Embed Size (px)
Citation preview

73
6
K. E. Havens ()Florida Sea Grant and University of Florida School of Forest Resources and Conservation, Gainesville, 32611 FL, USAe-mail: [email protected]
Abstract
The plankton food web comprises microscopic organisms that transport carbon and energy from producers to consumers in the water column of aquatic ecosystems. In lakes, the food web includes bacteria, phytoplankton, protozoa, micro- and macrozooplankton, and the tro-phic links between them. Structure of the plankton food web in lakes is affected in a predict-able manner by eutrophication: phytoplankton biomass becomes relatively greater, while macrozooplankton biomass declines. The relative biomass of protozoa also may increase. Food web function also is affected: ecological transfer efficiency is lowest in ultraoligo-trophic lakes and hypereutrophic lakes and highest in mesotrophic lakes. The relationship between the size of producers and grazers drives this unimodal pattern. In ultraoligotrophic lakes, cooccurrence of picoplankton with copepods that cannot directly graze such small particles results in long, energetically inefficient food webs with multiple steps includ-ing protozoa and microzooplankton. In hypereutrophic lakes, similar food webs occur, but owing to cooccurrence of cyanobacteria and small macrozooplankton that cannot handle those large and sometimes toxic phytoplankton. The dominance of large cyanobacteria in hypereutrophic lakes can be directly linked to enrichment with nutrients, particularly nitro-gen and phosphorus, while the small size of zooplankton is attributed most often to intense fish predation. Long-term studies indicate that the changes in plankton food web structure and function can be reversed if nutrient loads are substantially reduced.
Keywords
Eutrophication · Plankton food web · Lake management · Fish · Plants
eutrophication, is most often owing to an overenrichment of lakes with phosphorus (P) and/or nitrogen (N; Conley et al. 2009) and recent evidence suggests that its symptoms may be exacerbated by climate change (Moss et al. 2011) making the rehabilitation of culturally eutrophic lakes an even great-er challenge in the future. One impact of eutrophication on lakes is a change in the structure and function of their food webs. The aim of this chapter is to provide a concise over-view of how cultural eutrophication affects plankton food webs, drawing on research from the 1960s through studies completed in recent years. The chapter is divided into the
A. A. Ansari, S. S. Gill (eds.), Eutrophication: Causes, Consequences and Control, DOI 10.1007/978-94-007-7814-6_6, © Springer Science+Business Media Dordrecht 2014
Lake Eutrophication and Plankton Food Webs
Karl E. Havens
6.1 Introduction
Thousands of lakes around the world have been impacted by excessive inputs of nutrients from human-related uses of the land, and as a result have experienced changes in their ecological structure and function. This phenomenon, cultural

74 K. E. Havens
following sections: a description of the plankton food web; an evaluation of data regarding how biomass distribution among components of the plankton food web changes along a gradient from nutrient-poor to nutrient-rich lakes; a con-sideration of how changes in the structure of plankton food webs affects their function in regard to their ability to effec-tively transfer energy to fish and other biota at higher trophic levels; a brief discussion of how other changes in the lake ecosystem influence how the plankton food web responds to eutrophication; and implications for the management of culturally eutrophic lakes.
6.2 The Plankton Food Web
Limnologists’ view of the plankton food web has changed considerably since the mid-1900s when it was considered as a chain where phytoplankton fixed carbon that was then transferred directly to zooplankton and then to fish and other predators. This traditional view was supplemented by the concept of additional microbial pathways in the 1980s (Azam et al. 1983) and it is now widely recognized that bacteria and protozoa also play an important role in energy transfer and nutrient cycling in the plankton. Five major components and nine major links characterize the contemporary model of the plankton food web (Fig. 6.1). Phytoplankton fix car-bon by photosynthesis and are consumed by a variety of dif-ferent types of protozoa (including ciliates and flagellates) by microzooplankton such as rotifers and immature stages (nauplii) of copepods, and by macrozooplankton (cladocer-ans and copepods). A second route of energy flow occurs from protozoa to microzooplankton and macrozooplankton. Bacteria metabolize organic carbon that enters the lake from outside sources as well as carbon excreted into the water by phytoplankton, protozoa, and zooplankton. Bacteria are a food source for protozoa, microzooplankton, and macrozoo-plankton.
