Embed Size (px)
Citation preview

q 2002 The Paleontological Society. All rights reserved. 0094-8373/02/2804-0005/$1.00
Paleobiology, 28(4), 2002, pp. 474–507
Evolution of development of the vertebrate dermal andoral skeletons: unraveling concepts, regulatory theories,and homologies
Philip C. J. Donoghue
Abstract.—It has been contended that Reif’s odontode regulation theory is a rival and alternative toStensio and Ørvig’s lepidomorial theory as means of explaining the evolution of development of thevertebrate dermal and oral skeleton. The lepidomorial theory is a pattern-based theory that providesa homological framework that goes further than the odontode regulation theory in comparing dentalpapillae and their products, and it provides an explanatory mechanism for such relationships a pos-teriori. In contrast, the odontode regulation theory is process-based and observes only developmentalsimilarity, providing no means of identifying homologies beyond this. The lepidomorial theory issuperior to the odontode regulation theory in its ability to trace homology through the evolution ofdevelopment of the dermal and oral skeleton. The criteria proposed to identify homology betweenscales—either within a given individual or taxon, or between different individuals or taxa—are, pri-marily, vascular architecture and, secondarily, external morphology. External morphology may beexcluded on Reif’s argument for the overarching principle of differentiation, a hypothesis supportedby recent advances in the understanding of dental morphogenesis. Vascular architecture is potentiallyuseful but appears to be determined by tooth/scale morphology rather than reflecting historical (phy-logenetic) constraint. Data on the development of epithelial appendages, including teeth, scales, andfeathers, indicate that individual primordia develop through progressive differentiation of originallylarger, homogenous morphogenetic fields. Thus, there is no mechanism of ontogenetic developmentalconcrescence, just differentiation. Phylogenetic patterns of concrescence and differentiation are sim-ilarly achieved through ontogenetic developmental differentiation, or a lack thereof. In practice, how-ever, it is not possible to distinguish between patterns of phylogenetic concrescence and differenti-ation because there is no means of identifying homology between individual elements within a squa-mation, or a dentition (in almost all instances). Thus, phylogenetic patterns of increase and decreasein the numbers of elements constituting dentitions or dermal elements are best described as such;further attempts to constrain precise underlying patterns remain without constraint and outside therealms of scientific enquiry. The application of the homology concept in the dermal and visceral skel-etons is explored and it is determined that odontodes are serial homologs, conform only to the bio-logical homology concept at this level of observation, and are devoid of phylogenetic meaning. It isconcluded that Reif’s theory is close to a universal theory of the evolution of development for thedermoskeleton and dentition, and additional components of theory, including the regulatory basis oftemporal and spatial patterning, are tested and extended in light of data on the development of thechick feather array. Finally, the dermoskeleton is identified as an exemplary system for examining theregulatory basis of patterning and morphogenesis as it encompasses and surpasses the repertoire ofestablished model organ systems.
Philip C. J. Donoghue. Lapworth Museum of Geology, School of Geography, Earth and Environmental Sci-ences, University of Birmingham, Edgbaston, Birmingham B15 2TT, United Kingdom. E-mail:[email protected]
Accepted: 18 June 2002
Introduction
Teeth and scales are among the most abun-dant and most readily fossilizable compo-nents of the vertebrate skeleton and it is notsurprising, therefore, to discover that theyhave played a significant role in phylogeneticreconstruction and the classification of bothfossil and living vertebrates (e.g., Agassiz1833–1843; Owen 1845). Attempts to tracethese skeletal structures have developed inhand with the primary concept in compara-
tive anatomy, homology. Indeed, the mostpressing problem in comparative biology, thereconciliation of anatomical, developmental,and historical concepts of homology (e.g., deBeer 1971; Wagner 1989), was attendant in thelate nineteenth century when anatomical (pa-leontological) and embryological approachesto tracing homology in reptilian and mam-malian dentitions first clashed. Similar at-tempts had been underway in the dermal skel-eton, stemming from Williamson’s (1849,

475SKELETAL EVOLUTION OF DEVELOPMENT
1851) use of placoid scales from extant sharksas conceptual units in comparative analysis.Subsequently, various authors have formulat-ed evolutionary hypotheses in which placoidscales have figured centrally as conceptualbuilding blocks in the evolution and evolutionof development of vertebrate teeth and scales(e.g., Hertwig 1974a,b; Ørvig 1967, 1968, 1977;Reif 1982a).
Most notable among these debates are theissues of concrescence (assimilation/coales-cence) and differentiation (increased morpho-logical specialization and/or division of ele-ments) as patterns in the evolution of teethand scales, and the developmental mecha-nisms that underlie these patterns. Althoughon face value this a subject is of little morethan esoteric value, recent approaches to un-derstanding the basic principles of organo-genesis in vertebrates have broadened the sig-nificance of these issues and debates. The de-velopment of teeth, among the simplest of allvertebrate organs (Jernvall 1995; Stock et al.1997), has been studied at ever increasing res-olution such that the controls that directly un-derpin patterning and morphogenesis are be-ginning to be understood (e.g., Peters andBalling 1999). From what is known already, itis clear that many of the aspects of tooth de-velopment in laboratory animals, such as themouse, are common to the teeth of other mam-mals (e.g., Keranen et al. 1998, 1999), and tothe developmental programs of other epithe-lial-appendage organ systems (such as feath-ers, hair, and mammary, sebaceous, and sweatglands) of other vertebrates as well as inver-tebrate metazoans (e.g., Krejsa 1979; Thesleffet al. 1995; Chuong 1998). Given the degree ofconservation in induction and patterningmechanisms, we may be confident in impli-cating their activity in the patterning and mor-phogenesis of organisms that have yet to bestudied experimentally, and in those taxaknown solely from paleontological data. Thus,we may begin to use these new data in at-tempting to reconcile long-standing disputesregarding the evolution of patterning andmorphogenesis in these organ systems. Inturn, the fossil record provides data on extinctclades of vertebrates that are obviously notopen to experimental analysis. Most critically,
the organisms in which this developmentalsystem first evolved are entirely extinct andwe have no recourse but to paleontologicaldata in attempting to trace and understand theorigin and early evolution of this developmen-tal system.
Before reviewing established models forpatterning the vertebrate dermal and visceralskeleton, it is worth noting precisely what thisencompasses. The vertebrate skeleton is wide-ly perceived as two distinct systems that havequite different developmental and phyloge-netic backgrounds (Patterson 1977). The en-doskeleton, which is composed of the verte-brae and associated axial structures, the limbs,the brain case, and various components of theskull, is the skeleton that is most familiarlyvertebrate. However, the dermal and oral skel-eton, which is composed of the teeth, scales,fin spines, and dermal bones of vertebrates,particularly lower vertebrates, is primitivelythe most extensive and, although not the mostprimitive, is the first to appear in a mineral-ized form, both geologically and phylogenet-ically (Donoghue and Aldridge 2001; Dono-ghue and Sansom 2002). This paper concernsonly the evolution of development of the ver-tebrate dermal and oral skeleton.
In the recent literature on the evolution ofdevelopment of the vertebrate skeleton, theterms ‘‘dermoskeleton’’ and ‘‘exoskeleton’’have been used interchangeably. In this sense,the exoskeleton is used as an antonym to the‘‘endoskeleton,’’ but there are two main rea-sons why this is not an altogether accurate orappropriate description of this skeletal sys-tem. First, the vast majority of the ‘‘exoskele-ton’’ develops within the dermis and, thus,may more appropriately be assigned to thedermoskeleton (dermal skeleton) (Francillon-Vieillot et al. 1990). Strictly, the exoskeleton isrestricted to keratinous elements such ashorns, nails, claws, hairs, feathers, etc. Sepa-rating these into separate systems may be lit-tle more than semantic and may actually beentirely inaccurate from a historical, ratherthan a developmental, perspective. However,there remains potential for confusion throughsynonymy of the dermoskeleton and exoskel-eton. This stems from suggestions that the dif-ference in epithelial sources between true

476 PHILIP C. J. DONOGHUE
‘‘dermal’’ skeletal elements, such as scales,and endothelial teeth (and oral scales) impartsreal difference to two systems that are arguedto have been distinct since early in vertebratephylogeny (Smith and Coates 1998, 2000,2001). Thus, although experimental evidenceindicates that the patterning mechanisms un-derlying dental development are preciselysimilar to those controlling components of thedermoskeleton (see, e.g., Peterkova et al. 2000),the distinction is maintained.
Finally, it is worth noting that a further di-chotomy exists that cuts across any topologi-cal subdivision of the ‘‘exoskeleton’’ into oraland extra-oral systems. These are the odon-togenic and skeletogenic scleroblastic ecto-mesenchymal cell lineages that are responsi-ble for the dental elements (teeth and toothlikestructures) and bony plates (respectively) inthe dermal and oral skeleton. Although ex-perimental evidence in support of this hascome to light only relatively recently (Palmerand Lumsden 1987; Smith and Hall 1990,1993; Atchley and Hall 1991; Vaahtokari et al.1991), empirical evidence for separate devel-opmental backgrounds has been known forsome time (e.g., Westoll 1967). Quite surpris-ingly, established theories concern only theodontogenic component, addressing the ske-letogenic component as though it were only aminor component of a holistic odontogenicscleroblastic system. As this contribution at-tempts to test these theories, the skeletogenicsystem will not be addressed.
Historical Review of Patterning Conceptsand Theories
Early Work. The earliest attempts to unrav-el the nature of development and patterning ofthe vertebrate dermoskeleton and visceralskeleton place central importance upon thenature of the placoid scales in living sharks.These began with Williamson (1849, 1851)who was among the first students of paleohis-tology and the first to provide a detailed ac-count of the histology of Recent shark scales.On this basis, Williamson developed a theoryin which the cosmine scales of basal sarcop-terygians had developed through the amal-gamation, or ‘‘concrescence,’’ of units equiv-alent to placoid scales. However, and although
Williamson identified intermediates betweenplacoid and cosmine conditions, he did notnecessarily suggest that these represent anevolutionary sequence (which is not surpris-ing given that Williamson’s work was under-taken at least ten years before the publicationof The Origin). Williamson’s observation of‘‘analogy’’ between placoid and cosminestructure were given an evolutionary slant bysubsequent authors such as Hertwig (1874a,b;1876, 1879, 1882) and Goodrich (1907), whoappear to have assumed the primitiveness ofthe placoid condition. Hertwig and Goodrichturned Williamson’s hypothesis of concres-cence in the development of scales to one ofconcrescence in the evolution of developmentof scales and dermal bones. For instance, Her-twig’s main argument was that teleost scalesand large dermal bones developed throughthe concrescence of placoid scales such thatthe scale-bases fused into a single mass and,eventually, the surmounted denticles werelost. In contrast, Klaatsch (1890) appears tohave proposed the first differentiation hy-pothesis, arguing that scales and plates devel-op from a single denticle in which the basalplate has become much enlarged.
Despite the generally widespread accep-tance of concrescence mechanisms as expla-nations for the diversity of skeletal structuresmet with in the dermal skeleton, evolutionarystudies of dental patterning have had littletruck with concrescence as a transformationalmechanism. Nevertheless, the shift from sim-ple homodont coniform dentitions of mam-malian antecedents to complex heterodontmolariform dentitions of mammals has tempt-ed many authors to suggest that the cusps ofcomplex teeth arose through amalgamation ofsimpler teeth (e.g., Ameghino 1884, 1896,1899; Adloff 1916; Bolk 1912, 1922; Kukenthal1893; Rose 1892; Winge 1883). Concrescencecontinues to be raised as both a developmen-tal and a phylogenetic mechanism (e.g., Peter-kova et al. 2000).
The Lepidomorial Theory. During the middlepart of the twentieth century, Erik Stensio andTor Ørvig developed a theoretical model thatattempted to explain the patterning of the der-mal skeleton, to resolve between skeletal ele-ments both within and between taxa, and the

477SKELETAL EVOLUTION OF DEVELOPMENT
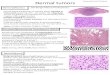
mechanism through which evolutionary trans-formations occurred. The ‘‘lepidomorial theo-ry’’ was developed from a study of Paleozoicchondrichthyan scales that was never pub-lished (Stensio and Ørvig 1951–1957); only themain conclusions of the theory ever made itinto print. The first reference to this theory wasmade by Jarvik (1948), and it was outlined ingreater depth by Ørvig (1951) and Stensio(1958, 1961, 1962, 1964). The basic tenet of thetheory (as published) is that the dermal andoral skeleton is, at its most basic level, the prod-uct of developmental modules termed lepido-moria. Each lepidomorium could be recognizedon the basis that it was composed of ‘‘an enam-el-coated crown of dentine and a basal plate sit-uated in the corium. To judge from the two ca-nals leading out from its pulp cavity each lep-idomorium arose ontogenetically from a sim-ple corium papilla formed around a singlevascular loop which ascended in a superficialdirection from the sub-epidermal vascularplexus of the corium’’ (Stensio 1961: p. 236)(Fig. 1A–F). Subsequent development was per-ceived to follow the pattern typified by placoidscales; enameloid develops immediately adja-cent to the enamel organ, constituting a rigidmold in which dentine begins to form; as such,the scale attains its definitive size and shape atonce. Dentine forms apically first, and basallysecond, and the plate tissues begin to developin coordination with the basal dentine.
As described, the lepidomorial theory is littlemore than a generalized description of the de-velopment of a placoid scale, as set out by Her-twig (1874a) and Klaatsch (1890). Although itis a concrescence-based model, it departs sig-nificantly from earlier theories in its interpre-tation of placoid scales. Modern shark scaleshad traditionally been held to reflect the prim-itive condition for the dermal skeleton, firstlybecause sharks were thought to be extremelyprimitive, but also because placoid scales arestructurally simple. However, after studyingthe scales of a group of poorly known Late Pa-leozoic chondrichthyans (Fig. 1G–K), Stensioand Ørvig (1951–1957) reached the conclusionthat the basic structural unit of the dermal skel-eton is a scale that is morphologically andstructurally simpler than the condition metwith in placoid scales. Most specifically, these
authors argued that the simplest of all scalesare vascularized by a single capillary loop thatenters the pulp cavity from below and exits viaa canal in the neck of the scale (Fig. 1G). Thus,because placoid scales possess supernumeraryneck canals (Fig. 1A–F)—Stensio (1961) cites arange of typically 3–15 or more per scale—theymust be derived from the coalescence of asmany ‘‘primitive’’ scales, or lepidomoria. In thisview, placoid scales are neither simple norprimitive, but extremely complex and highlyderived. Far from suggesting that scales actu-ally coalesced to form a single larger scale,Stensio and Ørvig (1951–1957) suggest that theprimordia, the lepidomoria of these scales, co-alesced (Fig. 1A) and, thus, there is no recordof this transformational event in the mineral-ized tissues, other than the occurrence of su-pernumerary neck-canals (Fig. 1B–F).
Not only does the lepidomorial theory ar-gue that morphologically complex scales andteeth arise through coalescence, but it also im-plicitly precludes differentiation as a mecha-nism for morphological change. Immediatelyafter its inception, the lepidomorial theory be-came extremely influential and provided thecriterion upon which entire orders of fish wereclassified (Stensio 1958, 1964). Further, Stensio(1961) argued that the lepidomorial theoryprovided the explanatory medium for evolu-tionary transformation between dental types,and most particularly for Cope (1883, 1889),Winge (1883) and Osborn’s (1907) theories ofincreased complexity of tooth morphology,through the amalgamation of teeth of simplemorphology.
The Odontode Concept. Beginning withØrvig (1967), a new concept, the ‘‘odontode’’was conceived. At an operational level, thereis little to distinguish between lepidomoriaand odontodes; Ørvig’s most precise defini-tion of the concept was (Ørvig 1977: p. 54)‘‘special hard tissue units, or dental units,which generally speaking have those devel-opmental and structural properties in com-mon with the teeth of the jaws (whenever suchare present) that they (a) each develop onto-genetically in a single, undivided dental papilla ofmesenchymal soft tissue, bounded at its cir-cumference by an epithelial dental organ in theadjoining dermis, (b) consist of dentine or, in
478 PHILIP C. J. DONOGHUE
FIGURE 1. A–K, The lepidomorial theory, from Stensio 1961 with the permission of Toronto University Press; L–O,The odontode regulation theory, from Reif 1982a. A–F, Diagrammatic representation of the hypothetical sequenceof development of a placoid (synchronomorial) scale as envisaged by Stensio and Ørvig (1951–1957). G–K, Sequenceof development of a cyclomorial scale based upon edestid scales from the Permian of Greenland. L–O represent thebasic tenets of the odontode regulation theory, in which scales suppress development of new scale primordia byexerting a ‘‘zone of inhibition’’ upon the adjacent dermis. New scales develop through growth of the dermis, leading

