and acyl moieties of the triglycerides, diacyl glyceryl ethers, choline phosphatides, and ethanolamine phosphatides are shown in Table VI. The 0-alkyl and 0-alk-1-enyl moieties of each lipid class consist mainly of 16:0, 18:0, and 18: 1 carbon chains. Since the acyl moieties in these experiments are derived from both position 1 and 2, a direct comparison with 0-alkyl moieties is not possible. However, the total acyl moieties of the same classes contain considerable quantities of CZO and C&Z chains and also exhibit a much higher degree of unsaturation, namely 18:2, 20:4, and 22:X chains.
DISCUSSION
Enzyme systems that produce 0-alkyl bonds have been pre- pared from both normal (19, 20) and Ehrlich ascites cells (23) as well as preputial gland tumors. Kapoulas and Thompson (43) and Friedberg (44) have reported similar findings in Tetra- hymena pyriformis, and Hajra (45) has reported an enzyme system that synthesizes 0-alkyl bonds in liver mitochondria of guinea pigs and in microsomes of mouse brain. In general, these systems that contain 0-alkyl-synthesizing enzymes have high quantities of ether-linked lipids (15, 46).
The identification of labeled 0-alkyldihydroxyacetone-P and
0-alkyldihydroxyacetone, suggested as intermediates in this system in our first paper (8) on the subject, has been documented in the preputial gland tumor (24) and in Ehrlich ascites cells (23). In Ehrlich ascites cells, 14C-, aH-, and “P-labeled DHAP and 14C- and aH-labeled fatty alcohols are incorporated into inter- mediates and 0-alkylglycerols in a 1: 1 molar ratio (23). 1J4C- l-aH-Labeled hexadecanol does not change its aH :I%! ratio when it is incorporated into the 0-alkyl chain (23), showing that the fatty alcohol chains are indeed incorporated as an intact unit. The formation of waxes from 14C-labeled fatty alcohols in cell- free systems that normally do not contain significant quantities of waxes has been noted before for homogenates of normal tissues (47,48).
We still do not know the mechanism of the initial step in the formation of 0-alkyl linkages, but, in view of the cofactor require- ments, it is possible to speculate. One possibility is that the ene-diol form of DHAP might enter the reaction as an activated CoA derivative. The fatty alcohols in the system could also exist as activated forms (AMP derivatives). The thiohemiacetal as depicted would be highly reactive. Current studies in our laboratories are directed toward exploring the feasibility of the proposed mechanisms:
OH Hz OH H OH
Y. fi HC-SCoA
F= +-+ OH
H&-O@ x
ATP, CoA-SII, Mg++ ’ A H OH
Hz -08 Ha a@ A
OH
H -SCoA
i: H OH + [ROH + Mg++ + ATP + ROAMP i- PPJ 3
H&O@
YH Hk-OR
H&OH + AMP + CoA-SH
H&O@
HO%OR HC-OR H) -OR -II,0
HOH F
i
A OH +
Hz -O@ Ha -08 i: 8
=0
H,C--O@
MECHANISM I
HXOH
i:
H&-SCoA
==O + CoA-SH ;z: ) +O
ECZCYA 4------ ;I;
s
2
OH + ROH -
Hz 48 F
=o + CoA-SH
Hz 48
MECHANISM II
Such mechanisms would permit the formation of a second ether bond in position 2 of glyceryl ethers in halophilic bacteria (49, 50) ; i.e. the dephosphorylated ketone intermediate could form an enol structure for a repetitive series of reactions as shown
(I and II). Our data (Table I) show that a reaction with dihy-
droxyacetone is feasible, but it was a poor acceptor and the
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1798 Glyceryl Ether Biosynthesis Vol. 245, No. 7
possibility remains that phosphorylation had occurred before it was used as a substrate.
A third mechanism for the biosynthesis of 0-alkyl bonds is that the DHAP might be necessary to attach to the enzyme, and that while it is enzyme-bound it is converted to glyceralde- hyde-3-P, which could then form a thiohemiacetal with CoA before conversion to the 0-alkyl compounds as initially proposed (8). The involvement of the ketone compounds in the various reaction sequences also permits proper stereochemical orientation of the hydroxyl group or second 0-alkyl moiety in the naturally occurring lipids.