The Dashed arrows in Fig. 6.1 roughly correspond to energy flows in the traditionally recognized grazing food chain links in the web and the solid arrows to the microbial food web links. Although quantitative evidence regarding the importance of microbial links emerged in the 1980s (Azam et al. 1983; Sherr et al. 1987; Porter et al. 1988), their importance was conceptually identified by Linde-man (1942) who developed an energy flow diagram that included bacteria and links to and from phytoplankton, zooplankton, and dissolved materials. Gliwicz (1969) first speculated that the importance of microbial links may in-crease with eutrophication, a prediction that was proven to be correct based on results of later studies (Munawar et al. 2011).
6.3 The Structure of Plankton Food Webs Versus Eutrophication
One typical and highly visible symptom of cultural eutrophi-cation is increased biomass and dominance of large inedible cyanobacteria (e.g., Smith et al. 2006). Coincidentally, there are changes in the biomass and composition of zooplank-ton—including a loss of large species of Daphnia, and in-creased dominance of small rotifers and copepods (Weisse and Stockner 1992; Havens et al. 1996). Auer et al. (2004) performed a comprehensive analysis of plankton food web structure along a eutrophication gradient, comparing plank-ton communities in 55 lakes in northern Germany that ranged from mesotrophic to hypereutrophic. All the components of the food web displayed an increase in their biomass with greater levels of enrichment, indexed by chlorophyll con-centration, although the rates of increase varied (Fig. 6.2). As a result, the proportional biomass changed: mesotrophic lakes had about 30 % of the plankton biomass in zooplankton and about 45 % in phytoplankton (Fig. 6.3a). Lesser amounts of biomass occurred in bacteria, heterotrophic flagellates, and ciliates. Over the gradient of enrichment, relative bio-mass of zooplankton dropped, to a low of around 15 % in
Fig. 6.1 A diagram of the plankton food web showing major compo-nents and links between them. Solid arrows correspond to links in the traditional grazing food chain and dashed arrows correspond to links in the microbial food web. Numerous species comprise each of the com-ponents shown here—individual species are shown as examples only
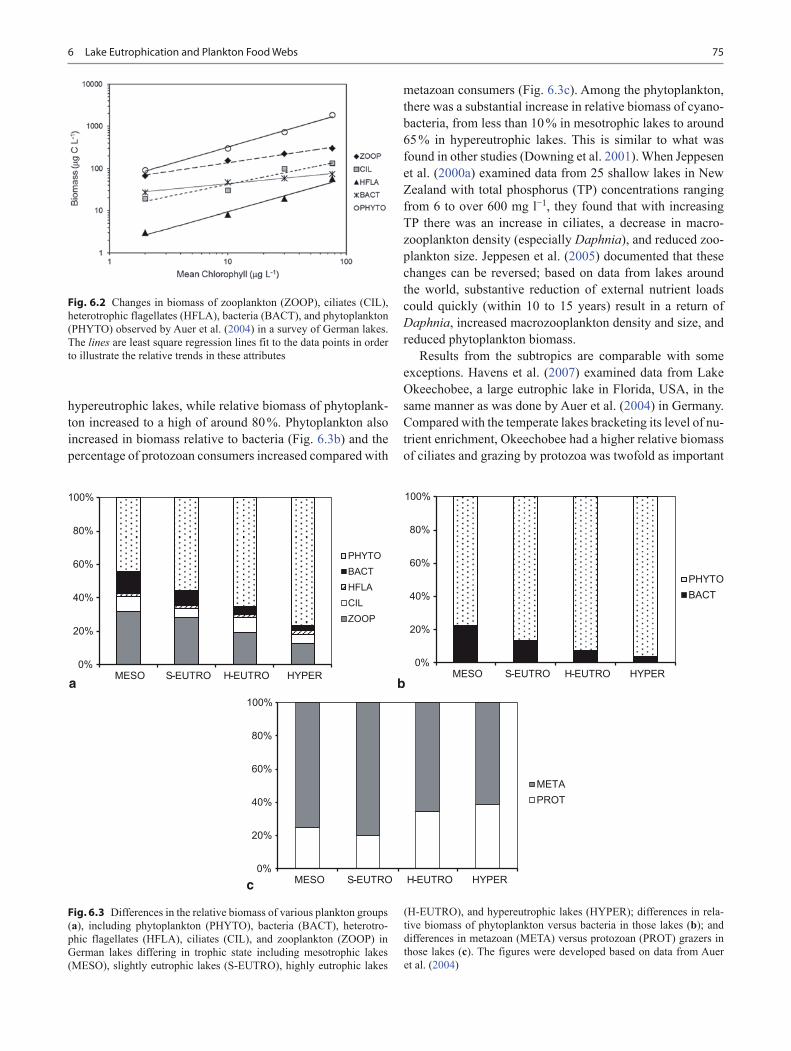
756 Lake Eutrophication and Plankton Food Webs
hypereutrophic lakes, while relative biomass of phytoplank-ton increased to a high of around 80 %. Phytoplankton also increased in biomass relative to bacteria (Fig. 6.3b) and the percentage of protozoan consumers increased compared with