479SKELETAL EVOLUTION OF DEVELOPMENT
←
to gaps in the zones of inhibition, through shedding of established scales, or through cessation in the inhibitoryeffects of established scales such that new primordia develop on top of existing scales. Reproduced with amendmentfrom Reif 1982a, with the permission of Plenum/Kluwer Publishing.
some forms, dentinous tissue . . . and (c) fre-quently (but not always) possess a superficiallayer of enameloid. At the same time, however,odontodes are in general terms also charac-terized by (d) not belonging to the dentitionsensu stricto but to other parts of the dermalskeleton (sometimes including those imme-diately adjoining the dentition), (e) not, as arule, fulfilling similar functions as teeth, (f) notforming in a submerged position in connec-tion with a dental lamina or single, ingrowingepidermis digitations but always in the super-ficial part of the corium . . . because of whichthey are not replaced from below or sidewaysin a manner of teeth in the great majority offishes, and (g) in many cases, not reachingnearly the same height (from top to base) asteeth.’’ It is clear from Ørvig’s later papersthat he was attempting to draw a distinctionbetween the odontode, as a concept, and lep-idomoria, as theoretical developmental units;i.e., he was attempting to distinguish patternfrom process. For instance, Ørvig (1977: p. 55)argues that the recognition of an odontode isa purely descriptive observation, that does‘‘not involve interpretation and can be appliedirrespective of whether or not one adopts linesof reasoning such as those e.g., the lepido-morial theory (sensu Stensio 1961). Odonto-des, for instance, can just as well be single lep-idomorial crowns as multi-lepidomorial for-mations of all degrees of complexity whenthey are analyzed on the basis of that theory.’’Although this was not stated explicitly at thepoint of inception of the odontode concept, itis clear from Ørvig’s usage of the concept inhis intervening papers that he intended thismeaning implicitly. For instance, in dealingwith the dermal skeleton of the placoderm Ro-mundina stellina, Ørvig states that ‘‘althoughall these tubercles certainly qualify as singleodontodes (sensu Ørvig 1967, 1968), there isevery reason to assume that each of them hasarisen phyletically by the fusion of a group ofconsiderably smaller odontodes of a kind ex-isting in the dermal skeleton of ancestral
forms. These latter odontodes are naturally nomore identifiable as such, but are neverthelessfrequently traceable by the nodules or cuspson the tubercles referred to above. Since thereare here, as mentioned, occasionally alsocusps associated with side-cusps, some of theoriginal odontodes by the coalescence ofwhich the tubercles formed were presumablybi- or tricuspidate elements’’ (Ørvig 1975: p.44). Nevertheless, Ørvig demonstrates reluc-tance to disassociate entirely pattern fromprocess and this is clear even in the paper inwhich he attempts to draw a firm distinctionbetween the recognition of morphogeneticunits (the odontodes), and the interpretationof their transformational relationships, i.e., be-tween a concept and a theory. In the case ofcosmine, Ørvig (1977: p. 66) argues that theodontodes have ‘‘fused so completely witheach other in a horizontal direction . . . thatthey lost their original ability to form sepa-rately, each in a dental papilla of its own, andall arose within one single large dental papillawith the same extent as the cosmine sheet asa whole. . . . In this case it seems far more ap-propriate to speak of component hard tissuesunits . . . than of component odontodes. It is,in fact, each individual cosmine sheet whichaccording to the definition above should de-serve to be referred to as an odontode, but thisusage is obviously awkward, and it is notadopted here.’’
In summary, Ørvig’s odontode is an oper-ational concept for the recognition of morpho-genetic units within mineralized dermal skel-etons. Although at first the distinction be-tween odontodes and their hypothetical trans-formational cousin, the lepidomoria, waspoor, Ørvig eventually massaged his odon-tode concept to one of pattern recognition,free of the transformational hypotheses thatburden the lepidomorial theory and its asso-ciated terminology.
The Odontode Regulation Theory. Subse-quently, Reif converted the odontode from aconcept to a theory of morphogenesis to rival

480 PHILIP C. J. DONOGHUE
the lepidomorial theory in its explanatorypower. Reif’s work began by testing the pre-mise upon which the lepidomorial theory wasbased: the existence of lepidomoria (Reif1973). Reif attempted this by examining thedevelopment of placoid scales, Stensio’s typeexample of a scale derived from the concres-cence of many lepidomoria. Reif (1973, 1974)found that at no stage in the development ofplacoid scales is there any evidence of differ-entiation of dental papillae into a series of dis-tinct ‘‘lepidomoria’’ and, thus, the basic as-sumption and explanatory mechanism of thelepidomorial theory would appear to be havebeen falsified (and with it the theory uponwhich it forms the basis).
Reif’s perception of the odontode is muchthe same as Ørvig’s, except on two key points.First, Reif (1982a) incorporates teeth into hisconcept of the odontode, whereas Ørvig re-served odontodes to encompass all toothlikestructures that are not actually teeth. Second,and more important, Reif rejects Ørvig’s con-tention that the odontode concept is tenable ir-respective of whether one accepts transfor-mational theories. Instead, Reif (1982a) devel-oped Ørvig’s (1967, 1968, 1977) atomisticodontode concept into a transformational the-ory by augmenting it with the axiom that mor-phological changes take place through differ-entiation. It is not surprising then that Reiffinds that ‘‘I do not agree with Ørvig’s (1977)statement that the odontode concept can beworked with irrespective of whether or notone accepts the Lepidomorial Theory. Myview is that the Odontode-Regulation Theoryis an alternative to the Lepidomorial Theory’’(Reif 1982a: p. 288). In other words, afterØrvig’s attempt to separate the collection ofmorphogenetic data from the interpretation oftransformational mechanisms, Reif (1982a,b)augmented the odontode concept with a newtransformational hypothesis, and then object-ed to Ørvig’s usage of the odontode concept.
Reif’s odontode regulation theory (Reif1982a,b), like the theories erected by William-son, Hertwig, and Goodrich, assumes the pri-macy of the placoid scale as reflecting the ba-sic morphogenetic unit within the vertebratedermal skeleton. Nevertheless, Reif’s theorydiffers from these in its contention that the
principal mechanism of evolutionary changeis differentiation of scale primordia. Thus,large or complex teeth and scales arisethrough changes in the morphogenesis of in-dividual primordia, rather than through theconcrescence of primordia. Reif (1982a,b) ar-gued that the disparity of dermoskeletal typesmet with in both extant and extinct verte-brates can be explained through a combina-tion of morphogenetic differentiation andtemporal and spatial changes in development(in particular, see Reif 1980a) (Fig. 1L–O).
Comparing and Evaluating theLepidomorial and Odontode-Regulation
Theories
Despite its influential nature, the lepido-morial theory has attracted criticism from em-bryologists, and comparative anatomistsstudying extinct and extant taxa alike (e.g.,Zangerl 1966, 1981; Peyer 1968; Gross 1973;Reif 1973, 1974, 1978a, 1982a; Maisey 1988).However, the greatest problems with the lep-idomorial theory lie with the fact that the dataand arguments upon which it is based werenever published, leaving to the published re-cord only short and confusing summaries ofthe implications and conclusions of the work(e.g., Ørvig 1951; Stensio 1961, 1962; Jarvik1960). And further, the theory ‘‘evolved’’through time such that its explanatory powerappears to have varied in scope. Thus, the the-ory has proven difficult to test and, as will bedemonstrated, has not been tested. Thiswould not be problematic if the theory couldbe rejected out of hand. But the theory contin-ues to be influential to this day, both formally(e.g., Karatajute-Talimaa 1992, 1998), and in-formally, in terms of the concepts that it en-capsulates (e.g., Peterkova et al. 2000). Further,as will be demonstrated below, its putative ri-val, the odontode regulation theory, requires arejection of the lepidomorial theory in its for-mulation. What is required, therefore, is anunambiguous explanation of how the lepido-morial theory was formulated, to provide botha means of erecting tests of the theory, and ameans of understanding its relationship to theodontode regulation theory.
A manuscript (Stensio and Ørvig 1951–1957) purporting to outline the foundations of

481SKELETAL EVOLUTION OF DEVELOPMENT
the lepidomorial theory has been cited manytimes in literature, both by the authors and byothers (e.g., Jarvik 1948; Ørvig 1951, 1957;Stensio 1961, 1962; Karatajute-Talimaa 1992,1998; Reif and Richter 2001), although few, ifany other than Erik Stensio and Tor Ørvig, canhave laid eyes upon it. The original manu-script survives as part of the Stensio Archivesat the Department of Palaeozoology, SwedishMuseum of Natural History, Stockholm. Thismanuscript concerns the scales of edestids(holocephalan elasmobranchs) from the Perm-ian of Greenland, which are divided into cat-egories based upon characteristics of the vas-cular architecture, morphology, and inferredhistogeny, and their relationship to the plac-oid scales of Recent elasmobranchs. The man-uscript provides clarity in a number of con-tentious issues. It is clear that the foundationof the theory lies not with existence of lepi-domoria, but in the study of edestid scales thatexhibit evidence of growth through augmen-tation with what are effectively individualscales in their own right (Fig. 1G–K). Stensioand Ørvig (1951–1957) attempted to comparethese ‘‘growing’’ scales to the ‘‘non-growing’’placoid scales of Recent sharks (Fig. 1A–F),and attempted to derive homology betweenthe two. In doing so they used two criteria.First, they compared the number of vascularcanals that supply the scales and, second, theycompared the morphology of the scale crownin these and other groups. Using these asframes of reference for identifying homology,Stensio and Ørvig (1951–1957) concluded thatplacoid scales are homologous not to one ofthe individual units that constitute the grow-ing scales of Paleozoic sharks, but to a numberof such units. (Retrospectively, it is possible todetermine that these points are also madeboth explicitly and implicitly by Ørvig [1951]and Stensio [1961, 1962].) The formulation ofsuch homologies is clearly pattern based andthe basic comparisons are valid regardless ofwhether the developmental explanation of thisphylogenetic pattern is correct. More intrigu-ingly, in this original formulation of the lepi-domorial theory, Stensio and Ørvig (1951–1957) use the lepidomorial concept in refer-ence to their basic unit of homology (vasculararchitecture). Therefore, and at least in its
original conception, a lepidomorium does notrefer to the hypothetical developmental unitsreferred to by Stensio (1961, 1962), and thereis little to distinguish the meaning of Ørvig’sodontode concept from this early conceptionof the lepidomorium. It is perhaps because ofthis changed meaning of the lepidomorialconcept that Ørvig erected the odontode con-cept, referring to the lepidomorial theory‘‘sensu Stensio 1961’’ (Ørvig 1977: p. 55), rath-er than sensu Stensio and Ørvig (1951–1957),as he had earlier (Ørvig 1951, 1957). However,Stensio and Ørvig (1951–1957) do also useboth meanings interchangeably.
In contrast to the lepidomorial theory, thefoundation of the odontode regulation theorylies with the development of placoid scales,and with the assumption that these scales rep-resent the basic, irreducible unit of develop-ment in the dermal and visceral skeleton. It isan axiom of this theory that hard-tissue units(odontodes in the sense of Ørvig 1977) com-parable in structure to placoid scales can beinferred to have developed in a directly com-parable manner. This axiom is supported bySchaeffer’s (1977: p. 26) argument that ‘‘Whenthe morphogenetic parameters for a particularorgan or structure have been establishedthrough experimentation in living forms, andwhen no significant deviation from these pa-rameters has been found, we may postulatethat the morphogenesis of homologous adultstructure occurred in extinct forms in thesame way.’’ Thus, in contrast to the pattern-based lepidomorial theory, the odontode reg-ulation theory is process based.
This difference in approach has significantimplications for the evolutionary patterns andprocesses perceived by the two theories. Forinstance, patterns of concrescence identifiedby the lepidomorial theory are evolutionarypatterns, whereas patterns of differentiationobserved through the odontode regulationtheory are ontogenetic patterns. Thus, in con-sideration of debate over whether the lepido-morial and odontode regulation theories aresynonymous (in whole or in part, e.g., Kara-tjute Talimaa 1998) or direct competitors (Reif1982a; Reif and Richter 2001), we may con-clude that neither situation obtains. Althoughboth theories draw upon the same data set,

482 PHILIP C. J. DONOGHUE
they address this data set in different ways. In-deed, it would appear that conflict with thelepidomorial and odontode regulation theo-ries is, to an extent, unnecessary and the twotheories have the potential to be complemen-tary, collectively constituting a universal the-ory of the evolution of development of the ver-tebrate dermal and oral skeleton.
However, the two theories do clash, and thisstems from the implicit assumption in the lep-idomorial theory that ontogenetic and phylo-genetic patterns equate; i.e., phylogenetic con-crescence results from ontogenetic concres-cence. This assumption is based partly on ob-servations of homology, using the vascularsystem and oral morphology of teeth andscales as a frame of reference, and partly onthe understanding that a scala naturae per-spective of vertebrate phylogeny is character-ized by a motif of skeletal reduction and as-similation. Given that there is no sclerochron-ological record of concrescence in the miner-alized tissues of such dermal elements,Stensio and Ørvig (1951–1957) were led to theconclusion that concrescence must have oc-curred ontogenetically at a stage prior to ini-tial mineralization. Reif’s (1973, 1974, 1978a,1982a) attempted refutation of this hypothesisis flawed on two counts. First, his observationthat placoid scales develop from individualuniform dental papillae is a corroboration ofStensio’s (1961: p. 243) own observations in theformulation of the lepidomorial theory. How-ever, it must be conceded that Stensio’s argu-ment, that the fusion of hitherto distinct den-tal papillae is obscured through phylogenysuch that they cannot be observed in Recentelasmobranchs and holocephalans, effectivelyprotects the hypothesis from falsification. Sec-ond, and more importantly, Reif failed to ad-dress the data on which the inference of on-togenetic concrescence was based, i.e., the in-ference of homology, as well as the use of vas-cular architecture as a criterion for tracinghomology, within the dermal or visceral skel-eton.
The odontode regulation theory requires re-jection of the lepidomorial theory on twocounts. First, unless it can be demonstratedthat there is no level of homology below thatof the odontode, the attempts of the odontode
regulation theory to articulate and explain theevolution of teeth and scales are at best incom-plete and at worst entirely spurious. Second,the use of vascular architecture as a means oftracing homology throughout the dermal andvisceral skeleton, independently of the effectsof both ontogenetic and phylogenetic concres-cence and differentiation, provides an inde-pendent test of the odontode regulation the-ory, a test that the theory fails.
Thus, it is imperative that the relationshipof the lepidomorial and odontode regulationtheories is resolved such that a single univer-sal theory of the evolution of development andpatterning in the vertebrate dermal and oralskeleton can be achieved either through amal-gamation or through rejection of one or boththeories. To this end, the assumptions under-lying these theories must be evaluated. Al-though these have been perennial questions inthe study of the evolution of the vertebrateskeleton, recent advances in developmental bi-ology and developmental genetics have begunto reveal the underlying basis of patterningand morphogenesis, and these data will beused in the evaluation of the lepidomorial andodontode regulation theories.
Before the implications of these experimen-tal data are used to test the lepidomorial andodontode regulation theories, however, it ispertinent to note that these experiments arelargely restricted to rodent dental develop-ment, and it is right to question how appro-priate they are to understanding skeletal vas-cular patterning and morphogenesis in theteeth and scales of lower vertebrates. For in-stance, Smith and Coates (1998, 2000, 2001)have argued that the divergence of dermaland oral skeletal systems is both ancient, pre-ceding the origin of jaws, and permanent.Thus, we might not expect tooth developmentin mammals to tell us very much about dermalscale development in, for example, Paleozoiclungfish. Furthermore, mammalian teeth ex-hibit a number of characteristics that are ex-clusively mammalian. Thus, we might also ex-pect that mammalian tooth development is oflittle relevance to understanding tooth devel-opment in any other vertebrate. However,there is good reason to assume that these datado provide an adequate test of the lepidomo-

