Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, June 1989, p. 1426-1434 Vol. 55, No. 60099-2240/89/061426-09$02.00/0Copyright © 1989, American Society for Microbiology
Expression by Soil Bacteria of Nodulation Genes fromRhizobium leguminosarum biovar trifolii
B. D. W. JARVIS,* L. J. H. WARD, AND E. A. SLADE
Department of Microbiology and Genetics, Massey University, Palinelston Nor-th, Newv Zealaknd
Received 13 December 1988/Accepted 28 February 1989
Gram-negative, rod-shaped bacteria from the soil of white clover-ryegrass pastures were screened for theirability to nodulate white clover (Trifolium repens) cultivar Grasslands Huia and for DNA homology withgenomic DNA from Rhizobium leguminosarum biovar trifolii ICMP2668 (NZP582). Of these strains, 3.2% wereable to hybridize with strain ICMP2668 and nodulate white clover and approximately 19% hybridized but wereunable to nodulate. Strains which nodulated but did not hybridize with strain ICMP2668 were not detected.DNA from R. leguminosarum biovar trifolii (strain PN165) cured of its symbiotic (Sym) plasmid and a specificnod probe were used to show that the relationship observed was usually due to chromosomal homology.Plasmid pPN1, a cointegrate of the broad-host-range plasmid R68.45 and a symbiotic plasmid pRtr5l4a, wastransferred by conjugation to representative strains of nonnodulating, gram-negative, rod-shaped soil bacteria.Transconjugants which formed nodules were obtained from 6 of 18 (33%) strains whose DNA hybridized withthat of PN165 and 1 of 9 (11%) strains containing DNA which did not hybridize with that of PN165. Thepresence and location of R68.45 and nod genes was confirmed in transconjugants from three of the strainswhich formed nodules. Similarly, a pLAFR1 cosmid containing nod genes from a derivative of R. leguminosa-rum biovar trifolii NZP514 formed nodules when transferred to soil bacteria.
The genus Rhizobiuim is "characteristically able to invadethe root hairs of temperate zone and some tropical zoneleguminous plants (family Leguminosae) and incite the pro-duction of root nodules wherein the bacteria occur asintracellular symbionts. . . which are normally involved infixing atmospheric nitrogen" (25). Thus, nodulation is re-garded as the characteristic function of Rhizobilam speciesand nitrogen fixation as a normal but not essential conse-quence of nodulation. This definition of the genus Rhizobinmis unsatisfactory because it usually depends on the presenceof a Sym plasmid which can be lost in strains maintained onlaboratory media (25). It would be preferable to definerhizobia in terms of DNA-DNA or rRNA-DNA homology toaccepted reference bacteria.
Primary isolation of Rhizobium strains is usually fromnodulated legumes (31, 38, 47, 50, 51). We believe that thishas tended to obscure relationships between rhizobia andother soil bacteria. Strains isolated from nodules on red(Trifoliium pratense) and white (Trifoliium repens) clover areall classified as Rhizobium leguminosariitm biovar trifolii(25), but they exhibit considerable genotypic (6, 21) andphenotypic (51, 52) diversity. Jarvis et al. (21) and Crow etal. (6) used recognized inoculum strains as sources ofreference DNA and found values for optimal DNA homol-ogy and divergence in related sequences consistent with thepresence of subspecies or perhaps multiple species by ac-cepted criteria (2, 23). Young and co-workers (51, 52) usedenzyme polymorphism to show the existence of a number ofgenetically distinct lineages among R. legiuminosarumn bio-vars viceae, trifolii, and phaseoli in soil. They furthershowed that distinct lineages could carry any of severaldifferent host-range determinants. They postulated that soilmight also contain bacteria which were electrophoreticallyindistinguishable from rhizobia but did not contain a Symplasmid and were therefore unrecognizable as rhizobia byconventional methods (51).
* Corresponding author.
Numerous reports have shown that Sym plasmids can betransferred between Rhizobiium strains in the laboratory.Some have involved mobilization by cointegrate formation(27, 40), others self-transmissible Sym plasmids (10, 19, 24,28). It has also been shown that nodule formation but notnitrogen fixation can be expressed on legume roots by Symplasmids in Agrobacteriium species (19, 27, 32), Pseiudomo-nas aeruiginosa, and Ligniobacter species (33).The effects of Sym plasmid transfer to Rhizobiium sp. have
been studied in relation to nodulation specificity (10, 12, 19,24, 28), nitrogen fixation (19, 24, 28), competitiveness (4),and symbiotic efficiency (7, 45). Several aspects of Symplasmid expression in R. legliminosarimn are partly depen-dent on chromosomal factors contributed by the recipientbacteria. These include rhizosphere competitiveness, nodu-lation competitiveness (4), the onset of nitrogen fixation, thecarbon cost of nitrogen fixation, and the host plant biomassresulting from the symbiosis (45).
In view of these considerations, it was decided to examinesoil for the presence of free-living Rhizobilln-related bacte-ria unable to nodulate white clover and to examine theexpression of symbiotic genes from R. leguminosarum bio-var trijolii in these bacteria.