metazoan consumers (Fig. 6.3c). Among the phytoplankton, there was a substantial increase in relative biomass of cyano-bacteria, from less than 10 % in mesotrophic lakes to around 65 % in hypereutrophic lakes. This is similar to what was found in other studies (Downing et al. 2001). When Jeppesen et al. (2000a) examined data from 25 shallow lakes in New Zealand with total phosphorus (TP) concentrations ranging from 6 to over 600 mg l−1, they found that with increasing TP there was an increase in ciliates, a decrease in macro-zooplankton density (especially Daphnia), and reduced zoo-plankton size. Jeppesen et al. (2005) documented that these changes can be reversed; based on data from lakes around the world, substantive reduction of external nutrient loads could quickly (within 10 to 15 years) result in a return of Daphnia, increased macrozooplankton density and size, and reduced phytoplankton biomass.
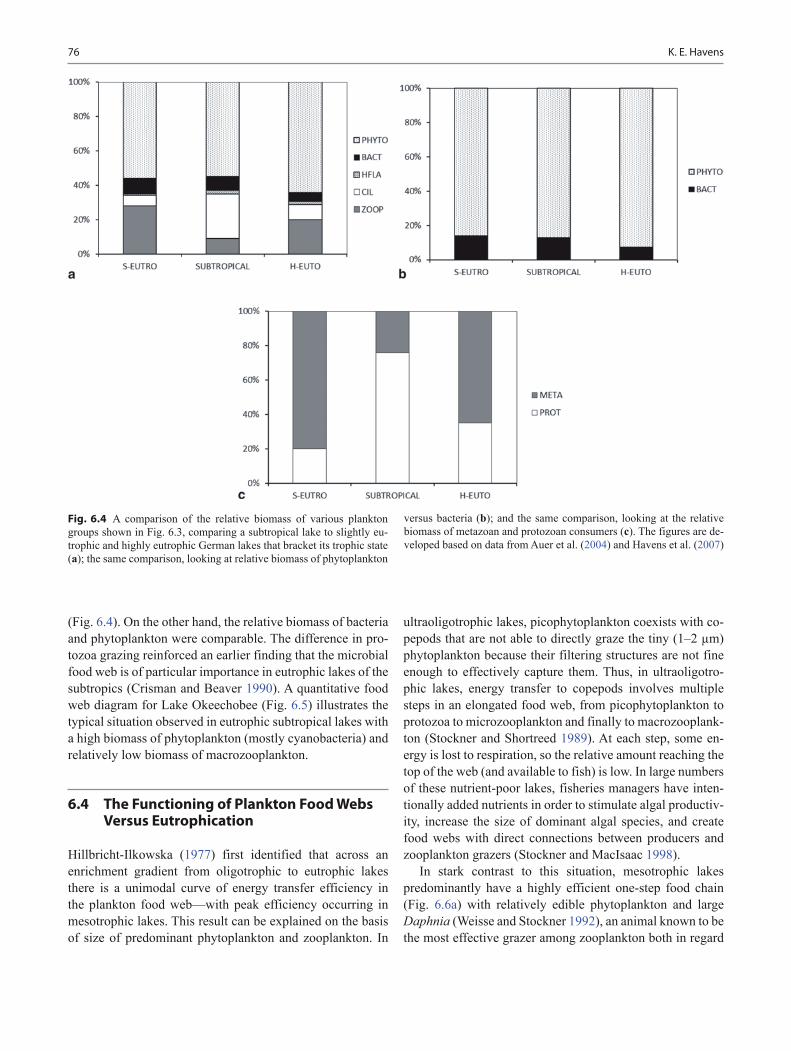
Results from the subtropics are comparable with some exceptions. Havens et al. (2007) examined data from Lake Okeechobee, a large eutrophic lake in Florida, USA, in the same manner as was done by Auer et al. (2004) in Germany. Compared with the temperate lakes bracketing its level of nu-trient enrichment, Okeechobee had a higher relative biomass of ciliates and grazing by protozoa was twofold as important
a b
c
Fig. 6.3 Differences in the relative biomass of various plankton groups (a), including phytoplankton (PHYTO), bacteria (BACT), heterotro-phic flagellates (HFLA), ciliates (CIL), and zooplankton (ZOOP) in German lakes differing in trophic state including mesotrophic lakes (MESO), slightly eutrophic lakes (S-EUTRO), highly eutrophic lakes
(H-EUTRO), and hypereutrophic lakes (HYPER); differences in rela-tive biomass of phytoplankton versus bacteria in those lakes (b); and differences in metazoan (META) versus protozoan (PROT) grazers in those lakes (c). The figures were developed based on data from Auer et al. (2004)
Fig. 6.2 Changes in biomass of zooplankton (ZOOP), ciliates (CIL), heterotrophic flagellates (HFLA), bacteria (BACT), and phytoplankton (PHYTO) observed by Auer et al. (2004) in a survey of German lakes. The lines are least square regression lines fit to the data points in order to illustrate the relative trends in these attributes
76 K. E. Havens
(Fig. 6.4). On the other hand, the relative biomass of bacteria and phytoplankton were comparable. The difference in pro-tozoa grazing reinforced an earlier finding that the microbial food web is of particular importance in eutrophic lakes of the subtropics (Crisman and Beaver 1990). A quantitative food web diagram for Lake Okeechobee (Fig. 6.5) illustrates the typical situation observed in eutrophic subtropical lakes with a high biomass of phytoplankton (mostly cyanobacteria) and relatively low biomass of macrozooplankton.
6.4 The Functioning of Plankton Food Webs Versus Eutrophication
