Embed Size (px)
Citation preview

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
1
1. TÍTULO: EXPRESIÓN Y PURIFICACIÓN DE ANTÍGENOS DE Mycobacterium bovis MEDIANTE LA TECNOLOGÍA DEL DNA RECOMBINANTE CON USO POTENCIAL EN DIAGNÓSTICO. Presenta: IBQ. Paulina Karely Beltran Medina Tutor Académico: Dr. Ángel Hilario Álvarez Herrera Asesor: Dr. Abel Gutiérrez Ortega

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
2
3. RESUMEN
La tuberculosis bovina (TBb) es causada por Mycobacterium bovis. El método de
diagóstico de rutina es la prueba de la tuberculina. Sin embargo, su sensibilidad y
especificidad son muy variables, ya que los antígenos del PPD no son específicos.
Algunos autores han buscado la sustitución de estos antígenos en las pruebas
serológicas con la utilización de proteínas recombinantes de M. bovis entre ellas están
ESAT6 y CFP10.
Por otro lado se ha descrito que algunas proteínas del complejo de Mycobacterium
tuberculosis cuya expresión se ha asociado a que se codifica en forma de regulón
(regulón DosR) son altamente inmunogénicos, debido a que muestran ser antígenos
fuertemente reconocidas por las células inmunes de humanos infectados por M.
tuberculosis, y además se ha demostrado que tienen un porcentaje de similitud muy alto
con los genes ortólogos de M. bovis, mientras que están ausentes o con muy poca
homología con micobacterias ambientales como M. avium.
El objetivo de este trabajo fue producir las proteínas recombinantes PfkB, HspX y
Mb1762c de M. bovis y evaluar su antigenicidad en sangre de bovinos naturalmente
expuestos a la micobacteria, mediante la inducción de la producción de IFN-γ, y
compararla con aquella provocada por el PPD bovino (PPD-B) y/o ESAT6 y CFP10.
Como resultado se logró producir dos proteínas recombinantes PfkB y HspX, las cuales
fueron probadas cada una por separado y en conjunto en una mezcla junto con el PPD-
B, para estimular sangre de bovinos con diversas prevalencias de infección. La mezcla
de PfkB, HspX y PPD-B demostró incrementar la sensibilidad y la especifidad, que
aquella dada por las proteínas probadas individualente y/o PPD-B solo.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
3
4. ÍNDICE DE CONTENIDO Páginas
5. ÍNDICE DE CUADROS, GRAFICAS Y FIGURAS 4
6. ANTECEDENTES 9
7. FUNDAMENTACIÓN 11
7.1. Tuberculosis Bovina 11
7.2.Transmisión 12
7.3.Patogenia 12
7.4. Epidemiología 14
7.5. Reacción de la hipersensibilidad tardía de la piel, bases del
diagnóstico de la tuberculosis bovina 16
7.6. Diagnóstico de la tuberculosis bovina 17
7.6.1. Prueba de la tuberculina 18
7.6.2. Prueba del Interferón Gamma (IFN-γ) 19
7.6.2.1. Ventajas prácticas de la prueba del IFN-γ 20
8. JUSTIFICACIÓN 22
9. DEFINICIÓN DEL PROBLEMA 23
10. HIPÓTESIS 25
11. OBJETIVOS 25
11.1. Objetivo General 25
11.2. Objetivos Específicos 25
12. MATERIALES Y MÉTODOS 26
12.1. Diseño de oligonucleótidos y amplificación de los genes. 26
12.2. Clonación en el vector pET101/D-TOPO para expresión usando
cultivos de E. coli. 27
12.3. Clonación en el vector pIVEX 2.3d para expresión en un sistema de
traducción libre de células. 28
12.4. Expresión de las proteínas recombinantes. 29
12.4.1. Análisis de las proteínas recombinantes mediante electroforesis en 30

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
4
gel de poliacrilamida (SDS-PAGE).
Páginas
12.4.2. Análisis de las proteínas recombinantes por Western blot con
anticuerpo anti-His6. 30
12.5. Purificación de las proteínas recombinantes mediante columnas de
afinidad. 31
12.6. Evaluación de la antigenicidad de las proteínas recombinantes en
sangre de bovino. 32
12.7. Análisis estadístico. 33
12.7.1. Cumplimiento de supuestos estadísticos 33
12.7.2. Comparación de grupos 33
12.7.3. Curvas ROC (Reciever Operating Characteristics) 33
13. RESULTADOS 35
13.1. Selección de antígenos 35
13.2 Amplificación de los genes seleccionados. 36
13.3. Clonación de gen pfkB en vector de expresión 37
13.4. Clonación de gen hspx en vector de expresión pIVEX2.3d 39
13.5. Síntesis de las proteínas pfkB y hspx en sistema RTS ProteoMaster 40
13.6. Purificación de la proteína PfkB sintetizada con el sistema RTS
proteomaster 40
13.7. Purificación de la proteína PfkB expresada en E. coli BL21 41
13.8. Clonación y purificación de la proteína Hspx en vector de expresión
pET101-TOPO 43
13.9. Clonación y expresión de la proteína Mb1762 en vector de expresión
pET101-TOPO 46
13.10. Evaluación de la antigenicidad de las proteínas recombinantes en
sangre de bovino 47
14. DISCUSION 52
15. CONCLUSION 54
16. REFERENCIAS
17.ANEXO 1 Cartas de Aceptación Congresos Nacionales 68

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
5
18. ANEXO 2 Copia de articulo de divulgación científica 70
5. Índice de Tablas y Figuras
Tablas
Páginas
12.1 Oligonucleótidos usados para amplificar los genes completos de las
proteínas de este estudio de acuerdo a los vectores de clonación. 27
13.1. Proteínas del regulón DosR de M. bovis consideradas para el estudio 35
13.2. Similitud de los antígenos DosR seleccionados de distintas
micobacterias 36
13.3. Sensibilidad de los antígenos y valores de corte requeridos. 49
Gráficas
13.1. Curva ROC. Determinación y comparación de la sensibilidad de la
mezcla de proteínas que contiene PPD-B, HspX y PfkB contra las proteínas
HspX, PfkB, CFP10/ESAT6 y el PPD-B proveniente del kit comercial
Bovigam 48
13.2. Comparación estadística de animales infectados 49
13.3. Detección de animales infectados con tuberculosis bovina empleando 51
Figuras
Fig. 13.1. Productos de amplificación por PCR de los genes hspX,
mb1762c y pfkB. 36
Fig. 13.2. Productos de digestión con enzimas de restricción NcoI y SmaI. 37
Fig. 13.3. Productos de digestión del vector recombinante pIVEX
2.3d+pfkB con las enzimas SmaI y NcoI. 38
Fig. 13.4. Producto de PCR de pIVEX+pfkB 38
Fig. 13.5. Digestión de pIVEX 2.3d+hspX con enzima SmaI 39
Fig. 13.6. Productos de PCR de pIVEX2.3d+hspX 39

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
6
Páginas
Fig. 13.7. Western-blot con anticuerpo anti-His6 de la proteína PfkB
sintetizada en RTS Proteo Master 40
Fig. 13.8. Gel de poliacrilamida teñido con azul de Coomassie de las
fracciones eluidas por la columna de afinidad donde se pasó la mezcla de
proteínas sintetizadas en el Proteo Master. 41
Fig. 13.9. Gel teñido con plata de las fracciones purificadas de la proteína
PfkB por columna de afinidad. 42
Fig. 13.10. Western blot con anti-His6 después de la purificación por una
columna de afinidad. 42
Fig. 13.11. PCR de clonas obtenidas en vector pET101/D-TOPO+hspX y
Mb1762c. 43
Fig. 13.12. Purificación de la proteína HspX en vector de expresión
pET101-D/TOPO.
(A) Gel teñido por Comassie de la proteína HspX expresada en E. coli
BL21 inducida con IPTG 1 mM. Fracción soluble de un cultivo de la cepa
recombinante sin inducir
(B) Western blot con anti-His6 de la proteína HspX expresada en E. coli
BL21 inducida con IPTG 1 mM. 44
Fig. 13.13. Gel teñido con plata de fracciones purificadas de la proteína
HspX por columna de afinidad. 45
Fig. 13.14. Western blot de la proteína HspX
(A) Western blot con anticuerpo monoclonal anti-HspX.
(B) Western blot con anticuerpo anti-His.. 45
Fg. 13.15. PCR de clonas obtenidas pET101/D-TOPO+Mb1762c. 46

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
7
Índice de Abreviaturas
BSA seroalbúmuna bovina
CMH Complejo Mayor de Histocompatibilidad
EDTA Ácido etilendiaminotetraacético
HIS histidina
IFN-γ Interferón-gamma
IPTG isopropil-β-D-tiogalactopiranósido
KCl Cloruro de potasio
kDa kilo daltones
KH2PO4 Fosfatode potasio monobásico
M. africanum Mycobacterium africanum
M. avium Mycobacterium avium
M. avium sp.
paratuberculosis
Mycobacterium avium subesp. paratuberculosis
M. bovis Mycobacterium bovis
M. canetti Mycobacterium canetti
M. microti Mycobacterium microti
M. tuberculosis Mycobacterium tuberculosis
Pb Pares de bases
PCR Reacción en cadena de la polimersa
PPD Derivado Protéico Purificado
PPD-A Derivado Protéico Purificado de M. avium
PPDD-B Derivado Protéico Purificado de M. bovis
ROC Receiver Operating Characteristics
SDS-PAGE Electroforesis en gel de poliacrilamida con dodecilsulfato de sodio
TBb Tuberculosis Bovina
TMB 3, 3´, 5, 5´-tetrametilbenzideno

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
8
6. ANTECEDENTES.
La Tuberculosis bovina (TBb) es una enfermedad que afecta al ganado bovino, la cual
es considerada una zoonosis ya que puede afectar a otras especies de mamíferos
inclusive al humano. El agente causal es Mycobacterium bovis miembro del complejo
M. tuberculosis causante de la tuberculosis en humanos. En países subdesarrollados
como en el caso de México, la TBb es un problema zoosanitario muy frecuente y es
muy importante realizar campañas de erradicación, puesto que al estar los animales
enfermos, predispone alganado a otras enfermedades, limitando el desarrollo de la
ganadería, lo cual causa fuertes perdidas económicas.
Dentro de los métodos de detección existentesla prueba intradérmicatambién conocida
como prueba de la “Tuberculina”, estipulada por la Norma Oficial Mexicana contra la
tuberculosis bovina, (NOM-031-ZOO-1995), la prueba se basa en la respuesta
inmunológica del animal a la inyección intradérmica de 0.1 ml de tuberculina en la
dermis del pliegue caudal derecho con un extracto proteínico purificado (PPD) de M.
bovis AN5 ó Valleé (Monaghan y col 1994), cualquier induración igual o mayor a 5 mm
se considera como una reacción positiva (animal PPD o tuberculina positiva). Se mide
la reacción de hipersensibilidad del tipo retardado al componente. Pero las pruebas no
son muy precisas ya que se dan muchos falsos positivos, debido a la falta de
especificidad, enmascarando si el animal esta realmente infectado por M. bovis o quizá
por otras micobacterias. Además se ha demostrado una disminución de un 5% por año
en su sensibilidad debido a la anergia inmunológica generada por múltiples
aplicaciones. En diferente estudios en el campo se ha determinado una sensibilidad de la
prueba entre el 63-99%, con una media de 83.9% (De la Rua Domenech y col 2006).
Aunado a esta prueba se realiza otra prueba confirmatoria que consiste en el cultivo
microbiológico, donde se puede aislar a M. bovis de biopsias provenientes de animales
sacrificados. Sin embargo, este examen tiene el inconveniente de ser muy tardía, ya que
el microorganismo demora entre 4 y 6 semanas para crecer en medios selectivos y sólo
puede realizarse después del sacrificio.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
9
Para tratar de disminuir los errores de interpretación de la prueba intradérmica, se
desarrollo una prueba comercia para medir directamente el interferón gamma (IFN-γ) en
la sangre del bovino (Bovine Gamma Interferon Test-Bovigam®). La prueba consiste
en incubar sangre completa de bovinos sospechosos de tuberculosis con PPD bovino
(PPD-B) bajo condiciones especiales. Si el animal ha estado en contacto con el
microorganismo, sus linfocitos liberarán IFN-γ, el cual será detectado a través de un
sistema de ELISA sándwich, donde los anticuerpos anti- IFN-γ unidos a una placa de 96
pozos capturan el IFN-γ bovino. La reacción se detecta por la adición de un segundo
anticuerpo específico anti- IFN-γ conjugado a una peroxidasa la cual reacciona con un
sustrato produciendo una coloración (Wood y col 2001).
Una desventajas importante de esta prueba, es que también está basada en la
sensibilización de los linfocitos usando el PPD-B para la identificación de los animales
infectados, por lo que algunos reportes indican la identificación de falsos positivos
producto de coinfecciones con micobacterias ambientales que cruzan antigénicamente
con el PPD (Aranaz y col 2006)., con una sensibilidad que varia entre el 69 y el 99%
con una media de 88%, por tales motivos se ha tratado de utilizar la prueba usando
antígenos mas específicos como ESAT-6 y CFP10 (Aagaard y col 2006).
Debido a que las pruebas de diagnóstico existentes hasta el momento, como son la
prueba de la tuberculina y la del IFN-γ, están basadas en la estimulación de la respuesta
inmune usando derivados proteícos complejos (PPD-B y PPD-A) para hacer un
diagnóstico diferencial, la especificidad se ve comprometida debido a que los PPD-B y
PPD-A comparten diversos antígenos comunes entre ambos microorganismos (Borsuk y
col 2009). Por ello, se ha sugerido usar proteínas que sean más especificas, así como
también la inclusión de nuevos criterios de interpretaciones (Cockle y col, 2006).
En los últimos años se ha debido un significativo progreso en la identificación de
algunas proteínas antigénicas de M. bovis, en un intento por identificar aquellas que
sean más especificas que las presentes en el PPD (Aagaard y col, 2003). La utilización
de algunas proteínas recombinantes en la prueba del IFN-γ como ESAT-6, CFP-10 y

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
10
MPB70, h permitido sugerir sustituiral PPD-B en las pruebas intrdérmica y del IFN-γ
(Whelan y col 2010, Casal y col, 2010.)
7. FUNDAMENTACIÓN
7.1. Tuberculosis Bovina
La tuberculosis bovina es una enfermedad crónica de los animales provocada por M.
bovis, bacteria que se considera GRAM-positiva y que guarda una estrecha relación con
la bacteria causante de la tuberculosis humana, siendo miembro del complejo M.
tuberculosis caracterzados por provocar infecciones con lesiones granlomatosas.
M. bovis pertenece a la clase Actinomycetes, Orden Actinomycetales, Familia
Mycobacteraceae, género Mycobacterium(Biet y col, 2005). Estas bacterias se
caracterizan por ser bacilos ácido-alcohol resistentes, es decir, resisten a la decoloración
con alcohol acidificado una vez que se han teñido con carbol-fucsina (tinción de Ziehl-
Neelsen).
El género está dividido en dos grandes grupos: 1) el complejo M. tuberculosis (M.
tuberculosis, M. bovis, M. canetti, M. microti,M. africanum, entre otras) y 2)
micobacterias no-tuberculosas (Biet y col,2005).
Estas bacterias principalmente se transmiten por vía aerógena, y en estados avanzados
de la infección pueden causar una enfermedad diseminada en la mayoría de órganos, sin
embargo la tuberculosis pulmonar es la más común (Ward 2005, Lopez de Buen y col,
2007, Flynn y col, 2001).
Las principales especies que afectan al ganado bovino son: M. bovis, M. tuberculosis,
M. avium y M. avium ssp paratuberculosis. Aunque se considera que el verdadero
hospedador de M. bovis es el ganado bovino, la tuberculosis bovina también se
considera una zoonosis , ya que también se ha encontrado la bacteria en los humanos
(López de Buen y col, 2007, de Ward 2005), y en otras especies de mamíferos, por
ejemplo en búfalos, venados, bisontes, ovejas, cabras, caballos, camellos, cerdos y
tejones. Por lo cual es una enfermedad que puede permanecer en reservorios naturales y

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
11
volver a infectar al ganado sano por lo que se considera muy difícil de erradicar (Aranaz
y col. 2006, Vordemeier y col. 2006).
7.2.Transmisión
En el bovino la vía más importante de infección por M. bovis es la respiratoria y esto
ocurriría por inhalación de partículas de polvo contaminados con el bacilo. En animales
adultos infectados naturalmente, se puede apreciar una marcada predominancia de
lesiones tisulares en ganglios retrofaríngeos, traqueobronquiales y/o mediastínicos
(Perdomo y col, 1986).
La infección por vía digestiva se da en casos en que los animales ingieren leche, agua o
forraje contaminado y se evidencia por la reacción inmunológica del complejo inmune
celular o primario en órganos digestivos y ganglios regionales (Perdomo y col, 1986).
A pesar de que la vía aerógena es la principal fuente de infección en el bovino, 80 a
90% de los casos, se estima que los bovinos expuestos se contagian entre un 10 a un
20% por vía digestiva. Esta forma de infección es presentada en terneros que se
alimentan de vacas tuberculosas o por leche contaminada no pasteurizada. De 1 a 3% de
vacas presentan mastitis tuberculosa, convirtiéndose en diseminadoras permanentes de
bacilos. También se contagian consumiendo alimentos y agua contaminados por los
bacilos que están en contacto con el suelo que contiene heces de animales infectados.
Otras vías no comunes pero probables son: la cutánea, la congénita y la genital.
(Abalosy col, 2004, Biet y col, 2005) .
Los factores de predisposición para el contagio de la tuberculosis bovina son: el
hacinamiento, la poca ventilación, descarga de secreciones respiratorias, acumulación
de heces fecales, descargas uterinas, monta natural y alimentación de becerras con leche
sin pasteurizar (de Ward 2005, Perdomo y col, 1986, Soto 2007).
7.3.Patogenia

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
12
En cuanto a la evolución de la tuberculosis se describen tres etapas clásicas, las cuales
no se dan todas obligatoriamente:
a) Período de primo-infección. La lesión inicia en el órgano que actúa como puerta de
entrada, en este lugar se establece el primer contacto de la bacteria en el organismo
(foco primario), en este punto donde el bacilo se aloja se desencadenan una serie de
reacciones histológicas y orgánicas. Posteriormente los bacilos se drenan por vía
linfática a los nódulos linfáticos de la zona, vehiculizados por los macrófagos. En
estados más avanzados, el bacilo puede penetra en los pulmones, se multiplica y se
disemina en el órgano, produciendo lesiones en forma de tubérculo al igual que
cuando infectan a los nódulos linfáticos bronquiales (Soto 2007).
b) Procesos post-primario. Las manifestaciones observadas en este período son:
- Reactivación de una lesión antigua, ya que el animal posee inmunidad adquirida
durante la primera infección ó la sobreinfección, es decir una reinfección sobre
agregada por los bacilos venido del exterior (reinfección exógena).
- La exacerbación de focos preexistentes de la primo infección con siembra por
vía hemolinfática de bacilos provenientes de esos focos.
- La formación de granulomas en los órganos donde se detienen, la extensión o
diseminación de las lesiones se puede realizar por vía linfática, sanguínea o por
contacto seroso.
La primo infección, rara vez se elimina en los animales, casi siempre progresa lenta o
rápidamente según la condición inmunológica del hospedero. La diseminación a través
del torrente sanguíneo y de las vías linfáticas puede ser generalizada y causar muerte
rápida en el animal. La mayoría de las veces la tuberculosis bovina tiene un curso
crónico y limitado a un sólo órgano: el pulmón (Manual de Merck de Veterinaria 2000).
Experimentalmente el foco primario visible se desarrolla en ocho días y la calcificación
en un plazo de tres semanas (Liébana y col, 1998).
c) Agravamiento
La resistencia adquirida o pre munición es vencida a consecuencia de ciertas
influencias (mala higiene, subalimentación, agotamiento, enfermedades

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
13
intercurrentes, etc.) llevando a la bacilemia, luego a la generalización y por último a
la muerte(Ward 2005, Soto 2007, Manual de Tuberculosis bovina de Uruguay
1986).
7.4. Epidemiología
Aproximadamente el 30% de los animales expuestos llegan a ser infectados, los bacilos
micobacterianos al entrar al organismo del animal, pueden ser fagocitados por
macrófagos; éstos pueden eliminar a las micobacterias o permitir que proliferen dentro
de ellos, en este ultimo caso, se puede formar un foco primario, ya que la bacteria
prolifera intracelularmente e induce a que las citocinas inicien la respuesta inflamatoria;
y esta respuesta caracteriza por una reacción de hipersensibilidad tipo IV, esta lesión es
formada por macrófagos muertos, rodeados por granulocitos, linfocitos y otras células
del sistema inmune (Aranaz y col 1996, Tizard 1995).
El organismo ha desarrollado mecanismos para evadir la eliminación de los bacilos
gracias a una fuerte respuesta inmune mediada por células. La resistencia natural del
animal destruye algunos bacilos, liberando ciertas moléculas que actúan como
antígenos. Cuando se activan los linfocitos estimulados inmunológicamente, los
macrófagos muestran actividad antimicobacteriana (Pollock y col 2005). Sin embargo
algunos bacilos no pueden ser destruidos, ya que durante la infección, el macrófago
juega un papel contradictorio ya que es la unidad primaria de defensa y además, el sitio
primario de la replicación de la bacteria (Flynn y col 2001).
Puesto que M. bovis persiste en los macrófagos dentro de un granuloma en los órganos
de los huéspedes infectados, estos bacilos son los que llegan a los nódulos vía vasos
linfáticos produciendo un proceso inflamatorio inespecífico hasta que el nivel de
hipersensibilidad sea lo suficientemente acentuado como para producir una necrosis
caseosa.
Los granulomas recién formados están compuestos por fagocitos mononucleares
inmaduros rodeados de células efectoras linfocíticas como las células T CD4+ y CD8+.
Durante el proceso de maduración a granulomas productivos, los fagocitos
mononucleares se diferencian en macrófagos y llegan a ser activados, agregándose en

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
14
células gigantes multinucleares y células “epiteloidales” (Flynn 2001, Waters 2011,
Welsh 2005, Pollock 2005).
Las células inmunológicamente competentes son los linfocitos T. El periodo de latencia
para alcanzar el pico de actividad requiere varios días a dos semanas frente a un
segunda exposición, se produce una respuesta lenta cuyo pico se alcanza entre 24 y 72
hrs. Los antígenos son procesados primero por los macrófagos análogos a los del
sistema humoral inmunitario y luego presentado a las células T. Estas células T
sensibilizadas atraen macrófagos adicionales que se acumulan en el lugar de reacción.
Dichos macrófagos son los componentes celulares principales de la respuesta
inflamatoria mediada por la inmunidad celular. Una vez, reconocido el antígeno, los
linfocitos liberan sustancias denominados linfocinas que atraen monocitos (macrófagos)
del torrente sanguíneo. Los linfocitos sensibilizados pueden participar directamente en
la destrucción del antígeno, (células T citotóxicas o killer cells) pero los macrófagos
atraídos también elaboran materiales citotóxicos y presentan su capacidad fagocítica
aumentada, requiriendo activación por la linfocinas. Esta reacción es una especifica
pero una vez activados los macrófagos se convierten en citotóxicos inespecíficos (, Roitt
y col 1998, Abbas y col 2004).
El granuloma tuberculoso contiene ambas células T CD4+ y CD8+ que probablemente
participan en la batalla continua para contener la infección dentro de los granulomas y
la prevención de la reactivación (Pollock 2005, Flynn 2001, Skinner 2003, Mcaulay
2003)
Se encontró que el interferón gama(IFN-γ) es un citocina clave en el control de la
infección por M. bovis. Esta citosina es producida tanto por linfocitos T CD4+ como por
CD8+ en la tuberculosis, así como por las células NK(Flynn y col 2001, Waters 2010,
Welsh 2005). El factor de necrosis tumoral alfa (TNF-α), aunque es ineficaz cuando se
usa solo, tiene un efecto sinérgico con el IFN-γ para inducir los efectos
antimicobacterianos de macrófagos. Un importante mecanismo efector responsable de
la actividad antimicobacteriana del IFN-γ y TNF-α, es la inducción de la producción de
la producción de óxido nítrico y intermediarios de nitrógeno reactivo (RNI) por lo
macrófagos a través,de la acción de la forma inducible de la sintetasa del óxido nítrico

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
15
(NOS2), de hecho, estas dos citocinas actúan de forma sinérgica para inducir la
expresión de la vía de generación de RNI (Flynn y col 2001, Pollock 2005).
La formación del fagolisosoma es otra función de los macrófagos, que es dirigida a
controlar la infección intracelular; principalmente el lisosoma es un complejo de
orgánulos vacuolares de la vía endocítica tardía. Dentro de las vacuolas lisosomales
existen potentes enzimas hidroliticas, capaces de degradar una amplia gamma de
macromoléculas incluyen microbios. La función óptima de estas enzimas es que
trabajan a un PH acido (una condición encontrada en el medio intralisosomal). El
lisosoma trabaja en un ambiente muy acido en las células animales: pH 4.5-5.0; este
ambiente acido se mantiene gracias a las bombas de membranas de protones
dependiente de ATP, la H+ATPasa (Flynn 2001, Welsh 2005).
7.5. Reacción de la hipersensibilidad tardía de la piel, bases del diagnóstico de la
tuberculosis bovina
El conocimiento de este mecanismo comenzó con la descripción que hizo Koch de un
fenómeno que lleva su nombre; en cobayos infectados por vía sistémica con
Mycobacterium tuberculosis, luego de 3 semanas aplicaba una nueva dosis de bacilos
por vía intradérmica Koch observó, que en el lugar de la segunda inoculación, después
de producirse un nódulo que alcanzaba su mayor tamaño luego de las 72 hrs, se curaba
espontáneamente aunque el cobayo moría por tuberculosis debida a la primera
inoculación (Soto 2007).
Esta inmunidad de manifestación local tiene un carácter diferente de la inmunidad por
anticuerpo circulantes y por el papel fundamental desempeñado por los leucocitos
mononucleares, se le ha denominado inmunidad celular o mediada por células.
También demostró que la inoculación intradérmica de tuberculina podría producir una
reacción local, que es manifestación de la hipersensibilidad retardada que se produce
luego de 3 semanas post-infección. Luego de la infección primaria, lo bovinos
desarrollan inmunidad celular e hipersensibilidad. La inmunidad adquirida puede
retardar la evolución de la lesión, pero no la detiene (Gómez y col, 1986).

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
16
Entre las especies de micobacterias más comunes que causan sensibilidad tuberculínica
en la ausencia de infección de M. bovis se pueden encontrar:
1. M. tuberculosis. Causa predominante de tuberculosis humana. Esta bacteria no
provoca enfermedad progresiva en el bovino, pero es una importante causa de
reactividad en bovinos expuestos a personas enfermas de tuberculosis.
2. Complejo M. avium. La exposición ambiental de bovinos con M. avium es una
causa muy importante de sensibilidad heteroespecífica en el bovino.
3. M. paratuberculosis (M. johnei). Es la causa de la paratuberculosis (enfermedad
de Johne) en bovino y es una causa de sensibilidad hetero-específica en el
bovino.
4. Otras micobacterias no patógenas( M. chelonae, M. abscessus), como las
especies de micobacterias que se encuentran en las lesiones de piel (dermatitis
tuberculosa), capaces de causar sensibilidad tuberculínica.
Cabe destacar, que la micobacteria tiene más de 1000 antiígenos (Ag) diferentes,
algunos inmunogénicos, otros inmunosupresores. Unos son presentados por el
Complejo Mayor de histocompatibilidad clase II de los macrófagos a los linfocitos
CD4+ y/o linfocitos B, otros por el CMH clase I., de células somátivas a los linfocitos
CD8+. Así algunas proteínas unidas a una fracción cérea pueden mediante inyección
inducir una respuesta inmune celular (sensibilidad a la tuberculina) y provocar la
formación de diversos anticuerpos. (Soto 2007).
7.6. Diagnóstico de la tuberculosis bovina
Existen métodos directos e indirectos para diagnosticar la tuberculosis bovina. En los
directos se determina la presencia del agente en el huésped, esto es en las plantas de
sacrificio, la inspección epidemiológica es una actividad básica en la vigilancia de la
tuberculosis bovina, ya que es ahí donde se detectan en los bovinos las lesiones
macroscópicas asociadas a la infección que se ven a simple vista. La presencia de
tumoraciones en los diferentes órganos. El foco de necrosis presenta una coloración
amarillenta con apariencia caseificada (como queso) y es posible detectar la presencia
de calcio; durante la necropsia se perciben como pequeñas granulaciones blanquecinas

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
17
que crepitan al cortar con el cuchillo. También puede observarse exudado de apariencia
purulenta en meninges.
La prueba confirmatoria de la tuberculosis bovina es el cultivo microbiológico donde se
puede aislar a M. bovis en biopsias de animales sacrificados. Sin embargo, esta prueba
tiene el inconveniente de ser muy tardía, ya que el microorganismo demora entre 4 y 6
semanas para crecer en medios selectivos y sólo puede ser realizada después del
sacrificio.
Los métodos indirectos son aquellos que determina la respuesta inmune del huésped al
agente, ya sea esta de tipo celular o humoral. La prueba más utilizada, como ya se
mencionó anteriormente y la cual es mundialmente practicada es la prueba de la
tuberculina o de intradermo-reacción con PPD bovina (es un derivado proteico
purificado producido a partir de cultivos inactivados de Mycobacterium bovis).
La Secretaria de agricultura, ganadería y desarrollo rural, mediante la Norma Oficial
Mexicana NOM-031-ZOO-1995, para la campaña nacional contra la Tuberculosis
Bovina (Mycobacterium bovis), declara que las pruebas diagnósticas autorizadas son:
a) Prueba de la tuberculina
b) Histopatología
c) Aislamiento bacteriológico e identificación
d) Tipificación
7.6.1. Prueba de la tuberculina
Es el método oficial para detectar a los animales con tuberculosis bovina, sin embargo,
esta prueba posee algunas desventajas relacionadas con la sensibilidad de la prueba. Se
basa en la inducción de una respuesta celular de hipersensibilidad tardía a la inyección
intradérmica de PPD. Cuando se inyecta PPD a un animal sensibilizado por la infección
con M. bovis o por la exposición a antígenos de reacción cruzada de micobacterias
ambientales, hay una respuesta inflamatoria e hinchazón en el sitio de inyección con
mayor intensidad 48-72 horas después de la inyección y es rápidamente reversible.
Debido a la reactividad cruzada con infecciones por micobacterias ambientales tiene
baja sensibilidad que aún provoca numerosos diagnósticos falsos positivos (de Ward

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
18
2005). En diferentes estudios en el campo se ha determinado una sensibilidad de la
prueba entre el 63-99%, con una media de 83.9% (De la Rua Domenech y col 2006).
Entre los incovenientes de la aolicación de la preuba y que son la causa de que en
animales PPD (+) no se muesten lesiones visibles, se destacan las siguientes causas:
- Muchos de estos animales pueden estar en estados iniciales de la enfermedad por
lo que son sensibles a la tuberculina pero no muestran lesiones visibles.
- Muchos de estos animales tienen lesiones localizadas en partes del cuerpo que
generalmente no se examinan durante el trabajo sistemático de inspección de
carnes.
- Los animales que han estado en contacto con el bacilo tuberculoso aviario,
ordinariamente muestran lesiones discretas o invisibles, siendo sensible a la
tuberculina.
- Puede haber anergia inmunológica al componenete PPD debido a coinfecciones
que abaten el sistema inmune, o también debido a múltiples aplicaciones de la
prueba.
7.6.2. Prueba del Interferón Gamma (IFN-γ)
Por otro lado, existe una prueba comercial, la cual consiste en incubar sangre completa
de bovinos sospechosos de tuberculosis diagnosticados con PPD bovino (PPD-B). Si el
animal ha estado en contacto con el microorganismo, sus linfocitos liberarán IFN-γ, el
cual será detectado a través de un sistema de ELISA sándwich. La desventaja es que la
prueba es comparativamente cara, y además, debido a que también usa el derivado PPD-
B para la identificación de los animales infectados, algunos reportes indican la
identificación de falsos positivos producto de co-infecciones con micobacterias
ambientales que cruzan antigénicamente con el PPD (Aranaz y col. 2006), sin embargo,
sólo se utiliza como prueba de diagnóstico complementario o confirmativa en países
como Australia, Nueva Zelanda y los EEUU (De Ward 2005).
La prueba del interferón gama (IFN-γ) (Bovine Gamma Interferon Test-BOVIGAM)
(Wood y Jones 2001) es una prueba especifica que evalúa la inmunidad celular que se
desarrollo para la detección del IFN- γ principalmente por parte de las células T, y no

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
19
tanto en la respuesta innata. La sangre completa del bovino sospechoso de estar
infectado primeramente se incuba con antígenos como el PPD bovino, PPD aviar y
control negativo, durante toda la noche y el IFN-γ liberado en el plasma sobrenadante,
es detectado por un ensayo inmuno-enzimático empleando anticuerpos monoclonales.
La infección de tuberculosis es indicada por la liberación predominante de IFN-γ en
respuesta al PPD-B (de la Rua Domenech 2006, Palmer 2006, Rangen 2009).
La desventaja importante de esta prueba es que también está basada en la
sensibilización de los linfocitos usando una mezcla compleja de antígenos, el PPD-B y
PPD-A, para la identificación de los animales infectados, por lo que algunos reportes
indican la identificación de falsos positivos producto de coinfecciones con
micobacterias ambientales que cruzan antigénicamente con el PPD (Aranaz y col 2006),
con una sensibilidad que varía entre el 69 y el 99%, con una media de 88%; por tales
motivos se ha tratado de utilizar la prueba usando antígenos más específicos como
ESAT-6 y CFP10 (Aagaard y col 2006).
7.6.2.1. Ventajas prácticas de la prueba del IFN-γ
La sensibilidad aunque es muy similar a la prueba de la tuberculina doble comparativa,
el tiempo necesario para desarrollar una respuesta es ligeramente menor. También
permite mayor repetición de la prueba, debido a que se puede utilizar la misma sangre
que se evaluó, no se inyecta la tuberculina, por lo tanto, no hay interferencia con el
sistema inmune del huésped; no es necesaria una segunda visita a la granja para leer la
prueba, lo cual permite una interpretación más objetiva y estandarizada de los resultados
puesto que todas las mediciones son basadas en el laboratorio; reduce muchos
problemas prácticos asociados a las prueba intradérmica (ejemplo: malas
interpretaciones, error de operador); se pueden incluir antígenos específicos definidos.
permitiendo la diferenciación de la respuesta de IFN-γ debido a la infección con
micobacterias patógenas de las micobacterias ambientales (Dela Rua Domenech 2006,
Vordermeier y col 1999; Pollock y col 2005, 2001; Buddle y col 1999; Waters y col
2006). Se menciona que la prueba tiene la flexibilidad para establecer los criterios de
interpretación y puntos de corte necesarios para obtener un resultado positivo más
apropiado para una situación endémica de tuberculosis bovina específica (Vordermeiery

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
20
col, 2008.); además, también se menciona que la prueba es susceptible de ser
enriquecida con la adición de antígenos que sean más específicos para M. bovis.
En los últimos años ha habido un significativo progreso en la identificación de algunas
proteínas antigénicas de M. bovis, en un intento por identificar aquellas que sean más
específicas que las presentes en el PPD (Aagaard y col 2003). La utilización de algunas
proteínas recombinantes en la prueba del IFN-gamma como ESAT-6, CFP10, MPB70
y Rv3615c, ha permitido sugerir sustituir al PPD-B en las pruebas intradérmica y del
IFN-γ (Whelan y col 2010, Casal y col 2012).
Ahora bien, en relación a la utilización de los antígenos ESAT-6 y CFP10 con la prueba
Bovigam®, algunos estudios han revelado que en regiones con bajas prevalencias de la
infección, es posible mejorar la especificidad (99%) de la prueba in vitro del IFN-γ de
los animales reactores positivos a la prueba de la tuberculina, aunque la sensibilidad
esté por debajo de un 90%(Pollock y col, 2000), sin embargo, en regiones con altas
prevalencias hasta un 85% de los animales positivos fueron confirmados utilizando
estos dos antígenos en la prueba del IFN-γ (Aagaard y col 2010). lo cual demuestra la
necesidad de seguir buscando nuevos antígenos que puedan aumentar la sensibilidad de
la prueba en sangre en hatos ganaderos independientemente del estatus de la infección
en la región.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
21
8. JUSTIFICACIÓN
Un estudio comparativo reciente de los genomas de varias micobacterias para estudiar
la presencia y homología de un grupo de genes denominado regulón DosR, reveló que
existen diferencias significativas entre los genomas de M. bovis y M. avium. Se observó
que aunque algunos de estos genes están presentes en M. avium spp. paratuberculosis,
tienen diversos grados de identidad, y otros están totalmente ausentes en su genoma
(Lin y col 2009), lo que los hace fuertes candidatos para ser utilizados para discriminar
una infección entre M. bovis y una de M. avium. Cabe hacer notar que algunos de los
antígenos DosR, mostraron tener antigenicidad al ser usados para estimular la respuesta
inmune de pacientes con tuberculosis humana (Leyten y col 2006), provocando
satisfactoriamente la proliferación de células T y liberación de IFN-gamma, haciendo
viable su uso para el diagnóstico de la infección por M. tuberculosis.
Debido a que aún falta definir una mezcla de antígenos que sea totalmente específica
para detectar una infección por M. bovis en el ganado bovino en este trabajo se optó por
producir los antígenos recombinantes Mb1762c, PfkB y HspX pertenecientes al
regulón DosR de M. bovis, con la finalidad de comprobar si estos son realmente
antigénicos promoviendo la estimulación de la respuesta inmune celular con uso
potencial en el diagnóstico de la TB bovina.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
22
9. DEFINICIÓN DEL PROBLEMA
La ganadería bovina y la industria de la carne y leche representan una de las principales
actividades del sector agropecuario de muchos estados de la República Mexicana. Las
pérdidas económicas en el sector agropecuario por infecciones como la tuberculosis en
el ganado han originado una constante preocupación de la industria alimenticia y
productores nacionales e internacionales porque genera muchas pérdidas en la
producción de carne y leche limitando el comercio internacional.
La experiencia acumulada indica que la tuberculosis bovina es controlable y erradicable
mediante la detección y eliminación de los animales infectados, sin embargo, la alta
incidencia de tuberculosis en el ganado productor de leche hace al sacrificio una medida
de control difícil de aplicar por las pérdidas económicas que representa al productor.
Durante años, la prueba oficial para detectar a los animales con tuberculosis bovina ha
sido la tuberculina, sin embargo, esta prueba posee algunas desventajas relacionadas
con la sensibilidad de la prueba. Métodos comerciales de detección más recientes están
basados básicamente en la sensibilización de los leucocitos de los animales infectados a
secretar IFN-gama por su estimulación in vitro con mezclas de antígenos
micobacterianos de M. bovis (PPD-B) o de M. avium (PPD-A), con una sensibilidad
igualmente variable.
Debido a que aún falta identificar nuevos antígenos de M. bovis que puedan ser
utilizados para detectar una población infectada de una forma más confiable, esta
propuesta plantea utilizar los antígenos específicos PfkB, HspX y Mb1762c de este
microorganismo para aumentar la sensibilidad y el valor predictivo de la prueba de
diagnóstico comercial contra la tuberculosis bovina que está basada en la detección de
la producción de IFN-γ en sangre.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
23
10. HIPÓTESIS
Los antígenos recombinantes PfkB, HspX y Mb1762c de M. bovis son los bastante
antigénicos para utilizarlos en el diagnóstico de la TB bovina.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
24
11. OBJETIVOS
11.1. Objetivo General
Producir las proteínas recombinantes PfkB, HspX y Mb1762c de M. bovis y evaluar su
antigenicidad en sangre de bovinos naturalmente expuestos a la infección con M. bovis
11.2. Objetivos Específicos
� Purificar las proteínas recombinantes Mb1762c, PfkB y HspX producidas en
cualquiera de dos sistemas de expresión de E. coli.
� Comprobar la antigenicidad de las proteínas recombinantes mediante la
inducción de la producción de IFN-γ en sangre de bovinos de hatos ganaderos
con diferentes prevalencias de la infección.
� Comparar la estimulación de la respuesta celular producida por las proteínas
recombinantes contra la provocada por los antígenos PPD-B, ESAT-6 y CFP10.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
25
12. MATERIALES Y MÉTODOS
12.1. Diseño de oligonucleótidos y amplificación de los genes.
Para sintetizar las proteínas de M. bovis, primero se diseñaron los oligonucleótidos
específicos utilizando el software CLC BIO DNA workbench (Tabla 1) para poder
amplificar los genes Mb1762c, pfkB y hspX completos de las proteínas. Los oligos
fueron diseñados de acuerdo a las recomendaciones del manual para poder integrar el
amplificado a los vectores de expresión correspondientes de acuerdo a las
especificaciones de cada kit de expresión. Los fragmentos de los genes Mb1762c, pfkB
y hspX, de 633 pb, 1020 pb y 435 pb, respectivamente, fueron amplificados a partir de
DNA genómico de M. bovis de la cepa AN5 por una reacción de PCR) de alta fidelidad
utilizando la enzima PfuUltra High-Fidelity DNA polimerasa (Stratagene). La mezcla
de reacción de la PCR conteniauna concentración final de 200 nM de cada uno de los
oligos F y R 10 µM, 100 ng/µl de DNA templado , 2.5U de PFU DNA polimerasa
(Stratagene). La mezcla de reacción fue llevada a un volumen final de 50 µl con agua
desionizada esteril. Las condiciones para la PCR fueron las siguientes:
desnaturalización inicial a 95ºC por 2 min, 30 ciclos de 95ºC por 30 seg, 60ºC por 30
seg y 72ºc por un min, y un paso final de extensión de 72ºC por 10 min. se analizaron
por electroforesis 10 µL de las mezclas resultantes, cargándose en un gel de agarosa al
1% preparado con SYBR-Safe (Invitrogen), y se corrieron durante 45 min a un voltaje
constante de 80 V. Las bandas de los productos de amplificación en el gel se
visualizaron mediante un transiluminador de luz UV y un sistema analizador de
imágenes GelDoc XR (Bio-Rad). Los fragmentos de DNA amplificados fueron
purificados directamente de la mezcla de reacción con el kit MinElute PCR purification
kit de QIAGEN.
Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
26
Tabla 12.1. Oligonucleótidos usados para amplificar los genes completos de las
proteínas de este estudio de acuerdo a los vectores de clonación.
Vector pIVEX2.3d Gen Oligo Forward Oligo Reverse hpsX ttttttccatggccaccacccttc ttttttgatatcgttggtggaccggatct pfkB tatantaccatggcggagccagcagcggcgtg tatatacccgggtggcgaggcttccgggttaac Mb1762 ttttttccatgggaatcgccacaacccgc ttttttcccgggccgctgcgtgcagaa
Vector pET101/D-TOPO
Gen Oligo Forward Oligo Reverse
hpsX caccatggccaccacccttc gttggtggaccggatct
pfkB caccatgacggagccagcgg tggcgaggcttccgg
Mb1762 caccatgatcgccacaacccg ccgctgcgtgcagaa
12.2. Clonación en el vector pET101/D-TOPO para expresión usando cultivos de E.
coli.
Los fragmentos amplificados de los genes completos Mb1762c, pfkB y hspX, fueron
sujetos a clonación en el vector pET101/D-TOPO (Invitrogen) de acuerdo a la técnica
del manual de inserción del propio vector, y posteriormente se transformó cada uno de
los vectores recombinantes a la cepa de E. coli DH5αmediante electroporación a 200
Ohms, 25 µF y 2.5 kV en celdas para electroporación de 0.1 cm (Sigma), utilizando un
electroporador Gene Pulser II (BioRad). Las células transformadas se incubaron en 1 ml
de caldo LB durante 1 hr y después se recuperaron por centrifugación para sembrarse
todas en medio LB sólido con ampicilina 100 mM y finalmente se incubaron por 16 h a
37°C.
Para la comprobación de la presencia del plásmido recombinante, se eligieron algunas
colonias al azar y se cultivaron toda la noche en 3 ml de caldo LB, posteriormente se les
extrajo el DNA plasmídico con el kit GeneJET Plasmid Miniprep (Fermentas). Se
realizó un PCR usando el kit DreamTaq PCR Master Mix (Fermentas), utilizando los
oligos específicos de cada gen con el DNA plasmídico de las colonias recombinantes

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
27
con los insertos de cada gen, de acuerdo a las siguientes condicione: La mezcla de
reacción del PCR contenía, 26 ng de DNA moldedel templado de M. bovis, 0.5 µl de
cada oligo 10 µM, y 10 µl de Dream Tag Master Mix 2X; la mezcla se llevó a un
volumen final de 20 µl con agua desionizada esteril. Las condiciones de PCR fueron las
siguientes: desnaturalización inicial a 95ºC por 5 min; 30 ciclos de 95ºC por 30 seg,
60ºC por 30 seg y 72ºC por 40 seg; y un paso final de extensión a 72ºC por 7 min.. Los
amplificados fueron analizados mediante electroforesis en gel de agarosa como se
describió anteriormente.
12.3. Clonación en el vector pIVEX 2.3d para expresión en un sistema de traducción
libre de células.
Simultaneamentese diseñaron los oligos usando el software CLC BIO DNA workbench
para la amplificación del gen pfkB y clonarlo en el vector de expresión pIVEX.2.3d
(Roche) para la expresión en un sistema de traducción in vitro de E. coli. El gen
completo se amplificó mediante PCR de alta fidelidad con el kit PlatinumTaq DNA
polimerase High Fidelity (Invitrogen), de acuerdo a las siguientes condiciones: La
mezcla de reacción del PCR constó de 150 ng/µl de DNA templado, 0.5 µl de oligo F y
R 10 µM, y 10 µl de Dream Tag Master Mix 2X. La mezcla se llevó a un volumen final
de 20 µl con agua desionizada esteril. Las condiciones de la PCR fueron las siguientes:
desnaruralización inicial a 95ºC por 5 min; 30 ciclos a 95ºC por 30 seg, 60ºC por 30
seg, y 72ºC por 40 seg; y un paso final de extensión a 72 ºC por 7 min.
Posteriormente, el fragmento completo del gen pfkB se insertó en el vector pCR-TOPO
2.1 (Invitrogen) mediante una reacción de inserción de acuerdo al manual de
procedimiento del propio vector, y se transformó a la cepa de E. coli DH5αmediante el
procedimiento descrito anteriormente. Se seleccionaron las cepas sugestivas de contener
el vector recombinante, en agar LB con ampicilina y se obtuvieron los plásmidos
recombinantes de un cultivo en medio líquido mediante el procedimiento descrito
anteriormente. Se cortaron los dos vectores (pCR2.1-TOPO recombinante y pIVEX
2.3d) con las enzimas NcoI y SmaI (Promega) incubándolos en un Termoblock
(Labnet): primero a 37 ºC por 1 hr, y luego a 30 ºC por 1 hr, respectivamente,. Los

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
28
productos de restricción se corrieron en un gel de agarosa como se describió
anteriormente, y los fragmentos de interés
Los fragmentos se ligaron en una proporción vector digerido-inserto de 1:4, es decir,
2.5 µl del vector pIVEX 2.3d y 10 µl de fragmento pfkB liberado del pCR2.1-TOPO,
además, la mezcla llevó 4 µl de buffer de ligación 5X, 2 µl de DNA ligasa T4
(Invitrogen) y se llevó a un volumen del 20 µl con agua desionizada esteril, se incubó
por 18 h a 16°C. Posteriormente, la mezcla de ligación resultante, se transformó
directamente a células electrocompetentes de la cepa E. coli DH5α mediante el
procedimiento de electroporación descrito; las cepas transformantes se seleccionaron
como ya se indicó en medio LB y ampicilina. Después de aislar los plásmidos
recombinantes, se usó el kit libre de células RTS 100 E. coli HY (Roche) en el equipo
ProteoMaster de Roche para la síntesis de 20 µg de proteína.
12.4. Expresión de las proteínas recombinantes.
Los plásmidos recombinantes resultantes de la clonación de los tres genes en el vector
pET101/D-TOPO se transformaron a la cepa de E. coli BL21 Star (DE3) (Invitrogen)
mediante electroporación de la forma descrita. Las transformantes se seleccionaron en
agar LB con ampicilina. De las clonas obtenidas, se seleccionó sólo una colonia, la cual
se comprobó que contuviera el plásmido mediante aislamiento de plásmidos. Las
colonias se inocularon en 5 ml de caldo LB con ampicilina y se incubaron a por 16 h a
37°C con agitación de 250 rpm. Posteriormente, los cultivos de 5 ml se transfirieron a
200 ml de caldo LB en un matraz de 1 lt y se continuaron incubando a 37°C con
agitación de 250 rpm, hasta llegar a una densidad óptica de 0.5 unidades medida a 600
nm en un espectrofotómetro Genesys 10uv (ElectronCorporation). Inmediátamente
después, se les adicionó isoprpil-β-D-tiogalactósido (IPTG) a una concentración de 1
mM, continuando con la incubación en agitación durante 4 h más. Posteriormente, se
consecharon las células por centrifugación a 5000 rpm por 15 min a 4°C en botellas de
100 ml para rotor de centrifuga Universal 320R (Itettich Zentrifugen). Las pastillas
celulares se lisaron en buffer de lisis PBS (buffer salino de fosfatos con triton X-100 al
0.1% y un coctel de inhibidores de proteasas (Leupeptina al 10 mM, 4-2 Aminoetil
benceno sulfonilfloruro hidrocloruro al 0.1 mM, Antipaina al 50 mM, EDTA al 10 mM)

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
29
mediante 4 pulsos de 1 min de sonicación en hielo con un sonicador de punta metálica
Misonix XL-2000 y se separó la fracción soluble de la insoluble por centrifugación a
5000 rpm y 4°C, la fracción soluble posteriormente se filtró por una membrana de 0.45
µm de diámetro del poro. Ambas fracciones se guardaron a -20°C hasta su posterior uso
para comprobar la presencia de la proteína expresada mediante electroforesis y Western
blot.
Además, la expresión de la proteína PfkB clonada en el vector pIVEX 2.3d se llevó a
cabo mediante el kit RTS 100 E. coli HY (Roche) como se describe brevemente a
continuación. Quinientos ng de plásmido recombinante disueltos en 10 µl, se mezclaron
con 12 µl de un lisado de E. coli, 10 µl de mezcla de reacción, 12 µl de mezcla de
aminoácidos, 1 µl de metionina y 5 µl de buffer de reconstitución (Roche,
proporcionados por el kit); la mezcla final (50 µl), se incubó durante 6 h a 37°C y 300
rpm en un equipo RTS ProteoMaster (Roche). Posteriormente, se tomaron 5 µl de la
reacción final para su análisis por electroforesis y Western blot como se describe a
continuación. Los 45 µl restantes se guardaron a -20°C hasta su uso.
12.4.1. Análisis de las proteínas recombinantes mediante electroforesis en gel de
poliacrilamida (SDS-PAGE).
Una alícuota de 20 µl de las fracciones soluble e insoluble de los lisados celulares
obtenidos después de la inducción con IPTG, se incubaron con buffer de Laemmli 2X
(Tris base 12M, SDS 0.12M, glicerol 2M, β-mercaptoetanol al 10% y azul de
bromofenol) y se calentaron en baño María hirviendo por 5 min y posteriormente se
corrieron en un gel de poliacrilamida al 12% mediante electroforesis desnaturalizante
(SDS-PAGE) por 1 h a un voltaje constante de 120 V en una cámara Mini-Protean II
(BioRad). El gel se tiñó con azul de Coomassie y después de desteñirse con una
solución de ácido acético al 10% y metanol al 40%, se observó en un analizador de
imágenes GelDoc XR (BioRad).
12.4.2. Análisis de las proteínas recombinantes por Western blot con anticuerpo anti-
His6.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
30
Una réplica del gel de poliacrilamida con las fracciones subcelulares obtenidas, se
colocó en un sándwich sobre una membrana de nitrocelulosa Hybond ECL (Amersham)
y se sometió a una electrotransferencia durante 1 h a 100 V constantes en buffer de
transferencia (Tris base 25 mM, glicina 192 mM y metanol al 20%). Posteriormente, la
membrana se bloqueó durante 1 h a 4oC con amortiguador PBS-Tween 20 al 0.1% con
leche sin grasa al 5% y después de algunos lavados con el mismo amortiguador sin
leche, se incubó 1 h con el anticuerpo IgG anti-His6 de ratón (Roche) en una
concentración de 0.2 µg/ml en PBS-Tween con leche al 5%. Posteriormente se lavó de
nuevo la membrana con el buffer PBS-Tween por 3 ocasiones y se incubó durante 2 h
con un segundo anticuerpo anti-IgG de ratón conjugado a peroxidasa (Sigma) en
dilución 1:2000 en amortiguador PBS-Tween 20 al 0.05% con leche al 2.5%. Después
se lavó por 3 ocasiones más y se reveló con 25 ml de solución reveladora de PBS con
sustrato TMB (3, 3´, 5, 5´-tetrametilbenzideno, BioRad) al 0.3%, metanol al 16% y 30
µl de peróxido de hidrógeno al 30%, hasta la aparición de las manchas de color marrón.
12.5. Purificación de las proteínas recombinantes mediante columnas de afinidad.
Las fracciones solubles de los lisados bacterianos, se pasaron por una columna de
afinidad BIO-SCALE Mini Profinity IMAC (Bio-Rad) de 5X5 ml. La columna de
afinidad se equilibró previamente con 5 volúmenes de buffer de equilibrio (KCl 300
mM, KH2PO4 50 mM e imidazol 5 mM). Después, se pasó por gravedad la fracción
soluble con la proteína recombinante, y posteriormente se lavó la columna con 6
volúmenes de buffer de lavado (KCl 300 mM, KH2PO4 50 mM e imidazol 10 mM), por
último, se agregaron 10 volúmenes de buffer de elución (KCl 300 mM, KH2PO4 50 mM
e imidazol 250 mM) para despegar la proteína, recolectándose en alícuotas de 5 ml. Las
fracciones de lavado y elución, se guardaron a -20°C hasta su uso, y solamente unas
alícuotas de 20 µl de cada fracción se analizaron mediante electroforesis SDS-PAGE en
geles de poliacrilamida al 12%, tinción de Coomassie y Western blot, usando el mismo
procedimiento descrito anteriormente.
Con el fin de eliminar cualquier contaminante de la proteína se realizó una
electroelución,se sometieron las fracciones de elusión a una concentración por
deshidratación en vacío mediante SpeedVac a 45°C hasta tener alícuotas de 200 µl

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
31
aproximadamente en tubos de 1.5 ml. Posteriormente, las fracciones concentradas se
corrieron en geles preparativos de SDS-PAGE al 10% a 120 V por 1 hr, el gel se tiñó
por la técnica reversible de Zn-imidazol para visualizar las bandas más ricas de la
proteína recombinante y estas se cortaron del gel para colocar los fragmentos de gel con
la proteína en una celda para electroelución armada con los frits de filtración (BioRad).
Para la electroleución se corrió primero con buffer de corrida 1X con SDS durante 4 hr
a 12 mA, posteriormente con buffer de corrida 1X sin SDS durante otras 2 hr. Por
último, se recuperó con micropipeta la suspensión concentrada de la proteína de la celda
de electroelución. La solución de proteína en buffer Tris se colocó en membrana para
diálisis (Sigma) con diámetro de poro de 10 kDa y se dializó contra buffer PBS frío
estéril (1:200 v/v) con 4 cambios de buffer de cada 4 hr cada uno a 4°C. Finalmente, la
proteína pura se concentró en el tubo de diálisis por ósmosis reversa con polvo
Aquacide II (Calbiochem). La suspensión de proteína en PBS se guardó hasta su uso a -
70°C en alícuotas de 100 µl a una concentración de proteína aproximada de 50 mg/ml,
estimada mediante el método de Bradford adaptado a microplaca. Para la cuantificación
se tomó en cuenta una curva de estándar de seroalbúmina bovina (BSA) con un rango
de concentración entre 0 y 400 µg/ml, calculando la concentración directamente por el
método de regresión lineal simple.
12.6. Evaluación de la antigenicidad de las proteínas recombinantes en sangre de
bovino.
Para medir la antigenicidad de las proteínas recombinantes mediante la estimulación de
la inmunidad celular, se usó el kit de diagnóstico complementario de la tuberculosis
bovina conocido como prueba de Bovigam (Prionics), el cual se basa en medir la
producción de IFN-γ en sangre de bovinos. Se tomaron en tubos de heparina de litio
alrededor de 6 a 10 ml de sangre de bovinos naturalmente expuestos a M. bovis,
habiéndose muestreado animales de zonas con diferente prevalencia de la infección
algunos de los cuales dieron negativa la tuberculina pero están expuestos a animales
positivos, y otros, que en su mayoría tuvieron una prueba de la tuberculina positiva. La
sangre se estimuló tal y como lo indica el manual con los propios antígenos del kit
(PPD-B y PPD-A), y con las proteínas recombinantes HspX y PfkB, a una

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
32
concentración de 4 µg/ml, así como también en mezcla con los antígenos ESAT6 y
CFP-10.
Posteriormente, se utilizó solamente el plasma que se obtuvo por centrifugación de la
sangre, para medir indirectamente la proporción de IFN-γ producido después del
estímulo con los antígenos, midiendo la densidad óptica (450 nm) de la muestra
mediante la prueba de ELISA tipo sándwich de doble anticuerpo monoclonal, tal y
como lo indica el manual del kit. Los resultados se expresaron como índices de la
densidad óptica obtenida por antígeno o mezcla de antígenos usados para la
estimulación.
12.7. Análisis estadístico.
12.7.1 Cumplimiento de supuestos estadísticos
Se sometieron los datos de absorbancia de cada antígeno a diferentes pruebas para ver si
cumplían con los supuestos estadísticos para poder ser analizados mediante estadística
inferencial. Se realizó la prueba de Shapiro-Wilks para determinar la normalidad de los
datos. Para determinar la homogeneidad de varianzas se uso la prueba de Bartlett. Los
datos con un valor de p<0.05 no cumplen con el supuesto de normalidad o de
homogeneidad de varianzas.
12.7.2 Comparación de grupos
Se analizó si existían diferencias estadísticas significativas entre las muestras
estimuladas con los diferentes antígenos del grupo de alta prevalencia de TBb con el
grupo de baja prevalencia. Este análisis se realizó mediante la prueba U de Mann-
Whitney, que es la versión no paramétrica de la habitual prueba de t de Student. Un
valor de p<0.05 fue considerado significativo. El análisis estadístico se realizó con el
software SigmaStat 8.0®.
12.7.3 Curvas ROC (Receiver Operating Characteristics)
Evaluamos mediante curvas ROC el potencial de diagnóstico, calculando la sensibilidad
y la DO de corte, tanto para el antígeno PPD-B del Bovigam®, como para las distintas
mezclas de antígenos recombinantes. Las curvas ROC (Receiving Operating

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
33
Characteristic Curve) son particularmente útiles para comparar el desempeño de dos o
más pruebas diagnósticas (Greiner y col, 2000).Las curvas ROC se realizaron con
SigmaPlot® 10.0 y un valor de p<0.05 se consideró significativo. Estas curvas
correlacionan la sensibilidad (proporción de verdaderos positivos) con la especificidad
(falsos positivos) en una prueba diagnóstica. El área bajo la curva indica la probabilidad
de clasificar correctamente un par de muestras tomadas al azar. El punto de corte y la
sensibilidad fueron calculados a una especificidad del 85% en la curva ROC de cada
antígeno.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
34
13. RESULTADOS
13.1. Selección de antígenos
Un estudio comparativo reciente de los genomas de varias micobacterias para estudiar
la presencia y homología de un grupo de genes denominado regulón DosR, reveló que
existen diferencias significativas entre los genomas de M. bovis y M. avium. Se observó
que aunque algunos de estos genes están presentes en M. avium spp. paratuberculosis,
tienen diversos grados de identidad, y otros están totalmente ausentes en su genoma
(Lin y col, 2009), lo que los hace fuertes candidatos para ser utilizados para discriminar
una infección entre M. bovis y una de M. avium. Cabe hacer notar que algunos de los
antígenos DosR, mostraron tener antigenicidad al ser usados para estimular la respuesta
inmune de pacientes con tuberculosis humana (Leyten y col, 2006), provocando
satisfactoriamente la proliferación de células T y liberación de IFN-gamma, haciendo
viable su uso para el diagnóstico de la infección por M. tuberculosis.
Así, se seleccionaron 3 proteínas del regulón DosR de M. bovis (Tabla 1) ya que están
ausentes o con muy poca homología en M. avium spp. paratuberculosis (Tabla 2), lo
que los hace fuertes candidatos para comprobar si poseen alguna antigenicidad en
sangre de bovino Infectado por M. bovis.
Tabla 13.1. Proteínas del regulón DosR de M. bovis consideradas para el estudio.
Antígeno de
M.
tuberculosis
Antígeno de
M. bovis
Gen Peso
molecular
(kDa)
Tamaño
del gen
(pb)
Producto
Rv1733c Mb1762c mb1762c 22.4 633 Probable proteína
transmembranal conservada
Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
35
Rv2029c Mb2054c pfkB 35.4 1020 Fosfofructocinasa
Rv2031c Mb2057c hspX, acr 16.3 435 Proteína de choque térmico
HspX ( α-cristalina)
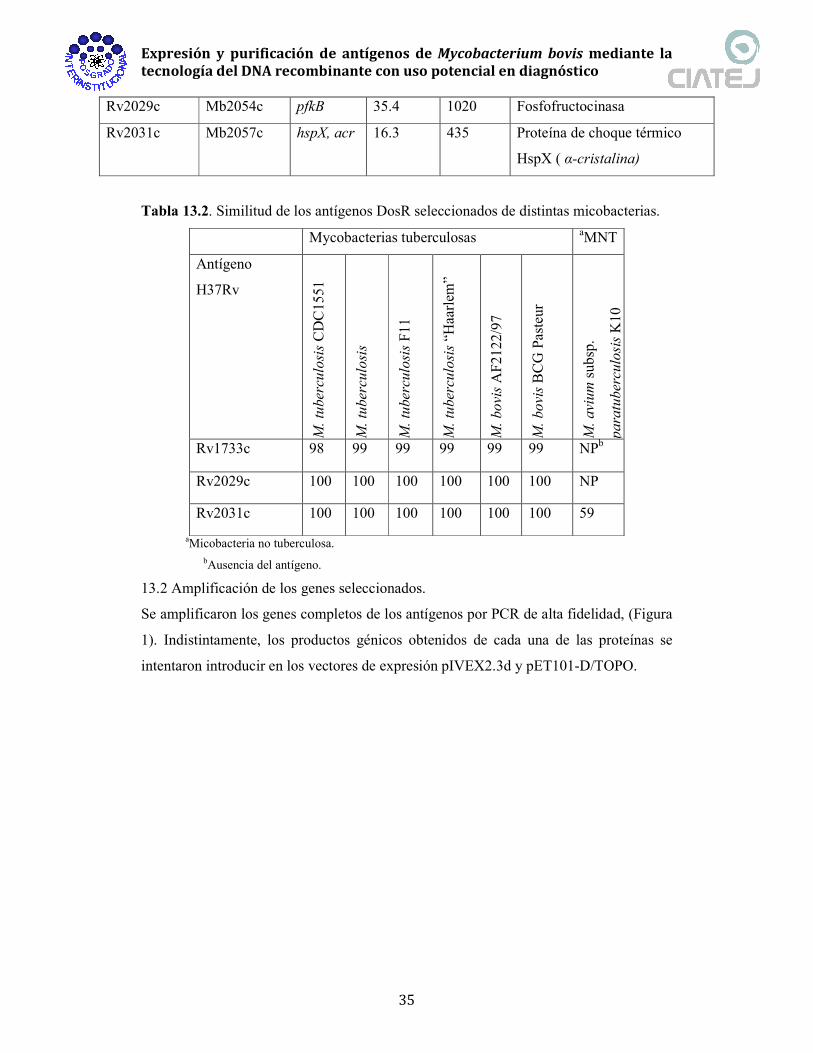
Tabla 13.2. Similitud de los antígenos DosR seleccionados de distintas micobacterias.
Mycobacterias tuberculosas aMNT
Antígeno
H37Rv
M. tuberculosis CDC15
51
M. tuberculosis
M. tuberculosis F1
1
M. tuberculosis “H
aarlem
”
M. bovis AF21
22/97
M. bovis BCG Pasteur
M. avium sub
sp.
paratuberculosis K10
Rv1733c 98 99 99 99 99 99 NPb
Rv2029c 100 100 100 100 100 100 NP
Rv2031c 100 100 100 100 100 100 59
aMicobacteria no tuberculosa. bAusencia del antígeno.
13.2 Amplificación de los genes seleccionados.
Se amplificaron los genes completos de los antígenos por PCR de alta fidelidad, (Figura
1). Indistintamente, los productos génicos obtenidos de cada una de las proteínas se
intentaron introducir en los vectores de expresión pIVEX2.3d y pET101-D/TOPO.
Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
36
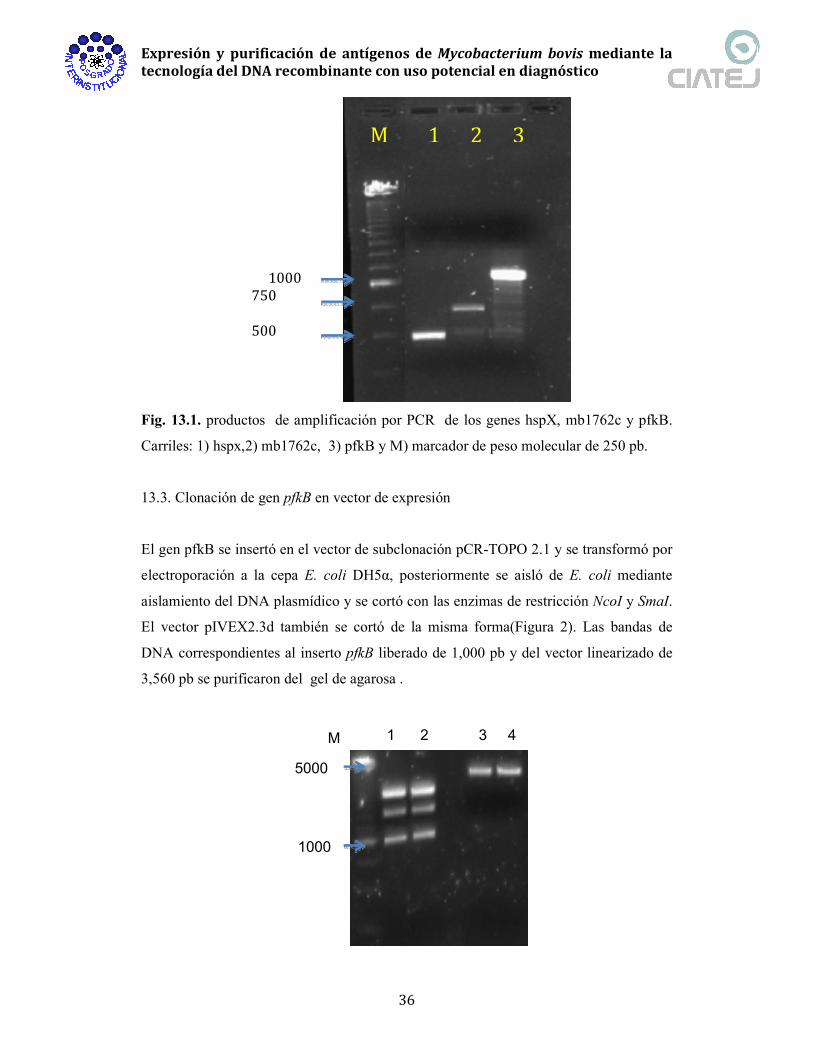
Fig. 13.1. productos de amplificación por PCR de los genes hspX, mb1762c y pfkB.
Carriles: 1) hspx,2) mb1762c, 3) pfkB y M) marcador de peso molecular de 250 pb.
13.3. Clonación de gen pfkB en vector de expresión
El gen pfkB se insertó en el vector de subclonación pCR-TOPO 2.1 y se transformó por
electroporación a la cepa E. coli DH5α, posteriormente se aisló de E. coli mediante
aislamiento del DNA plasmídico y se cortó con las enzimas de restricción NcoI y SmaI.
El vector pIVEX2.3d también se cortó de la misma forma(Figura 2). Las bandas de
DNA correspondientes al inserto pfkB liberado de 1,000 pb y del vector linearizado de
3,560 pb se purificaron del gel de agarosa .
5000
1000
1 2 3 4 M
M 1 2 3
1000
750
500

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
37
Fig. 13.2. productos de digestión con enzimas de restricción NcoI y SmaI. Carriles: 1 y
2) vector pCR-TOPO 2.1 con inserto del gen pfkB ; 3 y 4) vector pIVEX 2.3d y M)
marcador de peso molecular d 250 pb.
Posteriormente a la subclonación de pfkB en pIVEX2.3d, se procedió a comprobar la
construcción cortando el vector con la enzima de restricción SmaI esperando un
fragmento de 4500 pb correspondiente a pIVEX2.3d+pfkB (Figura 3). También se
comprobó el inserto liberando nuevamente el inserto pfkB del vector con las enzimas de
restricción SmaI y NcoI, (Figura 3).
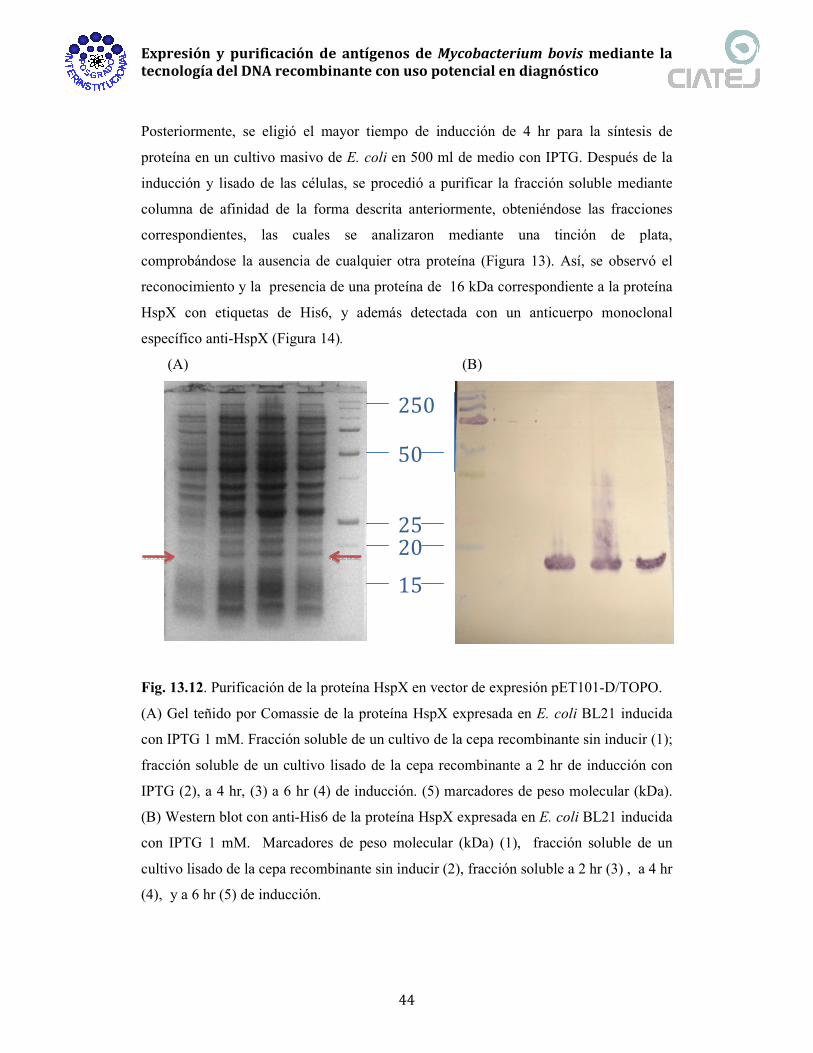
Fig. 13.3. Productos de digestión del vector recombinante pIVEX 2.3d+pfkB con las
enzimas SmaI y NcoI. El vector recombinanate se incubó durante 1 hr a 37°C con las
enzima de restricción. . Carriles: 1) pIVEX2.3d+pfkB digerido con SmaI y 2)
pIVEX2.3d+pfkB digerida con SmaI y NcoI. M) marcador de peso molecular de 1 kb.
A su vez, para confirmar la presencia del gen pfkB en el vector recombinante, se realizó
una PCR lo cual confirma la presencia del gen en el vector de expresión(Figura 4).
M 1 2
1000
3000
2000
M 1 2

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
38
Fig. 13.4. Producto de PCR de pIVEX+pfkB. Carriles: 1) pIVEX2.3d+pfkB, 2)
pIVEX2.3d. M) marcador de peso molecular de 250 pb.
13.4. Clonación de gen hspx en vector de expresión pIVEX2.3d
Por otro lado, después de amplificar por PCR el gen hspX, se clonó el fragmento
amplificado en el vector pIVEX2.3d linearizado, posteriormentese comprobó la
presencia del gen en el vector con la enzima de restricción SmaI observándose una sola
banda de….correspondiente al vector más inserto (Figura 5).
Fig. 13.5. Digestión de pIVEX 2.3d+hspX con enzima SmaI. Carriles: 1, 2 y 3) vector
recombinante pIVEX+hspX; M) marcador de peso molecular de 1 kb.
1000
500
250
1 2 3 M
3500
2000
1000

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
39
Por PCR confirmatorio para amplificar el gen hspX directamente del vector pIVEX2.3d-
hspX obtenido a fin de corroborar dicha construcción, se observó un amplificado de
435 pb, del gen hspX en el vector (Figura 6). Lo cual indica que si hubo inserción del
gen hspX en el vector de expresión.
Fig. 13.6. Productos de PCR de pIVEX2.3d+hspX. Carriles: 1, 2 y 3) pIVEX+hspX, 4)
pIVEX2.3d M) marcador de peso molecular de 250 pb.
13.5. Síntesis de las proteínas pfkB y hspx en sistema RTS ProteoMaster
Una alícuota de 10 µl de proteína sintetizada en el equipo RTS-Proteomaster (Roche)
usando cada uno de los dos vectores recombinantes pIVEX2.3d+pfkB y hspX fue
sometida a electroforesis en SDS-PAGE al 12% y transferida a membrana de
nitrocelulosa para poder ser detectada con anticuerpos anti-His6 mediante Western blot.
La proteína PfkB de 37 kDa pudo ser detectada, confirmando su síntesis in vitro (Figura
7), sin embargo la proteína HspX de 16 kDa no fue posible observarla mediante Western
blot (dato no mostrado), existiendo la posibilidad de que el gen no se haya insertado en
el marco de lectura adecuado para que se le incorporen las etiquetas His6.
M 1 2 3 4
1000
500
250

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
40
Fig. 13.7. Western-blot con anticuerpo anti-His6 de la proteína PfkB sintetizada en RTS
Proteo Master. M) marcadores de peso molecular (kDa).
13.6. Purificación de la proteína PfkB sintetizada con el sistema RTS Proteo Master
Ya que la proteína PfkB se sintetizó correctamente con la etiqueta de histidinas, se
realizó la purificación de acuerdo al manual de procedimiento de las columnas de
afinidad His-Spin Proteinminiprep (ZymoResearch). La proteína PfkB de 37 kDa de
interés no se purificó completamente pura ya que eluye con al menos otra proteína
quizás con una cierta afinidad a la matríz de afinidad (Figura 8). Por otro lado, también
observamos que la cantidad de proteína es muy pequeña ya que en la elución no la
podemos observar muy bien debido a que la banda es muy tenue.
250
75
50
37
25
kDa M PfkB
M 1 2 3 4 5 6 7

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
41
Fig. 13.8. Gel de poliacrilamida teñido con azul de Coomassie de las fracciones eluidas
por la columna de afinidad donde se pasó la mezcla de proteínas sintetizadas en el
Proteo Master. Carriles: M) marcador de peso molecular (kDa), 1) mezcla completa, 2)
fracción no unida, 3) fracción del lavado 1, 4) lavado 2, 5) lavado 3, 6) fracción de
elución 1 y 7) elución 2.
13.7. Purificación de la proteína PfkB expresada en E. coli BL21
Debido a que la cantidad de proteína PfkB obtenida en el sistema RTS-Proteo Master
fue inferior a la esperada, se decidió transformar por electroporación el vector
recombinante a E. coli BL21, para la producción de proteína. El vector pIVEX 2.3d
también posee los promotores de la transcripción para E. coli BL21, basados en el
sistema de transcripción de la RNA polimerasa del fago T7, para la expresión de las
proteínas in situ.
Ya que la proteína recombinante PfkB contiene etiquetas His6x y se realizó un cultivo
masivo de E. coli BL21, para su purificación se utilizarón columnas de afinidad Bio-
Scale Mini Profinity IMAC con capacidad de flujo para 5 ml . Se recolectó cada una de
las fracciones, , y se tomó una alícuota de 10 µl para comprobar la presencia de la
proteína PfkB mediante tinción y Western blot En el gel de poliacrilamida al 12%
teñido con plata se muestra la banfa de 37 kDa de PfkB totalmente pura (Figura 9).
37 PfkB

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
42
Fig. 13.9. Gel teñido con plata de las fracciones purificadas de la proteína PfkB por
columna de afinidad. L) lisado total de E. coli, 1) eluido 1, 2) eluido 2, 3) eluido 3, PM)
marcadores kDa
Posteriormente, mediante un Western blot se comprobó la presencia de la proteína en
las fracciones de elución, así como también en las fracciones previas de lavados, es
probable que la presencia en todas las fracciones de purificación sea debida a la alta
concentración de proteína recombinante producida en el cultivo de células masivo
(Figura 10).
Fig 13.10. Western blot con anti-His6 después de la purificación por una columna de
afinidad. Carriles: 1) extracto total, 2) fracción no unida, 3) lavado 1, 4) lavado 2, 5)
elución 1 y 6) elución 2. M) marcador de peso molecular (kDa),
L 1 M 2 3
50
15
20
25
37 PfkB
1 2 M 3 4 5 6
37 PfkB

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
43
13.8. Clonación y purificación de la proteína HspX en vector de expresión pET101-
D/TOPO.
La segunda opción para intentar producir las proteínas, fue clonar los genes hspX y
Mb1762c en el vector de expresión pET101-D/TOPO para la expresión en E. coli.
.Posterior a la clonación y transformación química a la cepa BL21, para comprobar si
realmente las colonias adquirieron el vector con el inserto del gen de interés, se realizó
el aislamiento de plásmidos de las clonas seleccionadas en agar LB con ampicilina y el
consecuente PCR, en las cuales se observó la amplificación de los genes Mb1762c y de
hspX (Figura 11).
Fig. 13.11. PCR de clonas obtenidas en vector pET101/D-TOPO+hspX y Mb1762c.
Carriles: 1) plásmido recombinante aislado de la clona 1, 2) plásmido recombinante
aislado de la clona 2; 3 y 4) PCR de clonas con gen Mb1762c; 5 y 6) PCR de clonas con
gen hspX . M) marcador de peso molecular de 1kb.
Una vez seleccionadas las clonas positivas de tener el gen hspX, se realizó una cinética
de expresión para determinar el tiempo óptimo de expresión de la proteína en una de
las clonas con el vector recombinante pET101-hspX. Se tomaron muestras de 1 ml del
cultivo a diferentes tiempos de la cinética de inducción, se lisó el cultivo y se corrió en
un gel de poliacrilamida comprobando la presencia de la banda de la proteína
recombinante expresada (Figura 12 y B).
M 1 2 3 4 5 6
5000
600
400
Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
44
Posteriormente, se eligió el mayor tiempo de inducción de 4 hr para la síntesis de
proteína en un cultivo masivo de E. coli en 500 ml de medio con IPTG. Después de la
inducción y lisado de las células, se procedió a purificar la fracción soluble mediante
columna de afinidad de la forma descrita anteriormente, obteniéndose las fracciones
correspondientes, las cuales se analizaron mediante una tinción de plata,
comprobándose la ausencia de cualquier otra proteína (Figura 13). Así, se observó el
reconocimiento y la presencia de una proteína de 16 kDa correspondiente a la proteína
HspX con etiquetas de His6, y además detectada con un anticuerpo monoclonal
específico anti-HspX (Figura 14).
(A) (B)
Fig. 13.12. Purificación de la proteína HspX en vector de expresión pET101-D/TOPO.
(A) Gel teñido por Comassie de la proteína HspX expresada en E. coli BL21 inducida
con IPTG 1 mM. Fracción soluble de un cultivo de la cepa recombinante sin inducir (1);
fracción soluble de un cultivo lisado de la cepa recombinante a 2 hr de inducción con
IPTG (2), a 4 hr, (3) a 6 hr (4) de inducción. (5) marcadores de peso molecular (kDa).
(B) Western blot con anti-His6 de la proteína HspX expresada en E. coli BL21 inducida
con IPTG 1 mM. Marcadores de peso molecular (kDa) (1), fracción soluble de un
cultivo lisado de la cepa recombinante sin inducir (2), fracción soluble a 2 hr (3) , a 4 hr
(4), y a 6 hr (5) de inducción.
20
25
50
250
15

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
45
Fig. 13.13. Gel teñido con plata de fracciones purificadas de la proteína HspX por
columna de afinidad. L) lisado total de E. coli, 1) fracción de elución 1, 2) fracción de
elución 2, 3) fracción de elución 3, 4) fracción de elución 4, M) marcadores de peso
molecular kDa.
Fig. 13.14. (A) Western blot con anticuerpo monoclonal anti-HspX. Carriles: 1) eluido
4, 2) eluido 3, 3) eluido 2, 4) eluido 1, M) marcador de peso molecular (kDa) y 5) lisado
total. (B) Western blot con anticuerpo anti-His.. Carriles: 1) lisado total, M) marcador
de peso molecular (kDa), 2) elución 1, 3) elución 2, 4) elución 3, y 5) elución 4.
250
37
20
a) b)
L M 1 2 3 4
10
15
20
25 37
HspX

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
46
13.9. Clonación y expresión de la proteína Mb1762c en vector de expresión pET101/D-
TOPO
Como ya se mencionó , después de la clonación y transformación a la cepa BL21, para
comprobar si realmente las colonias adquirieron el vector con el inserto del gen de
interés, se realizó una PCR con las clonas seleccionadas en las cuales se observó la
amplificación del fragmento completo del gen Mb1762c (Figura 15) .
Fig. 13.15. PCR de clonas obtenidas pET101/D-TOPO+Mb1762c. Carriles: 1)
Marcador de peso molecular de 250 pb, 2) amplificado de Mb1762c de la clona 1, 3)
amplificado de Mb1762c de la clona 2.
Para la expresión del la proteína recombinante se llevó a cabo una inducción piloto en
un matraz de 20 mL de caldo LB, después de lisar el paquete celular, se realizó el
análisis de la proteína; primeramente se analizó la parte soluble de la proteína inducida,
obteniendo resultados no satisfactorios ya que la proteína no se alcanzó a percibir por
tinción en geles de poliacrilamida, ni por Western blot con anticuerpo anti-His6 (dato
no mostrado).
Por otro lado, al analizar la fracción insoluble del lisado celular agregando hidrocloruro
de guanidina 6 M al buffer de lisis, para solubilizar la proteína en caso de que estuviera
en cuerpos de inclusión. Aún así tampoco observamos la presencia de la proteína (dato
no mostrado).
M 1 2
1000
500
250

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
47
Debido a que en diversos intentos, variando las condiciones de la inducción de la
expresión, no encontramos la proteína del tamaño esperado, como último paso se
mandó a secuenciar el plásmido pET101-Mb1762c; y lo que se encontró, después de
conocer la secuencia y aplicar un análisis BLAST y comparación con la base de datos
GeneBank (NCBI), es que el gen clonado fue el de la proteína PhdA (subunidad A de la
piruvato deshidrogenasa de M. bovis) (dato no mostrado).
13.10 Evaluación de la antigenicidad de las proteínas recombinantes en sangre de
bovino
Como ya se describió, la evaluación de la antigenicidad de las proteínas recombinantes
obtenidas, se llevo a cabo utilizando para tal fin el kit de diagnóstico BOVIGAM
(Prionics). Las muestras de sangre de bovinos de hatos con diferentes prevalencias, se
estimularon con PPD-B, PDD-A y con una solución fisiológica (PBS) de tal manera
que se usaron como estándares para comparar la estimulación de la producción de IFN-γ
por las proteínas recombinantes en la sangre. .
Actualmente de acuerdo a la prueba comercial Bovigam® para considerar un animal
positivo a tuberculosis, la relación de la densidad óptica (DO) a 450 nm, producto de la
estimulación del IFN-γ por el PPD-B, menos la DO de aquella obtenida por la adición
de PBS, debe ser mayor o igual a 0.1, y además, la relación entre la DO producto de la
estimulación del IFN-γ por el PPD-B, menos la DO provocada por la estimulación el
PPD-A, igualmente debe ser mayor o igual a 0.1.
Así, de acuerdo a los criterios antes mencionados, para la clasificación de bovinos
infectados y bovinos no infectados con tuberculosis bovina, además nosotros evaluamos
mediante curvas ROC el potencial de diagnóstico, calculando la sensibilidad y la DO de
corte, tanto para el antígeno PPD-B del Bovigam®, como para los antígenos
recombinantes PfkB, HspX y ESAT6/CFP10 solos y en mezcla.Por separado en cada
muestra de sangre, se estimuló con 4 µg/ml de proteínas recombinantes PfkB, HspX,
ESAT-6/CFP10, así como también por la mezcla compuesta por 2 µg/ml de PfkB, 2
µg/ml de HspX y 10 µg/ml de PPD-B.
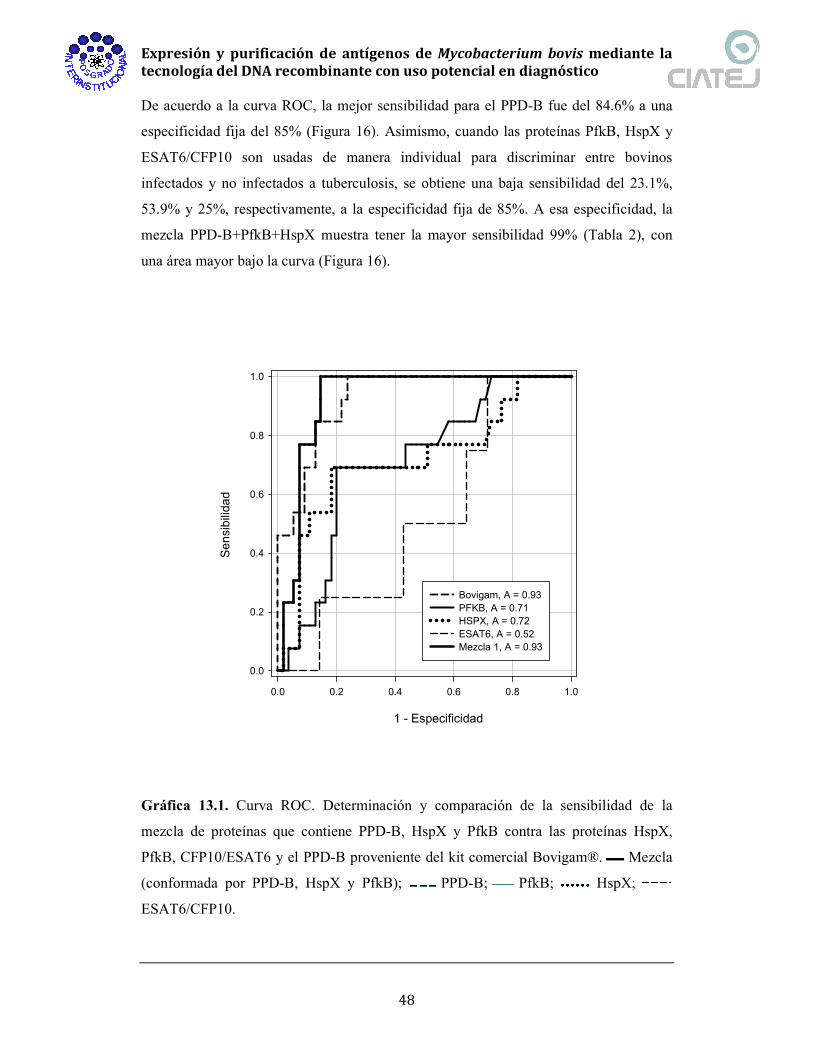
Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
48
De acuerdo a la curva ROC, la mejor sensibilidad para el PPD-B fue del 84.6% a una
especificidad fija del 85% (Figura 16). Asimismo, cuando las proteínas PfkB, HspX y
ESAT6/CFP10 son usadas de manera individual para discriminar entre bovinos
infectados y no infectados a tuberculosis, se obtiene una baja sensibilidad del 23.1%,
53.9% y 25%, respectivamente, a la especificidad fija de 85%. A esa especificidad, la
mezcla PPD-B+PfkB+HspX muestra tener la mayor sensibilidad 99% (Tabla 2), con
una área mayor bajo la curva (Figura 16).
0.0 0.2 0.4 0.6 0.8 1.0
1 - Especificidad
Sensibilidad
0.0
0.2
0.4
0.6
0.8
1.0
Bovigam, A = 0.93
PFKB, A = 0.71
HSPX, A = 0.72
ESAT6, A = 0.52
Mezcla 1, A = 0.93
Gráfica 13.1. Curva ROC. Determinación y comparación de la sensibilidad de la
mezcla de proteínas que contiene PPD-B, HspX y PfkB contra las proteínas HspX,
PfkB, CFP10/ESAT6 y el PPD-B proveniente del kit comercial Bovigam®. Mezcla
(conformada por PPD-B, HspX y PfkB); PPD-B; PfkB; HspX;
ESAT6/CFP10.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
49
Tabla 13.3. Sensibilidad de los antígenos y valores de corte requeridos.
Antígeno ______________________Especificidad (%)a__________________
_____________________________80 85 90 93__________
PPDB 84.6 (0.17) 84.6 (0.21) 69.2 (0.24) 53.9 (0.29)
PfkB 69.2 (0.18) 23.1 (0.31) 15.4 (0.36) 7.7 (0.38)
HspX 69.2 (0.19) 53.9 (0.22) 46.2 (0.25) 46.2 (0.26)
ESAT-6/CFP10 25.0 (0.34) 25.0 (0.35) 15.3 (0.52) 15.3 (0.52)
PPDB+PfkB+HspX 99.0 (0.23) 99.0 (0.25) 76.9 (0.29) 76.9 (0.34)
aSensibilidad de los antígenos expresada en % con los valores de corte requeridos
entre paréntesis (DO 450 nm), usando los datos obtenidos de los animales
sensibilizados en campo (n=68).
Así, cuando se utiliza la mezcla de proteínas conformada por PPD-B+HspX+PfkB, a la
especificidad de 85% y un punto de corte de 0.25, se obtiene una diferencia de 14.4% de
incremento en la sensibilidad (99%), con respecto a la sensibilidad obtenida por el PPD-
B (84.6%) usado de manera individual para estimular la sangre, lo cual indica que con
esta mezcla de antígenos hay una mayor probabilidad de encontrar a los animales
infectados realmente por M. bovis detectados como positivos (Figura 17).
Gráfica 13.2. Comparación estadística de animales infectados (A) de no infectados (B)
con la estimulación del IFN-γ por el PPD-B proveniente del kit comercial Bovigam®,
así como los animales infectados (C) de no infectados (D) debido a la estimulación del
IFN-γ con la mezcla que contiene PPD-B, HspX y PfkB. Se empleó la prueba U de
Mann-Whitney. Los asteriscos indican diferencias significativa (* < 0.001 y **
<.000001).

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
50
Una vez determinado el punto de corte, se procedió a determinar si existieron
diferencias estadísticas significativas en los ensayos de liberación de IFN-γ estimulados
con el PPD-B y con la mezcla PPD-B+HspX+PfkB usando el punto de corte de 0.25
obtenido, empleando la prueba no paramétrica U de Mann-Whitney. Los valores de p
<0.05 fueron considerados significativos. Los análisis estadísticos se realizaron con
SigmaStat® v8.0. Comparando los animales que dan positivos contra los que dan
negativos empleando la prueba de Bovigam®, se tiene una diferencia estadística
significativa con un valor de p < 0.001, sin embargo, como se puede notar, existe un
número de muestras que se traslapan entre los que deben ser positivos y los negativos
(Figura 17). Mientras que con la mezcla del PPD-B+HspX+PfkB, se tiene una
significancia estadística aún mayor con un valor de p <0.000001 dejando notar que
existe una mejor separación, y a su vez, ningún traslape entre los animales positivos y
los negativos con la nueva mezcla (Figura 17), lo cual también nos indica que hay
menor probabilidad de detectar animales falsos positivos usando la mezcla de PPD-
B+HspX+PfkB con el nuevo criterio de interpretación, que cuando se utiliza el único
componente antigénico (PPD-B) de la prueba comercial.
Una vez que se demostró que la adición de las proteínas HspX y PfkB al PPD-B
incrementan un 14.4% la sensibilidad del kit comercial Bovigam®, se procedió a
validar la prueba con muestras de 91 bovinos, comparando la mezcla de antígenos
(PPD-B+HspX+PfkB) con respecto al criterio del kit Bovigam®. Con el kit Bovigam®
se detectaron sólo 18 infectados, mientras que con la mezcla conformada por PPD-
B+HspX+PfkB y el nuevo criterio de interpretación a un punto de corte de D.O. de
0.25, se logran detectar 32 infectados (Figura 18); cabe mencionar que dentro de estos
32 detectados, 18 son los mismos que se logran detectar con el criterio del kit
Bovigam®.
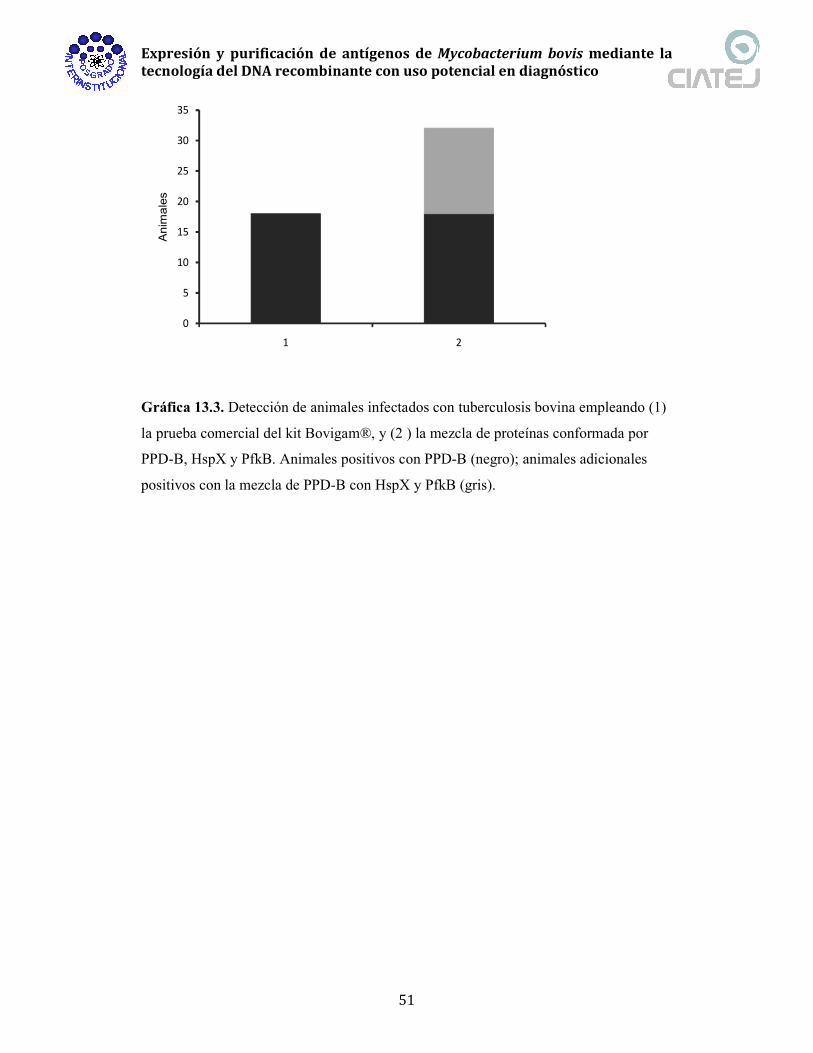
Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
51
0
5
10
15
20
25
30
35
1 2
Animales
Gráfica 13.3. Detección de animales infectados con tuberculosis bovina empleando (1)
la prueba comercial del kit Bovigam®, y (2 ) la mezcla de proteínas conformada por
PPD-B, HspX y PfkB. Animales positivos con PPD-B (negro); animales adicionales
positivos con la mezcla de PPD-B con HspX y PfkB (gris).

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
52
14. DISCUSION
Para evaluar el potencial de las proteínas recombinantes como marcadores de
diagnóstico, se compararon las características antigénicas de las diferentes proteínas
recombinantes con el antígeno comercial usado para el diagnóstico de la TBb (PPD-B).
Se incluyeron todos los animales muestreados, los cuales todos mostraron un resultado
positivo a la prueba de la Tuberculina intradérmica simple o TST (de sus siglas en
inglés), pero que no forzosamente dieron una prueba positiva a la prueba
complementaria del IFN-γ en sangre (BOVIGAM).
Dado que estos no se distribuyen normalmente, a los resultados obtenidos
mediante la prueba in vitro del BOVIGAM para medir la estimulación de la secreción
de IFN-γ en todas las muestras, se les realizó un análisis estadístico mediante el uso de
una prueba no paramétrica Mann-Whitney, al comparar la liberación de IFN-γ obtenida
con los antígenos PfkB, HspX, ESAT-6/CFP10, solos y en mezcla de antígenos con
PPD-B, revelando que hubo una mayor liberación de IFN-γ cuando la mezcla
conformada por PfkB, HspX y PPD-B se utilizó para estimular, que aquella dada solo
por el PPD-B.
La medición del IFN-γ producido por las proteínas recombinantes en el plasma,
muestran que la mezcla de proteínas dan mayor respuesta que el PPD-B. Nuestros
hallazgos concuerdan con lo reportado por otros al trabajar con proteínas puras, donde
hay mayor probabilidad de incrementar la antigenicidad frente al PPD-B (Whelan y
col. 2010).
Para calcular los valores de la nueva sensibilidad y especificidad de la prueba
optimizada con la mezca de antígenos recombinantes y PPD-B, se realizó una curva
ROC con los todos los datos obtenidos de la medición del IFN-γ con las distintas
opciones antigénicas evaluadas a una misma D.O. de 450 nm, donde se observa que en
comparación a todos los antígenos probados, la mezcla optimizada da un área bajo la
curva mayor. Dicha área bajo la curva puede interpretarse como la probabilidad de que
la nueva mezcla de PfkB, HspX y PPD-B al probarse con la sangre de cualquier animal
de un resultado mayor o parecido al PPD-B, aumentando su sensibilidad. Cuando las
D.O. de todos las muestras de sangre de bovinos fueron aplicadas al análisis ROC, se

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
53
observa que la mezcla de antíganos tuvo un área bajo la curva de 0.9259 (95% intervalo
de confianza (IC), Dando un valor muy parecido al PPD-B que mostró un valor de
0.9273, estimando una sensibilidad de 99% y una especificidad de 85% para la mezcla
y para el PPD-B se estimo una sensibilidad de 92% a una especificidad de 84.6%.
A su vez, de acuerdo a la curva ROC para la mezcla compuesta de PPD-B+HspX+PfkB,
se determinó un punto de corte de 0.25 de D.O. para obtener la sensibilidad del 99%, lo
cual no se contrapone a los principios de la prueba, ya que se menciona que la prueba
tiene la flexibilidad para establecer los criterios de interpretación y puntos de corte
necesarios para obtener un resultado positivo más apropiado para una situación
endémica de tuberculosis bovina específica (Vordermeier y col. 2008, Aranaz y col,
2006); además, también se menciona que la prueba es susceptible de ser enriquecida
con la adición de antígenos que sean más específicos para M. bovis, para lo cual se
tienen que modificar los criterios de interpretación basados en los puntos de corte
obtenidos a la mejor sensibilidad establecida (Aagaard y col. 2006, Cockle 2006).
En un trabajo muy reciente realizado por Casal y col. (2012), encontraron que al evaluar
a las proteínas ESAT-6, CFP10 y Rv3615c, en forma de péptidos o de proteínas
recombinantes, no hubo un aumento en la sensibilidad al probarlos tanto de una forma
intradérmica como de una forma in vitro, con sensibilidades entre el 52.2% para el
coctel peptídico y 47.8% para el coctel de proteínas. Lo cual demuestra que es necesario
seguir analizando más proteínas específicas para elevar la sensibilidad de las pruebas
usando antígenos específicos. En otro trabajo en regiones con bajas prevalencias de la
infección, se observó que un 60% de los animales positivos a las pruebas de la
tuberculina e IFN-γ, no respondieron a los antígenos específicos ESAT-6 y CFP10, sin
embargo, en regiones con altas prevalencias hasta un 85% de los animales positivos
fueron confirmados utilizando estos dos antígenos en la prueba (Aagaard y col. 2010),
lo cual demuestra la necesidad de seguir buscando nuevos antígenos que puedan
aumentar la sensibilidad de la prueba en sangre en hatos ganaderos independientemente
del estatus de la infección en la región.
En los últimos años ha habido un significativo progreso en la identificación de algunas
proteínas antigénicas de M. bovis, en un intento por identificar aquellas que sean más
específicas que las presentes en el PPD (Aagaard y col. 2003). La utilización de algunas

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
54
proteínas recombinantes en la prueba del IFN-γ como ESAT-6, CFP10 y MPB70, ha
permitido sugerir sustituir al PPD-B en las pruebas intradérmica y del INF-γ (Whelan y
col. 2010).
La prueba actual para el diagnóstico de la TBb en sangre (BOVIGAM) está basada en la
sensibilización de los linfocitos usando una mezcla compleja de antígenos, el PPD-B y
PPD-A, para la identificación de los animales infectados, por lo que algunos reportes
indican la identificación de falsos positivos producto de coinfecciones con
micobacterias ambientales que cruzan antigénicamente con el PPD (Aranaz y col.
2006), con una sensibilidad que varía entre el 69 y el 99%, con una media de 88%; por
tales motivos se ha tratado de utilizar la prueba usando los antígenos más específicos
como ESAT-6 y CFP10 (Aagaard y col. 2006), lo cual todavía no ha demostrado ser
una estrategia totalmente eficaz para el diagnóstico de M. bovis en bovinos.
Nuestros datos sugieren fuertemente que los antígenos PfkB y HspX, podrían ser
buenos candidatos para utilizarlos en pruebas basadas en la medición de IFN-γ ya sea en
mezcla con el PPD-B o con otros antígenos específicos como ESAT6/CFP10. Koo y
col. (2005) obtuvieron una sensibilidad de 98% y una especificidad de 96% utilizando
ESAT-6 y MPB70. Por otro lado, un coctel peptídico con las proteínas Rv3873,
Rv3879c, Rv0288 y Rv3019c demostró tener una mayor sensibilidad en la respuesta
celular que la dada por ESAT-6/CFP10 o PPD, destacando así su ventaja en la
identificación correcta de animales infectados (Cockle y col 2006). Nuestros datos
concuerdan con estas observaciones, en donde nuestras proteínas antigénicas
demuestran proporcionar una mayor sensibilidad que ESAT-6/CFP10.
En nuestro trabajo se estudiaron los antígenos recombinantes PfkB y HspX, los cuales
lograron detectar un mayor número de animales positivos a la prueba del IFN-γ, sin
embargo es necesario seguir en la búsqueda de nuevas mezclas de antígenos que sirvan
para enriquecer la sensibilidad y especificidad.
Con nuestros datos se sugiere incrementar el punto de corte de la prueba a una
absorbancia de 0.25 mediante el uso de la mezcla de proteínas recombinantes, a fin de
detectar con mayor sensibilidad los casos de animales verdaderamente infectados por
M. bovis, sin embargo, se necesita seguir evaluando si en distintas zonas afectadas por

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
55
la infección animal con diversas prevalencias si nuestros valores diagnósticos, nuestras
consideraciones son extrapolables a las distintas situaciones.
15. CONCLUSIONES
1. Las proteínas PfkB y HspX fueron correctamente producidas en E. coli mediante el uso
de dos vectores de expresión distintos.
2. La antigenicidad de las proteínas recombinantes fue mayor al utilizarse en mezcla con el
antígeno PPD-B que al utilizarse por separado.
3. Se logró detectar un aumento de 14.4% en la sensibilidad en la prueba comercial
mediante el uso de los antígenos recombinantes.
4. La mezcla de proteínas recombinantes PfkB y HspX con PPD-B demostró un valor
mayor de sensibilidad que el obtenido por los antígenos ESAT-6/CFP10.
5. Mediante el uso de la mezcla de proteínas recombinantes PfkB y HspX con PPD-B se
logra detectar un mayor número de animales positivos a la prueba del IFN-γ.
16. REFERENCIAS
AAGAARD C, Govaerts, Meikle M, Gutiérrez-Pabello J, McNair J, Andersen P,
Suárez-Güemes F, Pollock J,Espitia C, Cataldi A (2010) Detection of bovine
tuberculosis in herds with different disease prevalence and influence of
paratuberculosis infection on PPDB and ESAT-6/CFP10 specifity. Preventive
veterinary medicine 96:161-9
AAGAARD C, Govaerts M, Meikle V, Vallecillo AJ, Gutierrez-Pabello JA, Suarez-
Güemez F, McNair J, Cataldi A, Espitia C, Andersen P y Pollock JM (2006)
Optimizing antigen cocktails for detection of Mycobacterium bovis in herds with
different prevalences of bovine tuberculosis ESAT6-CFP10 mixture shows optimal
sensitivity and specificity. J. Clin. Microbiol. 44:4326-4335.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
56
AAGGARD C, Govaerts M, Meng-Okkels L, Andersen P, Pollock JM (2003) Genomic
approach to identification of Mycobacterium bovis diagnostic antigens in cattle.
Journal of clinical microbiology 41: 3719
ABBAS L (2004). Basic Immunology. 2º Ed , Elsevier. EUA.
ABEBE F, Holm-Hansen C, Wiker HG, Bjune G (2007) Progress in serodiagnosis of
Mycobacterium tuberculosis infection. Scandinavian J. of immunology 66:176-91
ALVAREZ A, Estrada-Chávez, Flores-Valdez M (2009), Molecular findings ans
aproaches spotlighting Mycobacterium bovis persistence in cattle, veterinary
research, 40, 3, 22.
AMADORI M, Lyashchenko KP, Gennaro ML, Pollock JM y Zerbini I (2002) Use of
recombinant proteins in antibody tests for bovine tuberculosis. Vet. Microbiol.
85:379-389.
ARANAZ A, De Juan L, Bezos J, Alvarez J, Romero B, Lozano F, Paramio JL, López-
Sánchez J, Mateos A y Domínguez L (2006) Assessment of diagnostic tools for
eradication of bovine tuberculosis in cattle co-infected with Mycobacterium bovis
and M. avium subsp. paratuberculosis. Vet. Res. 37:593-606.
AURTENETXE O, Barral M, Vicente J y otros (2008), Development and validation of
an enzyme-linked immunosorbent assay for antibodies against Mycobacterium
bovis in European wild bord, BMC veterinary research, 85, 4, 379-89.
BARRY C, Corbett D, Bakker D y otros (2011) The effect of Mycobacterium avium
complez infections on routine Mycobacterium bovis diagnostic test, 145092.
BARTEKA L, Rutherfordb R, Gruppoc V, Mortona R, Morrisd R, y col (2010), The
DosR regulon of M. tuberculosis and antibacterial tolerance, Tuberculosis (Edinb);
89(4): 310–316.
BIET F, Boschiroli L, Thorel M, Guilloteau L (2005) Zoonotic aspect of
Mycobacterium bovis and Mycobacterium avium-intracellular complex (MAC);
Vet. Res. 36,411-436.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
57
BLACK G, Thiel B, Ota M y otros (2009), Immunogenicity of novel DosR regulon
encoded candidate antigens of Mycobacterium tuberculosis in three high-burden
populations in africa, Clinical And Vaccine Immunology: CVI, 16, 8, 1203-12.
BORSUK S, newcombe J, Mendum T y otros (2009) Identification of proteins from
tuberculin purified protein derivative (PPD) by LC_-MS/MS, Tuberculosis
(Edinburgh, Scotlnad), 89, 6, 423-30.
BRADFORD MM (1976): A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding. Anal
Biochem. 72: 248-254.
BUDDLE B, Livingstone P, de Lisle G (2009) Advances in ante.mortem diagnosis of
tuberculosi in cattle, New Zeland Veterinary Journal, 57, 4, 173-80.
BUDDLE B, Parlene N; Keen D y otros (1999) Diferrentiation between Mycobacterium
bovis BCG-vaccinated and M. bovis-infected cattle by using recombinant
mycobacterial antigens, Clinical and Diagnostic Laboratory Immunology, 6, 1, 1-5.
BUDDLE B, Wilson T, Denis M y otros (2010) Sensitivity, specificity, and
confounding factors of novel serologcal test used for the rapid serological test used
for the rapid diagnosis of bovine tuberculosis in farmed red deer (cervus elaphus),
Clinical and Vaccine Immunology: CVI, 17, 4, 626-30.
BUDDLE B, M, McCarthy AR, Ryan TJ, Pollock JM, Vordermeier HM, Hewinson
RG, Andersen P, de Lisle GW (2003) Use of mycobacterial peptides and
recombinant proteins for the diagnosis of bovine tuberculosis in skin test-positive
cattle. Vet. Rec. 153:615-620.
CACCAMO N, Barera A, Di Sano C y otros (2003), Cytokine profile, HLA restriction
and PCR sequence analysis of human CD$+T clones specific for an
immunodominant epitope of Mycobacterium tuberculosis 16.kDa protein., Clinical
and Experimental Immunology; 133, 2, 260-6.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
58
CACCAMO N, guggino G, Joosten S y otros (2010) Multifunctional CD4+Y cells
correlate with active Mycobacterium tuberculosis infection, European Journal Of
Immunology; 40, 8, 2211-20.
CASAL C, Bezos J, Diez-Guerrier A, Alvarez J, Romero B, de Juan L, Rodriguez-
Campos S, Vordermeier M, Whelan A, Hewinson RG, Mateos A, Dominguez L,
Aranaz A (2012) Evaluation of two cocktails containing ESAT-6, CFP-10 and Rv-
3615c in the intradermal test and the interferon-γ assay for diagnosis of bovine
tuberculosis. Prev. Vet. Med. 105:149-154.
CFSPH The Center for Food Security & Public Health (2009) Bovine Tuberculosis.
http://www.cfsph.iastate.edu/Factsheets/pdfs/bovine_tuberculosis.pdf
CHAUHAN S, Sharma D, Singh A y otros (2001) Comprehensive insights into
Mycobacterium tuberculosis DevR (DosR) regulon activation switch, Nucleic
acids research; 1-15.
COCKLE P, Gordon S, Hewinson R y otros (2006), Field evaluation of a novel
differential diagnostic reagent for detection of Mycobacterium bovis cattle,
Clinical and Vaccine Immunology, CVI; 13, 10, 1119-24.
COCKLE P, Gordon S, Lavlani A, Buddle BM, Hewinson RG y Vordermeier HM
(2002) Identification of Novel Mycobacteium tuberculosis antigens with potential
as diagnostic reagents of subunit vaccine candidates by comparative genomics.
Infect. Immun. 70: 6996-7003.
COLLINS, D. M. (2011). Advances in molecular diagnostics for Mycobacterium bovis.
Veterinary Microbiology, 151(1-2), 2-7. Elsevier B.V.
doi:10.1016/j.vetmic.2011.02.019
COMMANDEUR S, Lin M, Van Mijgaarden K y otros (2001), Double. And
monofunctional CD$+ ans CD8+ T-cell responses to Mycobacterium tuberculosis
DosR antigens and peptides in long-term latenly infected individuals, European
Journal Of Immunology; 41, 10, 2925-36.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
59
COURTENAY O, Reilly L, Sweeney F y otros (2006), Is Mycobacterium bovis in the
enviroment important for the persistence od tuberculosis?, Biology Letters; 2, 3,
460-2.
DE LA RUA-DOMENECH R, Goodchild AT, Vordermeier HM, Hewinson RG,
Christiansen KH, Clifton-Hadley RS (2006) Ante mortem diagnosis of tuberculosis
in cattle: A review of the tuberculin tests, γ-interferon assay and other ancillary
diagnostic techniques. Res. Vet. Sci. 81:190-210.
DE WARD JH (2005) Tuberculosis bovina. En: Manual de Ganadería Doble Propósito.
González-Stagnaro C y Soto-Belloso E (Eds) pp. 346-369. ISBN 980-6863-00-3.
DEMISSIE A, Leyten E, Abebe m y otros (2006), Recognition of Stage-Specific
Mycobacterial Antigens Differentiates between Acute and Latent Infections with
Mycobacterium tuberculosis; Society; 13,2, 179-186
FLYNN, J. L., & Chan, J. (2001). Immunology of tuberculosis. Annual review of
immunology, 19, 93-129. doi:10.1146/annurev.immunol.19.1.93
GÓMEZ G, Sena I, Bove C, Blanco ML (1986). Tuberculosis y brucelosis en el
Uruguay. Editorial Serie de publicaciones misceláneas. Montevideo, Uruguay.
ISSBN-0534-5391.
GORMLEY E, doyle –m, Fitzsimons T y otros (2006), Diagnosis of Mycobacterium
bovis infection in cattle by use of the gamma-interferon (Bovigam) assay,
Veterinary microbiology.112, 2-4, 171-9.
GREINER M, Pfeiffer D, Smith RD (2000) Principles and practical application of the
receiver-operating characteristic analysis for diagnostic tests. Prev. Vet. Med.
45:23-41
HOPE J, Thom l, Villareal-Ramos b, HEwinson R, Vordermeier H, Howard C (2005),
Exposure to Mycobacterium avium induces low-level protection from
Mycobacterium bovis infection but compromise diagnosis of disease in cattle.
British Society Immunology, Clinical and Experimental Immunology; 141(3)432-
439.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
60
JONES G, Hewinson R, Vordermeier, H (2010) Screening of predicted secreted
antigens from Mycobacterium bovis identifies potential novel differential
diagnostic reagents. Clinical and vaccine immunology : CVI; 17, 9, 1344-8
KOO, H. C., Park, Y. H., Ahn, J., Waters, W. R., Palmer, M. V., Hamilton, M. J.,
Barrington, G., et al. (2005). Use of rMPB70 Protein and ESAT-6 Peptide as
Antigens for Comparison of the Enzyme-Linked Immunosorbent ,
Immunochromatographic , and Latex Bead Agglutination Assays for Serodiagnosis
of Bovine Tuberculosis. Society, 43(9), 4498-4506. doi:10.1128/JCM.43.9.4498
LALVANI, A., & Thillai, M. (2009). Diagnosis of tuberculosis: principles and practice
of using interferon-gamma release assays (IGRAs). Breathe, 5(4), 302-309.
doi:10.1183/18106838.0504.302
LALVANI A, Pareek M (2010) Interefon gamma reléase assays: principle and practice.
Enfermedades infecciosas y microbiología clínica.
http://www.ncbi.nlm.nih.gov/pubmed/19783328
LEYTEN EMS, lin MY, franken JLMC, Friggen AH, Pins C, van Meijgaarden KE,
Voskuil MI, Weldingh K, Anderson P, Schoolnick GK, Arend SM, Ottenhoff
THM, Klein MR (2006) Human T-cell responses to 25 novel antigens encoded by
genes of the dormansy regulon of Mycobacterium tuberculosis. Microb. Infect.
8:2052-2060.
LIEBANA E, Aranaz A, Urquia J, Mateos A y Dominguez L (1998). Evaluation of the
gamma interferon assay for eradication of tuberculosis in a goat herd. Aust. Vet. J.
76, 1, 50- 53.
LIN MY, Reddy TBK, Arend SM, Friggen AH, Franken KLMC, van Meijgaarden KE,
Verduyn MJC, Schoolnick GK, Klein MR y OttenhoffTHM (2009) Cross-reactive
immunity to Mycobacterium tuberculosis DosR regulon-encoded antigens in

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
61
individual infected with environmental, nontubeculous mycobacteria. Infect.
Immun. 77:5071-5079.
LIU, S., Guo, S., Wang, C., Shao, M., Zhang, X., Guo, Y., & Gong, Q. (2007). A novel
fusion protein-based indirect enzyme-linked immunosorbent assay for the
detection of bovine tuberculosis. Tuberculosis (Edinburgh, Scotland), 87(3), 212-
7. doi:10.1016/j.tube.2006.07.007
LOPEZ L, Celma AB, Bravo J, Zepeda H (2007) Tuberculosis bovina: ¿zoonosis
re.emergente? Rev Cien y Tec de Uni Vrz 20
LYASHCHENKO KP, Pollock JM, Colangeli R, Gennaro ML (1998) Diversity of
antigen recognition by serum antibodies in experimental bovine tuberculosis.
Infect. Immun. 66:5344-5349.
LYASHCHENKO K, Whelan O, Greenwald R, Pollock J, Andersen P, Hewinson R,
Vordermeier, H (2004, Association of Tuberculin-Boosted Antibody Responses
with Pathology and Cell-Mediated Immunity in Cattle Vaccinated with
Mycobacterium bovis BCG and Infected with M . bovis, Infection And Immunity;
2462–2467.
MALAGHINI M, Thomaz-Soccol V, Probst C, Krieger M, Preti H, Kritski A, Soccol C
(2011), Recombinant antigen production for assays of intradermoreaction for
diagnosis and surveillance of tuberculosis. Journal of biotechnology; 156, 1, 56-8.
MERCK Y CO INC.(2000) Manual Merck de Veterinaria. 3º ed. Editorial Centrum.
Barcelona, España.
MCAULAY, M., Chance, V., Martin, M., Morgan, J., & Howard, C. J. (2003).
Investigation of the Role of CD8 ϩ T Cells in Bovine Tuberculosis In Vivo.
Society, 71(8), 4297-4303. doi:10.1128/IAI.71.8.4297
MILIAN-SUAZO, F., Harris, B., Dı, C. A., Romero, C., Stuber, T., Alvarez, G.,
Morales, A., et al. (2008). Molecular epidemiology of Mycobacterium bovis :
Usefulness in international trade, 87, 261-271.
doi:10.1016/j.prevetmed.2008.04.004.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
62
MONAGHAN ML, Doherty ML, Collins JD, Kazda JF, Quinn PJ. The tuberculin test.
Vet Microbiol. 1994. 40: 111-124
MOVAHEDI, A. R., & Hampson, D. J. (2008). New ways to identify novel bacterial
antigens for vaccine development. Veterinary Microbiology, 131(1-2), 1-13.
doi:10.1016/j.vetmic.2008.02.011
Norma Oficial Mexicana NOM- 056- Z00- 1995 NORMA Oficial Mexicana NOM-056-
ZOO-1995, Especificaciones técnicas para las pruebas diagnósticas que realicen los
laboratorios de pruebas aprobados en materia zoosanitaria.
PALMER, M. V., Waters, W. R., Thacker, T. C., Greenwald, R., Esfandiari, J.,
Lyashchenko, K. P., (2006). Effects of Different Tuberculin Skin-Testing
Regimens on Gamma Interferon and Antibody Responses in Cattle Experimentally
Infected with Mycobacterium bovis. Society, 13(3), 387-394.
doi:10.1128/CVI.13.3.387
POLLOCK, J. M., Welsh, M. D., & McNair, J. (2005). Immune responses in bovine
tuberculosis: towards new strategies for the diagnosis and control of disease.
Veterinary immunology and immunopathology, 108(1-2), 37-43.
doi:10.1016/j.vetimm.2005.08.012
RANDO, E. H., & Castellanos-Serra, L. (2002), Precipitación selectiva de imidazolato
de zinc; aplicación a la detección de biomoléculas (proteínas, ácidos nucleicos y
glicolípidos bacterianos) separadas en geles de poliacrilamida, Biotecnología
Aplicada; 19:41-47.
RANGEN, S. a, Surujballi, O. P., Lutze-Wallace, C., & Lees, V. W. (2009). Is the
gamma interferon assay in cattle influenced by multiple tuberculin injections? The
Canadian veterinary journal. La revue vétérinaire canadienne, 50(3), 270-4.
Retrieved from
RENSHAW, P. S., Lightbody, K. L., Veverka, V., Muskett, F. W., Kelly, G., Frenkiel,
T. A., Gordon, S. V., et al. (2005). Structure and function of the complex formed

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
63
by the tuberculosis virulence factors CFP-10 and, 24(14), 2491-2498.
doi:10.1038/sj.emboj.7600732
RHODES SG, Gavier-Widen D, Buddle BM, Whelan AO, Singh M, Hewinson RG y
Vordermeier HM (2000) Antigen specificity in experimental bovine tuberculosis.
Infect. Immun. 68:2573-2578.
RANGEN, S. a, Surujballi, O. P., Lutze-Wallace, C., & Lees, V. W. (2009). Is the
gamma interferon assay in cattle influenced by multiple tuberculin injections? The
Canadian veterinary journal. La revue vétérinaire canadienne, 50(3), 270-4.
ROITT I. Brostoff J y Male D (2002). Immunology 10 ed. Editorial Medica
panamericana. México DF.
ROTH, U., Müller, S., & Hanisch, F.-G. (2006). Proteomic analysis of Dictyostelium
discoideum. Methods in molecular biology (Clifton, N.J.), 346, 95-109.
doi:10.1385/1-59745-144-4:95
ROUPIE, V., Romano, M., Zhang, L., Korf, H., Lin, M. Y., Franken, K. L. M. C.,
Ottenhoff, T. H. M., et al. (2007). Immunogenicity of eight dormancy regulon-
encoded proteins of Mycobacterium tuberculosis in DNA-vaccinated and
tuberculosis-infected mice. Infection and immunity, 75(2), 941-9.
doi:10.1128/IAI.01137-06
SCHILLER, I., Vordermeier, H. M., Waters, W. R., Whelan, A. O., Coad, M., Gormley,
E., Buddle, B. M., et al. (2010). Bovine tuberculosis: effect of the tuberculin skin
test on in vitro interferon gamma responses. Veterinary immunology and
immunopathology, 136(1-2), 1-11. Elsevier B.V.
doi:10.1016/j.vetimm.2010.02.007
SCHILLER, I., Waters, W. R., Vordermeier, H. M., Nonnecke, B., Welsh, M., Keck,
N., Whelan, A., et al. (2009). Optimization of a whole-blood gamma interferon
assay for detection of Mycobacterium bovis-infected cattle. Clinical and vaccine
immunology : CVI, 16(8), 1196-202. doi:10.1128/CVI.00150-09

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
64
SIDDERS, B., Pirson, C., Hogarth, P. J., Hewinson, R. G., Stoker, N. G., Vordermeier,
H. M., & Ewer, K. (2008). Screening of highly expressed mycobacterial genes
identifies Rv3615c as a useful differential diagnostic antigen for the
Mycobacterium tuberculosis complex. Infection and immunity, 76(9), 3932-9.
doi:10.1128/IAI.00150-08
SKINNER, M. a, Parlane, N., McCarthy, A., & Buddle, B. M. (2003). Cytotoxic T-cell
responses to Mycobacterium bovis during experimental infection of cattle with
bovine tuberculosis. Immunology, 110(2), 234-41. Retrieved from
SOPP, P., Coad, M., Hewinson, G., Howard, C. J., Cheallaigh, C. N., Keane, J., Harris,
J., et al. (2008). Development of a simple, sensitive, rapid test which discriminates
BCG-vaccinated from Mycobacterium bovis-infected cattle. Vaccine, 26(43),
5470-6. doi:10.1016/j.vaccine.2008.07.059
SOTO R L R, (2007). Test de gamma interferón como prueba confirmatoria para el
diagnostico de la tuberculosis bovna. Universidad Nacional Mayor de San Marcos .
Maestría.
STEMKE-HALE, K., Kaltenboeck, B., DeGraves, F. J., Sykes, K. F., Huang, J., Bu, C.-
hui, & Johnston, S. A. (2005). Screening the whole genome of a pathogen in vivo
for individual protective antigens. Vaccine, 23(23), 3016-25.
doi:10.1016/j.vaccine.2004.12.013
THACKER, T. C., Palmer, M. V., & Waters, W. R. (2007). Associations between
cytokine gene expression and pathology in Mycobacterium bovis infected cattle.
Veterinary immunology and immunopathology, 119(3-4), 204-13.
doi:10.1016/j.vetimm.2007.05.009
TIZARD I. (1995). Inmunología Veterinaria 5º ed. Editorial Medica Panamericana,
México DF.
THOLE, J. E., Dauwerse, H. G., Das, P. K., Groothuis, D. G., Schouls, L. M., & van
Embden, J. D. (1985). Cloning of Mycobacterium bovis BCG DNA and expression
of antigens in Escherichia coli. Infection and immunity, 50(3), 800-6.

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
65
VORDERMEIER HM, Cockle PC, Whelan A, Rhodes S, Palmer N, Bakker D y
Hewinson RG (1999) Development of diagnostic reagents to differentiate between
Mycobacterium bovis BCG vaccination and M. bovis infection in cattle. Clin. Diag.
Lab. Immunol. 6:675-682.
VORDERMEIER HM, Chambers MA, Buddle BM, Pollock JM y Hewinson RG (2006)
Progress in the development of vaccines and diagnostic reagents to control
tuberculosis in cattle. Vet. J. 171:229-244.
VORDERMEIER, H. M., Chambers, M. A., Cockle, P. J., Whelan, A. O., Simmons, J.,
& Hewinson, R. G. (2002). Correlation of ESAT-6-Specific Gamma Interferon
Production with Pathology in Cattle following Mycobacterium bovis BCG
Vaccination against Experimental Bovine Tuberculosis. Peptides, 70(6), 3026-
3032. doi:10.1128/IAI.70.6.3026
VORDERMEIER M, Whelan A, Hewinson G. The scientific case for the gamma
interferon “BOVIGAMTM” assay. Gov Vet J. 2008. 19:38-43
WATERS WR, Palmer MV, Thacker TC, Bannantine JP, Vordermeier HM, Hewinson
RG, Greenwald R, Esfandiari J, McNair J, Pollock JM, Andersen P y Lyashchenko
KP (2006) Early antibody responses to experimental Mycobacterium bovis
infection in cattle. Clin. Vacc. Immunol. 13:648-654.
WATERS, W. R., Whelan, a O., Lyashchenko, K. P., Greenwald, R., Palmer, M. V.,
Harris, B. N., Hewinson, R. G., et al. (2010). Immune responses in cattle
inoculated with Mycobacterium bovis, Mycobacterium tuberculosis, or
Mycobacterium kansasii. Clinical and vaccine immunology : CVI, 17(2), 247-52.
doi:10.1128/CVI.00442-09
WATERS, W. R., Palmer, M. V., Thacker, T. C., Davis, W. C., Sreevatsan, S.,
Coussens, P., Meade, K. G., et al. (2011). Tuberculosis Immunity : Opportunities
from Studies with Cattle. Clinical and Developmental Immunology, 2011.
doi:10.1155/2011/768542
WELSH, M. D., Cunningham, R. T., Corbett, D. M., Girvin, R. M., McNair, J., Skuce,
R. a, Bryson, D. G., et al. (2005). Influence of pathological progression on the

Expresión y purificación de antígenos de Mycobacterium bovis mediante la tecnología del DNA recombinante con uso potencial en diagnóstico
66
balance between cellular and humoral immune responses in bovine tuberculosis.
Immunology, 114(1), 101-11. doi:10.1111/j.1365-2567.2004.02003.
WHELAN AO, Clifford D, Upadhyay B, Breadon EL, McNair J, Hewinson GR y
Vordermeier MH (2010) Development of a skin test for bovine tuberculosis for
differentiating infected from vaccinated animals. J. Clin. Microbiol. 48:3176-3181.
WHELAN, C., Shuralev, E., Kwok, H. F., Kenny, K., Duignan, A., Good, M., Davis,
W. C., et al. (2011). Use of a multiplex enzyme-linked immunosorbent assay to
detect a subpopulation of Mycobacterium bovis-infected animals deemed negative
or inconclusive by the single intradermal comparative tuberculin skin test. Journal
of veterinary diagnostic investigation : official publication of the American
Association of Veterinary Laboratory Diagnosticians, Inc, 23(3), 499-503.
doi:10.1177/1040638711403410
WHELAN, C., Shuralev, E., O’Keeffe, G., Hyland, P., Kwok, H. F., Snoddy, P.,
O’Brien, A., et al. (2008). Multiplex immunoassay for serological diagnosis of
Mycobacterium bovis infection in cattle. Clinical and vaccine immunology : CVI,
15(12), 1834-8. doi:10.1128/CVI.00238-08
WHIPPLE, D. L., Palmer, M. V., Slaughter, R. E., & Jones, S. L. (2001). Comparison
of Purified Protein Derivatives and Effect of Skin Testing on Results of a
Commercial Gamma Interferon Assay for Diagnosis of Tuberculosis in Cattle.
Journal of Veterinary Diagnostic Investigation, 13(2), 117-122.
doi:10.1177/104063870101300204
WOOD PR y Jones L (2001) BOVIGAM: an in vitro cellular diagnostic test for bovine
tuberculosis. Tuberculosis 81:147-155.
WOOD, PR, & Rothel, J. S. (1994). In vitro immunodiagnostic assays for bovine
tuberculosis. Veterinary microbiology, 40(1-2), 125-35. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/8073620.