483SKELETAL EVOLUTION OF DEVELOPMENT
rial and odontode regulation theories. First,both theories have been applied throughoutthe dermal and oral skeleton of all vertebrates,including mammals. Further, mammalianteeth and the teeth and scales of other verte-brates exhibit precisely similar evolutionarypatterns of morphological, numerical, and to-pological change, and are composed of ho-mologous tissues that are arranged and de-velop in the same way. Finally, there is a com-pelling argument in favor of the ready appli-cability of mammalian tooth development toan understanding of dermal and oral skeletaldevelopment among a wide range of verte-brates. This centers on the fact that the earlystages of tooth morphogenesis exhibit a strik-ing comparison to other epithelial-appendageorganogenic systems such as scale, hair, kid-ney, limb, liver, lung, mammary gland, andsweat gland. This comparison extends beyondmorphological similarities in development,such as mesenchymal condensation and thethickening and folding of epithelial sheets(Krejsa 1979), to commonality of signalingand receptor molecules, transcription factors,cell adhesion, and extracellular matrix mole-cules that participate in the regulation of de-velopment (Thesleff et al. 1995; Chuong et al.2000b; Peterkova et al. 2000). Hence, the de-fective function of such molecules has beenimplicated as a causative role of multi-organimpaired development in such congenital syn-dromes as ectodermal dysplasias (Priolo et al.2000; Thesleff 2000). Despite extreme conser-vatism, there are rare examples where thecausative effects of such human syndromeshave different effects in other vertebrates. Forexample, cbfa1 loss of function in humans re-sults in cleidocranial dysplasia syndrome,where bone is hypoplastic and patients devel-op supernumerary teeth (oligodontia [Mun-dlos et al. 1997]). In mice, however, cbfa1 lossof function results in complete failure of bonedevelopment and in hypodontia, where toothdevelopment is arrested at cap stage (D’Souzaet al. 1999; Inohaya and Kudo 2000 have re-ported a similar function of cbfa1 in teleost de-velopment). Such rare examples notwith-standing, it would be extremely surprising ifthe regulatory networks controlling tooth pat-terning and morphogenesis in mammals were
significantly different from tooth and scaledevelopment in other vertebrates. Thus, wemay follow Schaeffer’s (1977) axiom inferringdevelopment in extinct organisms by compar-ison to generalism of development in extantorganisms.
Testing the Lepidomorial and OdontodeRegulation Theories
Experimental Data on Vascular Development.Considering the clinical implications, it isquite astonishing how little is known aboutthe development and architecture of the vas-cular system in the teeth of vertebrates. Moststudies of the development of dental vascula-ture are restricted to mammalian teeth, anddata are largely limited to inferences basedupon comparing late fetal and adult pheno-type. Lepkowski (1901) described the condi-tion of the vasculature in the tooth germ of aseventh-month fetus. He was able to discernthe branching of the inferior alveolar artery toeach tooth germ, where each branch entered atooth germ via the base and extended throughthe basal odontoblastic membrane to reach theforming dentine via a plexus of branchingcapillaries. Lepkowski observed that the vas-culature varied according to tooth type suchthat there was a direct correlation between thenumber of vessel-bundles and the number oftooth cusps, giving the impression that thetooth had developed from a correspondingnumber of distinct units.
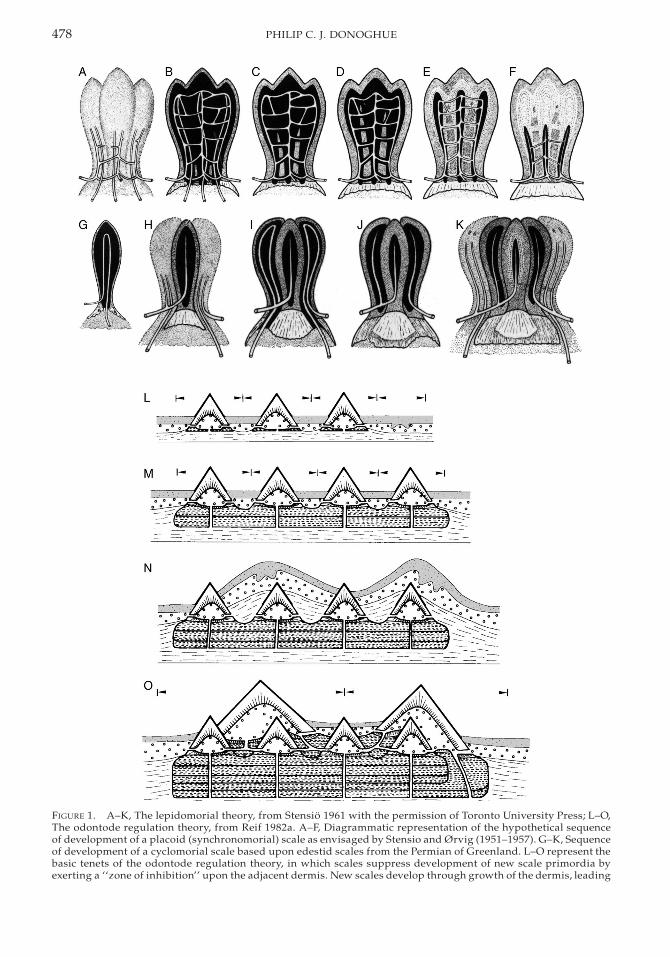
The most comprehensive data on vasculardevelopment during odontogenesis are basedupon incisors and molars of rats (Bernick1960, 1962; Yoshida 1991; Yoshida and Ohsh-ima 1996) (Fig. 2). Yoshida (1991) concerns thedevelopment of the vascular architecturewithin the dental papilla from E18.5 (E 5 em-bryonic day) to E22, representing the day be-fore birth. Bernick (1960) details the develop-ment of the vascular supply from bell stagethrough to early stages of occlusal function(from birth to approximately one month afterbirth) of rat teeth, molar teeth in particular.Bernick (1962) describes the development ofthe vascular architecture from one month toone year from birth. Yoshida and Ohshima(1996) detail the development of the periph-eral capillaries and their relationship to odon-
484 PHILIP C. J. DONOGHUE
FIGURE 2. Development of vascular architecture of Wistar rat, from A, E19; through B, E19.5; C, E20; D, E20.5; E,E21; F, E21.5; to G, E22. All figures reproduced from Yoshida 1991. Reproduced with the permission of the authorand publisher, Springer-Verlag.
485SKELETAL EVOLUTION OF DEVELOPMENT
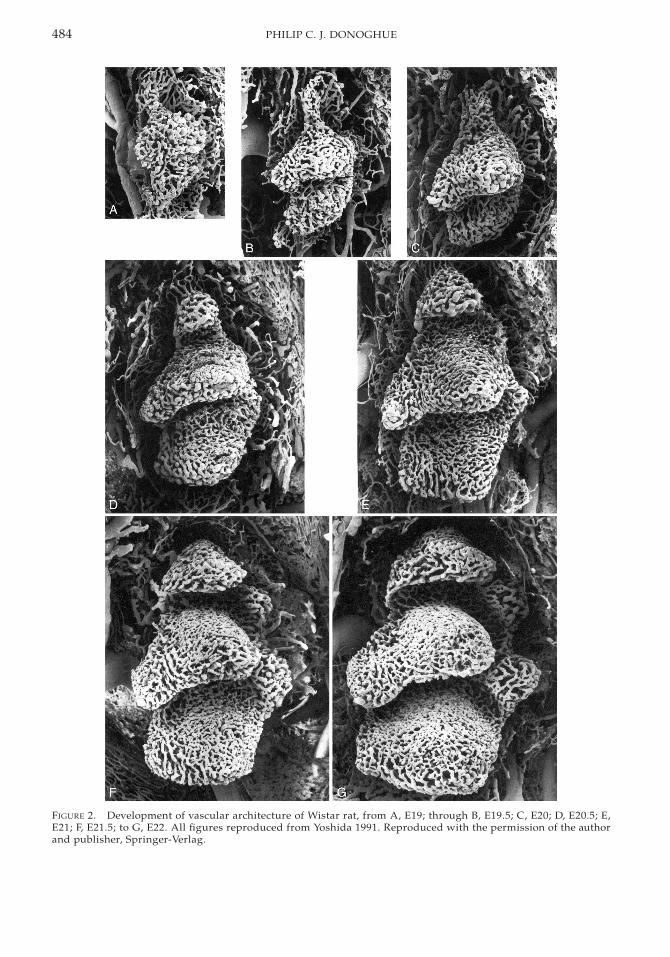
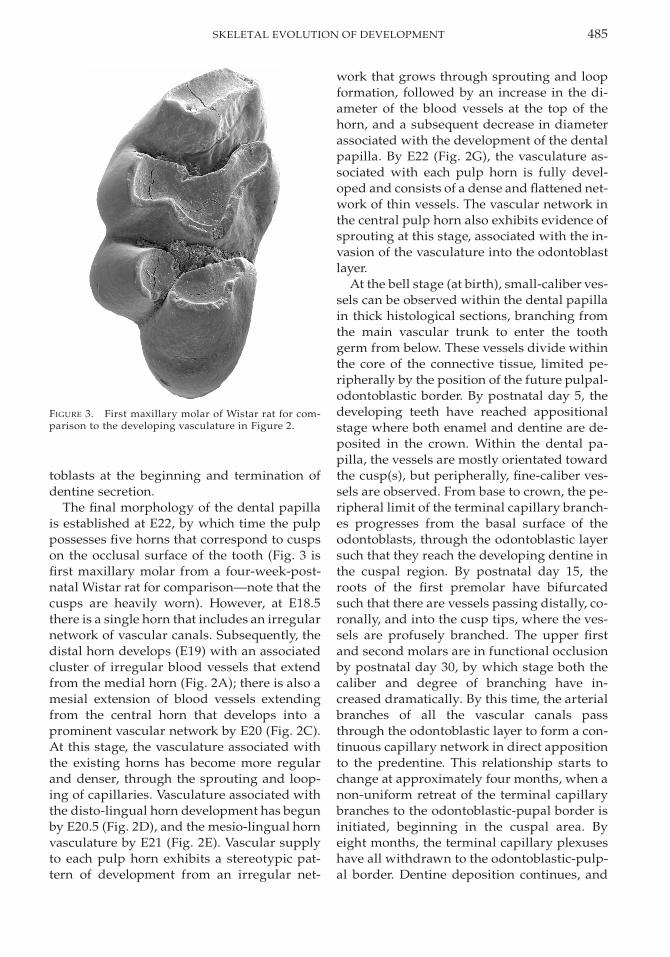
FIGURE 3. First maxillary molar of Wistar rat for com-parison to the developing vasculature in Figure 2.
toblasts at the beginning and termination ofdentine secretion.
The final morphology of the dental papillais established at E22, by which time the pulppossesses five horns that correspond to cuspson the occlusal surface of the tooth (Fig. 3 isfirst maxillary molar from a four-week-post-natal Wistar rat for comparison—note that thecusps are heavily worn). However, at E18.5there is a single horn that includes an irregularnetwork of vascular canals. Subsequently, thedistal horn develops (E19) with an associatedcluster of irregular blood vessels that extendfrom the medial horn (Fig. 2A); there is also amesial extension of blood vessels extendingfrom the central horn that develops into aprominent vascular network by E20 (Fig. 2C).At this stage, the vasculature associated withthe existing horns has become more regularand denser, through the sprouting and loop-ing of capillaries. Vasculature associated withthe disto-lingual horn development has begunby E20.5 (Fig. 2D), and the mesio-lingual hornvasculature by E21 (Fig. 2E). Vascular supplyto each pulp horn exhibits a stereotypic pat-tern of development from an irregular net-
work that grows through sprouting and loopformation, followed by an increase in the di-ameter of the blood vessels at the top of thehorn, and a subsequent decrease in diameterassociated with the development of the dentalpapilla. By E22 (Fig. 2G), the vasculature as-sociated with each pulp horn is fully devel-oped and consists of a dense and flattened net-work of thin vessels. The vascular network inthe central pulp horn also exhibits evidence ofsprouting at this stage, associated with the in-vasion of the vasculature into the odontoblastlayer.
At the bell stage (at birth), small-caliber ves-sels can be observed within the dental papillain thick histological sections, branching fromthe main vascular trunk to enter the toothgerm from below. These vessels divide withinthe core of the connective tissue, limited pe-ripherally by the position of the future pulpal-odontoblastic border. By postnatal day 5, thedeveloping teeth have reached appositionalstage where both enamel and dentine are de-posited in the crown. Within the dental pa-pilla, the vessels are mostly orientated towardthe cusp(s), but peripherally, fine-caliber ves-sels are observed. From base to crown, the pe-ripheral limit of the terminal capillary branch-es progresses from the basal surface of theodontoblasts, through the odontoblastic layersuch that they reach the developing dentine inthe cuspal region. By postnatal day 15, theroots of the first premolar have bifurcatedsuch that there are vessels passing distally, co-ronally, and into the cusp tips, where the ves-sels are profusely branched. The upper firstand second molars are in functional occlusionby postnatal day 30, by which stage both thecaliber and degree of branching have in-creased dramatically. By this time, the arterialbranches of all the vascular canals passthrough the odontoblastic layer to form a con-tinuous capillary network in direct appositionto the predentine. This relationship starts tochange at approximately four months, when anon-uniform retreat of the terminal capillarybranches to the odontoblastic-pupal border isinitiated, beginning in the cuspal area. Byeight months, the terminal capillary plexuseshave all withdrawn to the odontoblastic-pulp-al border. Dentine deposition continues, and

486 PHILIP C. J. DONOGHUE
the reduced volume of the pulp cavity resultsin convolution of the blood vessels, which aresignificantly reduced in number and of coars-er caliber. The architecture of the pulp cavityis also reduced to a series of coarse channelsthat surround the remaining blood vessels.Advance and retreat of the vascular networkrelative to the odontoblasts is positively relat-ed to the activity of the odontoblasts (Yoshidaand Ohshima 1996).
Experimental Data on Dental Patterning andMorphogenesis. Although a significant timeperiod has elapsed since Reif’s main work ontooth and scale morphogenesis, we remain ina position where little is known regarding theregulation of morphogenesis in the dermalskeleton. In contrast, there has been an explo-sion in our understanding of molecular basisof patterning and morphogenesis of teeth, andthis can be used specifically to test hypothesesof concrescence versus differentiation in theorigin of complex heterodonty in early mam-mal evolution.
The earliest stages of tooth development aremarked by thickening of the epithelium andby condensation of mesenchyme that has beendemonstrated to be of neural crest origin inmouse (Lumsden 1984, 1987, 1988) and am-phibian (Chibon 1966, 1967, 1970; Cassin andCapuron 1979). A series of epithelial-mesen-chymal interactions ensue in which mesen-chyme induces oral epithelium to proliferate,which in turn induces the proliferation of den-tal mesenchyme and the development of thedental papilla, which induces dental epitheli-um to form an enamel organ including prea-meloblasts; the preameloblasts induce differ-entiation of cells of the dental papilla intopreodontoblasts and, ultimately, odontoblaststhat induce preameloblasts to differentiateinto ameloblasts, which synthesize and de-posit enamel and induce odontoblasts to syn-thesize and deposit dentine (see, e.g., Lums-den 1987). The roles of epithelium and mes-enchyme in tooth development have been ex-tensively studied (Butler 1995; Thesleff et al.1995), but the source of the initial inductivesignal remains equivocal. In particular, Pax9expression has been implicated, as it specifi-cally marks the mesenchymal regions of allteeth prior to any morphological manifesta-
tion of development (Neubuser et al. 1997),and Pax9-deficient mice lack all teeth (amongother developmental defects [Peters et al.1998]). Furthermore, temporal changes in Pax9expression can now be integrated with classicstudies demonstrating the relative roles of oralepithelium and mesenchyme in murine toothinitiation (Kollar and Baird 1969; 1970; Kollarand Mina 1987, 1991; Lumsden 1988). Theseexperiments demonstrated that initially (E9–11) oral epithelium has the potential to inducetooth formation, after which time the induc-tive potential is transferred to the underlyingmesenchyme. These observations can be cor-related with the results of tissue-recombina-tion experiments, which indicate that untilE11.5 epithelial signals are required to main-tain the expression of Pax9 in mesenchyme(Neubuser et al. 1997). The spatial control onPax9 mesenchymal expression appears to becontrolled by the antagonistic effects of Bmp4and Bmp2 on the inducing activity of Fgf8.Thus, Neubuser et al. (1997) have proposedthat Pax9 is induced only at sites in which Fgf8is expressed in the overlying epithelium andwhere Bmp4/2 signaling does not interferewith the Pax9-inducing activity of Fgf8.
Tooth shape itself is dictated by the mor-phology of the dental epithelium and, thus,tooth morphogenesis is effectively regulatedby the controls on epithelial growth and dif-ferentiation (Fig. 4). Initial development oftooth buds in the epithelium is associatedwith the appearance of transient signalingcenters in the epithelium whose formation ap-pears to be regulated by mesenchymal signal-ing (Keranen et al. 1998). Although the life his-tory of these signaling centers is poorly un-derstood, Bmp4 (Vainio et al. 1993; Bei andMaas 1998) and Activin bA (Ferguson et al.1998) have been implicated in the initiation ofbud formation; signals in all four signaling-molecule families are expressed by the sig-naling center. Subsequent growth of the den-tal epithelium into a caplike structure occursthrough invagination about its tip such thatthe lateral margins of the developing epithe-lium extend into the mesenchyme to surroundthe developing dental papilla. This transitionis controlled by a second signaling center,known as the primary enamel knot; this knot
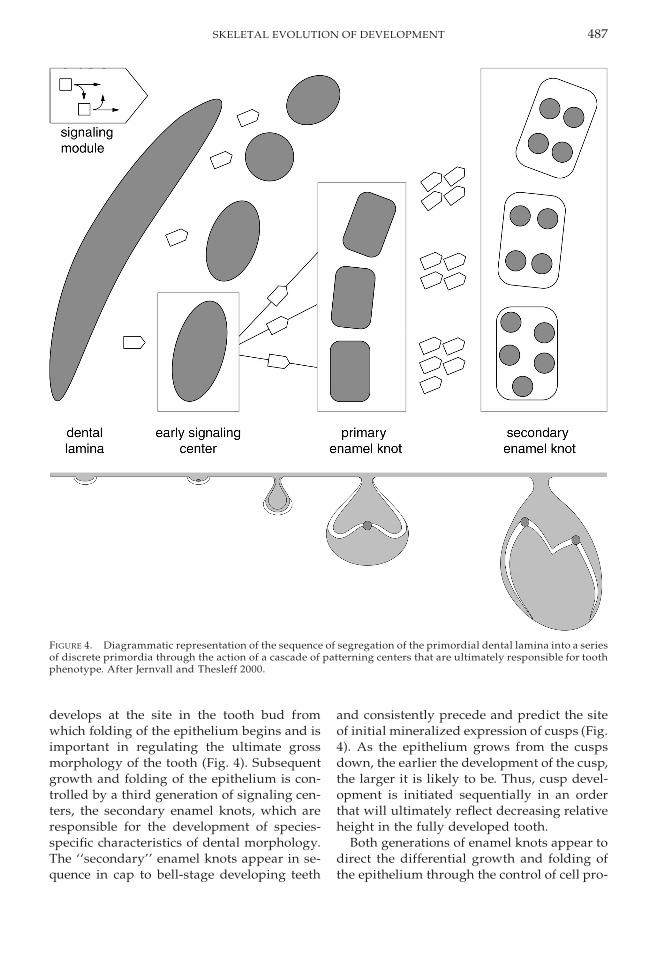
487SKELETAL EVOLUTION OF DEVELOPMENT
FIGURE 4. Diagrammatic representation of the sequence of segregation of the primordial dental lamina into a seriesof discrete primordia through the action of a cascade of patterning centers that are ultimately responsible for toothphenotype. After Jernvall and Thesleff 2000.
develops at the site in the tooth bud fromwhich folding of the epithelium begins and isimportant in regulating the ultimate grossmorphology of the tooth (Fig. 4). Subsequentgrowth and folding of the epithelium is con-trolled by a third generation of signaling cen-ters, the secondary enamel knots, which areresponsible for the development of species-specific characteristics of dental morphology.The ‘‘secondary’’ enamel knots appear in se-quence in cap to bell-stage developing teeth
and consistently precede and predict the siteof initial mineralized expression of cusps (Fig.4). As the epithelium grows from the cuspsdown, the earlier the development of the cusp,the larger it is likely to be. Thus, cusp devel-opment is initiated sequentially in an orderthat will ultimately reflect decreasing relativeheight in the fully developed tooth.
Both generations of enamel knots appear todirect the differential growth and folding ofthe epithelium through the control of cell pro-

488 PHILIP C. J. DONOGHUE
FIGURE 5. Schematic representation of the expressiondomains of Barx-1, Msx-1, Bmp-4, and Fgf-8. In the de-veloping mandibular arch of the mouse. After Tucker etal. 1998.
liferation by mitogens, including members ofthe Fgf signal family (Kettunen and Thesleff1998) around the non-dividing cells of theenamel knot. In vitro studies of the effect ofBmp4 beads upon isolated dental epithelium(Jernvall et al. 1998) have implicated Bmp4 inboth the induction and the demise of enamelknots.
Because a common suite of genes is in-volved in the development of all cusps, it isunlikely that cusp-specific locational infor-mation determines the identity of specificcusps (Keranen et al. 1998). Rather, the se-quential inception of cusps is envisaged as theproduct of a patterning cascade wherein dif-ferences in tooth morphology arise solelyfrom heterochronic patterns of cusp initiation(Weiss et al. 1998). Further, it has been consid-ered that all teeth in a dentition are serial ho-mologs (Stock et al. 1997), and differences be-tween specific teeth in a dentition arise fromlocal modifications in the expression patternsof shared developmental regulatory genes.This is supported by evidence of homeoticchanges across tooth classes, such as the mo-larization of premolars in horses (Butler 1978;see also Butler 1967). However, the signifi-cance of such changes is difficult to determinein the absence of data on their underlying ba-sis. Recently, this view of teeth as serial ho-mologs has received experimental supportfrom a study in which transformation of toothidentity from incisor to molar was achievedthrough Noggin-mediated inhibition of Bmp4signaling in the distal mesenchyme of murinemandibular arch (Tucker et al. 1998). Bmp4otherwise inhibits expression of Barx1, re-stricting it to proximal presumptive molarmesenchyme (Fig. 5), and the experimental in-
hibition of Bmp4 resulted in a distal expansionof the Barx1 expression domain, resulting in arelative transformation of mesenchymal iden-tity from incisor to molar. This transformationis limited, however, to a narrow temporal win-dow prior to E11. Furthermore, this study of-fers a refinement of the widely appreciatedview that tooth identity is conferred by mes-enchyme (Kollar and Baird 1969, 1970) by im-plying that neural crest cells in the mandibu-lar arch are equally responsive to epithelialsignals and, thus, are not prepatterned but arespecified by contact with epithelial signals.This equates well with evidence that neuralcrest cells participating in the formation of thefirst branchial arch are derived from the mid-brain and are not pre-patterned—in contrastto neural crest cells involved in the develop-ment of the other branchial arches, which arederived from the hindbrain and are patternedby members of the Hox family of homeobox-containing genes (Lumsden et al. 1991). Nev-ertheless, the first branchial arch does expressa number of homeobox genes including Msx1/2, Dlx1/2/3/5/6/7, Barx1, Otlx2, Lhx6/7 in dis-tinct spatial patterns prior to morphologicalmanifestation of tooth development. This hasled to the suggestion of an odontogenic ho-meobox code (Sharpe 1995; Thomas et al.1998; Thomas and Sharpe 1998; Tucker andSharpe 1999) in which the dental classes aredefined by spatial combinatorial expression ofsome of the homeobox-containing genes thatare expressed in the developing jaw. Thomaset al. (1998) suggested that molar expressiondomains are patterned by Barx1, and by Dlx1/2 in the upper molars versus Dlx5/6 in thelower molars, and the incisor expression do-main is defined by the expression of Msx1/2.
Discussion
Vascular Architecture and Morphology. Al-though Stensio and Ørvig (1951–1957; Ørvig1951; Stensio 1961, 1962) appear to have beenunaware of the contemporary work on dentalvascular development, dramatic comparisonscan be drawn between their observations andinferences on placoid scales, and what isknown of vascular architecture and its devel-opment in mammalian dentitions. For instance,detailed developmental studies corroborate,

489SKELETAL EVOLUTION OF DEVELOPMENT
rather than refute, Lepkowski’s (1901) viewthat the correlation between tooth cusps andthe more or less discrete vascular loops suggestthat teeth develop from a number of discreteunits. And if it were assumed, as did Stensio(1961, 1962), that vascular canals provide alandmark for identifying homology that is in-dependent of concrescence, the multicuspedteeth of mammals would be homologous to anumber of distinct teeth or scales in less-de-rived vertebrates (e.g., Ameghino 1884). How-ever, data on the development of the vascula-ture (outlined above) raise several reasons forconcern. First, the vascular architecture of teethis extremely dynamic, both increasing and de-creasing in its extent during development. Fur-thermore, the assumption that pulp cavity ar-chitecture faithfully reflects vascular architec-ture (Stensio and Ørvig 1951–1957) is un-founded not only because of the dynamicnature of the vascular network, but also be-cause the pulp cavity contains other systemssuch as nerve networks, odontoblasts, and thepulp itself. Finally, the different vascular loopsdevelop interdependently, such that they budsuccessively in a serial cascade, rather than ex-hibiting separate developmental histories asmight be expected of discrete developmentalunits. Data on the development of the vascularnetwork itself indicate that there is reason todoubt the reliability of vascular architecture inidentifying homology.
Data on the morphogenesis of the tooth it-self reveal that morphology is conferred andestablished after the discrete identity of eachtooth primoridium has been established,through a hierarchical series of patterningcenters. Hence, the presumptive tooth germsof single-cusped teeth have equipotential todevelop into multicusped teeth, at least withina narrow window of early development, andthis identity is conferred by gene expressionwithin the epithelial field, rather than by anyinnate clone-patterning. Unfortunately, nodata on the vascular architecture of the ho-meotic shift from presumptive incisor to mo-lar phenotype from Tucker et al. (1998) areavailable and, thus, it is not possible to testthis null model. Nevertheless, the parallel se-quential budding characteristic of largest tosmallest tooth cusps and vascular horns in-
dicate that the two are closely correlated.However, data on vascular development indi-cate that the development of the vascular ar-chitecture follows the establishment of toothmorphology, as determined by the final fold-ing of the dental epithelium and the differ-entiation of the odontoblasts that the vascularnetwork supplies. Further, the architecture ofthe vascular network is determined by the ar-chitecture of the pulp cavity, and subsequentchanges in the vascular architecture occur inresponse to the narrowing of the pulp cavity,as well as to decreased activity of the odon-toblasts. Thus, it appears most likely that thearchitecture of the dental vascular system isconstrained not by independent patterningbut by the morphology of the tooth itself; thus,it is not phylogenetically constrained anddoes not provide us with a means of discrim-inating homology independently of morphol-ogy, as required by the lepidomorial theory.With the rejection of its underlying scheme ofhomology, the lepidomorial theory collapses.
Ontogenetic Concrescence and Differentiation.An understanding of the morphogenetic basisof tooth development provides a means oftesting both Stensio’s use of morphology as aguide to homology in the composition of teethand scales and Reif’s assumption that teethand scales develop through differentiationrather than a mechanism of concrescence, asargued by the lepidomorial theory. The dataindicate that although multicuspid mamma-lian teeth do indeed develop from, and arepatterned by, separate signaling centers (sec-ondary enamel knots), they do so only aftersecondary differentiation of a coherent toothgerm. No existing ontogenetic developmentaldata can be interpreted in support of the viewthat complex teeth arose phylogeneticallythrough the amalgamation of numerous teethof simple morphology (contra Ameghino1884, 1896, 1899; Bolk 1912; Kukenthal 1893;Rose 1892; Stensio 1961). Instead, the cuspswithin the developing tooth arise as the resultof a patterning cascade of control centers thatultimately direct the position, and timing, ofboth onset and offset of development. Weiss etal. (1998; Zhao et al. 2000) have gone so far asto argue that although tooth cusps can be ho-mologized within and between taxa, the lack

490 PHILIP C. J. DONOGHUE
of specific genetic coding for their develop-ment is sufficient to cast doubt on the under-lying basis of their homology.
Given that tooth development—from theinitial development of the dental lamina,through its patterning and division into dis-tinct tooth classes and, ultimately, patterningof the specific teeth—is a process of progres-sive differentiation (a point emphasized by ex-perimental division of tooth germs; [Glas-stone 1952; Coin et al. 2000]), there does notappear to be any scope for a mechanism of on-togenetic concrescence. This process supportsthe axiom of the odontode regulation theorythat assumes that changes in morphology areachieved through the differentiation of toothgerms. Indeed, on this basis it is possible toreject the implicit axiom of the lepidomorialtheory, that all change occurs through onto-genetic concrescence, as well as the principlethat ontogenetic concrescence is possible.
Teratological examples of putative dentalconcrescence, (concrescence, connation, dou-ble teeth, fusion, gemination, incomplete di-chotomy, odontopagy, and synodonty) cover amultitude of types, from teeth that are fusedtogether solely by cementum or by their roots,to, more rarely, teeth with fused crowns (e.g.,Smith et al. 1953; Hitchin and Morris 1966;Miles and Grigson 1990; Law et al. 1994; Pav-lica et al. 2001). There is considerable debateover their etiology in the literature, though itwould appear that the most substantive ex-amples result from the failure of differentia-tion of dental papillae, rather than from theconcrescence of differentiated papillae (al-though there is some evidence of herniation ofenamel organs in closely spaced developingteeth, resulting in connate teeth that are par-tially joined by the enamel layer but maintaindistinct pulps and remain recognizable as dis-tinct teeth). This finding is in accord with theobservation that examples of ‘‘concrescence’’do not cross the primary dental divisions inmammals.
Phylogenetic Concrescence and Differentiation.Even though concrescence, as an ontogeneticdevelopmental pattern, can be rejected, thisdoes not preclude the possibility of concres-cence in a phylogenetic context. However, be-fore the question of how such patterns can be
constrained, the issue of how a pattern of con-crescence could be produced through onto-genetic developmental mechanisms that pre-clude ontogenetic concrescence will be ex-plored.
Given that the differentiation of the dentallamina, into the stereotypical tooth suites ofincisor, canine, and molar, is progressive, theonly conceivable mechanism of (evolutionary)concrescence is through the lack/failure of in-hibitory process that underlies differentiation.In the model system, rodent teeth, spatial con-trol on development is conferred by Bmp2 andBmp4 inhibition of the Pax9-inducing ability ofFgf8. Thus, evolutionary concrescence of oth-erwise distinct dentitions could occur throughthe absence of Bmp2/4 expression. Such amechanism has been demonstrated in pattern-ing of another epithelial-appendage organo-genic system, the feather array (a particularlyappropriate analogy for squamation pattern-ing). Feather primorida are induced and pat-terned by the antagonistic effects of Bmp2/4 onFgf4 (Fig. 6A,B). Fgf4-soaked beads placedonto explants of the developing feather arrayresult in a local breakdown in placode spacingand the development of fewer, larger featherprimordia (Jung et al. 1998). Jung et al. (1998)and other authors (Peterkova et al. 2000) in-terpret this as fusion of placodes, but in on-togenetic developmental terms, it is a failureof differentiation that provides a mechanismfor patterns of concrescence apparent fromcomparative phenotypes at the intra- or inter-taxon level.
The question of how to distinguish whetherloss (differentiation) or fusion (concrescence)best explains phylogenetic patterns in Recentand/or extinct organisms is both old and prob-lematic. One of the best examples concerns theidentification of homologies in the evolution ofdermal bone patterns in the skull roofs of os-teichthyans. Homologous skull roof bones areidentified by their relationship to adjoiningbones whose homologies can similarly be con-strained. Additional lines of evidence can bemarshaled, including the relationship of indi-vidual bones to lateral line canals or grooves, alink that is apparently supported by evidencethat developing neuromasts of the lateral linesactually induce the development of dermal
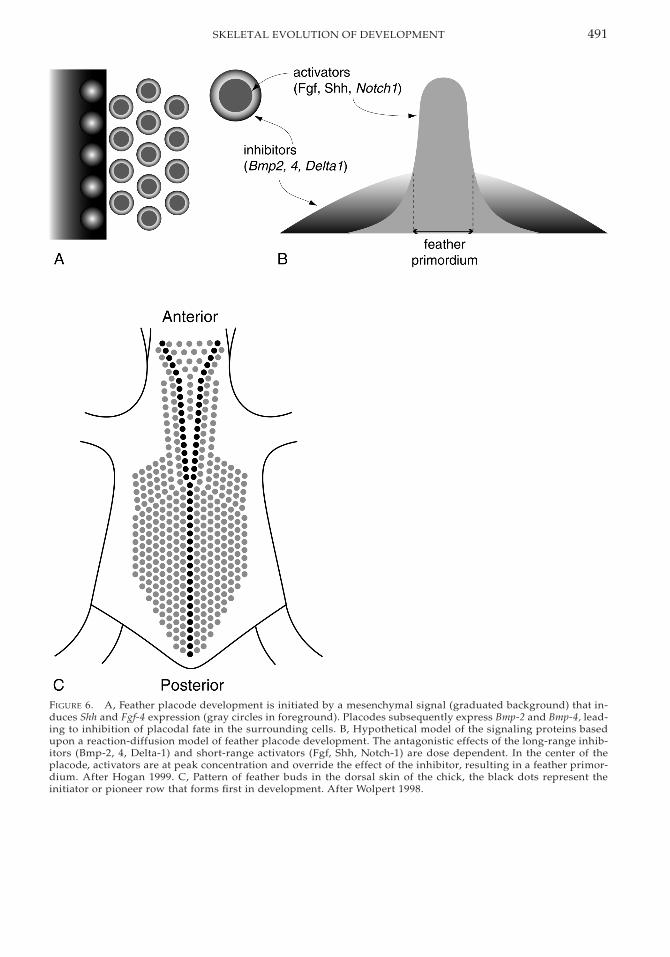
491SKELETAL EVOLUTION OF DEVELOPMENT
FIGURE 6. A, Feather placode development is initiated by a mesenchymal signal (graduated background) that in-duces Shh and Fgf-4 expression (gray circles in foreground). Placodes subsequently express Bmp-2 and Bmp-4, lead-ing to inhibition of placodal fate in the surrounding cells. B, Hypothetical model of the signaling proteins basedupon a reaction-diffusion model of feather placode development. The antagonistic effects of the long-range inhib-itors (Bmp-2, 4, Delta-1) and short-range activators (Fgf, Shh, Notch-1) are dose dependent. In the center of theplacode, activators are at peak concentration and override the effect of the inhibitor, resulting in a feather primor-dium. After Hogan 1999. C, Pattern of feather buds in the dorsal skin of the chick, the black dots represent theinitiator or pioneer row that forms first in development. After Wolpert 1998.

492 PHILIP C. J. DONOGHUE
bones (Allis 1888; Pehrson 1922; Devillers1947). Association of dermal bones with char-acteristics of the neurocranium (Westoll 1938,1943; Romer 1941, 1945; Parrington 1967; Gra-ham-Smith 1978) as well as the functional rolesof differing elements (Thomson 1993) havebeen cited as sources of phylogenetic data thatcan help constrain homology of the differentbony plates that constitute the skull roof. Nev-ertheless, despite the many and varied poten-tial landmarks and guides that may assist inconstraining homology, the increase or de-crease in the number of dermal elements form-ing the skull comes down to the same old prob-lem. That is, in the example of a numerical de-crease, did two (or more) plates fuse together,or has one plate been suppressed and its ter-ritory invaded by an adjacent bone (or bones)?Jardine (1969) wrestled with this problem andoutlined the different approaches that may betaken in attempting to resolve between phylo-genetic loss and fusion. For instance, we mayconsider the development of dermal bone pat-terns within a well-constrained phylogeneticcontext (Fig. 7A,B). However, we are never inpossession of a complete phylogenetic series,and the discovery of additional intermediateforms is likely to upset any hypothesis of phy-logenetic loss or fusion. This obtains regardlessof whether hypotheses are formulated withinan ancestor-descendent framework or com-posed from reconstructed character states ofhypothetical common ancestors in a cladisticframework.
As Jardine (1969) argues, this difficultymight be resolved if phylogenetic change isconsidered not in terms of sequential changesin adult morphology, but as modifications inontogenetic processes. Thus, we might onlyconsider phylogenetic fusion should there beevidence of fusion in an ontogenetic sequence(Fig. 7C). However, this shifts the balance ofevidence required such that the burden ofproof lies with fusion, and loss (and hence dif-ferentiation) supersedes fusion as the pre-ferred explanation in all instances where suchproof is not forthcoming. This is problematicbecause loss is concluded even in the absenceof evidence. Potentially more problematic,however, is the assumption that changes in theontogenetic program are more prevalent at
late, rather than early, developmental stages.This ‘‘biogenetic law’’ (Haeckel 1866) has fa-mously been refuted on the basis that changesin development most often appear to havebeen effected through the alteration, ratherthan in the superseding, of developmentalprograms (Sedgwick 1894; de Beer 1958;Gould 1977). Thus, the ontogenetic-phyloge-netic link still cannot preclude the possibilitythat progenetic, peramorphic changes in thetiming of fusion of dermal bones during de-velopment will be misinterpreted as loss anddifferentiation in the absence of proof of fu-sion.
In practice, many authors have simplyadopted an axiom that reduction in the totalnumber of cranial bones reflects either loss(Watson 1921; Moy-Thomas 1938; Parrington1949, 1956, 1967; Romer 1945) or fusion (Sten-sio 1921, 1947; Save-Soderbergh 1933, 1935,1941; Jarvik 1944, 1948, 1950, 1952). Westoll(1938, 1943, 1949) took the approach that al-though phylogenetic fusion may occur, it didso only within the subsets of lateral-line-bear-ing bones or bones that do not encompass lat-eral lines.
Clearly, it is extremely difficult to reconcilewhether phylogenetic differentiation or con-crescence is responsible for a decrease in thenumber of skeletal elements. This is vividly ap-parent from the example of osteichthyan cra-nial bones where, in almost all instances, theindividual elements can be determined as ho-mologs of elements of cranial bones in other in-dividuals and other taxa. Even in mammalianteeth, where individual teeth are both morpho-logically and positionally distinctive, identify-ing precise homologies where there has beenan increase or decrease in the number of teeth,and thereby reconciling phylogenetic differen-tiation and concrescence, can be impossible(Bateson 1892, 1894; Van Valen 1964, 1982).Thus, the situation in much of the remainder ofthe dermal skeleton, where individual ele-ments are indistinguishable morphologicallyor topologically, ensures that discussion of pat-terns such as fusion or loss are entirely vacu-ous. An example of such an intractable prob-lem is provided by a developmental study offeather patterning. Feathers, like teeth andscales, develop as epithelial appendages and,
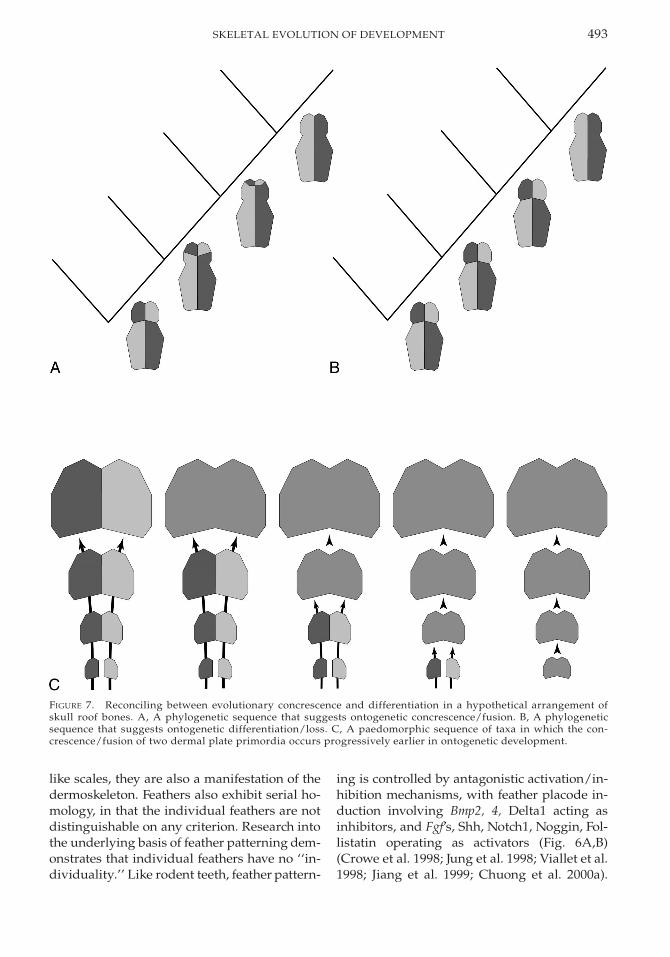
493SKELETAL EVOLUTION OF DEVELOPMENT
FIGURE 7. Reconciling between evolutionary concrescence and differentiation in a hypothetical arrangement ofskull roof bones. A, A phylogenetic sequence that suggests ontogenetic concrescence/fusion. B, A phylogeneticsequence that suggests ontogenetic differentiation/loss. C, A paedomorphic sequence of taxa in which the con-crescence/fusion of two dermal plate primordia occurs progressively earlier in ontogenetic development.
like scales, they are also a manifestation of thedermoskeleton. Feathers also exhibit serial ho-mology, in that the individual feathers are notdistinguishable on any criterion. Research intothe underlying basis of feather patterning dem-onstrates that individual feathers have no ‘‘in-dividuality.’’ Like rodent teeth, feather pattern-
ing is controlled by antagonistic activation/in-hibition mechanisms, with feather placode in-duction involving Bmp2, 4, Delta1 acting asinhibitors, and Fgf’s, Shh, Notch1, Noggin, Fol-listatin operating as activators (Fig. 6A,B)(Crowe et al. 1998; Jung et al. 1998; Viallet et al.1998; Jiang et al. 1999; Chuong et al. 2000a).

494 PHILIP C. J. DONOGHUE
These act randomly within an initially uni-formly competent feather field and their activ-ity is such that the resulting pattern conformsto close packing. Thus, the precise (and, to adegree, the relative) position of a feather pri-mordium, or interprimordium, is an issue ofprobability rather than predetermination(Jiang et al. 1999). The limited available data onthe underlying basis of scale patterning (Quil-hac 2000) suggest that this system is compa-rable to patterning of the feather array, andtherefore these conclusions have broad rele-vance.
Thus, even when the number of elementsconstituting an array of feathers or a squa-mation of scales remains constant, the absenceof distinguishing characteristics of individualelements, in terms of either morphology or to-pology, precludes the possibility of identify-ing homologous elements between one indi-vidual, or taxon, and another. Attempts to dis-criminate between phylogenetic patterns ofconcrescence or differentiation among seriallyhomologous elements are therefore entirelyfutile. However, before we all give up, it isworth examining the nature of conclusionssuch as phylogenetic concrescence and differ-entiation. Other than in the context of ances-tor-descendent relationships, the identifica-tion of ‘‘phylogenetic’’ concrescence or differ-entiation has no real meaning, because theydo not describe actual evolutionary events butmerely articulate the nature of the relationshipbetween one condition and another. And, giv-en the intangible nature of ancestor-descen-dent relationships, we should not, therefore,be despondent that we cannot reconcile phy-logenetic concrescence and differentiation.
Homology. A significant problem is inher-ent in the concept of homology as it is appliedto studying the evolution of development ofthe vertebrate dermo-visceral skeleton: pre-cisely what is meant by the assertion that teethand scales are homologous? Similarities ofcomposition and development compose thedata marshaled to support this, the ‘‘odonto-de’’ hypothesis. This is homology in the senseof the ‘‘biological homology’’ concept (Roth1988; Wagner 1989), rather than in the histor-ical or phylogenetic sense, as has classicallybeen perceived by the term ‘‘homolog’’ since
it was ‘‘evolutionized’’ by Darwin. Biologicalhomologs are useful in terms of individual-izing morphogenetic anatomical buildingblocks, and in attempting to reconcile why dif-ferent organs with very different phylogenetichistories have such similar developmentalbackgrounds (Wagner 1999). However, biolog-ical homologs are of limited use if the objec-tive is to trace the evolution, or the evolutionof development, of a phenotypic structurethrough phylogeny, as it is here. Critically, bi-ological homologs lack the phylogenetic con-straint that identifies ‘‘historical’’ homologs asthe ‘‘same,’’ rather than merely ‘‘similar,’’ indifferent individuals. Thus, stating that teethand scales are the same (i.e., homologous) onthe basis of their development is little morethan a truism; as a theory it makes no predic-tions and implies nothing more than the for-mulation on which it is based. This stands ir-respective of hypotheses contending thatteeth evolved from dermal scales, for example,through co-option of a dermal scale cover thatinvaded the mouth (cf. Halstead Tarlo andHalstead Tarlo 1965), or through heterotopy(cf. Hall 1998).
Research into the underlying basis of thesecomparisons, traditionally considered ‘‘serialhomologs,’’ has revealed that homology is en-tirely contingent upon the dynamic quantita-tive, temporal, and spatial interaction of vari-ous signaling factors. This has been offered asa counsel of despair by some developmentalbiologists who doubt that, in the absence of di-rect genetic control of morphogenesis, anysuch comparisons can represent true homo-logs (Zhao et al. 2000). However, we must re-member the distinction between explanans andexplanandum in our concept of homology. Theoperational criteria on which we propose ho-mology have changed little since the conceptwas first formalized outside of an evolution-ary context by Owen. A hypothesis of homol-ogy is the explanandum, the phenomenon thatis to be explained; evolution provides the ex-planans, the explanation of the phenomenon.But evolution provides only one explanatoryperspective on the phenomenon and devel-opment must provide the explanation of theontogenetic mechanism that underlies thesimilarities and differences between proposed

495SKELETAL EVOLUTION OF DEVELOPMENT
homologs. When this explanation is not whatwas originally expected, this does not erasethe phenomenon but suggests that our under-standing of the mechanisms that underlie con-served similarity are not quite as universal asoriginally thought.
It is tempting to conclude from the relative-ly random nature in which the topological po-sition of serially homologous elements are pat-terned that our inability to recognize homol-ogy within serially homologous elements isnot simply a result of limitations in compar-ative anatomy, but a reflection of fact (cf. Jianget al. 1999; Chuong et al. 2000a). However, thediscovery that individually identifiable ho-mologous elements have a common develop-mental basis in patterning and morphogenesisas serially homologous elements (that, by def-inition, lack individual identity), togetherwith the recognition that identity can be con-ferred entirely epigenetically, demonstratesthat homology is all about identity. Thus, ascomparative anatomists, we are limited solelyby our ability to individualize identity.
Toward a Universal Theory of theEvolution of Development of the Vertebrate
Dermoskeleton and Dentition?
A single universal theory of the evolution ofdevelopment of the vertebrate dermal and oralskeleton is possible. However, it is clear fromthe experimental data outlined above thatsuch a theory does not lie in the direction ofthe lepidomorial theory. The ontogenetic de-velopment of the dermal and visceral skeletonfollows a pattern of progressive differentia-tion and, thus, there is no scope for concres-cence, other than in a phylogenetic context.The lepidomorial and odontode regulationtheories concern homology only at a concep-tual level—comparative analysis of an ab-straction of structure, development, and pat-terning in teeth and scales. This is because inalmost all instances, it is simply not possibleto identify the same element in the squama-tion or dentition of different individuals ordifferent taxa. And given the lack of homolo-gy of individual elements in the dermal andoral skeleton of most vertebrates, attempts todiscriminate between patterns of concres-cence and differentiation are futile and vacu-
ous. An assertion that differentiation is a moreparsimonious interpretation of phylogeneticpatterns than is concrescence, as argued by theodontode regulation theory (Reif 1982a: p.348), implies certainty and data where thereare none. These problems notwithstanding,Reif’s odontode regulation theory is close to auniversal theory of the evolution of develop-ment of the vertebrate dermal and oral skele-ton. However, Reif’s ‘‘differentiation theorycan only explain changes in numbers of scalesand teeth but not changes in number’’ (Reif1982a: p. 348), and a weakness of the theorylies in its inability to trace homology belowthe level of the odontode. The experimentaldata summarized herein indicate that theodontode regulation theory can be supple-mented with the axiom that patterning, aswell as morphogenesis, is achieved throughdifferentiation. Thus, there is no homology be-low the level of the odontode. It is in manyways unfortunate that we cannot go furtherand attempt to explain changes in the numberof elements constituting the squamation anddentition of vertebrates, but such subjects lieoutside of the sphere of scientific enquiry. Auniversal theory of the evolution of develop-ment of the vertebrate dermal and oral skele-ton should therefore be restricted in its remitto addressing ontogenetic development (cf.Reif 1980a) and its relation to phylogeny.
The Molecular Basis of RegulatoryPatterning Mechanisms
Setting aside the characteristics of the basicpatterning unit in the dermoskeleton and den-tition, the odontode regulation theory alsoconcerns the regulatory mechanisms respon-sible for induction, arrangement, and main-tenance through ontogeny of skeletal elementswithin these skeletal systems, with varyingdegrees of success. Although Reif addressedthe problem of how topological informationmay be conveyed in the developing squama-tion or dentition, he was unable to arrive atany satisfactory conclusions. Similarly, hemade little advance in understanding the in-duction and establishment of squamations, al-though he undertook significant work on theestablishment of dental patterning in sharks.One area in which Reif did make significant

496 PHILIP C. J. DONOGHUE
headway was in developing a regulatorymechanism for the local spatial and temporalrelationship between individual elements.The ‘‘zone of inhibition’’ model (Reif 1980a)became the central regulatory mechanism inthe ‘‘odontode regulation theory,’’ and itholds that the development of individual teethand scales is constrained by lateral ‘‘zones ofinhibition’’ that preclude the development ofnew skeletal units in the vicinity of existing el-ements, both during initial development and,subsequently, in the regulation of patterningduring replacement. This was developed fromReif’s earlier work on tooth replacement (Reif1976), squamation ontogeny (Reif 1974), andwound repair in the dermoskeleton (Reif1978c). In the case of scales, Reif observed thatthe regular close-packing arrangement pre-cluded development of additional scales un-less new space was created over an above-the-normal interscale spacing, either through anincrease in surface area of the dermis concom-itant with volumetric growth of the animal,through shedding of old scales, or throughwounding in which portions of the dermiswere removed. In extant sharks, space in thesquamation is taken up by new scales that de-velop between, and distinct from, those al-ready present. However, in Paleozoic sharks,and many other groups, new space can be tak-en up through the augmentation of existingscales with new scales that either are attachedto the side of or completely envelope the ex-isting scales. The inhibitory field concept wasdeveloped to account for the available facts re-lating to the ontogeny of the dermal skeletonand dentition (Reif 1974, 1976, 1978c, 1980a).However, considering modifications to its tim-ing extent and interactions with other mech-anisms, such as shedding, Reif (1982a,b) ex-tended the model as a mechanistic explana-tion for the differences between the universeof dermoskeletal and dental conditions en-countered among living and fossil lower ver-tebrates. Although Reif did not speculate onthe basis of such zones of inhibition, he sug-gested that they would function through dif-fusion from the developing and adult scale ortooth. Taking scales as an example, the radialextent of the inhibition zone relative to the ra-dius of the scale structure determines the
proximity of contemporaneous scales to oneanother, and new scales can be initiated onlyif portions of the dermis fall outside the pe-rimeter of overlapping inhibition zones exert-ed by adjacent scales. This degree of complex-ity accounts only for the maintenance of squa-mations in which the component scales are ofnongrowing type, as in modern shark scales.To account for growing scales, which is themost general condition met with in lower ver-tebrates, Reif (1980a) suggested that the extentand influence of zones of inhibition might de-grade over time and, depending upon the‘‘half-life’’ of the inhibition zone relative to thelifetime of the animal, growth of the squa-mation would keep pace with surface area in-crease in the dermis, through either scale aug-mentation (growing scales) in the case of ashort half-life or the addition of discretely po-sitioned scales in the case of a long half-life.
Reif’s inhibitory zone model is entirely em-pirically based, and although it has yet to betested with data other than those on which themodel was originally based, it provides notonly an excellent explanation of the availablefacts, but also a mechanistic explanation ofhow the universe of dental and dermoskeletalconditions encountered among fossil and liv-ing lower vertebrates might appear. How doesthis compare to the regulatory mechanismsunderlying development in better-understoodsystems? As has already been seen, develop-ment of the chick feather array exhibits a num-ber of very detailed parallels to tooth devel-opment. But many of the phenomena that Reifattempted to account for in the dermoskeletonand dentition of sharks and other lower ver-tebrates are the same as those classically ad-dressed in experimental attempts to under-stand the establishment and regulation of thechick feather array during development. Thus,further comparison may prove insightful inassessing Reif’s model of inhibition zones reg-ulating scale and tooth patterning. Further-more, data on the regulatory basis of devel-opment in the chick feather array may providea new perspective on the dermoskeleton in ar-eas where the odontode regulation theory isdelimited, such as in terms of how positionalinformation is conveyed during development

497SKELETAL EVOLUTION OF DEVELOPMENT
and in maintenance during subsequent ontog-eny.
The pattern of induction of the overall feath-er array is quite different from scale inductionin Reif’s model system, living sharks. For in-stance, Reif’s documentation of scale induc-tion in carcharinid sharks indicates that allfirst-generation scales within a given regionform simultaneously and are originally ran-domly arranged (Reif 1980b). Reif (1980b)contrasted this with the situation in the feath-er array, where feather buds first appear as asingle dorsal longitudinal ‘‘initiator’’ row, or‘‘file,’’ after which successive rows are addeddorsolaterally (Fig. 6C). Subsequent work hasdemonstrated that the appearance of the ini-tiator file is preceded in development by ex-pression of a single solid strip of low-levelFgf4 and Shh expression within the dermis,which is subsequently subdivided into a rowof cell groups that exhibit high-level expres-sion of these genes and, subsequently, of Bmp2and Bmp4 in both the epidermis and meso-derm. The same antagonistic interactive roleof these genes in delimiting the developingdentition has been replicated through the ec-topic application of their transcripts and im-plicated in the developing feather array (Junget al. 1998). Shh and Fgf4 promote feather plac-ode induction as short range activators,whereas Bmp’s, emanating from the samepoint source, act as long-range inhibitors con-straining feather bud shape and preventinginduction of new, adjacent feather buds (Fig.6A,B). Feather buds are induced and their sizeis regulated through the concentration-depen-dent interaction of activators and inhibitorssuch that over-threshold concentration of theactivator (Fgf4) relative to inhibitor (Bmp’s) cir-cumscribes the feather placode; the remainingarea of below-threshold activator adopts thefate of an interbud area (Fig. 6A,B). Featherbud development is further constrained viacell-cell signaling molecules such as in theNotch-Delta pathway, and although the fullrepertoire has yet to be determined, as haveprecise roles of those already implicated,Crowe et al. (1998) have, for instance, dem-onstrated that Delta1 both promotes featherbud development in the overlying ectodermand inhibits placode fate laterally. Feather in-
duction is ultimately constrained by a wave ofcompetence that, within the primordial stripe,is at first concentrated at the posterior end andsubsequently propagates anteriorly, and thenlaterally, allowing the cells to react to the ac-tivators; the timing and polarity of compe-tence differs for different regions of the der-mis.
Although there are clear parallels to the in-duction of complex deciduous tooth families,Reif (1980b) ruled out the relevance of the de-veloping feather array to understanding squa-mation induction in sharks because of the dif-ference between tightly regulated patterningin the chick and apparently topologically ran-dom induction of scales in carcharinids. It isnot possible to test this comparison any fur-ther for lack of developmental data, but it isinteresting to note that the condition in livingsharks is quite unrepresentative of lower ver-tebrates (‘‘fishes’’) in general and is possiblyunique. In all other groups of lower verte-brates with a dermal skeleton, and for whichdata are available, induction of the squamationbegins with a row or field of pioneer scale pri-mordia (Fig. 8A–D). Although the topology ofthe pioneer primordia varies, in many taxa itis the rows of lateral line scales, for example,in acanthodians (Watson 1937; Zidek 1985),sarcopterygians (Neoceratodus forsteri [AnneKemp personal communication 2002]), basalactinopterygians (‘‘palaeonisciforms’’ [Schul-tze and Bardack 1987; Hutchinson 1973]), andteleosts (Salmo [Jollie 1984], cyprinids [Mc-Crimmon and Swee 1967; Andrews 1970],Brachydanio rerio [Armstrong 1973; Sire et al.1997]). Among the exceptions, scale row in-duction sweeps posterior to anterior with a‘‘,’’ front (apex oriented rostrally) and eitheris initiated by the lateral line (e.g., Pomoxis[Ward and Leonard 1952]; Aplodinotus [Priegel1966]), initiates as a mid-lateral rostro-caudalrow independent of the lateral line (e.g., Cen-trarchus [Conley and Witt 1966]), or otherwiseinitiates with scales associated with fin bases(Pomoxis [Cooper 1971]). Variation notwith-standing, there is clear evidence of a commontheme among lower vertebrates, bar livingsharks, of scale induction beginning with asingle pioneer row or field, as in the chickfeather array. Indeed, just as regulation of the

498 PHILIP C. J. DONOGHUE
FIGURE 8. Squamation development in the Triassic actinopterygian Brookvalia gracilis Wade (basal crown-groupactinopterygian, stem-group actinopteran). The specimens are reproduced at constant frame width (and, thus, dif-ferent magnifications) such that they best convey the sequence of squamation development; note that the scales ofboth sides of the body are preserved within the same plane. Specimens from The Natural History Museum, London.A, A 42-mm individual exhibiting a single, partially developed paired row of scales flanking the main lateral line;NHM P15808; note that this image has been digitally reversed along horizontal axis to maintain an orientationconsistent with the other figured specimens. B, A 52-mm individual exhibiting a single complete paired row, and

499SKELETAL EVOLUTION OF DEVELOPMENT
←
the rostral end of three new ventrolateral scale rows; NHM P15806. C, A 58-mm individual exhibiting four or fivecomplete scale rows, in both ventrolateral and dorsolateral positions, with the rostral portion of a number of ad-ditional scale rows; NHM P24721. D, An 82-mm individual with an almost complete squamation; NHM P15800.
chick feather array is distinct in different partsof the body, patterns of scale induction appearto belie distinctly regulated body and finsquamation fields. Although there are veryfew data on squamation development in Pa-leozoic sharks, there is some evidence to sug-gest that the condition met with in livingsharks is a derived peculiarity. Microverte-brate collections indicate that the earliestknown chondrichthyans had an extensive der-moskeleton (Sansom et al. 1996), but most Pa-leozoic chondrichthyans known from articu-lated remains, for example, Stethacanthus(Lund 1985) and Diademodus (Zangerl 1981),almost entirely lack a dermoskeleton, exceptfor scales in the head region and in associationwith the lateral line, i.e., those scale rows thatappear first during development and are mostlikely to be retained upon reduction of thedermoskeleton during phylogeny. Althoughthese taxa appear to be basal stem-holoce-phalans (Coates and Sequeira 2001) and adultprimitive chondrichthyans were fully envel-oped by dermal scales (Young 1982), Stetha-canthus and its kin nevertheless provide a con-straint on squamation ontogeny among basalchondrichthyans. Thus, at least plesiomorph-ically, it is likely that chondrichthyans alsoconform to a pattern of squamation develop-ment that is general for crown-group gnathos-tomes at the very least. The establishment ofthe main squamation in lower vertebrates as awhole therefore appears to be more compa-rable to the chick feather array than Reif(1980b) envisaged in his study of carcharinidsalone.
Experimental manipulation of developmentin the chick feather array also provides in-sights into how positional information may beconveyed in patterning the squamation suchthat the correct scale morphology develops inthe correct topological position. Heterotopicgrafting of chick paraxial mesoderm and over-lying ectoderm results in the development ofa feather pattern that is characteristic of theorientation and axial level from which the
graft was removed (Mauger 1972). This im-plies that there is an underlying patterningmechanism conferring positional informationalong the anterior-posterior axis during de-velopment of derivatives of paraxial meso-derm (dermotome). Axial patterning of scler-otomal derivatives of paraxial mesoderm suchas the vertebral (endo)skeleton of amniotes,where positional and, thus, morphologicalidentity is conveyed, is conferred by the Hoxfamily of homeotic genes (Gaunt 1994; Burkeet al. 1995). Recent in situ hybridization ex-periments have confirmed that some Hoxtranscripts maintain structural colinearityeven after the dermatome-derived dermalcells are in position, in both chick and mouse(Kanzler et al. 1994, 1997; Reid and Gaunt2002) (although there are differences betweenthese taxa in terms of which Hox genes main-tain structural colinearity). Structural colin-earity of the dorsolateral dermis of chick hasso far been explored in relatively few Hoxgenes, and among these only a small fractionhave been observed to follow this rule (Reidand Gaunt 2002). Nevertheless, expression do-mains of at least one of these three (Hoxb4,Hoxa7, Hoxc8) is known to coincide with thetransition between two different morphologi-cal compartments of spinal pteryla (Reid andGaunt 2002).
The role of Hox genes in axial patterninghas not been demonstrated among ‘‘fishes,’’although they exhibit comparable, albeit con-densed, expression boundaries in the zebra-fish (Prince et al. 1998) that correlate to con-sistent patterns of axial innervation (Burke etal. 1995; Burke 2000). Further, the role of Hoxparalogs in axial patterning of arthropods in-dicates that this is a symplesiomorphic char-acter of bilaterians (Carroll 1995; Tabin et al.1999). Intriguingly, phylogenetic constraint ontheir roles in development within the chordatelineage is conferred by Branchiostoma, whereHox genes are not expressed in paraxial me-soderm (Garcia-Fernandez and Holland 1994)and, thus, expansion of the role of Hox genes

500 PHILIP C. J. DONOGHUE
in patterning the dermo- and axial skeletalsystems must be a craniate or vertebrate syn-apomorphy (cf. Sharman and Holland 1998),occurring within the gnathostome stem line-age at the very latest (cf. Coates and Cohn1998). This does not by any means provide thedefinitive answer to how positional identity isconveyed to the developing dermoskeleton,and many other regulatory genes must be im-plicated in constraining topologically depen-dent expression of the dermoskeleton, but itdoes go some way to providing an under-standing of how topological information isconferred. In the case of the dentition, Hoxgenes are not implicated because their expres-sion domains do not extend rostrally as far asthe mandibular arch and, as has already beendiscussed, other regulatory gene families,such as Barx, Dlx, Lhx, Msx and Otlx havebeen implicated in providing spatial identityduring dental development (e.g., Sharpe1995), at least within mammals.
There are thus clear parallels between pat-terns of induction, development, and patternregulation of the chick feather array and scalesquamation of lower vertebrates, sufficient, in-deed, to use the better-known feather array asa model for assessing the veracity of Reif’spattern regulation mechanism. Good evidenceexists for a molecular basis to Reif’s ‘‘zones ofinhibition’’; the mechanics that underlie thephenomena that Reif described are more com-plex than he envisaged, however, and involvea number of regulatory mechanisms concern-ing the progressive restriction and refinementof originally uniform expression domains of avariety of signaling molecules, to eventuallycircumscribe the sites of primordial elements.Reif’s implicit assumption, that the epitheliumis uniformly competent at the outset and thatperiodic patterns self-organize from an ini-tially random state, seems at best oversimpli-fied and, at worst, unjustified. Although theinitial expression domains may be uniform,they do not initially encompass the entire de-veloping field, and they undergo progressiverestriction before the cell in which the signal-ing molecules are expressed achieves compe-tence (cf. Jung et al. 1998). Further, althoughperiodic patterning appears to be self-orga-nizing, this is true only to a degree. Jung et al.
(1998) argue that the available evidence indi-cates that patterning is achieved in the mannerenvisaged by Turing (1952), through the effectof random interaction between multiple pointsources of differentially diffusing activatorsand inhibitors. The overall patterning of thefeather array is, nevertheless, tightly con-strained by compartmentalization into dis-tinct fields, in which induction proceeds onlyafter achieving competence in a controlledand progressive pattern of induction.
The only molecular data on scale develop-ment are limited to regeneration (Quilhac 2000),but they nevertheless indicate that the samegenes involved in induction and development ofthe initial feather array in the chick are also in-volved in the induction and patterning of re-placement scales in the zebrafish. It is not so ob-vious how the chick model could be interpretedto account for patterns of growth akin to thegrowing scales of Paleozoic sharks and mostother ‘‘fish’’ groups; this problem is particularlysignificant given that scale ‘‘growth’’ appears tobe the plesiomorphic condition for the dermo-skeleton (Donoghue and Sansom 2002). How-ever, in this matter it should be rememberedthat scale ‘‘growth’’ is just a modified form ofscale replacement in which the existing scale re-mains in place and the ‘‘replacement’’ scaleadopts a position adjacent to the existing scalerather than a distinct position between existingscales. A model akin to Reif’s (1980a) diminish-ing inhibition zone could be developed from theactivation-inhibition model of feather array pat-terning to account for this phenomenon throughincreasing the relative concentration of the ac-tivator to inhibitor and, thus, expanding thezone of scale induction beyond the limits of theexisting scale. This model would stand in con-trast to the regular pattern of scale replacementin which a portion of the dermis falls outside theinfluence of inhibitors as a result of growth ofthe animal. Increased topological dominance ofthe activator could be achieved through increas-ing absolute concentration of the activator or de-creasing inhibitor-dosage, and depending uponwhether this effect was polarized or equant,scale augmentation would occur through mar-ginal or areal growth.
Does the developmental model provided bythe chick feather array provide a better frame-

501SKELETAL EVOLUTION OF DEVELOPMENT
work for understanding the evolution of thevertebrate dermoskeleton from a developmen-tal perspective than does Reif’s inhibitionzone theory? This is not an either-or debate;Reif’s theory is entirely empirically based andthe main mechanisms that underlie it are alsoa grossly simplified view of the regulatory ba-sis of feather array development. More appro-priately, the model of feather array patterningand induction should be used as a predictivemodel in experimental analyses of squama-tion development in lower vertebrates. Initialforays into constraining the molecular basis ofsquamation and scale development have be-gun with the zebrafish (Quilhac 2000). This isan ideal model animal in which to test the ve-racity of the chick model in detail becausemany or all of the regulatory genes implicatedin development of the feather array have al-ready been cloned in the zebrafish. The rele-vance of this reaches beyond the understand-ing of dermoskeletal development to examin-ing the phylogenetic distribution of regulato-ry gene networks responsible for patterningperiodic elements and exploring the nature ofevolution and homology within such net-works.
Concluding Remarks
As mentioned in the introduction, this con-tribution has been restricted, almost exclu-sively, to testing established theories that seekto explain the evolution of development of thevertebrate dermal and oral skeleton. Thus, incommon with these theories, it has addresseddirectly only the odontogenic component ofthe oral and dermal skeleton. Research intothe molecular patterning controls on the de-velopment of the skeletogenic system remainsat a relatively early stage and the regulatorygene networks have yet to be explored. How-ever, what little is known (see, e.g., Opperman2000 for cranial sutures; Quilhac 2000 for theonly available data on patterning of elasmoidscales) suggests that such development is con-strained by comparable controls; furthermore,many of the examples and data used in thisanalysis demonstrate that the same patterningphenomena encountered in the odontogenicsystem are also found in products of the ske-letogenic system. Thus, we may extend the
conclusions reached above to the skeletoge-netic scleroblastic system, with the caveat thatthe odontogenic and skeletogenic systems areoften independently patterned (judging fromempirical evidence from the distribution oftheir products in living and fossil vertebrates[e.g., Westoll 1967]). And although both sys-tems are ubiquitous in the dermal skeleton ofthe extinct groups of ‘‘armored’’ jawless ver-tebrates in which the skeletal system firstevolved, the odontogenic and skeletogenicsystems are, in large part, mutually exclusivein the dermoskeletons of living vertebrates(exceptions include the lower actinopterygi-ans and lower sarcopterygians). Attempts totrace the evolution of development of the der-mal and oral skeleton have, in the past, con-flated these two scleroblastic systems. Futureanalyses must unravel the contribution ofthese two distinct systems and the relative in-fluence that patterning of either system hasupon the final skeleton.
The first attempts to test the applicability ofpatterning models based on other epithelial-appendage systems, such as teeth, feather,lung etc., on the patterning of the dermal skel-eton have begun (Opperman 2000; Quilhac2000). It will be interesting to see how similarthe underlying basis of patterning and mor-phogenesis seen in scales is to that in their pu-tative closest system, the oral skeleton, ascompared to other organ systems within theepithelial appendage family (Chuong 1998).Wider taxonomic analysis of model systemslies at the heart of our attempts to unravel theevolution of developmental systems. But it isonly in combination with further explorationof the diversity of organogenic systems withinthe epithelial-appendage family that will webegin to discover whether their similarity indevelopment results from conservation (e.g.,genetic piracy Roth 1984) or more simply, aconservatism in developmental programs.
Acknowledgments
First and foremost I would like to expressmy deep gratitude to C. Franzen-Bengtson(Department of Palaeozoology, Swedish Mu-seum of Natural History) who took greatpains to locate the original manuscript on thelepidomorial theory and the development of

502 PHILIP C. J. DONOGHUE
placoid scales by E. Stensio and T. Ørvig. G.Sperber (University of Alberta), P. Butler (Uni-versity of London), and A. Smith (Universityof Birmingham) provided sources of literatureon the development of dental vasculature inrodents and humans. P. E. Ahlberg, P. L. Forey(both Natural History Museum, London), andA. Kemp (University of Queensland) provideddata, material, and/or sources of literature ofsquamation development. A. Smith and G.Smith (University of Birmingham) providedand prepared Wistar rat molar teeth for anal-ysis; J. Jernvall (Helsinki University) and W. E.Reif (Tubingen) provided comments on adraft manuscript; P. Janvier (Museum Nation-al d’Histoire Naturelle, Paris) and B. K. Hall(Dalhousie University) provided helpful re-views of the submitted manuscript. Images inFigure 1 were reproduced from Stensio 1961and Reif 1982a with the permission of TorontoUniversity Press and Plenum/Kluwer Pub-lishing Corporation, respectively. Figure 2was reproduced from Yoshida 1991 with kindpermission of the author and Springer-Verlag.Finally, I would like to express my gratitudeto I. J. Sansom (University of Birmingham) forenduring incessant discussion on this topicover the past few years. This work was fundedby Natural Environment Research Councilpostdoctoral research fellowship GT5/99/ES/2.
Literature CitedAdloff, P. 1916. Die Entwicklung des Zahnsystems der Sauge-
tiere und des Menschen. Verlag von Hermann Meusser, Ber-lin.
Agassiz, L. 1833–1843. Recherches sur les Poissons Fossiles. Im-primerie de Petitpierre, Neuchatel.
Ahlberg, P. E., ed. 2001. Major events in vertebrate evolution:palaeontology, phylogeny, genetics and development. Taylorand Francis, London.
Allis, E. P. 1888. On the morphology of certain of the bones ofthe cheek and snout of Amia. Journal of Morphology 1:425–466.
Ameghino, F. 1884. Filogenia.———. 1896. Sur l’evolution des dents des mammiferes. Boletın
de la Academia Nacional de Ciencias, Cordoba, Argentina 14:381–517.
———. 1899. On the primitive type of the plexodont molars ofmammals. Proceedings of the Zoological Society, London1899:555–571.
Andrews, A. K. 1970. Squamation chronology of the flatheadminnow, Pimephales promelas. Transactions of the AmericanFisheries Society 99:429–432.
Andrews, S. M., R. S. Miles, and A. D. Walker, eds. 1977. Prob-lems in vertebrate evolution. Linnean Society Symposium Se-ries 4. Academic Press, London.
Armstrong, J. G. 1973. Squamation chronology of the zebrafish(Cyprinidae), Brachydanio rerio. Copeia 1973:823–824.
Atchley, W. R., and B. K. Hall. 1991. A model for developmentand evolution of complex morphological structures and itsapplication to the mammalian mandible. Biological Reviews66:101–157.
Bateson, W. 1892. On numerical variation in teeth, with a dis-cussion of the concept of homology. Proceedings of the Zoo-logical Society, London 1892:102–115.
———. 1894. Materials for the study of variation treated withespecial regard to discontinuity in the origin of species. Mac-millan, London.
Bei, M., and R. Maas. 1998. FGFs and Bmp4 induce both Msx1-independent and Msx1-dependent signaling pathways in ear-ly tooth development. Development 125:4325–4333.
Bernick, S. 1960. Vascular supply to the developing teeth of rats.Anatomical Record 137:141–151.
———. 1962. Ages changes in the blood supply to molar teethof rats. Anatomical Record 144:265–274.
Bolk, L. 1912. On the structure of the reptilian dentition and itsrelationship to the mammalian dentition. Anatomischer An-zeiger 41(Suppl.).
———. 1922. Odontological essays. 4. On the relation betweenreptilian and mammalian teeth. Journal of Anatomy 56:107–136.
Burke, A. C. 2000. Hox genes and the global patterning of thesomitic mesoderm. Current Topics in Developmental Biology47:155–181.
Burke, A. C., B. A. Nelson, and C. Tabin. 1995. Hox genes andthe evolution of vertebrate axial morphology. Development121:333–346.
Butler, P. M. 1967. Dental merism and tooth development. Jour-nal of Dental Research 46:845–850.
———. 1978. The ontogeny of mammalian heterodonty. Journalde Biologie Buccale 6:217–227.
———. 1995. Ontogenetic aspects of dental evolution. Interna-tional Journal of Developmental Biology 39:25–34.
Carroll, S. B. 1995. Homeotic genes and the evolution of arthro-pods and chordates. Nature 376:479–485.
Cassin, C., and A. Capuron. 1979. Buccal organogenesis in Pleu-rodeles waltii Michah (urodele amphibian), study by intrablas-tocoelic transplantation and in vitro culture. Journal de Biol-ogie Buccale 7:61–76.
Chibon, P. 1966. Analyse experimentale de la regionalisation etdes capacites morphogenetiques de la crete neurale chezl’amphibien urodele Pleurodeles waltll Michah. Memoires de laSociete Zoologique de France 36:1–107.
———. 1967. Marquage nucleaire par la thymidine tritee desderives de la crete neurale chez l’amphibien urodele Pleuro-deles waltll Michah. Journal of Embryology and ExperimentalMorphology 18:343–358.
———. 1970. Capacite de regulation des excedents dans la creteneurale d’Amphibien. Journal of Embryology and Experimen-tal Morphology 24:479–496.
Chuong, C.-M., ed. 1998. Molecular basis of epithelial append-age morphogenesis. R. G. Landes, Austin.
Chuong, C.-M., R. Chodankar, R. B. Widelitz, and T.-X. Jiang.2000a. Evo-Devo of feather and scales: building complex ep-ithelial appendages. Current Opinion in Genetics and Devel-opment 10:449–456.
Chuong, C.-M., N. Patel, J. Lin, H.-S. Jung, and R. B. Widelitz.2000b. Sonic hedgehog signaling pathway in vertebrate epi-thelial appendage morphogenesis: perspectives in develop-ment and evolution. Cellular and Molecular Life Sciences 57:1672–1681.
Coates, M. I., and M. J. Cohn. 1998. Fins, limbs, and tails: out-growths and axial patterning in vertebrate evolution. Bio-Essays 20:371–381.