Hillbricht-Ilkowska (1977) first identified that across an enrichment gradient from oligotrophic to eutrophic lakes there is a unimodal curve of energy transfer efficiency in the plankton food web—with peak efficiency occurring in mesotrophic lakes. This result can be explained on the basis of size of predominant phytoplankton and zooplankton. In
ultraoligotrophic lakes, picophytoplankton coexists with co-pepods that are not able to directly graze the tiny (1–2 µm) phytoplankton because their filtering structures are not fine enough to effectively capture them. Thus, in ultraoligotro-phic lakes, energy transfer to copepods involves multiple steps in an elongated food web, from picophytoplankton to protozoa to microzooplankton and finally to macrozooplank-ton (Stockner and Shortreed 1989). At each step, some en-ergy is lost to respiration, so the relative amount reaching the top of the web (and available to fish) is low. In large numbers of these nutrient-poor lakes, fisheries managers have inten-tionally added nutrients in order to stimulate algal productiv-ity, increase the size of dominant algal species, and create food webs with direct connections between producers and zooplankton grazers (Stockner and MacIsaac 1998).
In stark contrast to this situation, mesotrophic lakes predominantly have a highly efficient one-step food chain (Fig. 6.6a) with relatively edible phytoplankton and large Daphnia (Weisse and Stockner 1992), an animal known to be the most effective grazer among zooplankton both in regard
Fig. 6.4 A comparison of the relative biomass of various plankton groups shown in Fig. 6.3, comparing a subtropical lake to slightly eu-trophic and highly eutrophic German lakes that bracket its trophic state (a); the same comparison, looking at relative biomass of phytoplankton
versus bacteria (b); and the same comparison, looking at the relative biomass of metazoan and protozoan consumers (c). The figures are de-veloped based on data from Auer et al. (2004) and Havens et al. (2007)

776 Lake Eutrophication and Plankton Food Webs
to its quantity of grazing and wide size range of grazed par-ticles. In hypereutrophic lakes, size again interferes with direct energy transfer in the grazing chain. Large cyanobac-teria such as Anabaena coexist with small zooplankton such as Bosmina that cannot eat those phytoplankton (Fig. 6.6b). The cyanobacteria excrete carbon that is taken up by bac-teria and then is routed up through a microbial web to the zooplankton.
Quantitative data documenting the unimodal pattern in food web efficiency were presented by Jeppesen et al. (2003) based on a comprehensive survey of 466 lakes spanning the temperate to arctic zones and encompassing a broad range of trophic states. Other studies documenting the predominance of a microbial food web in eutrophic lakes include Riemann and Sondergaard (1986), who compared the plankton in two lakes differing in degree of nutrient enrichment, and Cole et al. (1988), who compared the plankton in 24 lakes across an eutrophication gradient.