MATERIALS AND METHODSBacterial strains and plasmids. The bacteria and plasmids
used in this study are listed in Table 1.Isolation of soil bacteria. Soil cores 70 to 100 mm long were
collected, placed in plastic bags, and stored at 4°C until theywere cultured. Samples were diluted in 0.05% tryptone brothand immediately placed on yeast-mannitol-glucose (YMG)agar (47) or on soil extract (SE) agar (34). Cultures wereincubated for 5 to 7 days at 28°C. Isolated colonies wererestreaked on YMG or SE agar and reincubated. The ORstrains were gram-negative rod-shaped bacteria from theseplates. The NR strains were gram-negative rod-shaped bac-teria from Rhizobiium sp.-like colonies grown on YMG agar.Cultures were maintained on YMG at 4°C or as cell suspen-
1426
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from

V1od GENE EXPRESSION IN SOIL BACTERIA 1427
TABLE 1. Bacteria and plasmids used in this study
Bacterial strains and plasmids Characteristics" Source or reference
Rlhizobilon legiuminosairimbiovar trifolii
ANU843ICMP2668 (NZP582)
NZP514
NZP561PN100PN 165
Escherichia c oliATCC 9637
HB101RR1PN200PN291PN362PN600PN298
Soil bacteriaNR41
NR64OR168
Spontaneous antibiotic-resistant dervatives
BJ1BJ2BJ3BJ4
Transconjugant bacteriaSJ20 to SJ25SJ1 to SJ6SJ30, SJ32, SJ33SJ40, SJ41SJ43, SJ44
PlasmidspBR328R68.45pRK290pRK2013pRtrSl4a
pRt572
pPN1pLAFR1pPN26
Nod' Fix' on white and subterranean cloversNod' Fix'
Nod+
Nod' Fix'Nod+ Fix+ Str-1 Rif-1Sym plasmid-cured derivative of NZP561
Source of nonrhizobial DNA for colony hybrid-izations and dot blots
pro leiu thi lacY Str-1 recA r mpro lel thi lacY Str-1 r- m-HB101(pPN1)RR1(pRt572)HB101 (pRK2013)HB101(pPN26)HB101(R68.45)
Gram-negative rod from a Rhizobiltin-like colonyon YMG agar
As for NR41Gram-negative rod
NR41Rif`NR64 RifOR168 RifPN165 Rift
Rif KmrRif' KmrRif' KmrRif' KmrRif' Kans
Ap' Tc' Cm'Km" Tc' Cb' IncPDerivative of RK2 TcrKm"Nod' symbiotic plasmid from R. legium7inosaruim
biovar trifolii NZP514R. legiuminosarulm biovar trifolii ANU 843 7.2-kbEcoRl niod fragment cloned in pBR328
pRtrSl4a: :R68.45Acos derivative of pRK290pLAFR1 cosmid containing niod region of PN100
41DSIR culture collection (Palmerston North, New
Zealand) isolated from white clover inoculatedwith ICMP2153 (NZP560)
DSIR culture collection41DSIR culture collectionSpontaneous mutant of NZP514 (41)35
Massey University culture collection (PalmerstonNorth, New Zealand)
4013537, 43D. B. ScottD. B. ScottD. B. Scott
This work
This workThis work
This workThis workThis workThis work
This work (BJ1 xThis work (BJ2 xThis work (BJ3 xThis work (BJ3 xThis work (BJ3 x
PN200)PN200)PN200 nodule 1)PN200 nodule 2)PN200 nodule 2)
1168845
37, 42, 43
411442
" Abbreviations: Ap, ampicillin; Cm, chloramphenicol; Km, kanamycin; Rif. rifampin: Tc, tetracycline.
sions in TY broth (0.5% tryptone, 0.3% yeast extract, 0.13%YCaCl, * 6H,O) containing 50% glycerol at -20°C.
Nodulation tests. Nodulation was tested on sterile whiteclover seedlings (T. repens cv. Grasslands Huia) growing onThornton agar with trace elements (17) in the enclosedculture method described by Vincent (47). In initial screen-ing experiments, two seedlings were inoculated with eachsoil bacterium and seedlings inoculated with R. legiumlinosa-ru(m biovar trifjoii ICMP2668, TAl, or ATCC 10004 andEscherichia coli W were used as positive and negativecontrols, respectively. In transconjugation experiments,three seedlings were used for each transconjugant ortransconjugation mixture and for each donor and recipientstrain. Strain ICMP2668 was used as a positive control. Alltubes were inserted in wooden blocks and grown under
controlled environment conditions at 22°C under artificiallight (180 W/m2) with a 12-h photoperiod. Plants wereexamined for nodule number, shoot length, and color atweekly intervals.
Isolation of bacteria from nodules. For isolation of bacteriafrom nodules, the methods described by Vincent (47) wereused.DNA preparation. Genomic DNA was isolated by a mod-
ification (40) of the method of Fischer and Lerman (13). Soilbacteria were examined for plasmids by a modification (41)of the method of Eckhardt (11) except that a 0.6% agarosegel was used and DNA was separated by electrophoresis (20V for 1 h and then 100 V for 16 h) in a horizontal electro-phoresis apparatus (210 by 160 by 3 mm) (29). Strains of E.coli were not treated with Sarkosyl before lysis. Small
VOL. 55, 1989
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
preparations of plasmids in E. coli were obtained by themethod of Holmes and Quigley (18). Plasmid DNA wasamplified by the method of Clewell (5) when applicable andisolated by the method of Ish-Horowicz and Burke (20). n1odprobes were prepared from pRt572 or pPN26 by digestionwith restriction enzyme EcoRI, separation by agarose gelelectrophoresis, and extraction of the appropriate fragmentby electroelution (30).DNA hybridization procedures. For colony hybridization,
strains were grown on Biodyne A nylon membranes (PallUltrafine Filtration Corp.) on TY agar and incubated for 48 hat 28°C (24 h for E. coli). The original Grunstein and Hogness(15) method was used for lysis, DNA denaturation, andfixation, except that the membrane was incubated in 100 p.gof proteinase K (Boehringer Mannheim Biochemicals, Indi-anapolis, Ind.) per ml for 30 min at 37°C and membraneswere air dried for 30 min and baked in a vacuum oven for 1h at 80°C. Dot blots were obtained by diluting 2 ,ug of DNAsolution (1 mg/ml) in an equal volume of 1.0 M NaOHcontaining 3.0 M NaCl and making three twofold dilutions inthis solution. Each dilution was held at room temperature for5 min and then spotted on a nitrocellulose filter (BA85;Schleicher & Schuell, Inc., Keene, N.H.) on a pad ofWhatman 3MM paper soaked in 0.5 M Tris-hydrochloridebuffer (pH 7.5) containing 1.5 M NaCl and left for at least 5min. The filter was placed on a filter holder, washed twicewith 90% ethanol (25 ml) under vacuum, air dried about 20min, and baked at 80°C for 2 h in a vacuum oven. PlasmidDNA was transferred from Eckhardt gels to nitrocellulose(BA85) by the method of Southern (46). Membranes wereexposed to 3 x 106 to 4 x 101 cpm of 32P-labeled probe DNAprepared by the random primer method (49) and incubated at55°C (dot blots) or 65°C (colony hybridizations and Southerntransfers) for 24 h. After hybridization, colony filters werewashed three times with agitation for 5 min each time in 2xSSC (1x SSC is 0.15 M NaCI plus 0.015 M sodium citrate)containing 0.1% (wt/vol) sodium dodecyl sulfate at roomtemperature and two times for 15 min each time in 0.1xSSC-0.1% sodium dodecyl sulfate at 50°C. Dot blots andSouthern transfers were washed with agitation twice in 2xSSC for 15 min at room temperature, once in 0.1x SSC for15 min at room temperature, and once in 0.1x SSC for 15min at 42°C (dot blots) or 50°C (Southern transfers). Allmembranes were exposed to X-ray film (Agfa-Gevaert,Curix RP2) in the presence of an intensifying screen (CronexLightning-Plus) at -70°C for periods of up to 6 days anddeveloped in a Kodak automatic film processor.