503SKELETAL EVOLUTION OF DEVELOPMENT
Coates, M. I., and S. E. K. Sequeira. 2001. Early sharks and prim-itive gnathostomes interrelationships. Pp. 241–262 in Ahlberg2001.
Coin, R., R. Schmitt, H. Lesot, J.-L. Vonesch, and J.-V. Ruch. 2000.Regeneration of halved embryonic lower first mouse molars:correlation with the distribution pattern of non dividing IDEcells, the putative organizers of morphogenetic units, thecusps. International Journal of Developmental Biology 44:289–295.
Conley, J. M., and A. Witt. 1966. The origin and development ofscales in the Flier, Centrarchus macropterus (Lacepede). Trans-actions of the American Fisheries Society 95:433–434.
Cooper, J. A. 1971. Scale development as related to growth ofjuvenile black crappie, Pomoxis nigromaculatus Lesueur. Trans-actions of the American Fisheries Society 100:570–572.
Cope, E. D. 1883. On the trituberculate type of molar teeth inthe Mammalia. Proceedings of the American PhilosophicalSociety 21:324–326.
———. 1889. Synopsis on the families of the Vertebrata. Amer-ican Naturalist 23:1–29.
Crowe, R., D. Henrique, D. Ish-Horowicz, and L. Niswander.1998. A new role for Notch and Delta in cell fate decisions:patterning the feather array. Development 125:767–775.
de Beer, G. R. 1958. Embryos and ancestors. Oxford UniversityPress, London.
———. 1971. Homology: an unsolved problem. Oxford Univer-sity Press, London.
Devillers, C. 1947. Recherches sur le crane dermique des teleos-teens. Annales de Paleontologie 33:1–94.
Donoghue, P. C. J., and R. J. Aldridge. 2001. Origin of a miner-alized skeleton. Pp. 85–105 in Ahlberg 2001.
Donoghue, P. C. J., and I. J. Sansom. 2002. Origin and early evo-lution of vertebrate skeletonization. Microscopy Research andTechnique 59:352–372.
D’Souza, R. N., T. Aberg, J. Gaikwad, A. Cavender, M. Owen, G.Karsenty, and I. Thesleff. 1999. Cbfa1 is required for epithe-lial-mesenchymal interactions regulating tooth developmentin mice. Development 126:2911–2920.
Ferguson, B. M., A. S. Tucker, L. Christensen, A. L. Lau, M. M.Matzuk, and P. T. Sharpe. 1998. Activin is an essential earlymesenchymal signal in tooth development that is required forpatterning of the murine dentition. Genes and Development12:2636–2649.
Francillon-Vieillot, H., V. de Buffrenil, J. Castanet, J. Gerauldie,F. J. Meunier, J.-Y. Sire, L. Zylberberg, and A. de Ricqles. 1990.Microstructure and mineralization of vertebrate skeletal tis-sues. Pp. 471–530 in J. G. Carter, ed. Skeletal biomineraliza-tion: patterns, processes, and evolutionary trends. Van Nos-trand Reinhold, New York.
Garcia-Fernandez, J., and P. W. H. Holland. 1994. Archetypal or-ganisation of the amphioxus Hox gene cluster. Nature 370:563–566.
Gaunt, S. J. 1994. Conservation of the Hox code during morpho-logical evolution. International Journal of Developmental Bi-ology 38:549–552.
Glasstone, S. 1952. The development of halved tooth germs: astudy in experimental embryology. Journal of Anatomy 87:12–15.
Goodrich, E. S. 1907. On the scales of fish, living and extinct,and their importance in classification. Proceedings of the Zoo-logical Society, London 2:751–774.
Gould, S. J. 1977. Ontogeny and phylogeny. Belknap Press ofHarvard University Press, Cambridge.
Graham-Smith, W. 1978. On the lateral lines and dermal bonesin the parietal region of some crossopterygian and dipnoanfishes. Philosophical Transactions of the Royal Society of Lon-don B 282:41–105.
Gross, W. 1973. Kleinschuppen, flossenstacheln und Zahne von
Fischen aus Europaischen und Nordamerikanischen bone-beds des Devons. Palaeontographica, Abteilung A 142:51–155.
Haeckel, E. 1866. Generelle Morphologie der Organismen: All-gemeine Grundzuge der organischen Formen-Wissenschaft,mechanisch begrundet durch die von Charles Darwin refor-mirte Descendenz-Theorie. 2 vols. Georg Reimer, Berlin.
Hall, B. K. 1998. Evolutionary developmental biology. Chapmanand Hall, London.
Halstead Tarlo, B. J., and L. B. Halstead Tarlo. 1965. The originof teeth. Discovery 26:20–26.
Hertwig, O. 1874a. Ueber Bau und Entwickelung der Placoid-schuppen und der Zahne der Selachier. Jenaische Zeitschriftfur Naturwissenschaftliche 8:221–404.
———. 1874b. Ueber das Zahnsystem der Amphibien und seineBedeutung fur die Genese des Skelets der Mundhohle. Einevergleichend anatomische, entwickelungsgeschichtliche Un-tersuchung. Archiv fur mikroskopische Anatomie 11(Suppl.).
———. 1876. Ueber das Hautskelet der Fische. MorphologischesJahrbuch 2:328–395.
———. 1879. Ueber das Hautskelet der Fische. 2: Das Hautskeletder Ganoiden (Lepidosteus und Polypterus). MorphologischesJahrbuch 5:1–21.
———. 1882. Ueber das Hautskelet der Fische. 3: Daus Haut-skelet der Pediculati, der Discoboli, der Gattung Diana, derCentriscidae, einiger Gattungen aus der Familie der Triglidaeund der Plectognathen. Morphologisches Jahrbuch 7:1–42.
Hitchin, A. D., and I. Morris. 1966. Geminated odontome-con-nation of the incisors in the dog—its etiology and ontogeny.Journal of Dental Research 45:575–583.
Hogan, B. L. M. 1999. Morphogenesis. Cell 96:225–233.Hutchinson, P. 1973. A revision of the redfieldiiform and per-
leidiform fishes from the Triassic of Bekker’s Kraal (South Af-rica) and Brookville (New South Wales). Bulletin of the BritishMuseum (Natural History), Geology 22:233–354.
Inohaya, K., and A. Kudo. 2000. Temporal and spatial patternsof cbfa1 expression during embryonic development in the tel-eost, Oryzias latipes. Development, Genes and Evolution 210:570–574.
Jardine, N. 1969. The observational and theoretical componentsof homology: a study based on the morphology of the dermalskull-roofs of rhipidistian fishes. Biological Journal of the Lin-nean Society 1:327–361.
Jarvik, E. 1944. On the dermal bones, sensory canals and pit-organs of the skull in Eusthenopteron foordi Whiteaves, withsome remarks on E. save-soderberghi Jarvik. Kungliga SvenskaVetenskapsakademiens Handlingar 21:1–48.
———. 1948. On the Middle Devonian crossopterygians fromthe Hornelen Field in Western Norway. Universitet i BergenArbok 1948.
———. 1950. Middle Devonian vertebrates from Canning Landand Wegeners Halvø (East Greenland), Part II. Crossoptery-gii. Meddelelser om Grønland 96:1–132.
———. 1952. On the fish-like tail in the ichthyostegid stegoce-phalians. Meddelelser om Grønland 114:1–90.
———. 1960. Theories de l’evolution des vertebres reconsider-ees a la lumiere des recentes decouvertes sur les vertebres in-ferieurs. Masson et Cie, Paris.
Jernvall, J. 1995. Mammalian molar cusp patterns: developmen-tal mechanisms of diversity. Acta Zoologica Fennica 198:1–61.
Jernvall, J., and I. Thesleff. 2000. Reiterative signaling and pat-terning during mammalian tooth morphogenesis. Mecha-nisms of Development 92:19–29.
Jernvall, J., T. Aberg, P. Kettunen, S. Keranen, and I. Thesleff.1998. The life history of an embryonic signaling center: BMP-4 induces p21 and is associated with apoptosis in the mousetooth enamel knot. Development 125:161–169.
Jiang, T.-X., H.-S. Jung, R. B. Widelitz, and C.-M. Chuong. 1999.

504 PHILIP C. J. DONOGHUE
Self-organization of periodic patterns by dissociated feathermesenchymal cells and the regulation of size, number andspacing of primordia. Development 126:4997–5009.
Jollie, M. 1984. Development of the head skeleton and pectoralgirdle of salmons, with a note on the scales. Canadian Journalof Zoology 62:1757–1778.
Jung, H.-S., P. H. Francis-West, R. B. Widelitz, T.-X. Jiang, S.Ting-Berreth, C. Tickle, L. Wolpert, and C.-M. Chuong. 1998.Local inhibitory action of BMPs and their relationships withactivators in feather formation: implications for periodic pat-terning. Developmental Biology 196:11–23.
Kanzler, B., J. P. Viallet, H. Le Mouellic, E. Boncinelli, D. Du-boule, and D. Dhouailly. 1994. Differential expression of twodifferent homeobox gene families during mouse tegumentmorphogenesis. International Journal of Developmental Bi-ology 38:633–640.
Kanzler, B., F. Prin, J. Thelu, and D. Dhouailly. 1997. CHOXC-8and CHOXD-13 expression in embryonic chick skin and cu-taneous appendage specification. Developmental Dynamics210:274–287.
Karatajute-Talimaa, V. 1992. The early stages of dermal skeletonformation in chondrichthyans. Pp. 223–231 in E. Mark-Kurik,ed. Fossil fishes as living animals. Institute of Geology, Tal-linn.
———. 1998. Determination methods for the exoskeletal re-mains of early vertebrates. Mitteilungen ausdem Museum furNaturkunde in Berlin, Geowissenschaftliche Reihe 1:21–52.
Keranen, S. V. E., T. Aberg, P. Kettunen, I. Thesleff, and J. Jern-vall. 1998. Association of developmental regulatory geneswith the development of different molar tooth shapes in twospecies of rodents. Development, Genes and Evolution 208:477–486.
Keranen, S. V. E., P. Kettunen, T. Aberg, I. Thesleff, and J. Jern-vall. 1999. Gene expression patterns associated with suppres-sion of odontogenesis in mouse and vole diastema regions.Development, Genes and Evolution 209:495–506.
Kettunen, P., and I. Thesleff. 1998. Expression and function ofFGFs-4, -8 and -9 suggest functional redundancy and repeti-tive use as epithelial signals during tooth morphogenesis. De-velopmental Dynamics 211:256–268.
Klaatsch, H. 1890. Zur Morphologie der Fischshcuppen und zurGeschichte der Hautsubstanzgewebe. Morphologisches Jahr-buch 16:97–196, 209–258.
Kollar, E. J., and G. R. Baird. 1969. The influence of the dentalpapilla on the developmment of tooth shape in embryonicmouse germs. Journal of Embryology and Experimental Mor-phology 21:131–148.
———. 1970. Tissue interactions in embryonic mouse toothgerms. II. The inductive role of the dental papilla. Journal ofEmbryology and Experimental Morphology 24:173–186.
Kollar, E. J., and M. Mina. 1987. The induction of odontogenesisin non-dental mesenchyme combined with early murine man-dibular arch epithelium. Archives of Oral Biology 32:123–127.
———. 1991. Role of the early epithelium in the patterning ofthe teeth and Meckel’s cartilage. Journal of Craniofacial Ge-netics and Developmental Biology 11:223–228.
Krejsa, R. J. 1979. The comparative anatomy of the integumentalskeleton. Pp. 112–191 in M. H. Wake, ed. Hyman’s compara-tive vertebrate anatomy. University of Chicago Press, Chica-go.
Kukenthal, W. 1893. Entwickelungsgeschichtliche Untersuchun-gen am Pinnipediergebisse. Jenaische Zeitschrift fur Natur-wissenschaftliche 28:76–118.
Law, L., G. Fishelberg, J. E. Skribner, and L. M. Lin. 1994. End-odontic treatment of mandibular molars with concrescence.Journal of Endodontics 20:562–564.
Lepkowski, W. 1901. Die Verteilung der Gefasse in den Zahnendes Menschen. Anat. Hefte 17:183–195.
Lumsden, A. G. S. 1984. Tooth morphogenesis: contributions ofthe cranial neural crest cells in mammals. In A. B. Belcour andJ.-V. Ruch, eds. Tooth morphogenesis and differentiation. Col-loque Inserm 125:29–40. INSERM, Paris.
———. 1987. The neural crest contribution to tooth develop-ment in the mammalian embryo. Pp. 261–300 in P. F. A. Mad-erson, ed. Developmental and evolutionary aspects of theneural crest. Wiley, New York.
———. 1988. Spatial organisation of the epithelium and the roleof neural crest cells in the initiation of the mammalian toothgerm. Development 1988(Suppl.):155–169.
Lumsden, A. G. S., N. Sprawson, and A. Graham. 1991. Seg-mental origin and migration of neural crest cells in the hind-brain region of the chick embryo. Development 113:1281–1291.
Lund, R. 1985. The morphology of Falcatus falcatus (St. John andWorthen), a Mississippian stethacanthid chondrichthyanfrom the Bear Gulch Limestone of Montana. Journal of Ver-tebrate Paleontology 5:1–19.
Maisey, J. G. 1988. Phylogeny of early vertebrate skeletal induc-tion and ossification patterns. Evolutionary Biology 22:1–36.
Mauger, A. 1972. Role du mesoderme somitique dans le devel-oppement du plumage dorsal chez l’embryon de Poulet. II.Regionalisation du mesoderme plumigene. Journal of Embry-ology and Experimental Morphology 28:343–366.
McCrimmon, H. R., and U. B. Swee. 1967. Scale formation as re-lated to growth and development of young carp, Cyprinus car-pio L. Journal of the Fisheries Research Board of Canada 24:47–51.
Miles, A. E. W., and C. Grigson. 1990. Colyer’s variations anddiseases of the teeth of animals. Cambridge University Press,Cambridge.
Moy-Thomas, J. A. 1938. The problem of the evolution of the der-mal bones in fishes. Pp. 305–319 in G. R. de Beer, ed. Evolu-tion, essays on aspects of evolutionary biology. Clarendon,Oxford.
Mundlos, S., F. Otto, J. B. Mundlos, A. S. Aylsworth, S. Albright,D. Lindhout, W. G. Cole, W. Henn, J. H. M. Knoll, M. J. Owen,R. Mertelsmann, B. U. Zabel, and B. R. Olsen. 1997. Mutationsinvolving the transcription factor CBFA1 cause cleidocranialdysplasia. Cell 89:773–779.
Neubuser, A., H. Peters, R. Balling, and G. R. Martin. 1997. An-tagonistic interactions between FGF and BMP signaling path-ways: a mechanism for positioning the sites of tooth forma-tion. Cell 90:247–255.
Opperman, L. A. 2000. Cranial sutures as intramembranousbone growth sites. Developmental Dynamics 219:472–485.
Ørvig, T. 1951. Histologic studies of ostracoderms, placodermsand fossil elasmobranchs. 1. The endoskeleton, with remarkson the hard tissues of lower vertebrates in general. Arkiv forZoologi 2:321–454.
———. 1957. Remarks on the vertebrate fauna of the lower up-per Devonian of Escuminac Bay, P. Q., Canada, with specialreference to the Porolepiform Crossopterygians. Arkiv forZoologi 10:367–427.
———. 1967. Phylogeny of tooth tissues: evolution of some cal-cified tissues in early vertebrates. Pp. 45–110 in A. E. W. Miles,ed. Structural and chemical organisation of teeth. AcademicPress, New York.
———. 1968. The dermal skeleton: general considerations. Pp.374–397 in T. Ørvig, ed. Current problems of lower vertebratephylogeny. Almquist and Wiksell, Stockholm.
———. 1975. Description, with special reference to the dermalskeleton, of a new radotinid arthrodire from the Gedinnian ofArctic Canada. In J.-P. Lehman, ed. Problemes actuels de pa-leontologie: evolution des vertebres. Colloques Internation-aux du Centre National de la Recherche Scientifique 218:41–71.