Phosphoenolpyruvate did not serve as a substrate in the for- mation of alkyl glyceryl ethers, yet it is well established as a precursor of ether bonds in a number of other compounds (51-53). The enol form of DHAP in the mechanism that we propose thus relates one common feature that might be necessary for formation of ether bonds in diverse compounds.
An intriguing hypothesis is that the 0-alkyl moieties might be precursors of the plasmalogens as Thompson (54) originally proposed. Thompson (54) discovered that the slug converts *H ,14C-labeled 0-alkylglycerols to plasmalogens ; the plasmalo- gens had the same 3H:14C ratio as the precursor. Furthermore, Keenan, Brown, and Marks (lo), Friedberg and Greene (13), and Wood and Healy (55) have demonstrated that long chain fatty alcohols can be incorporated into plasmalogens better than fatty acids or fatty aldehydes. Experimental data obtained with 14C-ethanolamine (56) have also been compatible with Thomp- son’s observation (54), and this is strengthened by the similarity between the 0-alkyl chains and the 0-alk-1-enyl chains found in the preputial gland tumor and other neoplasms (5).
At least two possible reactions (III or IV) could be envisioned for the conversion of 0-alkyl to 0-alk-1-enyl moieties.
Hz -OCH,R
8
Reductase
=O(OH or acyl) / \ * XP xmItFI+ (III)
H&!-O@ (OH, P-choline, or P-ethanola- mine)
H&O@(OH, choline, or ethanola- mine)
The XP represents a hydrogen acceptor such as NAD, NADP, or FAD.
H,y-OCHpR
HY
I =O(OH
8
or acyl) + b=O(OH or acyl)
HZ -O@ (OH, P-choline, or Hz cl3 -O@(OH, P-choline, P-ethanolamine) or P-ethanolamine)
HH (IV)
+ HY
P-ethanolamine)
The substituent “Y” on the 0-alkyl moiety in Reaction IV could be an OH, -OCH, -OCH&H3, NH2, SH or similar moiety. Such interconversions, however, have not been shown in a cell- free system, and we have not detected any plasmalogens in our microsomal system under the conditions of incubation used, but we have recently found an unidentified compound that has
chromatographic properties similar to /3-hydroxy-0-alkylglycerol (57, 58) in the microsomal system described and also in z&o in Ehrlich ascites cells after administering aH,14C-chimyl alcohol. Direct hydroxylation of alkyl ether moieties is thought to occur in the cleavage of glyceryl ethers (59), and a similar mechanism could be involved in the biosynthesis of plasmalogens.
Our negative results on the formation of plasmalogens in this system may be related to the need for other cofactors or con- ditions for the conversion of 0-alkyl to 0-alk-1-enyl linkages. The similarities of 0-alkyl and 0-alk-1-enyl moieties could also be explained by interconversions of the aliphatic moieties that serve as precursors. Currently, little is known about the inter- conversions of fatty acids, fatty aldehydes, and fatty alcohols, but some data on these interconversions have been obtained (25, 59) from investigations on the biocleavage of glyceryl ethers.
0-Alkyl linkages in preputial gland tumors are formed by a new metabolic pathway (DHAP and fatty alcohols) unrelated to the well known a-glycerophosphate and acyl-CoA pathway for acylated glycerolipids (60). It is the first cell-free system known to make 0-alkyl bonds in lipids.
Acknowledgments-We want to thank Dr. Ten-thing Lee for help with the zonal centrifugation procedure and Edgar A. Cress and Nelson Stephens for setting up some of the incubation studies reported. We also express our gratitude to Dr. Fred Hartman for supplying us with triose-P isomerase inhibitor.
REFERENCES
1. SNYDER, F., in R. T. HOLMAN (Editor), Progress in the chem- istry of fats and other lipids, Vol. X, Pergamon Press, Ox- ford, 1969, p. 287.
2. THOMPSON, G. A., JR., AND KAPOULAS, V. M., in J. M. LOWEN- STEIN (Editor); Methods in enzymology, vol. 14, Academic Press. New York. 1969.~~668.