Because complex food webs lead to greater energy loss compared with one-step chains where Daphnia directly graze phytoplankton, eutrophication-induced changes in food web efficiency have implications for higher trophic lev-els. At some point, the upward flow of energy may become insufficient to support an additional trophic level, such as piscivorous fish (Persson et al. 1988). Havens et al. (2000) documented the low ecological transfer efficiency that is typical of highly eutrophic lakes. They measured transfer
efficiency of plankton when cyanobacteria dominated the phytoplankton and copepods dominated the zooplankton using a radiotracer method developed by Ducklow et al. (1986). Havens et al. (2000) inoculated whole water samples including indigenous plankton with 14C-labeled glucose or bicarbonate (carbon sources for bacteria and phytoplankton, respectively). In both cases, they found that just 0.1 % to 1.0 % of the carbon was transferred to >200 m size particles (macrozooplankton) after 4 h of incubation. The same results were obtained when the experiment was repeated four more times over a year-long period. Similarly, when Sommaruga (1995) developed a carbon budget for a eutrophic lake in Uruguay dominated by filamentous cyanobacteria and small zooplankton, they measured extreme low ecological trans-fer efficiencies. This situation may explain, in part, why it is common to find hypereutrophic lakes dominated by fish, such as gizzard shad and carp, which can feed low in the food web, i.e., graze directly on phytoplankton and detritus (Bays and Crisman 1983; Allen et al. 2000), and thereby cir-cumvent the inefficient plankton food web.
6.5 Changes in Plants and Effects on Plankton
Other profound changes, described in this book, occur in lakes as they undergo eutrophication, and some of them can affect the plankton food web. One notable change is the loss of submerged aquatic vegetation (SAV) when biomass of cy-anobacteria becomes extremely high in the phytoplankton. Cyanobacteria blooms block light penetration to the extent that they shade out the SAV. The phenomenon is most com-mon in shallow eutrophic lakes and it has been suggested that it can happen quickly, with a lake quickly transitioning from a clear water phase with abundant SAV and low phyto-plankton biomass and a turbid water phase with little or no SAV and high phytoplankton biomass as the lake crosses a nutrient threshold (Scheffer et al. 1993). When SAV is lost from a lake, so too is the refuge they provide for Daphnia
Daphnia
Flagellates
DOC
BACT
CO2
Bosmina
Anabaena
DOC
BACT
CO2a b
Fig. 6.6 Simple diagrams of C flow in food webs contrasting domi-nance of Daphnia and edible flagellates (a) with dominance by Bos-mina and inedible cyanobacteria (b). The thickness of arrows corre-sponds to relative amounts of C flow
PHYT(600)
HFL(60)
PFL(80)
BAC(70)
CIL(70)
MIC(2)
MAC(30)Metazoan consumer
Protozoan consumerPrimary producerDecomposer
Size of boxes correspond toCarbon biomass (mg C L-1)And arrows indicate trophicLinks identified by Work et al.(2005)
Fig. 6.5 Partitioning of carbon among various components of the plankton food web of a subtropical eutrophic lake, Lake Okeechobee, Florida. The data are based on Havens et al. (2007). MAC macrozoo-plankton, MIC microzooplankton, CIL ciliates, HFL heterotrophic flagellates, PFL phototrophic flagellates, BAC bacteria, PHYT phyto-plankton

78 K. E. Havens
and other large zooplankton against plankton-eating fish, or refuge for those fish against larger fish predators. Depend-ing on the role that the plants were playing in the particular lake, loss of SAV could lead to a reduction or increase in the zooplankton. It should be noted that there is some debate as to whether the rapid switch from clear to turbid is typical or unusual. Sayer et al. (2010), for example, studied SAV and phytoplankton dynamics in 39 shallow lakes in the UK and Denmark over a broad nutrient gradient. They documented a long-term loss of SAV in many lakes that were affected by cultural eutrophication and suggested that there was a gradu-al replacement of plants by phytoplankton over a time period of 10 to 100 years, rather than the sudden switch predicted by the alternative states model. Other recent studies support this view (e.g., James et al. 2005). Yet, there are cases where transitions happen more quickly. In Lake Okeechobee, Flor-ida, for example, we observed a switch from turbid water with dense cyanobacteria blooms to clear water with wide-spread SAV ( Chara) in just 1 month (Havens et al. 2001). To a large extent, the rate of change may depend on the bot-tom morphology. In the case of Lake Okeechobee, the switch happened across a large area of shallow water with a nearly flat bottom versus the gradually sloping shoreline that is common in many lakes.
6.6 Changes in Fish and Effects on Plankton
Changes in fish assemblages also occur with eutrophica-tion and this may be owing, in part, to the afore-mentioned changes in the plankton food web; similarly, fish may have strong top-down impacts on the plankton. In particular, shal-low eutrophic lakes support high densities of planktivorous and omnivorous fish (Jeppesen et al. 2007) including species that are voracious sight-feeding predators of zooplankton, as well as filter-feeding omnivores such as gizzard shad that consume benthos, phytoplankton, and zooplankton (Cris-man and Beaver 1990). Warm-water lakes in the subtropics appear to have particularly high predation pressure on the zooplankton and have extreme low ratios of zooplankton to phytoplankton biomass, absence of Daphnia (e.g., Jeppesen et al. 2007; Havens et al. 2009), and lack of control of the phytoplankton by zooplankton grazers (Havens et al. 1996). In these lakes, most energy flow happens in microbial path-ways (Crisman and Beaver 1990; Work et al. 2005).
Although some (e.g., Hart 2011) suggest that large ined-ible phytoplankton are responsible for these characteristics, most research indicates that the paucity of zooplankton is a direct response to fish predation (Meerhoff et al. 2007; Igle-sias et al. 2008; Jeppesen et al. 2007; Havens and Beaver 2012). Jeppesen et al. (2000a) documented that in New Zea-land lakes, there is a strong positive correlation between fish
and phytoplankton biomass, but no such correlation between zooplankton and phytoplankton. These results indicate top-down control of the zooplankton by fish predation. Jeppesen et al. (2003) also examined data from 466 temperate to arc-tic lakes and found a strong pattern: fish control over large zooplankton is strongest in oligotrophic and highly eutrophic lakes and weakest in mesotrophic lakes (consistent with the findings about unimodal relationships discussed earlier). Jeppesen et al. (2000b) found that across a TP gradient of < 50 to > 400 g l−1, there was a decline in piscivores, an increase in cyprinids, a shift to smaller zooplankton, and a reduction in mean body weight of cladocerans from 5.1 to 1.5 g. The ratio of zooplankton to phytoplankton biomass dropped from 0.5 to 0.1 as biomass of phytoplankton increased 15-fold. In three lakes where fish kills occurred, the trends in plankton size and biomass quickly reversed, similar to the findings noted earlier in lakes that underwent reductions in external nutrient loads. Similarly, Auer et al. (2004) found that in-creases in rotifers, ciliates, and nanoflagellates seen in hy-pereutrophic lakes occur because of relaxed grazing pressure by larger Cladocera, which have been eliminated by fish. In Lake Okechobee, Florida, we recently found that seasonal changes in the ratio of zooplankton to phytoplankton bio-mass are almost entirely driven by seasonal changes in bio-mass of the zooplankton and that those changes coincide with periods of high versus reduced fish predation.
6.7 Implications for Lake Management
Reduction of external nutrient loads is a prerequisite for re-versing cultural eutrophication. At the same time, the process may be hastened if it is possible to perform some ecological engineering of the food web. This sometimes is done by ma-nipulating the density of planktivorous or omnivorous fish in order to reduce grazing pressure on Daphnia, so that animal can become more abundant and graze down the phytoplank-ton (e.g., Shapiro and Wright 1984; Carpenter et al. 1987). The goal is to reduce the amount of phytoplankton per unit of P in the lake, via an animal that can filter large volumes of water and feed on nearly the entire plankton food web, from phytoplankton to bacteria. The challenge is maintaining the lowered biomass of planktivorous and omnivorous fish. At some point in the rehabilitation process, where the lake reestablishes an oxygenated deep water zone in summer, it may be possible to introduce piscivorous fish to naturally keep planktivores under control. These approaches are not as effective in the subtropics, where large Daphnia generally do not occur, and where the microbial food web is more important than in temperate lakes of a similar degree of en-richment (Crisman and Beaver 1990; Havens et al. 2007). Thus, in the subtropics, control of external nutrient loading

796 Lake Eutrophication and Plankton Food Webs
is of greater importance for reversal of eutrophication. Al-though this situation is now restricted to the subtropics, with global warming, the same challenges could migrate north-ward into temperate lakes (Moss et al. 2011).
References
Allen MS, Hoyer MV, Canfield DE et al (2000) Factors related to giz-zard shad and threadfin shad occurrence and abundance in Florida lakes. J Fish Biol 57:291–302
Auer B, Elzer U, Arndt H et al (2004) Comparison of pelagic food webs in lakes along a trophic gradient and with seasonal aspects: influ-ence of resource and predation. J Plankton Res 26:697–709
Azam F, Fenchel T, Field JG et al (1983) The ecological role of water column microbes in the sea. Mar Ecol Prog Ser 10:257–263
Bays JS, Crisman TL (1983) Zooplankton and trophic state relation-ships in Florida lakes. Can J Fish Aquatic Sci 40:1813–1819
Carpenter SR, Kitchell JF, Hodgson JR, Cochran PA, Elser JJ, Elser MM, Lodge DM, Kretchmer D, He X, von Ende CN et al (1987) Regulation of lake ecosystem primary productivity by food web structure in whole-lake experiments. Ecology 68:1863–1876
Crisman TL, Beaver JR (1990) Applicability of biomanipulation for managing eutrophication in the subtropics. Hydrobiology 200:177–181
Cole JJ, Findlay S, Pace ML et al (1988) Bacterial production in fresh and saltwater ecosystems: a cross-system overview. Mar Ecol Prog Ser 43:1–10
Conley DJ, Paerl HW, Howarth RW, Boesch DF, Seitzinger SP, Havens KE, Lancelot C, Likens GE et al (2009) Controlling eutrophication: nitrogen and phosphorus. Science 323:1014–1015
Downing JA, Watson SB, McCauley E et al (2001) Predicting cyano-bacteria dominance in lakes. Can J Fish Aquat Sci 58:1905–1908
Ducklow HW, Purdie DA, Williams PJL, Davis JM et al (1986) Bac-terioplankton: a sink for carbon in a coastal marine plankton com-munity. Science 232:865–867
Gliwicz ZM (1969) Studies on the feeding of pelagic zooplankton in lakes of varying trophy. Ekol Pol 17:663–708
Hart RC (2011) Zooplankton biomass to chlorophyll ratios in relation to trophic status within and between ten South African reservoirs: causal inferences, and implications for biomanipulation. Water SA 37:513–521
Havens KE, East TL, Beaver JR et al (1996) Experimental stud-ies of zooplankton-phytoplankton-nutrient interactions in a large subtropical lake (Lake Okeechobee, Florida, USA). Freshw Biol 36:579–597
Havens KE, Work KA, East TL et al (2000) Relative efficiencies of car-bon transfer from bacteria and algae to zooplankton in a subtropical lake. J Plankton Res 22:1801–1809
Havens KE, Jin KR, Rodusky AJ, Sharfstein B, Brady MA, East TL, Iricanin N, James RT, Harwell MC, Steinman AD (2001) Hurricane effects on a shallow lake ecosystem and its response to a controlled manipulation of water level. ScientificWorldJournal 1:44–70
Havens KE, Beaver JR, East TL et al (2007) Plankton biomass par-titioning in a eutrophic subtropical lake: comparison with results from temperate lake ecosystems. J Plankton Res 29:1087–1097
Havens KE, Elia AC, Taticchi MI, Fulton RS et al (2009) Zooplantkton-phytoplankton relationships in shallow subtropical lakes Apopka (Florida, USA) and Trasimeno (Umbria, Italy). Hydrobiologia 628:165–175
Hillbricht-Ilkowska A (1977) Trophic relations and energy flow in pelagic plankton. Pol Ecol Stud 3:3–98
Iglesias C, Mazzeo N, Goyenola G, Fosalba C, Teixeira De Mello F, Garcia S, Jeppesen E (2008) Field and experimental evidence of the
effect of Jenynsiamultidentata, a small omnivorous-planktivorous fish, on the size distribution of zooplankton in subtropical lakes. Freshw Biol 53:1797–1807
James C, Fisher J, Russel V, Collings S, Moss B et al (2005) Nitrate availability and hydrophyte species richness in shallow lakes. Freshw Biol 50:1059–1063
Jeppesen E, Lauridsen TL, Mitchell SF, Christoffersen K, Burns CW et al (2000a) Trophic structure in the pelagial of 25 shallow New Zealand lakes: changes along nutrient and fish gradients. J Plankton Res 22:951–968
Jeppesen E, Jensen JP, Sondergaard M, Lauridsen T, Landkildehus F (2000b) Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient. Freshw Biol 45:201–218
Jeppesen E, Jensen JP, Jensen C, Faafeng B, Hessen DO, Sondergaard M, Lauridsen T, Brettum P, Christoffersen K (2003) The impact of nutrient state and lake depth on top-down control in the pelagic zone of lakes: a study of 466 lakes from the temperate zone to the Arctic. Ecosystems 6:313–325
Jeppesen E, Sondergaard M, Jensen JP, Havens KE et al (2005) Lake responses to reduced nutrient loading—an analysis of contempo-rary data from 35 European and North American long term studies. Freshw Biol 50:1747–1771
Jeppesen E, Meerhoff M, Jacobson BA, Hansen RS, Sondergaard M, Jensen JP, Lauridsen TL, Mazzeo N, Branco CWC et al (2007) Restoration of shallow lakes by nutrient control and biomanipu-lation—the successful strategy varies with lake size and climate. Hydrobiologia 581:269–285
Lindeman RL (1942) The trophic-dynamic aspect of ecology. Ecology 23:399–418
Meerhoff M, Clemente JM, Texiera De Mello F, Iglesias C, Pedersen AR, Jeppesen E (2007) Can warm climate-related structure of lit-toral predator assemblages weaken the clear water state in shallow lakes? Global Change Biol 13:1888–1897
Moss B, Kosten S, Meerhoff M, Battarbee RW, Jeppesen E, Mazzeo N, Havens K, Lacerot G, Liu Z, De Meester, L, Paerl H, Scheffer M et al (2011) Allied attack: climate change and eutrophiction. Inland Waters 1:101–105
Munawar M, Fitzpatrick M, Niblock H, Lorimer J et al (2011) The rela-tive importance of autotrophic and heterotrophic microbial commu-nities in the planktonic food web of the Bay of Quinte, Lake Ontario 2000–2007. Aq Ecos Health Manage 14:21–32
Persson L, Andersson G, Hamrin SF, Johansson L et al (1988) Predator regulation and primary production along the productivity gradient of temperate lake ecosystems. In: Carpenter SR (ed) Complex inter-actions in lake communities. Springer-Verlag, New York
Porter KG, Paerl H, Hodson R, Pace M, Priscu J, Rieman B, Scavia D, Stockner J et al (1988) Microbial interactions in lake food webs. In: Carpenter SR (ed) Complex interactions in lake communities. Springer-Verlag, New York
Riemann B, Sondergaard M (1986) Carbon dynamics in eutrophic tem-perate lakes. Elsevier, New York
Sayer CD, Davidson TA, Jones JI et al (2010) Seasonal dynamics of macrophytes and phytoplankton in shallow lakes: a eutrophication-driven pathway from plants to plankton? Freshw Biol 55:500–513
Scheffer M, Hosper SH, Meijer ML, Moss B, Jeppesen E et al (1993) Alternative equilibria in shallow lakes. Trends Ecol Evol 8:275–279
Shapiro J, Wright DI (1984) Lake restoration by biomanipulation: Round Lake, Minnesota, the first two years. Freshw Biol 14:371–383
Sherr EB, Sherr BF, Albright LJ et al (1987) Bacteria: link or sink? Science 235:88
Smith VH, Joye SB, Howarth RW (2006) Eutrophication of freshwater and marine ecosystems. Limnol Oceanogr 51:351–355
Sommaruga R (1995) Microbial and classical food webs: a visit to a hypereutrophic lake. FEMS Microbiol Ecol 17:257–270

80 K. E. Havens
Stockner JG, MacIsaac EA (1998) British Columbia lake enrich-ment programme: two decades of habitat enrichment for Sockeye Salmon. Regulated Rivers: Res and Manage 12:547–561
Stockner JG, Shortreed KS (1989) Algal picoplankton production and contribution to food webs in oligotrophic British Columbia lakes. Hydrobiology 175:151–166
Work K, Havens K, Sharfstein B, East T et al (2005) How important is bacterial carbon to planktonic grazers in a turbid, subtropical lake? J Plankton Res 27:357–372
Weisse T, Stockner JG (1992) Eutrophication: the role of microbial food webs. Mem Ist Ital Idrobiol 52:133–150





