505SKELETAL EVOLUTION OF DEVELOPMENT
———. 1977. A survey of odontodes (‘dermal teeth’) from de-velopmental, structural, functional, and phyletic points ofview. Pp. 53–75 in Andrews et al. 1977.
Osborn, H. F. 1907. Evolution of mammalian molar teeth to andfrom the triangular type. Macmillan, New York.
Owen, R. 1845. Odontography. Hippotype Bailliere, London.Palmer, R. M., and A. G. S. Lumsden. 1987. Development of peri-
odontal ligament and alveolar bone in homografted recom-binations of enamel organs and papillary, pulpal and follic-ular mesenchyme in the mouse. Archives of Oral Biology 32:281–289.
Parrington, F. R. 1949. A theory of the relations of lateral linesto dermal bones. Proceedings of the Zoological Society, Lon-don 119:65–78.
———. 1956. The patterns of dermal bones in primitive verte-brates. Proceedings of the Zoological Society, London 127:389–411.
———. 1967. The identification of the dermal bones of the head.Journal of the Linnean Society (Zoology) 47:231–239.
Patterson, C. 1977. Cartilage bones, dermal bones and mem-brane bones, or the exoskeleton versus the endoskeleton. Pp.77–121 in Andrews et al. 1977.
Pavlica, Z., V. Erjavec, and M. Petelin. 2001. Teeth abnormalitiesin the dog. Acta Veterinaria Brno 70:65–72.
Pehrson, T. 1922. Some points in the cranial development of te-leostomian fishes. Acta Zoologica 3:1–63.
Peterkova, R., M. Peterka, L. Viriot, and H. Lesot. 2000. Denti-tion development and budding morphogenesis. Journal ofCraniofacial Genetics and Developmental Biology 20:158–172.
Peters, H., and R. Balling. 1999. Teeth: where and how to makethem. Trends in Genetics 15:59–65.
Peters, H., A. Neubuser, K. Kratochwil, and R. Balling. 1998.Pax9-deficient mice lack pharyngeal pouch derivatives andteeth and exhibit craniofacial and limb abnormalities. Genesand Development 12:2735–2747.
Peyer, B. 1968. Comparative odontology. University of ChicagoPress, Chicago.
Priegel, G. R. 1966. Early scale development in the freshwaterdrum, Aplodinotus grunniens Rafinesque. Transactions of theAmerican Fisheries Society 95:434–436.
Prince, V. E., L. Joly, M. Ekker, and R. K. Ho. 1998. Zebrafish hoxgenes: genomic organization and modified colinear expres-sion patterns in the trunk. Development 125:407–420.
Priolo, M., M. Silengo, M. Lerone, and R. Ravazzolo. 2000. Ec-todermal dysplasias: not only ‘skin’ deep. Clinical Genetics58:415–430.
Quilhac, A. 2000. Etudes des interactions epidermo-dermiqueslors du developpement du dermosquelette des osteichthyens:le modele ecaille. Bulletin de la Societe Zoologique de France125:321–332.
Reid, A. I., and S. J. Gaunt. 2002. Colinearity and non-colinearityin the expression of Hox genes in developing chick skin. In-ternational Journal of Developmental Biology 46:209–215.
Reif, W.-E. 1973. Ontogenese des Hautskelettes von Heterodontusfalcifer (Selachii) aus dem Untertithon. Stuttgarter Beitragezur Naturkunde B 7:1–16.
———. 1974. Morphogenese und Musterbildung des Haut-zahnchen-Skelettes von Heterodontus. Lethaia 7:25–42.
———. 1976. Morphogenesis, pattern formation and function ofthe dentition of Heterodontus (Selachii). Zoomorphologie 83:1–47.
———. 1978a. Types of morphogenesis of the dermal skeletonin fossil sharks. Palaontologische Zeitschrift 52:110–128.
———. 1978b. Shark dentitions: morphogenetic processes andevolution. Neues Jahrbuch fur Geologie und Palaontologie,Abhandlungen 157:107–115.
———. 1978c. Wound healing in sharks: form and arrangementof repair scales. Zoomorphologie 90:101–111.
———. 1980a. A model of morphogenetic processes in the der-mal skeleton of elasmobranchs. Neues Jahrbuch fur Geologieund Palaontologie, Abhandlungen 159:339–359.
———. 1980b. Development of dentition and dermal skeleton inembryonic Scyliorhinus canicula. Journal of Morphology 166:275–288.
———. 1982a. Evolution of dermal skeleton and dentition invertebrates: the odontode regulation theory. Evolutionary Bi-ology 15:287–368.
———. 1982b. Morphogenesis and function of the squamationin sharks. Neues Jahrbuch fur Geologie und Palaontologie,Abhandlungen 164:172–183.
Reif, W.-E., and M. Richter. 2001. Revisiting the lepidomorialand odontode regulation theories of dermo-skeletal morpho-genesis. Neues Jahrbuch fur Geologie und Palaontologie, Ab-handlungen 219:285–304.
Romer, A. S. 1941. Notes on the crossopterygian hyomandibularand braincase. Journal of Morphology 69:141–160.
———. 1945. Vertebrate paleontology. University of ChicagoPress, Chicago.
Rose, C. 1892. Uber die Entstehung und Formabanderungen dermenschlichen Molaren. Anatomischer Anzeiger 7:393–421.
Roth, V. L. 1984. On homology. Biological Journal of the LinneanSociety 22:13–29.
———. 1988. The biological basis of homology. Pp. 1–26 in C. J.Humphries, ed. Ontogeny and systematics. British Museum(Natural History), London.
Sansom, I. J., M. P. Smith, and M. M. Smith. 1996. Scales of thel-odont and shark-like fishes from the Ordovician. Nature 379:628–630.
Save-Soderbergh, G. 1933. The dermal bones of the head and thelateral line system in Osteolepis macrolepidotus Ag., with re-marks on the terminology of the lateral line system and on thedermal bones in certain other crossopterygians. Nova ActaRegiae Societatis Scientarium Upsaliensis, series 4, 9:1–123.
———. 1935. On the dermal bones of the head in labyrinthodontstegocephalians and primitive Reptilia with special referenceto Eotriassic stegocephalians from East Greenland. Medde-lelser om Grønland 98:1–211.
———. 1941. Notes on the dermal bones of the head in Osteolepismacrolepidotus Ag., and the interpretation of the lateral linesystem in certain primitive vertebrates. Zoologiska Bidragfran Uppsala. 20:523–541.
Schaeffer, B. 1977. The dermal skeleton in fishes. Pp. 25–54 inAndrews et al. 1977.
Schultze, H.-P., and D. Bardack. 1987. Diversity and size chang-es in palaeonisciform fishes (Actinopterygii, Pisces) from thePennsylvanian Mazon Creek Fauna, Illinois, U.S.A. Journal ofVertebrate Paleontology 7:1–23.
Sedgwick, A. 1894. On the law of development commonlyknown as von Baer’s Law, and on the significance of ancestralrudiments in embryonic development. Quarterly Journal ofMicroscopical Science 36:35–52.
Sharman, A. C., and P. W. H. Holland. 1998. Estimation of Hoxgene cluster number in lampreys. International Journal of De-velopmental Biology 42:617–620.
Sharpe, P. T. 1995. Homeobox genes and orofacial development.Connective Tissue Research 32:17–25.
Sire, J.-Y., F. Allizard, O. Babiar, J. Buourguignon, and A. Quil-hac. 1997. Scale development in zebrafish (Danio rerio). Jour-nal of Anatomy 190:545–561.
Smith, M. A., A. d. A. Bellairs, and A. E. W. Miles. 1953. Obser-vations on the premaxillary dentition of snakes with specialreference to the egg-tooth. Journal of Linnean Society (Zool-ogy) 42:260–268.
Smith, M. M., and M. I. Coates. 1998. Evolutionary origins of the

506 PHILIP C. J. DONOGHUE
vertebrate dentition: phylogenetic patterns and developmen-tal evolution. European Journal of Oral Science 106(Suppl. 1):482–500.
———. 2000. Evolutionary origins of teeth and jaws: develop-mental models and phylogenetic patterns. Pp. 133–151 in M.F. Teaford, M. W. J. Ferguson, and M. M. Smith, eds. Devel-opment, function and evolution of teeth. Cambridge Univer-sity Press, Cambridge.
———. 2001. The evolution of vertebrate dentitions: phyloge-netic pattern and developmental models. Pp. 223–240 in Ahl-berg 2001.
Smith, M. M., and B. K. Hall. 1990. Development and evolution-ary origins of vertebrate skeletogenic and odontogenic tis-sues. Biological Reviews 65:277–373.
———. 1993. A developmental model for evolution of the ver-tebrate exoskeleton and teeth: the role of cranial and trunkneural crest. Evolutionary Biology 27:387–448.
Stensio, E. A. 1921. Triassic fishes from Spitsbergen, Part I.Adolf Holzhausen, Vienna.
———. 1947. The sensory lines and dermal bones of the cheekin fishes and amphibians. Kungliga Svenska Vetenskapsaka-demiens Handlingar 24:1–195.
———. 1958. Les Cyclostomes fossiles ou Ostracodermes. Pp.173–425 in P.-P. Grasse, ed. Traite de Zoologie. Masson, Paris.
———. 1961. Permian vertebrates. Pp. 231–247 in G. O. Raasch,ed. Geology of the Arctic. University of Toronto, Toronto.
———. 1962. Origine et nature des ecailles placoıdes et desdents. In J. P. Lehman, ed. Problemes actuels de paleontologie:(evolution des vertebres). Colloques Internationaux du CentreNational de la Recherche Scientifique 104:75–85.
———. 1964. Les cyclostomes fossiles ou ostracodermes. Pp. 96–382 in J.-P. Lehman, ed. Traite de paleontologie, Vol. IV.L’origine des vertebres: leur expansion dans les eaux douceset le milieu marin, Part 1, Agnathes. Masson et Cie, Paris.
Stensio, E. A., and T. Ørvig. 1951–1957. On the scales of Edes-tids. Unpublished manuscript, Stensio archives, Swedish Mu-seum of Natural History, Stockholm.
Stock, D. W., K. M. Weiss, and Z. Zhao. 1997. Patterning of themammalian dentition in development and evolution. Bio-Essays 19:481–490.
Tabin, C. J., S. B. Carroll, and G. Panganiban. 1999. Out on alimb: parallels in vertebrate and invertebrate limb patterningand the origin of appendages. American Zoologist 39:650–663.
Thesleff, I. 2000. Genetic basis of tooth development and dentaldefects. Acta Odontologica Scandinavia 58:191–194.
Thesleff, I., A. Vaahtokari, and A.-M. Partanen. 1995. Regulationof organogenesis: common molecular mechanisms regulatingthe development of teeth and other organs. International Jour-nal of Developmental Biology 39:35–50.
Thomas, B. L., and P. T. Sharpe. 1998. Patterning of the murinedentition by homeobox genes. European Journal of Oral Sci-ence 106(Suppl. 1):48–54.
Thomas, B. L., A. S. Ticker, C. Ferguson, M. Qiu, J. L. R. Rub-enstein, and P. T. Sharpe. 1998. Molecular control of odonto-genic patterning: positional dependent initiation and mor-phogenesis. European Journal of Oral Science 106(Suppl. 1):44–47.
Thomson, K. S. 1993. Segmentation, the adult skull, and theproblem of homology. Pp. 36–68 in J. Hanken and B. K. Hall,eds. The skull, Vol. 3. Patterns of structural and systematic di-versity. University of Chicago Press, Chicago.
Tucker, A. S., and P. T. Sharpe. 1999. Molecular genetics of toothmorphogenesis and patterning: the right shape in the rightplace. Journal of Dental Research 78:826–834.
Tucker, A. S., K. L. Matthews, and P. T. Sharpe. 1998. Transfor-mation of tooth type induced by inhibition of BMP signaling.Science 282:1136–1138.
Turing, A. M. 1952. The chemical basis of morphogenesis. Pro-ceedings of the Royal Society of London B 237:37–72.
Vaahtokari, A., S. Vainio, and I. Thesleff. 1991. Associations be-tween transforming growth factor b1 RNA expression and ep-ithelial-mesenchymal interactions during tooth morphogen-esis. Development 113:985–994.
Vainio, S., I. Karavanova, A. Jowett, and I. Thesleff. 1993. Iden-tification of BMP-4 as a signal mediating secondary inductionof between epithelial and mesenchymal tissues during earlytooth development. Cell 75:45–58.
Van Valen, L. M. 1964. Nature of the supernumerary molars ofOtocyon. Journal of Mammalogy 45:284–286.
———. 1982. Homology and causes. Journal of Morphology 173:305–312.
Viallet, J. P., F. Prin, I. Olivera-Martinez, E. Hirsinger, O. Pour-quie, and D. Dhouailly. 1998. Chick Delta-1 gene expressionand the formation of the feather primordia. Mechanisms ofDevelopment 72:159–168.
Wagner, G. P. 1989. The biological homology concept. AnnualReview of Ecology and Systematics 20:51–69.
———. 1999. A research program for testing the biological ho-mology concept. Pp. 125–140 in G. R. Bock and G. Cardew,eds. Homology (Novartis Foundation Symposium 222). Wiley,Chichester, England.
Ward, H. C., and E. M. Leonard. 1952. Order of appearance ofscales in the Black Crappie, Pomoxis nigromaculatus. Proceed-ings of the Oklahoma Academy of Science 33:138–140.
Watson, D. M. S. 1921. On the coelacanth fish. Annals of theMagazine of Natural History 8:319–337.
———. 1937. The acanthodian fishes. Philosophical Transac-tions of the Royal Society of London B 228:49–146.
Weiss, K. M., D. W. Stock, and Z. Zhao. 1998. Dynamic inter-actions and the evolutionary genetics of dental patterning.Critical Reviews in Oral Biology and Medicine 9:369–398.
Westoll, T. S. 1938. Ancestry of the tetrapods. Nature 141:127–128.
———. 1943. The origin of the tetrapods. Biological Reviews 18:78–98.
———. 1949. On the evolution of the Dipnoi. Pp. 121–184 in G.L. Jepsen et al., eds. Genetics, paleontology, and evolution.Princeton University Press, Princeton, N.J.
———. 1967. Radotina and other tesserate fishes. Journal of Lin-nean Society (Zoology) 47:83–98.
Williamson, W. C. 1849. On the microscopic structure of thescales and dermal teeth of some ganoid and placoid fish. Phil-osophical Transactions of the Royal Society of London B 139:435–475.
———. 1851. Investigations into the structure and developmentof the scales and bones of fishes. Philosophical Transactionsof the Royal Society of London B 141:643–702.
Winge, H. 1883. Om Patteddyrenes Tandskifte isaer med Hen-syn til Taendernes Former. Videnskabelige Meddelelser fraDansk Naturhistoriska Forening 4:15–69.
Wolpert, L. 1998. Pattern formation in epithelial develop-ment: the vertebrate limb and feather bud spacing. Philo-sophical Transactions of the Royal Society of London B353:871–875.
Yoshida, S. 1991. A scanning electron microscope study of vas-cular development in the dental papilla of prenatal rat molars.Anatomy and Embryology 183:379–384.
Yoshida, S., and H. Ohshima. 1996. Distribution and organiza-tion of peripheral capillaries in dental pulp and their rela-tionship to odontoblasts. Anatomical Record 245:313–326.
Young, G. C. 1982. Devonian sharks from south-eastern Austra-lia. Palaeontology 25:817–843.
Zangerl, R. 1966. A new shark of the family Edestidae, Ornitho-prion hertwigi from the Pennsylvanian Mecca and LoganQuarry Shales of Indiana. Fieldiana (Geology) 16:1–43.

507SKELETAL EVOLUTION OF DEVELOPMENT
———. 1981. Handbook of Paleoichthyology, Vol. 3A. Chondrich-thyes I (Paleozoic Elasmobranchii). Gustav Fischer, Stuttgart.
Zhao, Z., K. M. Weiss, and D. W. Stock. 2000. Development andevolution of dentition patterns and their genetic basis. Pp.152–172 in M. F. Teaford, M. M. Smith, and M. W. J. Ferguson,
eds. Development, function and evolution of teeth. Cam-bridge University Press, Cambridge.
Zidek, J. 1985. Growth in Acanthodes (Acanthodii: Pisces)data and implications. Palaontologische Zeitschrift 59:147–166.