3. SNYDER, F., in G. PATA& AND A. NIEDERWIESER (Editors), Progress in thin-layer chromatography and related methods, Ann Arbor-Humphrey Science Publishers, Inc., Ann Arbor, in press.
4. MANGOLD, H. K., AND BAUMANN, W. J., in G. V. MARINETTI (Editor), Lipid chromatographic analysis, Vol. 1, Marcel Dekker, Inc., New York, 1967, p. 339.
5. SNYDER, I?., in w. L. HOLMES, L. k: CARLSON, AND R. PAOLETTI (Editors), Advances in experimental medicine and biology, VoZ. 4, Plenum Publishing Corporation, New York, 1969, p. 609.
6. TSUJIMOTO, M., AND TOYAMA, Y., Chem. Umschau, 29, 27 (1922).
7. SNYDER, F.,MALONE, B., AND WYKLE, R. L., Biochem. Biophys. Res. Commun., 34, 40 (1969).
8. SNYDER, F.. WYKLE, R. L.. AND MALONE. B., Biochem. Biophys. Res. dbimun., 34, 315 (1969). ’
_ -
9. SNYDER. F.. WYKLE. R. L.. AND MALONE. B.. Fed. Proc.. 28. 595 (i969j. ' '
, , I I
10. KEENAN, R. W., BROWN, J. B., AND MARKS, B. H., Biochim. Biophys. Acta, 61, 226 (1961).
11. THOMPSON, G. A.. JR.. J. Biol. Chem., 240, 1912 (1965). 12. MALINS, D. C., Biochem. J., 101, 39P’(1966). 13. FRIEDBERG. S. J.. AND GREENE. R. C.. J. Biol. Chem.. 242.
5709 (196j). ’ I .
14. ELLINGBOE, J., AND KARNOVSKY, M. L., J. Biol. Chem., 242, 5693 (1967).
15. SNYDER, F., AND WOOD, R., Cancer Res., 28, 972 (1968). 16. SNYDER, F., AND BLANK, M. L., Arch. Biochem. Biophys., 130,
101 (1969). 17. BLANK, M. L., AND SNYDER, F., Lipids, in press. 18. FRIEDBERG, S. J., AND GREENE, R. C., Biochim. Biophys.
Acta, 164, 602 (1968). 19. SNYDER, F., MALONE, B., AND BLANK, M. L., Biochim. Biophys.
Acta, 187, 302 (1969).
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of April 10, 1970 F. Snyder, B. Malone, ancl M. L. Blank 1799
20. SNYDER, F., MALONE, B., AND GUMMING, R. B., Can. J. Bio- 41. BOXER, G. E., AND SHONK, C. E., Biochim. Biophys. Acta, 37, them., 48, 212 (1970). 194 (1960).
21. WYKLE, 1~. L., AND SNYDER, F., Biochem. Biophys. Res. Com- mun., 37, 658 (1969).
22. HARTMAN, F. C., Biochem. Biophys. Res. Commun., 33, 888 (1968).
23. 24.
WYKLE, R. L., AND SNYDER, F., J. Biol. Chem., in press. SNYDER, F., BLANK, M. L.,‘MA~oNE, B., AND WY<LE, R. L.,
J. Biol. Chem.. 245. 1800 (1970).
42. LEE, T-C., SWARTZENDRUBER, D. C., MORRIS, H. P., AND SNYDER, F., in Proceedings of the Tenth International Cancer Congress, Houston, Texas, May 1970, in press.
43. KAPOULAS, V. M., AND THOMPSON, G. A., JR., Biochim. Biophys. Acta, 187, 594 (1969).
25. PFLEGIZR, R. C.,’ PIA~TADO&, C:, AND SNYDER, F., Biochim.
44. FRIEDBERG, S. J., AND GREENE, R. C., J. Amer. Oil Chem. Sot., 46, Abstract 82 (1969).
26. Biophyi. Acta; 144, 633 (1967).
LEE. T-C.. SWARTZENDRUBER. D. C.. AND SNYDER. F.. Bio-
27. chkm. Biophys. Res. Commun., 36, 748 (1969). ’ ’
OSWALD, E. O., PIANTADOSI, C., ANDERSON, C. E., AND SNYDER F., Lipids, 1, 241 (1966).
45. HAJRA, A. K., Biochem. Biophys. Res. Commun., 37,486 (1969). 46. THOMPSON, G. A., JR., Biochemistry, 6, 2015 (1967). 47. FRIEDBERG, S. J., AND GREENE, R. C., J. Biol. Chem., 242,
234 (1967).
28. BLIGH, E. G., AND DYER, W. J., Can. J. Biochem. Physiol., 37, 911 (1959).
29.
30.
LOTVRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL, R. J., J. Biol. Chem., 193, 265 (1951).
BEISENHERZ, G., in S. P. COLOWICIC AND N. 0. KAPLAN (Edi- tors), Methods in enzymology, Vol. 1, Academic Press, New York, 1955, p. 387.
48. SNYDER, F., MALONE, B., AND SOODSMA, J., J. Amer. Oil Chem. Sot., 46, No. 2, Abstract 9 (1969).
49. SEHGAL, S. N., KATES, M., AND GIBBONS, N. E., Can. J. Biochem. Physiol., 40, 69 (1962).
50. KATES, M., SASTRY, P. S., AND YENGOYAN, L. S., Biochim. Biophys. Acta, 70, 705 (1963).
51. STROMINGER, J. L., Biochim. Biophys. Acta, 30, 645 (1968). 52. LEVIN, J. G., AND SPRINSON, D. B., J. Biol. Chem., 239, 1142
(1964). 31. WOOD, R., AND SNYDER, F., Lipids, 3, 129 (1968). 53. GUNETILEKE, K. G., .~ND ANWAR, R. A., J. Biol. Chem., 243, 32. HANAHAN, D. J., EKHOLM, J., AND JACKSON, C. M., Biochem- 5770 (1968).
istry, 2, 630 (1963). 33. WOOD, It., Lipids, 2, 199 (1967). 34. SNYDER, F., AND CRESS, E. A., Radiat. Res., 19, 129 (1963). 35. WOOD, R., AND SNYDER, F., Lipids, 2, 161 (1967). 36. PRIVETT, 0. S., BLANK, M. L., CODDING, D. W., AND NICKELL,
54. THOMPSON, G. A., JR., Biochim. Biophys. Acta, 152, 409 (1968). 55. WOOD, R., AND HEALY, K., Biochem. Biophys. Res. Commun.,
38, 205 (1970).
E. C., J. Amer. Oil Chem. Sot., 42, 381 (1965). SNYDER, F., AND KIMBLE, H., Anal. Biochem., 11, 510 (1965). SNYDER, F., AND SMITH, I)., Separ. Sci., 1, 709 (1966). OESPER, P., AND MEYERHOF, O., Arch. Biochem., 27,223 (1950). RAO, G. A., SORRELS, M. F., AND REISER, R., Biochem.Biophys.
Res. Commun., 31, 252 (1968).
56. HORROCKS, L. A., AND ANSELL, G. B., Lipids, 2, 329 (1967). 57. WYKLE, R. L., BLANK, M. L., PIANTSDOSI, C., AND SNYDER,
F., Fed. Proc., 29, 674 (1970). 37. 38. 39. 40.
58. BLANK, M. L., WYKLE, R. L., PIANTADOSI, C., AND SNYDER, F., &oc. Amer. Ass..Cancei Res., 11, 9 (1970).
59. TIETZ. A.. LINDRERG. M.. AND KENNEDY. E. P.. J. Biol. Chem.. 239,‘40dl (1964). ’ ’
, I
60. KENNEDY, E. P., Fed. Proc., 16, 847 (1957).
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Fred Snyder, Boyd Malone and M. L. Blank-Alkyl Bonds in GlycerolipidsOEnzymic Synthesis of
1970, 245:1790-1799.J. Biol. Chem.
http://www.jbc.org/content/245/7/1790Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/245/7/1790.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