RESULTS
Isolation and preliminary screening of soil bacteria. Strainsof gram-negative soil bacteria were obtained from 18 soilcores taken from ryegrass-white clover permanent pastureson river flats consisting of recent alluvial soil and twoneighboring terrace soils which consisted of yellow-greyearths derived from loess and alluvial material. These pas-tures had no recent history of Rhizobiumn inoculation butwere probably inoculated when they were sown down.Cultures on YMG agar or SE agar were incubated for 5 to 7days at 28°C, and colonies were purified by streaking. TheOR series comprised 200 strains of gram-negative rod-shaped bacteria with various colony morphologies, and theNR series comprised 46 strains of gram-negative rod-shapedbacteria whose colony morphology on YMG agar was simi-lar to that of Rhizobiium sp. Both series were tested for theirability to nodulate and fix atmospheric nitrogen on white
A
Idt0
14
24~
FIG. 1. Autoradiograms of DNA from colonies of soil bacteria(NR series) grown on Biodyne membranes and hybridized with32P-labeled genomic DNA from R. leguminiosarum biovar trifoliiICMP2668. Exposure was for 21 h at -70°C. Hybridizing strains inpanel A (NR9, NR-12, NR-16, and NR-19) nodulated white clover,but NR64 in panel B did not.
clover. Three (1.5%) OR strains and 5 (10.9%) NR strainsformed effective (nitrogen-fixing) nodules. The remainderwere unable to nodulate white clover.Colony hybridization was used to screen bacteria from the
OR and NR series for ability to hybridize with genomic DNAfrom R. legltninosari-in biovar trifolii ICMP2668 or PN165(Fig. 1). When cells were lysed on the nitrocellulose mem-brane, cellular material was released which bound 32p_labeled probe DNA. The method described above removesthis debris and minimizes nonspecific binding. A positivecontrol (ICMP2668) and a negative control (E. coli) wereincluded on each membrane, and autoradiograms were ex-posed so that hybridization with E. coli was negligible. In theOR series, 38 of 188 (20.2%) strains hybridized with probeDNA. Quantitative estimates of relatedness are not avail-able, but hybridization with strains previously shown tonodulate was stronger than that with nonnodulating strains(Fig. 1). All strains showing detectable homology wereincluded in the above estimate. No strains which nodulatedbut did not hybridize with the probe DNA were detected. Inthe NR series, 6 of 46 strains hybridized with the probeDNA. Five of these had nodulated white clover but one(NR64) had not. Based on the OR data, it is suggested that20% of the culturable gram-negative, nonsporeforming, rod-shaped bacteria in these soils had a detectable geneticrelationship with R. legiuminosarium biovar trifolii but only1.5% were capable of nodulation.Genomic homology between R. leguminosarum biovar trifo-
lii and other soil bacteria. The proportion of soil bacteriawhich did not nodulate but showed a relationship with R.legiiininosaruin biovar trifolii was unexpectedly high. Toconfirm homologies observed and examine the nature of thehybridizing DNA, we prepared dot blots from DNA ex-
1428 JARVIS ET AL.
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from
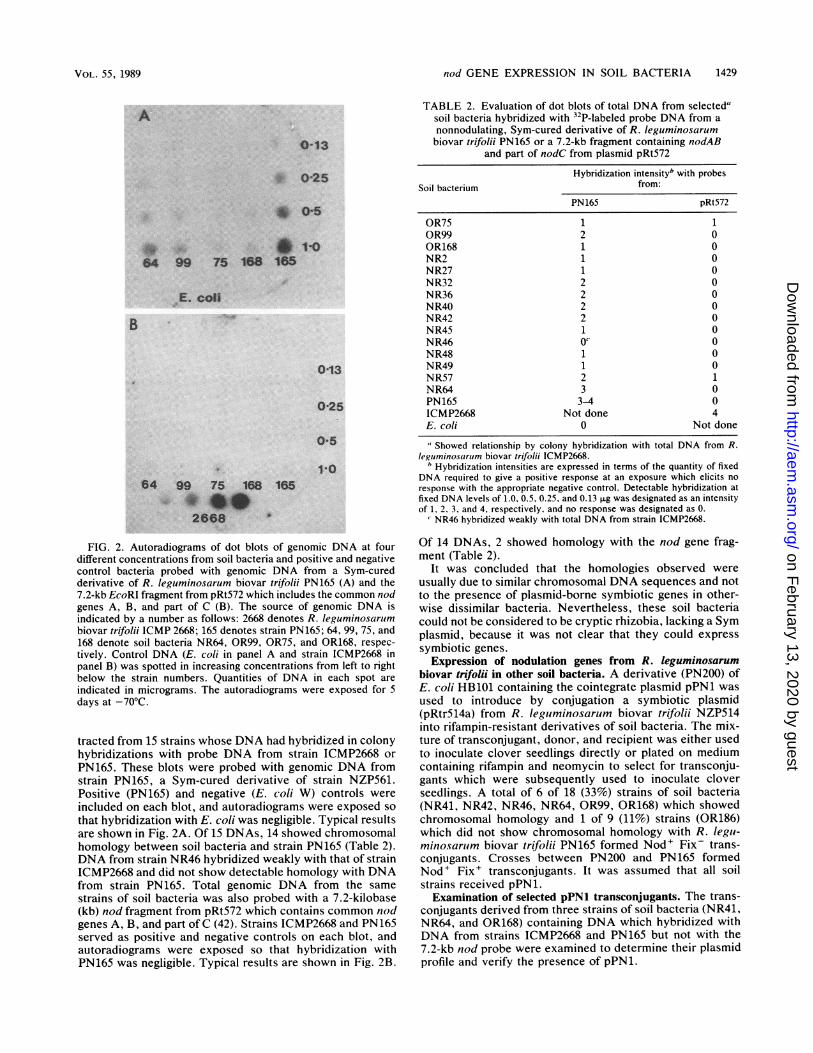
nod GENE EXPRESSION IN SOIL BACTERIA 1429
FIG. 2. Autoradiograms of dot blots of genomic DNA at fourdifferent concentrations from soil bacteria and positive and negativecontrol bacteria probed with genomic DNA from a Sym-curedderivative of R. leguminosarum biovar trifolii PN165 (A) and the7.2-kb EcoRI fragment from pRt572 which includes the common nodgenes A, B, and part of C (B). The source of genomic DNA isindicated by a number as follows: 2668 denotes R. leguminosarumbiovar trifolii ICMP 2668; 165 denotes strain PN165; 64, 99, 75, and168 denote soil bacteria NR64, OR99, OR75, and OR168, respec-
tively. Control DNA (E. coli in panel A and strain ICMP2668 inpanel B) was spotted in increasing concentrations from left to rightbelow the strain numbers. Quantities of DNA in each spot are
indicated in micrograms. The autoradiograms were exposed for 5days at -70°C.
tracted from 15 strains whose DNA had hybridized in colonyhybridizations with probe DNA from strain ICMP2668 or
PN165. These blots were probed with genomic DNA fromstrain PN165, a Sym-cured derivative of strain NZP561.Positive (PN165) and negative (E. coli W) controls were
included on each blot, and autoradiograms were exposed so
that hybridization with E. coli was negligible. Typical resultsare shown in Fig. 2A. Of 15 DNAs, 14 showed chromosomalhomology between soil bacteria and strain PN165 (Table 2).DNA from strain NR46 hybridized weakly with that of strainICMP2668 and did not show detectable homology with DNAfrom strain PN165. Total genomic DNA from the same
strains of soil bacteria was also probed with a 7.2-kilobase(kb) nod fragment from pRt572 which contains common nodgenes A, B, and part of C (42). Strains ICMP2668 and PN165served as positive and negative controls on each blot, andautoradiograms were exposed so that hybridization withPN165 was negligible. Typical results are shown in Fig. 2B.
TABLE 2. Evaluation of dot blots of total DNA from selected"soil bacteria hybridized with 32P-labeled probe DNA from anonnodulating, Sym-cured derivative of R. leguminosarumbiovar trifolii PN165 or a 7.2-kb fragment containing nodAB
and part of nodC from plasmid pRt572
Hybridization intensityb with probesSoil bacterium from:
PN165 pRt572
OR75 1 1OR99 2 0OR168 1 0NR2 1 0NR27 1 0NR32 2 0NR36 2 0NR40 2 0NR42 2 0NR45 1 0NR46 0o 0NR48 1 0NR49 1 0NR57 2 1NR64 3 0PN165 3-4 0ICMP2668 Not done 4E. coli 0 Not done
" Showed relationship by colony hybridization with total DNA from R.leguminosarum biovar trifolii ICMP2668.b Hybridization intensities are expressed in terms of the quantity of fixedDNA required to give a positive response at an exposure which elicits noresponse with the appropriate negative control. Detectable hybridization atfixed DNA levels of 1.0, 0.5, 0.25, and 0.13 p.g was designated as an intensityof 1. 2. 3, and 4, respectively, and no response was designated as 0.
' NR46 hybridized weakly with total DNA from strain ICMP2668.
Of 14 DNAs, 2 showed homology with the nod gene frag-ment (Table 2).
It was concluded that the homologies observed wereusually due to similar chromosomal DNA sequences and notto the presence of plasmid-borne symbiotic genes in other-wise dissimilar bacteria. Nevertheless, these soil bacteriacould not be considered to be cryptic rhizobia, lacking a Symplasmid, because it was not clear that they could expresssymbiotic genes.
Expression of nodulation genes from R. leguminosarumbiovar trifolii in other soil bacteria. A derivative (PN200) ofE. coli HB101 containing the cointegrate plasmid pPN1 wasused to introduce by conjugation a symbiotic plasmid(pRtrSl4a) from R. legiuminosarum biovar trifolii NZP514into rifampin-resistant derivatives of soil bacteria. The mix-ture of transconjugant, donor, and recipient was either usedto inoculate clover seedlings directly or plated on mediumcontaining rifampin and neomycin to select for transconju-gants which were subsequently used to inoculate cloverseedlings. A total of 6 of 18 (33%) strains of soil bacteria(NR41, NR42, NR46, NR64, OR99, OR168) which showedchromosomal homology and 1 of 9 (11%) strains (OR186)which did not show chromosomal homology with R. legiu-minosarum biovar trifolii PN165 formed Nod' Fix- trans-conjugants. Crosses between PN200 and PN165 formedNod' Fix' transconjugants. It was assumed that all soilstrains received pPN1.
Examination of selected pPN1 transconjugants. The trans-conjugants derived from three strains of soil bacteria (NR41,NR64, and OR168) containing DNA which hybridized withDNA from strains ICMP2668 and PN165 but not with the7.2-kb nod probe were examined to determine their plasmidprofile and verify the presence of pPN1.
VOL. 55, 1989
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
8 * 2 3 4 5 6 I a ".
:ii
Cl2 3 4 5 6 7 S -J
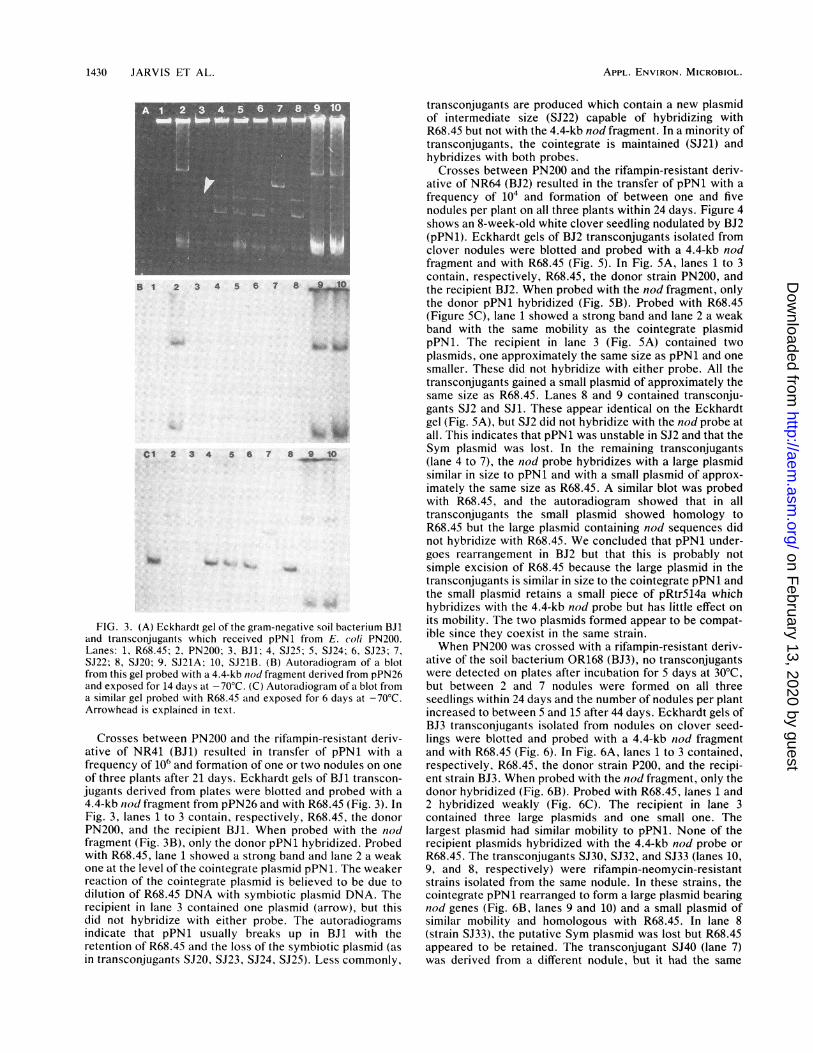
FIG. 3. (A) Eckhardt gel of the gram-negative soil bacterium BJland transconjugants which received pPNI from E. (0/i PN200.
Lanes: 1, R68.45; 2, PN200; 3, BJ1; 4, SJ25; 5, SJ24; 6, S123: 7,
SJ22; 8, SJ20; 9. SJ21A; 10, SJ21B. (B) Autoradiogram of a blot
from this gel probed with a 4.4-kb niod fragment derived from pPN26
and exposed for 14 days at -70'C. (C) Autoradiogram of a blot from
a similar gel probed with R68.45 and exposed for 6 days at -70'C.Arrowhead is explained in text.
Crosses between PN200 and the rifampin-resistant deriv-
ative of NR41 (BJI) resulted in transfer of pPN1 with a
frequency of 106 and formation of one or two nodules on one
of three plants after 21 days. Eckhardt gels of BJ1 transcon-
jugants derived from plates were blotted and probed with a
4.4-kb niod fragment from pPN26 and with R68.45 (Fig. 3). In
Fig. 3, lanes 1 to 3 contain, respectively, R68.45, the donor
PN200, and the recipient BJI. When probed with the nod
fragment (Fig. 313), only the donor pPN1I hybridized. Probed
with R68.45, lane showed a strong band and lane 2 a weak
one at the level of the cointegrate plasmid pPN1I. The weaker
reaction of the cointegrate plasmid is believed to be due to
dilution of R68.45 DNA with symbiotic plasmid DNA. The
recipient in lane 3 contained one plasmid (arrow), but this
did not hybridize with either probe. The autoradiogramsindicate that pPN1 usually breaks up in BJI with the
retention of R68.45 and the loss of the symbiotic plasmid (as
in transconjugants SJ20, SJ23, SJ24, SJ25). Less commonly,
transconjugants are produced which contain a new plasmidof intermediate size (SJ22) capable of hybridizing withR68.45 but not with the 4.4-kb nod fragment. In a minority oftransconjugants, the cointegrate is maintained (SJ21) andhybridizes with both probes.
Crosses between PN200 and the rifampin-resistant deriv-ative of NR64 (BJ2) resulted in the transfer of pPN1 with afrequency of 104 and formation of between one and fivenodules per plant on all three plants within 24 days. Figure 4shows an 8-week-old white clover seedling nodulated by BJ2(pPN1). Eckhardt gels of BJ2 transconjugants isolated fromclover nodules were blotted and probed with a 4.4-kb nodfragment and with R68.45 (Fig. 5). In Fig. 5A, lanes 1 to 3contain, respectively, R68.45, the donor strain PN200, andthe recipient BJ2. When probed with the nod fragment, onlythe donor pPN1 hybridized (Fig. 5B). Probed with R68.45(Figure SC), lane 1 showed a strong band and lane 2 a weakband with the same mobility as the cointegrate plasmidpPN1. The recipient in lane 3 (Fig. 5A) contained twoplasmids, one approximately the same size as pPN1 and onesmaller. These did not hybridize with either probe. All thetransconjugants gained a small plasmid of approximately thesame size as R68.45. Lanes 8 and 9 contained transconju-gants SJ2 and SJ1. These appear identical on the Eckhardtgel (Fig. SA), but SJ2 did not hybridize with the nod probe atall. This indicates that pPN1 was unstable in SJ2 and that theSym plasmid was lost. In the remaining transconjugants(lane 4 to 7), the nod probe hybridizes with a large plasmidsimilar in size to pPN1 and with a small plasmid of approx-imately the same size as R68.45. A similar blot was probedwith R68.45, and the autoradiogram showed that in alltransconjugants the small plasmid showed homology toR68.45 but the large plasmid containing nod sequences didnot hybridize with R68.45. We concluded that pPN1 under-goes rearrangement in BJ2 but that this is probably notsimple excision of R68.45 because the large plasmid in thetransconjugants is similar in size to the cointegrate pPN1 andthe small plasmid retains a small piece of pRtrS14a whichhybridizes with the 4.4-kb nod probe but has little effect onits mobility. The two plasmids formed appear to be compat-ible since they coexist in the same strain.When PN200 was crossed with a rifampin-resistant deriv-
ative of the soil bacterium OR168 (BJ3), no transconjugantswere detected on plates after incubation for 5 days at 30°C,but between 2 and 7 nodules were formed on all threeseedlings within 24 days and the number of nodules per plantincreased to between 5 and 15 after 44 days. Eckhardt gels ofBJ3 transconjugants isolated from nodules on clover seed-lings were blotted and probed with a 4.4-kb nod fragmentand with R68.45 (Fig. 6). In Fig. 6A, lanes 1 to 3 contained,respectively, R68.45, the donor strain P200, and the recipi-ent strain BJ3. When probed with the nod fragment, only thedonor hybridized (Fig. 6B). Probed with R68.45, lanes 1 and2 hybridized weakly (Fig. 6C). The recipient in lane 3contained three large plasmids and one small one. Thelargest plasmid had similar mobility to pPN1. None of therecipient plasmids hybridized with the 4.4-kb nod probe orR68.45. The transconjugants SJ30, SJ32, and SJ33 (lanes 10,9, and 8, respectively) were rifampin-neomycin-resistantstrains isolated from the same nodule. In these strains, thecointegrate pPN1 rearranged to form a large plasmid bearingnod genes (Fig. 6B, lanes 9 and 10) and a small plasmid ofsimilar mobility and homologous with R68.45. In lane 8(strain SJ33), the putative Sym plasmid was lost but R68.45appeared to be retained. The transconjugant SJ40 (lane 7)was derived from a different nodule, but it had the same
1430 JARVIS ET AL.
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from
nod GENE EXPRESSION IN SOIL BACTERIA 1431
ml 21111I 11
C,, 1oft
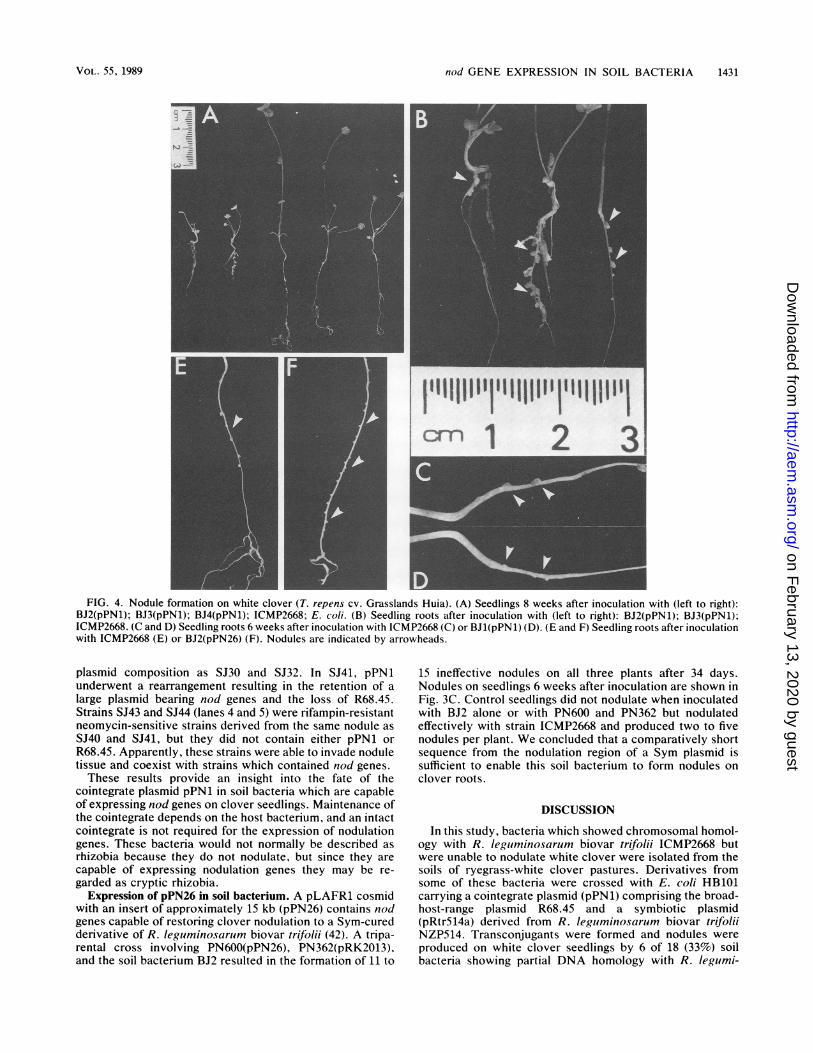
FIG. 4. Nodule formation on white clover (T. repens cv. Grasslands Huia). (A) Seedlings 8 weeks after inoculation with (left to right):BJ2(pPN1); BJ3(pPN1); BJ4(pPN1); ICMP2668; E. coli. (B) Seedling roots after inoculation with (left to right): BJ2(pPN1); BJ3(pPN1);ICMP2668. (C and D) Seedling roots 6 weeks after inoculation with ICMP2668 (C) or BJ1(pPN1) (D). (E and F) Seedling roots after inoculationwith ICMP2668 (E) or BJ2(pPN26) (F). Nodules are indicated by arrowheads.
plasmid composition as SJ30 and SJ32. In SJ41, pPN1underwent a rearrangement resulting in the retention of alarge plasmid bearing nod genes and the loss of R68.45.Strains SJ43 and SJ44 (lanes 4 and 5) were rifampin-resistantneomycin-sensitive strains derived from the same nodule asSJ40 and SJ41, but they did not contain either pPN1 orR68.45. Apparently, these strains were able to invade noduletissue and coexist with strains which contained nod genes.These results provide an insight into the fate of the
cointegrate plasmid pPN1 in soil bacteria which are capableof expressing nod genes on clover seedlings. Maintenance ofthe cointegrate depends on the host bacterium, and an intactcointegrate is not required for the expression of nodulationgenes. These bacteria would not normally be described asrhizobia because they do not nodulate, but since they arecapable of expressing nodulation genes they may be re-garded as cryptic rhizobia.
Expression of pPN26 in soil bacterium. A pLAFR1 cosmidwith an insert of approximately 15 kb (pPN26) contains nodgenes capable of restoring clover nodulation to a Sym-curedderivative of R. leguminosarilm biovar trifolii (42). A tripa-rental cross involving PN600(pPN26), PN362(pRK2013),and the soil bacterium BJ2 resulted in the formation of 11 to
15 ineffective nodules on all three plants after 34 days.Nodules on seedlings 6 weeks after inoculation are shown inFig. 3C. Control seedlings did not nodulate when inoculatedwith BJ2 alone or with PN600 and PN362 but nodulatedeffectively with strain ICMP2668 and produced two to fivenodules per plant. We concluded that a comparatively shortsequence from the nodulation region of a Sym plasmid issufficient to enable this soil bacterium to form nodules onclover roots.
DISCUSSION
In this study, bacteria which showed chromosomal homol-ogy with R. legunzinosarum biovar trifolii ICMP2668 butwere unable to nodulate white clover were isolated from thesoils of ryegrass-white clover pastures. Derivatives fromsome of these bacteria were crossed with E. coli HB101carrying a cointegrate plasmid (pPN1) comprising the broad-host-range plasmid R68.45 and a symbiotic plasmid(pRtrS14a) derived from R. legurninosarum biovar trifoliiNZP514. Transconjugants were formed and nodules wereproduced on white clover seedlings by 6 of 18 (33%) soilbacteria showing partial DNA homology with R. legumi-
VOL. 55, 1989
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
_~~~~~~~~~~~~~~~~~
5 1 2 3 4 5 6 7 8 9 10
A* _00 g: *j
ci~~~~ ~ ~~~~~~~~~24.ai
C1 2 3 4 5 6 7 8 9 10
bg
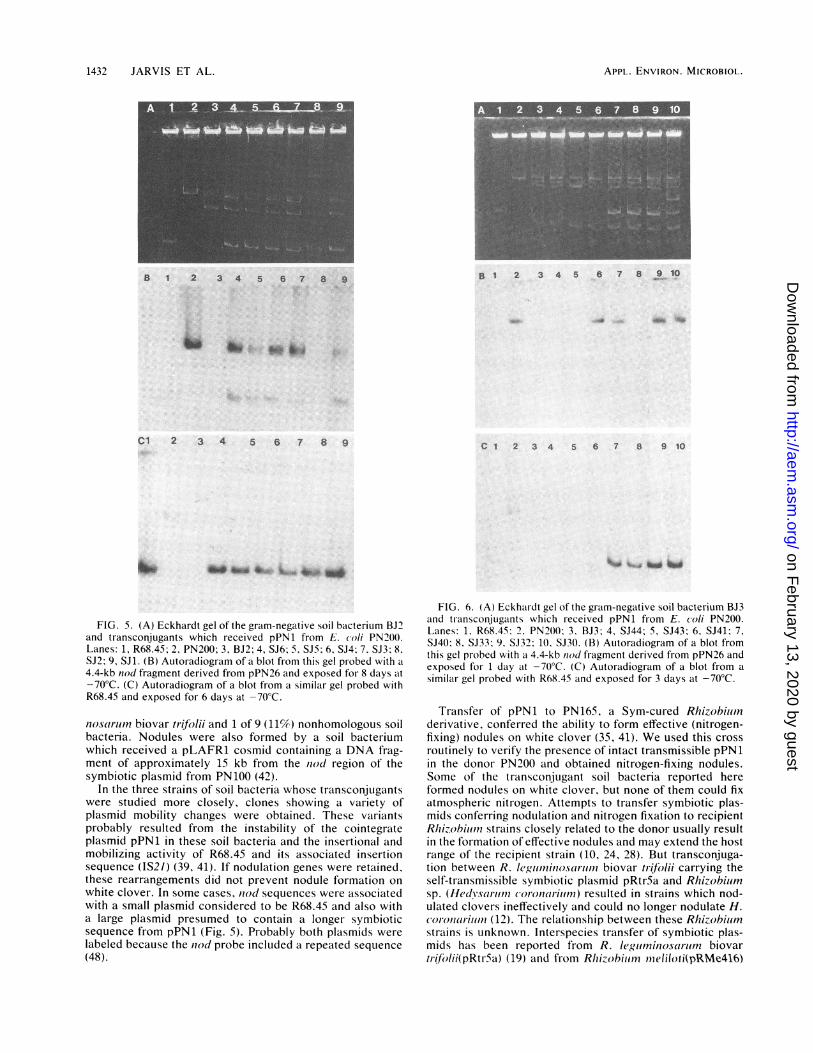
FIG. 5. (A) Eckhardt gel of the gram-negative soil bacterium BJ'and transconjugants which received pPN1 from E. coli PN')0.Lanes: 1, R68.45; 2, PN200; 3, BJ2; 4, SJ6; 5, SJ5; 6, SJ4; 7. SJ3; 8.SJ2; 9, SJ1. (B) Autoradiogram of a blot from this gel probed with a
4.4-kb nod fragment derived from pPN26 and exposed for 8 days at-70°C. (C) Autoradiogram of a blot from a similar gel probed withR68.45 and exposed for 6 days at -70°C.
nosaruin biovar t-ifolii and 1 of 9 (11%) nonhomologous soilbacteria. Nodules were also formed by a soil bacteriumwhich received a pLAFRI cosmid containing a DNA frag-ment of approximately 15 kb from the notd region of thesymbiotic plasmid from PN100 (42).
In the three strains of soil bacteria whose transconjugantswere studied more closely, clones showing a variety ofplasmid mobility changes were obtained. These variantsprobably resulted from the instability of the cointegrateplasmid pPN1 in these soil bacteria and the insertional andmobilizing activity of R68.45 and its associated insertionsequence (1S21) (39, 41). If nodulation genes were retained,these rearrangements did not prevent nodule formation on
white clover. In some cases, niod sequences were associatedwith a small plasmid considered to be R68.45 and also witha large plasmid presumed to contain a longer symbioticsequence from pPN1 (Fig. 5). Probably both plasmids were
labeled because the niod probe included a repeated sequence
(48).
FIG. 6. (A) Eckhardt gel of the gram-negative soil bacterium BJ3and transconjugants which received pPN1 from E. (oli PN200.Lanes: 1. R68.45: 2. PN200() 3. BJ3; 4, SJ44; 5, SJ43; 6, SJ41; 7,SJ40); 8, SJ33; 9. SJ32; 10, SJ30. (B) Autoradiogram of a blot fromthis gel probed with a 4.4-kb ntod fragment derived from pPN26 andexposed for 1 day aIt -70°C. (C) Autoradiogram of a blot from asimilar gel probed with R68.45 and exposed for 3 days at -70°C.
Transfer of pPN1 to PN165, a Sym-cured Rhizobiiinderivative, conferred the ability to form effective (nitrogen-fixing) nodules on white clover (35, 41). We used this crossroutinely to verify the presence of intact transmissible pPN1in the donor PN200 and obtained nitrogen-fixing nodules.Some of the transconjugant soil bacteria reported hereformed nodules on white clover, but none of them could fixatmospheric nitrogen. Attempts to transfer symbiotic plas-mids conferring nodulation and nitrogen fixation to recipientRhiz-obijino1 strains closely related to the donor usually resultin the formation of effective nodules and may extend the hostrange of the recipient strain (10, 24, 28). But transconjuga-tion between R. legitininosalrumi91 biovar t'if'olii carrying theself-transmissible symbiotic plasmid pRtr5a and Rhizobiumsp. (Hedvlysarun (cor-onai-iuin) resulted in strains which nod-ulated clovers ineffectively and could no longer nodulate H.corionaiIlm (12). The relationship between these Rhizobiiumstrains is unknown. Interspecies transfer of symbiotic plas-mids has been reported from R. legiumnincosaruirin biovart/ijolii(pRtr5a) (19) and from Rhizobiurn mne1i(oti(pRMe416)
404004*40
1432 JARVIS ET AL.
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from

nod GENE EXPRESSION IN SOIL BACTERIA 1433
(27) to Agrobacterium tumefaciens. These species are asclosely related as some rhizobia (22), and there is evidencefor a close ancestral relationship between the plasmids ofRhizobium and Agrobacterium species based on plasmidincompatibility studies (32). Nevertheless, the nodulesformed by Agrobacterium transconjugants were ineffective.Similarly, the cointegrate plasmid pPN1 was used to transfera Sym plasmid from R. leguminosarum biovar trifolii(pRtrSl4a) to Lignobacter strain K17 and P. aeruginosaPAO5 by conjugation, and ineffective nodules were formed(33). It is concluded that symbiotic plasmids from Rhizobiumspecies may be expressed by other bacteria but that expres-sion will most probably be confined to the formation ofineffective nodules on legume roots.
Skot et al. (45) studied the efficiency of nitrogenase-linkednodule respiration when three different symbiotic plasmidswere expressed in three different strains of R. leguminosa-rum and found that the bacterial genomic background had alarger effect than the Sym plasmids tested on the carboncosts of N2 fixation, host plant biomass, and the time ofonset of nitrogenase activity. These results suggest thatspecific determinants in the bacterial genome are requiredfor the efficient expression of plasmid-borne symbioticgenes. Random conjugation with susceptible soil bacteria isrelatively unlikely to achieve the required combination ofgenetic determinants, so that even though significant num-bers of transconjugant bacteria can be generated in soilmicrocosms within a day of inoculation with a donor popu-lation (26), genetic exchange which results in the formationof nitrogen-fixing transconjugants may be a relatively rareevent. A more probable outcome is the formation of apopulation of ineffective or poorly effective strains whichmay or may not be capable of competing with the inoculumstrain in the formation of nodules on the host plant.The distinguishing characteristic of Rhizobium and
Bradyrhizobium strains is their ability to form nodules on theroots of legumes (25), but it is known that this character isfrequently carried on a plasmid and several self-transmis-sible symbiotic plasmids have been reported (10, 24, 28).These plasmids have been expressed in Agrobacteriumstrains (19), and a symbiotic cointegrate plasmid was mobi-lized and expressed in Pseudomonas and Lignobac terstrains (33). Agrobacterium plasmids have been expressed inRhizobium species (33). We found that approximately 20%of the gram-negative, nonsporeforming, rod-shaped bacteria(OR series) isolated from three pasture soils contained DNAwhich gave detectable hybridization with R. leguminosariumbiovar trifolii and that 1.5% were able to nodulate whiteclover. Eighteen of the strains which hybridized but did notnodulate were crossed with PN200, and six strains gainedthe ability to form nodules. This suggested that aboutone-third of the 18.5% of strains which hybridized but didnot nodulate were capable of expressing nodulation genes.We concluded that the ability to express nodulation genes isnot sufficient, in itself, to define a biological entity whichshould be recognized by a taxonomic name, such as Rhizo-bium. Nodulation may prove to be a characteristic of strainsfrom several bacterial genera (19, 22, 33). If this is the case,it should be recognized and reflected in the nomenclature.From a practical viewpoint, it would be convenient anduseful to redefine rhizobia as those bacteria capable ofexpressing symbiotic nitrogen fixation, but many steps areinvolved in the formation of an effective, nitrogen-fixingnodule (9). Malfunctions in host or bacterial metabolism caninterfere with the process. Thus, reliance on the formation ofnitrogen-fixing nodules as the criterion would probably ex-
clude many root nodule bacteria and so hinder understand-ing of the biotic interactions among microorganisms in soil.These problems are avoided by defining bacterial genera interms of chromosomal homology with specific type strainsand several chromosomally borne phenotypic characters, ashas been done for Legionella (3, 44), or by using ribosomalcistron homologies (reviewed by Schleifer and Stackebrandt[36]).
ACKNOWLEDGMENTS
We thank Barry Scott for the provision of strains, Robert Cleaverfor technical advice, both Barry Scott and Brian Mansfield forconstructive criticism of the work, and Eric Terzaghi for criticalreading of the manuscript.
This work was supported by the Massey University ResearchFund.
LITERATURE CITED1. Bolivar, F., R. