Embed Size (px)
Citation preview

Exposing hidden endemism in a Neotropical forestraptor using citizen science
ALEXANDER C. LEES1* & ROBERT W. MARTIN2
1Coordenac�~ao de Zoologia, Museu Paraense Em�ılio Goeldi, C.P. 399, CEP 66040-170, Bel�em, PA, Brazil2BirdLife International, Wellbrook Court, Cambridge, CB3 0NA, UK
Gaps in our knowledge of the geographical distribution of species represent a funda-mental challenge to biogeographers and conservation biologists alike, and are particu-larly pervasive in the tropics. Here we highlight the case of the Rufous-thighed KiteHarpagus diodon, a South American raptor commonly mapped as resident across halfthe continent. Recent observations at migration watch points have indicated it may bepartially migratory in the southernmost parts of its range. To investigate this possibility,we collated contemporary and historical specimen records, published sight records and‘digital vouchers’ – photographs and sound-recordings archived online (from citizen sci-ence initiatives) – and explored the spatiotemporal distribution of records. We wereunable to trace any documented records of this species from Amazonia during the aus-tral summer (October–March), or records from the Atlantic Forest biome during thepeak of the Austral winter (June–August), and all proven breeding records stem fromthe Atlantic Forest region. We compared this pattern with that of a ‘control’ species,the congeneric Double-toothed Kite H. bidentatus, again using specimens and digitalvouchers. For this species we found no evidence of seasonality between biomes and candisregard spatiotemporal variation in observer effort as a cause of seasonal biases. Weconsider that all populations of Rufous-thighed Kites are fully migratory, wintering inEquatorial forests in the Amazonian basin. We provide evidence that this pattern waspreviously obscured by erroneous undocumented records and poor or erroneous speci-men metadata, and its discovery was primarily facilitated by digital vouchers. This dis-covery requires a reassessment of the species’ global conservation status as an AtlanticForest breeding endemic, threatened by habitat loss and degradation, as it was previ-ously considered to be resident across large swathes of undisturbed Amazonian Foreston the Guiana Shield. The bulk of the digital voucher data used to elucidate thispattern were extracted from a Brazilian citizen science initiative WikiAves, which mayserve as a model for collating biodiversity data in megadiverse countries and helpcatalyse environmental awareness.
Keywords: forest fragmentation, historical, migration, Neotropical avifauna, raptors,undocumented records.
Gaps in our knowledge of species distribution, the‘Wallacean shortfalls’ of Lomolino and Heaney(2004), are the greatest impediment to the accu-rate assessment of the conservation status of anygiven taxon (Rodrigues et al. 2006). Even formany bird species, the best known of terrestrial
biota, this information can be very poor at anygiven scale, particularly in the tropics (Winker1996, Newbold 2010). Historically, in the absenceof systematic surveys, the only data sources avail-able with which to construct distribution mapswere the presence-only data from museum speci-mens and opportunistic citations of species in thetechnical and scientific literature (Anderson 2012).Citizen science initiatives offer much promise in
*Corresponding author.Email: [email protected]
© 2014 British Ornithologists’ Union
Ibis (2014), doi: 10.1111/ibi.12207

filling this data vacuum, enlisting the general pub-lic in gathering large volumes of scientific informa-tion (Dickinson et al. 2010). The lowlandNeotropics have long been considered to be amajor cold-spot for biodiversity data (e.g. Haffer1985, Feeley & Silman 2011), with significantnumbers of new bird species still being describedfrom South America, predominantly from thesuper species-rich and poorly inventoried Amazonbasin (e.g. Whitney & Cohn-Haft 2013). Forest-dwelling Neotropical raptors are a particularlypoorly known group (Bierregaard 1995) giventheir typically low population densities and oftencryptic sub-canopy habits.
The Rufous-thighed Kite Harpagus diodon is agood example of a raptor for which natural historyinformation is sparse. According to Birdlife Interna-tional (2014), the species occupies 3 730 000 km2
between northern Argentina and French Guianaand most major reference works indicate that it issedentary (e.g. Thiollay 1994, Ferguson-Lees &Christie 2001). However, Bildstein (2004) consid-ered the species to be an ‘irregular or local migrant’based on Hayes et al. (1994) and Ridgely andGreenfield (2001). More recently, Cabanne andSeipke (2005) observed 129 Rufous-thighed Kitesapparently in active migration during seven surveysat Itatiaia National Park, in the Brazilian state ofRio de Janeiro in March 2005. They concluded thatthis added further support for the ‘migratory statusof the Rufous-thighed Kite in the southern extremeof the species’ range’, backed up by an apparentabsence of records in the southern cone in the aus-tral winter (e.g. Hayes et al. 1994, Azevedo et al.2003) and later reinforced by observations of smallnumbers of migrating birds in Bolivia (Juhant2012). However, Cabanne and Roesler (2007)assumed northern populations to be non-migratorygiven the historical collection of juveniles inAmazonia in the austral winter; quantitative studiesfrom French Guiana that generated estimates ofpopulation density (Thiollay 1989) bolster this con-clusion. Residency for Amazonian populations hasnot been assumed by all authors; Cohn-Haft et al.(1997) considered the species to be an australmigrant to Manaus in the Brazilian Amazon, andBraun et al. (2000) listed the species as a possibleaustral migrant even in Guyana.
During a review of distribution maps for BirdlifeInternational, R.M. noticed significant inconsisten-cies in published texts for the distribution andmigratory status of the Rufous-thighed Kite. He
contacted A.C.L., who consulted the avianphoto database WikiAves www.wikiaves.com.br (arecently established Brazilian citizen-science initia-tive) in which a previously undescribed pattern oftemporal distribution was readily apparent. Herewe carry out a spatiotemporal analysis of the globaldistribution of Rufous-thighed Kite comparingresults obtained using historical data from museumspecimens with new distributional data archivedonline by an ever-growing band of amateur orni-thologists, with the purpose of evaluating the distri-bution and migratory phenology of this poorlyknown raptor.
METHODS
Data collection
We used the digital database ORNIS (www.ornisnet.org/) to search for Rufous-thighed Kite speci-mens held in 33 North American institutions,requested specimen data from the Museu de Zoolo-gia Universidade de S~ao Paulo, S~ao Paulo, Brazil(MZUSP), and the Museu Nacional Rio de Janeiro(MNRJ) and visited the collections of the MuseuParaense Em�ılio Goeldi, Bel�em, Brazil (MPEG),and the Natural History Museum at Tring, UK(NHM), to look at specimens. Collecting localitieswere located using Paynter (1982, 1989, 1992,1994, 1995), Paynter and Traylor (1991) and Ste-phens and Traylor (1985). This search of museumcollections was accompanied by a review of previ-ous published ornithological inventories listing Ruf-ous-thighed Kites from South America (seeliterature list in Supporting Information AppendixS1), a request for data from eBird http://ebird.org/and an extensive search for digital vouchersarchived online, images on WikiAves, Fauna Para-guay www.faunaparaguay.com and the Internet BirdCollection http://ibc.lynxeds.com/ and soundrecordings on xeno-canto www.xeno-canto.org andthe Macaulay Library http://macaulaylibrary.org/.The minimum requirement for the inclusion of arecord was date information to at least month andlocality below the country level. For image-basedrecords (WikiAves, Internet Bird Collection) theExchangeable image file format (Exif) metadataassociated with the images was taken as a verifica-tion of the date of a record; images lacking Exif datawere coded separately and considered to lack dateverification. We incorporated images into the analy-sis posted up until 25 February 2014. Multiple
© 2014 British Ornithologists’ Union
2 A. C. Lees & R. W. Martin

registrations from the same location within thesame month were discarded unless evidence con-firmed multiple individuals. All unequivocal evi-dence of breeding (eggs, occupied nests anddependent chicks) associated with each record wasalso coded.
To calculate estimates of the seasonal geogra-phical range of Rufous-thighed Kites, we geo-referenced all records. Convex hull minimumconvex polygons (MCPs) were created in ARCGIS
10 (ESRI, Redlands, CA, USA) around all docu-mented records (those supported by a specimen ordigital voucher) for both the peak breeding seasonand the peak non-breeding season. The peak breed-ing season was determined as the middle 4 monthswithin the duration of all confirmed breedingrecords, in order to maximize the number of recordsthat relate to actively breeding individuals ratherthose individuals arriving in breeding areas late orleaving early. The opposite 4 months on the calen-dar were considered the peak non-breeding season.A 95% convex hull MCP was generated by remov-ing the 5% of documented records most distant(Euclidean distance) from the median centre ofall documented records. The MCPs were clippedto a base layer derived from the WWF TerrestrialEcoregions of the World (Olson et al. 2001). Ker-nel density analysis to investigate the spatial distri-bution of the documented records was performedusing the ‘ks’ package in R (Duong 2014, R CoreTeam 2014) using a Gaussian kernel, the plug-inestimator for bandwidth ‘h’ (Wand & Jones1994), cell size of 0.1 decimal degrees, and un-weighted points. Using the kernel density surfacecreated, 50% and 90% isopleth contours wereproduced using the ‘isopleth’ command in soft-ware GME (Beyer 2012).
We used a Loess-smoothing algorithm in SIGMA
PLOT 10.0 (Systat Software Inc., Chicago, IL, USA)to display the underlying structure in the spatio-temporal relationships derived from different datasources. The parameters set for the Loesssmoother were span (D) of 0.4 (determining thedegree of smoothing) and polynomial degree of 1(i.e. linear local regression). We analysed recordsbased on (1) specimens, (2) undocumentedrecords (reports without accompanying testimonialmaterial such as specimen, image, recording) and(3) digital vouchers (photos, sound-recordings)separately to see whether spatiotemporal patternswere maintained between different data sources.We compared mean latitudes and longitudes of
records of Rufous-thighed Kites and Double-toothed Kite H. bidentatus in the peak breedingand peak non-breeding periods using unpaired t-tests with the GRAPH-PAD PRISM software, version4.02 (GraphPad Software, San Diego, CA, USA).
To control for potential differences in observercoverage between regions and between seasons,we also analysed records of the congeneric Dou-ble-toothed Kite, which occurs sympatrically withRufous-thighed Kites in the northern Atlantic For-est and in Amazonia, and for which there is noevidence of migratory behaviour (e.g. Thiollay1994, Schulze et al. 2000, Ferguson-Lees & Chris-tie 2001) using the same methodology. We onlycompared specimen and photo records of Double-toothed Kites from within the corresponding rangeof Rufous-thighed Kite to adjust for possible sea-sonal influence in observer distribution.
RESULTS
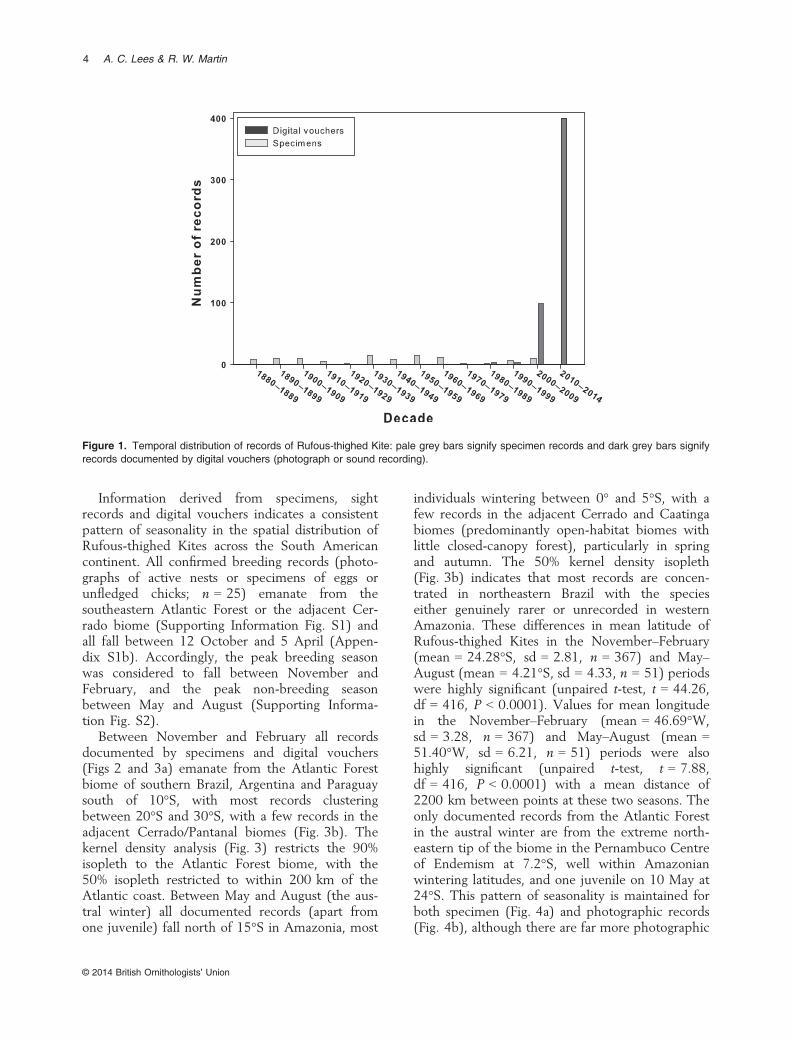
Our systematic search recovered data from 114museum specimens, 66 retrieved from ORNISfrom 13 North American institutions and 48 fromthe collections of MPEG, MZUSP, MNRJ andNHM (Supporting Information Appendix S1). Ofthese, 86 included sufficient metadata (date andplace below the country level) to permit spatio-temporal analysis. A total of 839 sight or audiorecords were obtained from online sources: 678from WikiAves, 124 from eBird, 14 from xeno-canto, six from IBC, three from Fauna Paraguay,one from the Macaulay Library and 13 from pub-lished literature (Ridgely 1980, Hayes et al. 1994,Cabanne & Seipke 2005, Cabanne & Roesler2007, Pereira et al. 2008, Klavins & Fari~na 2013).Two duplicates were then removed from xeno-canto, 174 duplicates from WikiAves and 25 dupli-cates from eBird. Of the remaining 638 sight oraudio records, two photographic records werefound to have been identified incorrectly (andwere re-assigned on the database). Specimens werecollected throughout the entire survey period1880 to the present, but digital vouchers onlyappeared in the last decade and quickly became byfar the most abundant source of information onRufous-thighed Kite distribution (Fig. 1). A totalof 722 records (15 were aural-only records)included sufficient date and locality information tobe analysed together (Appendix S1), including106 ‘undocumented’ records without supportingvoucher documentation.
© 2014 British Ornithologists’ Union
Endemism in a Neotropical forest raptor 3
Information derived from specimens, sightrecords and digital vouchers indicates a consistentpattern of seasonality in the spatial distribution ofRufous-thighed Kites across the South Americancontinent. All confirmed breeding records (photo-graphs of active nests or specimens of eggs orunfledged chicks; n = 25) emanate from thesoutheastern Atlantic Forest or the adjacent Cer-rado biome (Supporting Information Fig. S1) andall fall between 12 October and 5 April (Appen-dix S1b). Accordingly, the peak breeding seasonwas considered to fall between November andFebruary, and the peak non-breeding seasonbetween May and August (Supporting Informa-tion Fig. S2).
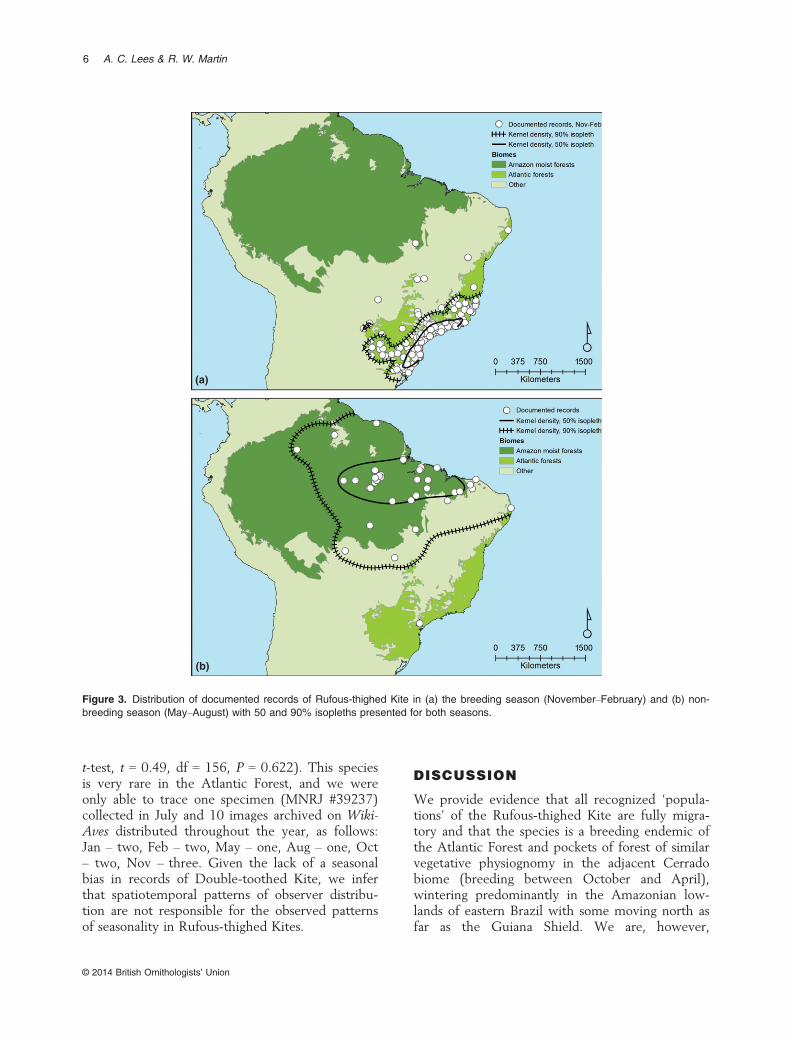
Between November and February all recordsdocumented by specimens and digital vouchers(Figs 2 and 3a) emanate from the Atlantic Forestbiome of southern Brazil, Argentina and Paraguaysouth of 10°S, with most records clusteringbetween 20°S and 30°S, with a few records in theadjacent Cerrado/Pantanal biomes (Fig. 3b). Thekernel density analysis (Fig. 3) restricts the 90%isopleth to the Atlantic Forest biome, with the50% isopleth restricted to within 200 km of theAtlantic coast. Between May and August (the aus-tral winter) all documented records (apart fromone juvenile) fall north of 15°S in Amazonia, most
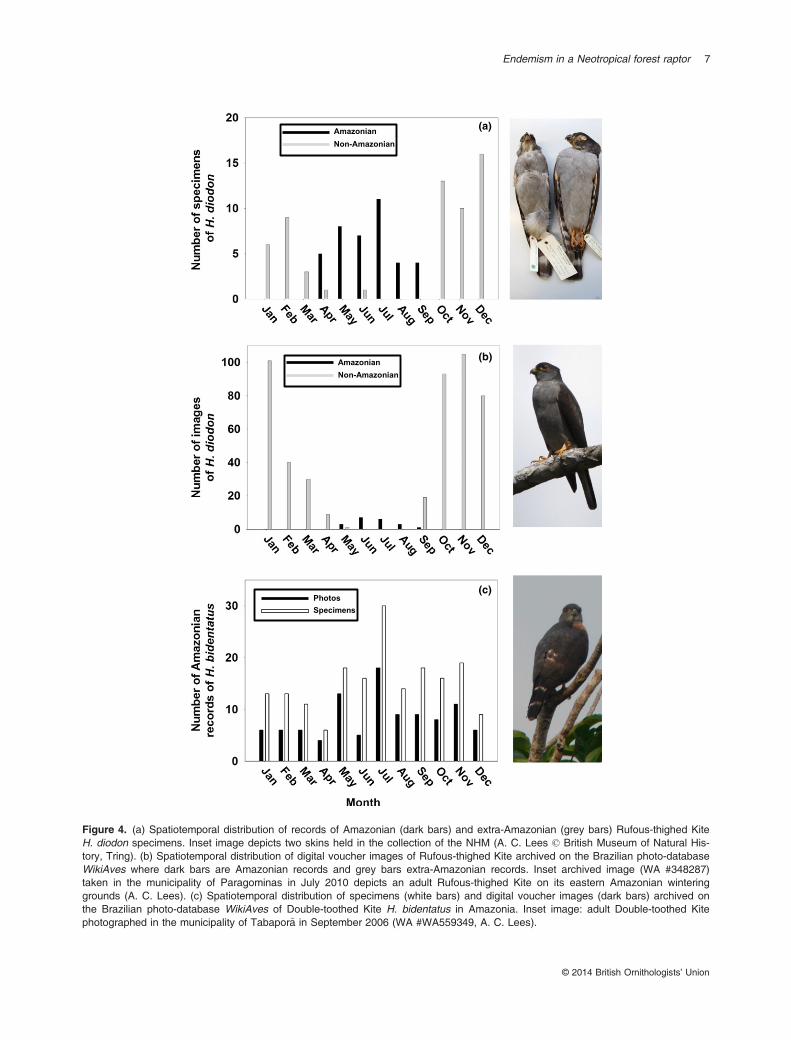
individuals wintering between 0° and 5°S, with afew records in the adjacent Cerrado and Caatingabiomes (predominantly open-habitat biomes withlittle closed-canopy forest), particularly in springand autumn. The 50% kernel density isopleth(Fig. 3b) indicates that most records are concen-trated in northeastern Brazil with the specieseither genuinely rarer or unrecorded in westernAmazonia. These differences in mean latitude ofRufous-thighed Kites in the November–February(mean = 24.28°S, sd = 2.81, n = 367) and May–August (mean = 4.21°S, sd = 4.33, n = 51) periodswere highly significant (unpaired t-test, t = 44.26,df = 416, P < 0.0001). Values for mean longitudein the November–February (mean = 46.69°W,sd = 3.28, n = 367) and May–August (mean =51.40°W, sd = 6.21, n = 51) periods were alsohighly significant (unpaired t-test, t = 7.88,df = 416, P < 0.0001) with a mean distance of2200 km between points at these two seasons. Theonly documented records from the Atlantic Forestin the austral winter are from the extreme north-eastern tip of the biome in the Pernambuco Centreof Endemism at 7.2°S, well within Amazonianwintering latitudes, and one juvenile on 10 May at24°S. This pattern of seasonality is maintained forboth specimen (Fig. 4a) and photographic records(Fig. 4b), although there are far more photographic
Figure 1. Temporal distribution of records of Rufous-thighed Kite: pale grey bars signify specimen records and dark grey bars signifyrecords documented by digital vouchers (photograph or sound recording).
© 2014 British Ornithologists’ Union
4 A. C. Lees & R. W. Martin
records from the Atlantic Forest than fromAmazonia, whereas specimen records are moreevenly distributed between these two biomes. Thebias in photographic records from Atlantic Forestreflects a higher density of observers in this biomethan in Amazonia; for instance, as of 1 July 2014
there were 5427 WikiAves users based in theAtlantic Forest state of S~ao Paulo, versus 197 inthe Amazonian state of Par�a.
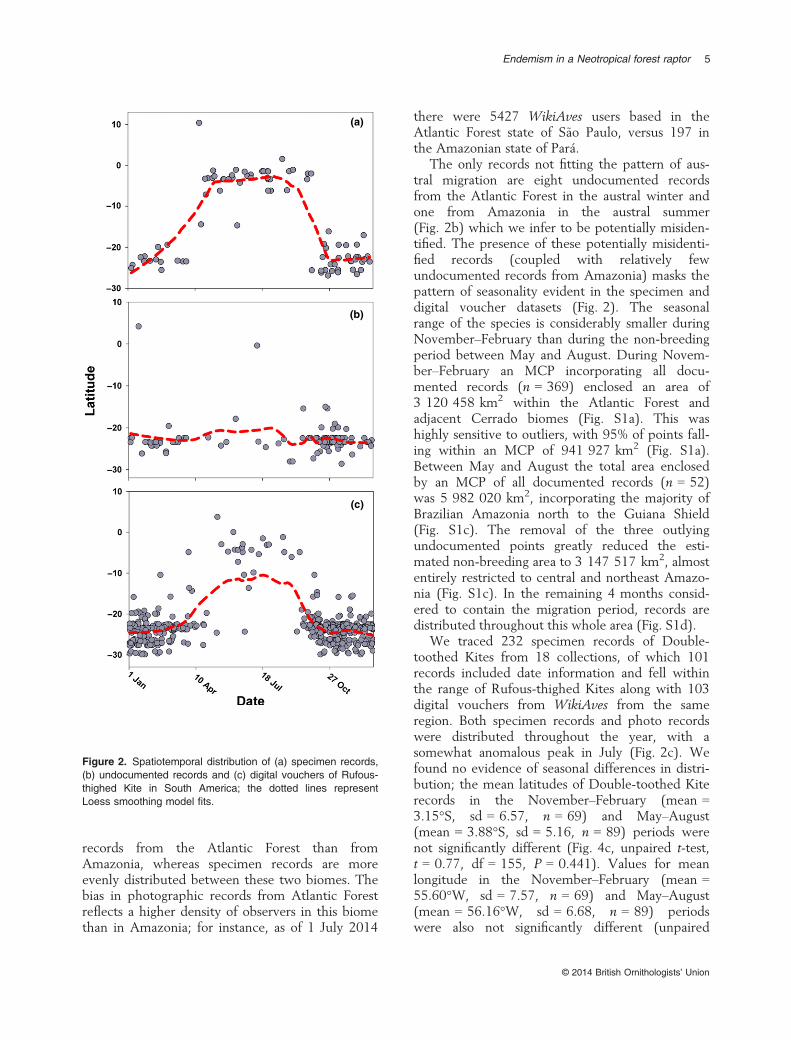
The only records not fitting the pattern of aus-tral migration are eight undocumented recordsfrom the Atlantic Forest in the austral winter andone from Amazonia in the austral summer(Fig. 2b) which we infer to be potentially misiden-tified. The presence of these potentially misidenti-fied records (coupled with relatively fewundocumented records from Amazonia) masks thepattern of seasonality evident in the specimen anddigital voucher datasets (Fig. 2). The seasonalrange of the species is considerably smaller duringNovember–February than during the non-breedingperiod between May and August. During Novem-ber–February an MCP incorporating all docu-mented records (n = 369) enclosed an area of3 120 458 km2 within the Atlantic Forest andadjacent Cerrado biomes (Fig. S1a). This washighly sensitive to outliers, with 95% of points fall-ing within an MCP of 941 927 km2 (Fig. S1a).Between May and August the total area enclosedby an MCP of all documented records (n = 52)was 5 982 020 km2, incorporating the majority ofBrazilian Amazonia north to the Guiana Shield(Fig. S1c). The removal of the three outlyingundocumented points greatly reduced the esti-mated non-breeding area to 3 147 517 km2, almostentirely restricted to central and northeast Amazo-nia (Fig. S1c). In the remaining 4 months consid-ered to contain the migration period, records aredistributed throughout this whole area (Fig. S1d).
We traced 232 specimen records of Double-toothed Kites from 18 collections, of which 101records included date information and fell withinthe range of Rufous-thighed Kites along with 103digital vouchers from WikiAves from the sameregion. Both specimen records and photo recordswere distributed throughout the year, with asomewhat anomalous peak in July (Fig. 2c). Wefound no evidence of seasonal differences in distri-bution; the mean latitudes of Double-toothed Kiterecords in the November–February (mean =3.15°S, sd = 6.57, n = 69) and May–August(mean = 3.88°S, sd = 5.16, n = 89) periods werenot significantly different (Fig. 4c, unpaired t-test,t = 0.77, df = 155, P = 0.441). Values for meanlongitude in the November–February (mean =55.60°W, sd = 7.57, n = 69) and May–August(mean = 56.16°W, sd = 6.68, n = 89) periodswere also not significantly different (unpaired
(a)
(b)
(c)
Figure 2. Spatiotemporal distribution of (a) specimen records,(b) undocumented records and (c) digital vouchers of Rufous-thighed Kite in South America; the dotted lines representLoess smoothing model fits.
© 2014 British Ornithologists’ Union
Endemism in a Neotropical forest raptor 5
t-test, t = 0.49, df = 156, P = 0.622). This speciesis very rare in the Atlantic Forest, and we wereonly able to trace one specimen (MNRJ #39237)collected in July and 10 images archived on Wiki-Aves distributed throughout the year, as follows:Jan – two, Feb – two, May – one, Aug – one, Oct– two, Nov – three. Given the lack of a seasonalbias in records of Double-toothed Kite, we inferthat spatiotemporal patterns of observer distribu-tion are not responsible for the observed patternsof seasonality in Rufous-thighed Kites.
DISCUSSION
We provide evidence that all recognized ‘popula-tions’ of the Rufous-thighed Kite are fully migra-tory and that the species is a breeding endemic ofthe Atlantic Forest and pockets of forest of similarvegetative physiognomy in the adjacent Cerradobiome (breeding between October and April),wintering predominantly in the Amazonian low-lands of eastern Brazil with some moving north asfar as the Guiana Shield. We are, however,
(a)
(b)
Figure 3. Distribution of documented records of Rufous-thighed Kite in (a) the breeding season (November–February) and (b) non-breeding season (May–August) with 50 and 90% isopleths presented for both seasons.
© 2014 British Ornithologists’ Union
6 A. C. Lees & R. W. Martin
PhotosSpecimens
AmazonianNon-Amazonian
AmazonianNon-Amazonian
(a)
(b)
(c)
Figure 4. (a) Spatiotemporal distribution of records of Amazonian (dark bars) and extra-Amazonian (grey bars) Rufous-thighed KiteH. diodon specimens. Inset image depicts two skins held in the collection of the NHM (A. C. Lees © British Museum of Natural His-tory, Tring). (b) Spatiotemporal distribution of digital voucher images of Rufous-thighed Kite archived on the Brazilian photo-databaseWikiAves where dark bars are Amazonian records and grey bars extra-Amazonian records. Inset archived image (WA #348287)taken in the municipality of Paragominas in July 2010 depicts an adult Rufous-thighed Kite on its eastern Amazonian winteringgrounds (A. C. Lees). (c) Spatiotemporal distribution of specimens (white bars) and digital voucher images (dark bars) archived onthe Brazilian photo-database WikiAves of Double-toothed Kite H. bidentatus in Amazonia. Inset image: adult Double-toothed Kitephotographed in the municipality of Tabapor~a in September 2006 (WA #WA559349, A. C. Lees).
© 2014 British Ornithologists’ Union
Endemism in a Neotropical forest raptor 7

unable to confirm the status of this species in thenortheastern Brazilian Atlantic Forest, where thereare single documented records in both the australsummer and the austral winter (Pereira et al.2008, Sonntag 2011) that may pertain to vagrantsor even a small resident population. Rufous-thighed Kite is not unique in this migration strat-egy; Chesser (1994) listed 24 species of australmigrants that winter predominantly in Amazonia,although most of these breed in the SouthernCone and move north, rather than breed in theAtlantic Forest and move northwest. The speciesthat shares the most similar pattern of movementis a passerine, the Atlantic Forest breeding ende-mic Rufous-tailed Attila Attila phoenicurus, whichhas been recorded in the austral winter as farnorth as Venezuela (Aleixo et al. 2011).
Rufous-thighed Kites are predominantly insec-tivorous (Cabanne & Roesler 2007). The speciesbreeds in the austral summer, which coincideswith the wet season in the southeast Atlantic For-est, when invertebrate biomass is likely to be high-est (e.g. Wolda 1978), so migration may be drivenby food shortages in the dry season. Migrantsreaching northern Amazonia in April–May (theaustral winter) will arrive at the start of the wetseason (Hammond 2005). We do not have enoughdata to discern whether most of the Kites winterin Southern or Northern Amazonia, as there arestrong spatial data-collection biases, with mostfieldwork taking place historically along the Ama-zon River (around 1–3°S). Tracking Kites with theaid of satellite tags would be an extremely interest-ing exercise to uncover patterns of movementsbetween and within their breeding and winteringranges.
Our conclusions are reinforced by a comparisonwith our control species, the Double-toothed Kite,which is partially sympatric in tropical forests withRufous-thighed Kite and which we consider to beequally detectable in these habitats. We do notbelieve that the spatiotemporal differences in theoccurrence of Rufous-thighed Kites reflect differ-ences in observer effort, as the Double-toothedKite was recorded throughout the year in Amazo-nia, albeit with a peak in July, which is the dryseason when visitation rates for both historicalcollectors and contemporary eco-tourists arehigher. Although data are scant for the AtlanticForest, vouchered records of this species are alsodistributed throughout the year and there are noextralimital records indicative of migration.
Why did this Wallacean shortfall persistfor 190 years?
We identify five potential contributing factors thatmay have obscured the migratory status and nar-row-breeding distribution of the Rufous-thighedKite:1 The rise of web-based biodiversity initiatives, such
as ORNIS uniting museum data and WikiAveshosting digital vouchers, is a 21st centuryphenomenon (Fig. 1a). Previously ornithologistshave had to try to unite patchy information fromdisparate sources, involving time-consuming visitsto institutions in different countries, and identi-fying errors was thus much more challenging.The digital voucher revolution ushered in anera of evidence-based qualitative ornithologythat permits both instant peer-review of birdrecords (Lees et al. 2014) and an exponentialexpansion in the total number of bird recordsas amateur ornithological observations areamassed (Dickinson et al. 2010). We echo thehope of Jetz et al. (2012) that these new initia-tives will precede a repository of global,quality-assessed and temporally explicit geogra-phical distributions for thousands of species.
2 The chances of detecting a low-density, unob-trusive forest-dwelling raptor as it disappearsinto the vastness of lowland Amazonia are rela-tively slim, and thus observer coverage and alack of ‘concentration effects’ across habitatthat is potentially uniformly suitable is likely toexplain the relative paucity of records. Forinstance, there are only two records from theAlta Floresta region of northern Mato Grosso,probably the most heavily inventoried South-ern Amazonian site, despite a history of24 years of ornithological fieldwork there (Leeset al. 2013). Given the lack of baseline naturalhistory knowledge and a pervasive assumptionthat intra-tropical migration is rare (Chesser1994), many previous authors have assumedyear-round residency for this species.
3 Although deforestation in the Atlantic Foreststarted as long ago as 1500, rates of forest lossincreased most strongly after 1900 (da Fonseca1985). Considering the extent of habitat lossafter 1900, populations of Rufous-thighedKites are likely to have been at least 10 timeshigher at the end of the 19th century thantoday, when northern South America was amajor source of bird specimens exported to
© 2014 British Ornithologists’ Union
8 A. C. Lees & R. W. Martin

European collections (Mearns & Mearns 1998).This spatial collecting bias may have given afalse impression that the species was morecommon in northern South America than else-where in its range, although Penard and Penard(1908) still regarded it as being rarer thanDouble-toothed Kite in Guyana at this time.
4 The value of old specimens is often heavilycompromised by imprecise geographical loca-tions and either erroneous or inappropriatemetadata that may compromise knowledge ofspecies distributions (Remsen 1995). Forinstance, location data for many of the speci-mens held at the NHM simply stated ‘SouthAmerica’, ‘Cayenne’ or ‘Brazil’, and only threespecimens listed date information other thanthe year. Of these, one juvenile that formedpart of the McConnell Collection (NHM#1922.3.5.979, Supporting Information Fig.S3) was listed as having been collected on theMazaruni River, British Guiana (now Guyana)in November 1898, and would represent theonly documented record from northern SouthAmerica at this season. However, this specimencould not have been collected in November, asthis 1898 expedition took place betweenAugust and October (Brown 1901), a seasonduring which occurrences of Rufous-thighedKites would be more expected; the specimenlabel was therefore probably replaced or datedonce the specimen had returned to the UK.
5 Historically, another factor obscuring themigratory status of Rufous-thighed Kites waslikely to be confusion with other resident andsympatric raptors such as the Bicoloured HawkAccipiter bicolor pileatus, Sharp-shinned HawkAccipiter striatus or juvenile and sub-adult Dou-ble-toothed Kites (Supporting Information Fig.S4). The close resemblance between adultBicoloured Hawks and Rufous-thighed Kites,adults of which are similarly grey with rufousthighs and underwings (Supporting InformationFig. S5), was first noted 140 years ago by Wal-lace (1870) and is likely to be a case of Bates-ian mimicry (Willis 1976). Thus, out-of-seasonreports of Rufous-thighed Kites without sup-porting documentation may refer to misidenti-fied Bicoloured Hawks or other small raptors.It is possible that some such misidentifiedrecords fed into quantitative assessments of thespecies status in French Guiana (e.g. Thiollay1989). The species has subsequently been
removed from the national list, as there are nounambiguously documented records with anaccompanying photo, sound recording or speci-men (Comit�e d’Homologation de Guyane, perO. Claessens, J.M. Thiollay in litt.). We antici-pate that this species will eventually be con-firmed as a rare austral migrant to this country;there are two records from adjacent Suriname,one undated 1843 specimen and one (AMNH#470008) collected near Paramaribo on 11 July1902 (Haverschmidt & Mees 1994) and origi-nally wrongly assigned to Double-toothed Kite.
Conservation status
Our reappraisal of the natural history of the Ruf-ous-thighed Kite should precipitate a reconsidera-tion of the species’ conservation status. PreviousRed List assessments have considered the species tobe resident across nearly half the South AmericanContinent, a distribution that encompasses hugetracts of relatively undisturbed forest habitatincluding countries such as Suriname, which is stillover 90% forested (CBD 2014). This is in starkcontrast to the situation in the Atlantic Forest,where Ribeiro et al. (2009) found that only 11.7%(16 377 472 ha) of the original forest vegetationremained, with 80% of fragments < 50 ha andalmost half < 100 m from its edges. Of the 55 spe-cies endemic to the Atlantic Forest, 28 (51%) areglobally threatened (Birdlife International 2014)but there are no quantitative data on the habitatrequirements or responses to land-use change ofRufous-thighed Kites. Cabanne and Roseler (2007)noted that it occupied ‘fragmented landscapes butneeds patches of native forest’ and Eduardo et al.(2007) only found the species ‘in semi-naturalareas’. We recommend that detailed modellingstudies be undertaken (e.g. Schnell et al. 2013) tore-evaluate the species’ current status and likelyfuture trends in the Atlantic Forest biome to whichwe consider it a breeding endemic, and in its east-ern Amazonian wintering areas, which are subjectto the highest absolute rates of forest loss and deg-radation worldwide and now increasingly vulnera-ble to climate change (e.g. Coe et al. 2013).
CONCLUSION
We strongly encourage ornithologists embarkingupon or carrying out meta-analyses of regionalspecies inventories in the tropics to take great care
© 2014 British Ornithologists’ Union
Endemism in a Neotropical forest raptor 9

to ensure that records are documented by speci-mens or digital vouchers with high-quality meta-data to guard against misidentifications and thesubsequent error cascades they may generate,which may impede conservation assessments(McKelvey et al. 2008, Lees et al. 2014). It is possi-ble that the status of other ‘partial migrant’ or even‘resident’ Neotropical bird species may need to berevised. Archived digital vouchers collated by citi-zen scientists, here championed by the WikiAvesinitiative (which as of July 2014 had over 18 000users and over 1 million digital vouchers archived),have here proven to be an extremely useful tool toelucidate the status of a poorly known tropical birdand represent an excellent focal point to organicallyfoster a generation of home-grown amateurNeotropical ornithologists.
We thank Reinaldo Guedes for providing us with datafrom WikiAves, Lu�ıs F�abio Silveira (MZUSP), Guilher-me Renzo Rocha Brito (MNRJ), Alexandre Aleixo &Fatima Lima (MPEG), and Mark Adams (NHM) forallowing access or proving data from their respective col-lections, Ciro Albano, Jos Barlow, Olivier Claessens,Jean M. Thiollay and Joe Tobias for comments andinformation on the status in the Guianan Shield, andtwo anonymous reviewers for comments on the manu-script. A.C.L. thanks CNPq for funding through a BolsaJovem Talento.
REFERENCES
Aleixo, A., Poletto, F. & Lima, M.D.F.C. 2011. Notas sobreos vertebrados do norte do Par�a, Brasil: uma parteesquecida da Regi~ao das Guianas, II. Avifauna. Bol. Mus.Para. Emilio Goeldi. Cienc. Nat. 6: 11–65.
Anderson, R.P. 2012. Harnessing the world’s biodiversitydata: promise and peril in ecological niche modeling ofspecies distributions. Ann. N. Y. Acad. Sci. 1260: 66–80.
Azevedo, M.A.G., Machado, D.A. & Albuquerque, J.L.B.2003. Aves de rapina na Ilha de Santa Catarina, SC:Composic�~ao, freq€uencia de ocorrencia, uso de habitat econservac�~ao. Ararajuba 11: 75–81.
Beyer, H.L. 2012. Geospatial Modelling Environment (Version0.7.2.0). Available at: www.spatialecology.com/gme(accessed 20 March 2014).
Bierregaard, R.O. 1995. The biology and conservation statusof Central and South American Falconiformes: a survey ofcurrent knowledge. Bird Conserv. Int. 5: 325–340.
Bildstein, K.L. 2004. Raptor migration in the Neotropics:patterns, process, and consequences. Ornitol. Neotrop. 15:83–99.
Birdlife International 2014. Birdlife Data Zone. Available at:www.birdlife.org/datazone/home (accessed 25 January 2014).
Braun, M.J., Finch, D.W., Robbins, M.B. & Schmidt, B.K.2000. A Field Checklist of the Birds of Guyana. Washington,DC: Smithsonian Institution.
Brown, N.E. 1901. I. Report on two Botanical Collectionsmade by Messrs. FV McConnell and JJ Quelch at MountRoraima in British Guiana. Trans. Linn. Soc. London 2ndSer. Bot. 6: 1–107.
Cabanne, G.S. & Roesler, I. 2007. A description of a nestand nestlings of the Rufous-thighed Kite (Harpagus diodon),with additional comments on diet and behavior. Ornitol.Neotrop. 18: 469–476.
Cabanne, G.S. & Seipke, S.H. 2005. Migration of the Rufous-thighed Kite (Harpagus diodon) in southeastern Brazil.Ornitol. Neotrop. 16: 547–549.
Chesser, R.T. 1994. Migration in South America: an overviewof the austral system. Bird Conserv. Int. 4: 91–107.
Coe, M.T., Marthews, T.R., Costa, M.H., Galbraith, D.R.,Greenglass, N.L., Imbuzeiro, H.M., Levine, N.M., Malhi,Y., Moorcroft, P.M., Muza, M.N., Powell, T.L., Saleska,S.R., Solorzano, L.A. & Wang, J. 2013. Deforestation andclimate feedbacks threaten the ecological integrity of south–southeastern Amazonia. Philos. Trans. R. Soc. Lond., B,Biol. Sci. 368: 20120155.
Cohn-Haft, M., Whittaker, A. & Stouffer, P.C. 1997. A newlook at the ‘species-poor’ central Amazon: the avifaunanorth of Manaus, Brazil. Ornithol. Monogr. 48: 205–235.
Convention on Biological Diversity (CBD) 2014. CountryProfiles. Convention on Biological Diversity Secretariat.Available at www.cbd.int/countries (accessed 19 March 2014).
Dickinson, J.L., Zuckerberg, B. & Bonter, D.N. 2010. Citizenscience as an ecological research tool: challenges andbenefits. Annu. Rev. Ecol. Evol. Syst. 41: 149–172.
Duong, T. 2014. ks: Kernel smoothing. R package version1.9.0. Available at: http://CRAN.R-project.org/package=ks(accessed 19 March 2014).
Eduardo, C., Carvalho, A. & Marini, M.A. 2007. Distributionpatterns of diurnal raptors in open and forested habitats insouth-eastern Brazil and the effects of urbanization. BirdConserv. Int. 17: 367–380.
Feeley, K.J. & Silman, M.R. 2011. The data void in modellingcurrent and future distributions of tropical species. Glob.Change Biol. 17: 626–630.
Ferguson-Lees, J. & Christie, D.A. 2001. Raptors of theWorld. New York, NY: Houghton Mifflin Company.
da Fonseca, G.A.B. 1985. The vanishing Brazilian Atlanticforest. Biol. Conserv. 34: 17–34.
Haffer, J. 1985. Avian zoogeography of the Neotropicallowlands. Ornithol. Monogr. 36: 113–146.
Hammond, D.S. 2005. Biophysical features of the GuianaShield. In Hammond, D.S. (ed.) Tropical Forests of theGuiana Shield: 15–194. Wallingford, UK: CABI Publishing.
Haverschmidt, F. & Mees, G.F. 1994. Birds of Suriname.Paramaribo: Vaco N.V.
Hayes, F.E., Scharf, P.A. & Ridgely, R.S. 1994. Austral birdmigrants in Paraguay. Condor 96: 83–97.
Jetz, W., McPherson, J.M. & Guralnick, R.P. 2012.Integrating biodiversity distribution knowledge: toward aglobal map of life. Trends Ecol. Evol. 27: 151–159.
Juhant, M.A. 2012. Raptor migration at Concepci�on, Bolivia.Wilson J. Ornithol. 124: 636–640.
Klavins, J. & Fariña, N. 2013. Milano de Corbata (Harpagusdiodon) en la provincia de Corrientes, Argentina. NuestrasAves 58: 51–52.
Lees, A.C., Zimmer, K.J., Marantz, C.M., Whittaker, A.,Davis, B.J.W. & Whitney, B.M. 2013. Alta Floresta
© 2014 British Ornithologists’ Union
10 A. C. Lees & R. W. Martin

revisited: an updated review of the avifauna of the mostintensively surveyed site in south-central Amazonia. Bull.Brit. Orn. Cl. 133: 178–239.
Lees, A.C., Naka, L.N., Aleixo, A., Cohn-Haft, M.,Piacentini, V.Q., Santos, M.P.D. & Silveira, L.F. 2014.Conducting rigorous avian inventories: Amazonian casestudies and a roadmap for improvement. Rev. Bras. Ornitol.22: 107–120.
Lomolino, M.V. & Heaney, L.R. (eds) 2004. Frontiers ofBiogeography: New Directions in the Geography of Nature.Sunderland, MA: Sinauer Associates.
McKelvey, K.S., Aubry, K.B. & Schwartz, M.K. 2008. Usinganecdotal occurrence data for rare or elusive species: theillusion of reality and a call for evidentiary standards.Bioscience 58: 549–555.
Mearns, B. & Mearns, R. 1998. The Bird Collectors. SanDiego, CA: Academic Press.
Newbold, T. 2010. Applications and limitations of museumdata for conservation and ecology, with particular attentionto species distribution models. Prog. Phys. Geogr. 34: 3–22.
Olson, D.M., Dinerstein, E., Wikramanayake, E.D., Burgess,N.D., Powell, G.V., Underwood, E.C., Itoua, I., Strand,H.E., Morrison, J.C., Loucks, C.J., Allnutt, T.F., Ricketts,T.H., Kura, Y., Lamoreux, J.F., Wettengel, W.W., Hedao,P. & Kassem, K.R. 2001. Terrestrial ecoregions of theWorld: a new map of life on Earth. Bioscience 51: 933–938.
Paynter, R.A. Jr. 1982. Ornithological Gazetteer ofVenezuela. Cambridge, MA: Museum of ComparativeZoology, Harvard University.
Paynter, R.A. Jr. 1989. Ornithological Gazetteer of Paraguay.2nd edn. Cambridge, MA: Museum of Comparative Zoology,Harvard University.
Paynter, R.A. Jr. 1992. Ornithological Gazetteer of Bolivia.2nd edn. Cambridge, MA: Museum of Comparative Zoology,Harvard University.
Paynter, R.A. Jr. 1994. Ornithological Gazetteer of Uruguay.2nd edn. Cambridge, MA: Museum of Comparative Zoology,Harvard University.
Paynter, R.A. Jr. 1995. Ornithological Gazetteer of Argentina.2nd edn. Cambridge, MA: Museum of Comparative Zoology,Harvard University.
Paynter, R.A. Jr & Traylor, M.A. Jr. 1991. OrnithologicalGazetteer of Brazil. Cambridge, MA: Museum of ComparativeZoology, Harvard University.
Penard, F.P. & Penard, A.P. 1908. De Vogels van Guyana:Suriname. Cayenne en Demerara (Vol. 1), Paramaibo: F.P.Penard.
Pereira, G.A., Whittaker, A., Whitney, B.M., Zimmer, K.J.,Dantas, S.M., Roda, S.A., Bevier, L.R., Coelho, G.,Hoyer, R.C. & Albano, C. 2008. Novos registros de avespara Pernambuco, Brasil, com notas sobre algumasesp�ecies pouco conhecidas no Estado. Rev. Bras. Ornitol.16: 47–53.
R Core Team. 2014. R: A Language and Environment forStatistical computing. Vienna: R Foundation for StatisticalComputing. Available at: www.R-project.org (accessed 1January 2014).
Remsen, J.V., Jr 1995. The importance of continuedcollecting of bird specimens to ornithology and birdconservation. Bird Conserv. Int. 5: 146–180.
Ribeiro, M.C., Metzger, J.P., Martensen, A.C., Ponzoni, F.J.& Hirota, M.M. 2009. The Brazilian Atlantic Forest: howmuch is left, and how is the remaining forest distributed?Implications for conservation. Biol. Conserv. 142: 1141–1153.
Ridgely, R.S. 1980. Notes on some rare or previouslyunrecorded birds in Ecuador. Amer. Birds 34: 242–248.
Ridgely, R.S. & Greenfield, J.A. 2001. The Birds of Ecuador.Ithaca, NY: Cornell University Press.
Rodrigues, A.S.L., Pilgrim, J.D., Lamoreux, J.F., Hoffmann,M. & Brooks, T.M. 2006. The value of the IUCN Red Listfor conservation. Trends Ecol. Evol. 21: 71–76.
Schnell, J.K., Harris, G.M., Pimm, S.L. & Russell, G.J.2013. Quantitative analysis of forest fragmentation in theAtlantic forest reveals more threatened bird species than thecurrent Red List. PLoS One 8: e65357.
Schulze, M.D., C�ordova, J.L., Seavy, N.E. & Whitacre, D.F.2000. Behavior, diet, and breeding biology of Double-toothed Kites at a Guatemalan lowland site. Condor 102:113–126.
Sonntag, F.A. 2011. WA369853, Harpagus diodon(Temminck, 1823). Wiki Aves – A Enciclop�edia das Aves doBrasil. Available at www.wikiaves.com/369853 (accessed 19March 2014).
Stephens, L. & Traylor, M.A. Jr. 1985. OrnithologicalGazetteer of the Guianas. Cambridge, MA: Museum ofComparative Zoology, Harvard University.
Thiollay, J.M. 1989. Area requirements for the conservation ofrain forest raptors and game birds in French Guiana.Conserv. Biol. 3: 128–137.
Thiollay, J.M. 1994. Familiy Accipitridae (Hawks and eagles).In del Hoyo, J., Elliot, A. & Sartagal, J. (eds) Handbook ofthe Birds of the World. Volume 2: New World Vultures toGuineafowl: 52–205. Barcelona: Lynx Edicions.
Wallace, A.R. 1870. Contributions to the Theory of NaturalSelection, a Series of Essays. London: Macmillan.
Wand, M.P. & Jones, M.C. 1994. Multivariate pluginbandwidth selection. Comput. Stat. 9: 97–116.
Whitney, B.M. & Cohn-Haft, C. 2013. Fifteen new species ofAmazonian birds. In del Hoyo, J., Elliott, A. & Christie, D.(eds) Handbook of the Birds of the World, Special Volume:New Species and Global Index: 225–239. Barcelona: LynxEdicions.
Willis, E.O. 1976. A possible reason for mimicry of a bird-eating hawk by an insect-eating kite. Auk 93: 841–842.
Winker, K. 1996. The crumbling infrastructure of biodiversity:the avian example. Conserv. Biol. 10: 703–707.
Wolda, H. 1978. Seasonal fluctuations in rainfall, food andabundance of tropical insects. J. Anim. Ecol. 47: 369–381.
Received 15 April 2014;revision accepted 24 August 2014.Associate Editor: Arjun Amar.
© 2014 British Ornithologists’ Union
Endemism in a Neotropical forest raptor 11

SUPPORTING INFORMATION
Additional Supporting Information may be foundin the online version of this article:
Appendix S1a. List of all records and sources ofRufous-thighed Kite records that have accompany-ing date information.
Appendix S1b. Documented confirmed breed-ing records of Rufous-thighed Kites in SouthAmerica, all of which are from Brazil.
Figure S1. Records of Rufous-thighed Kites:geographical distribution of (a) documented (filledcircle) and undocumented (crossed circles) in theaustral summer (November–February) with alldocumented (diagonal lines) and 95% documented(hatched) minimum convex polygons; (b) all con-firmed breeding records (marked by a star); (c)documented (filled circle) and undocumented(crossed circles) in the austral summer (Novem-ber–February) with all documented (diagonallines) and 95% documented (hatched) minimumconvex polygons; (d) documented (filled circle)and undocumented (crossed circles) records in themigration period (March–April and September–October).
Figure S2. Frequency distribution of breedingrecords of Rufous-thighed Kites based on indepen-dent observations of either reports of active nests,eggs or unfledged chicks.
Figure S3. Juvenile Rufous-thighed Kite (Harp-agus diodon) NHM #1922.3.5.979 collected on theMazaruni River, British Guiana (now Guyana),between August and October 1898 (not Novem-ber as indicated on the label) (A. C. Lees, © Brit-ish Museum of Natural History, Tring).
Figure S4. Top, juvenile male Rufous-thighedKite (Harpagus diodon) MPEG #15345 collectedat Santar�em, Par�a, Brazil (central-eastern Amazo-nia), on 4 July 1956; below, juvenile female Dou-ble-toothed Kite Hawk (H. bidentatus) MPEG#48030 collected at Cruzeiro do Sul, Acre, Brazil(western Amazonia), on 22 February 1992.
Figure S5. Top, adult male Rufous-thighed Kite(Harpagus diodon) MPEG #46532 collected atBel�em, Par�a, Brazil (eastern Amazonia), on 10June 1989; below, adult female Bicoloured Hawk(Accipiter bicolor pileatus) MPEG #75631 collectedat Jacarecanga, Par�a, Brazil (eastern Amazonia), on9 October 2012.
© 2014 British Ornithologists’ Union
12 A. C. Lees & R. W. Martin





























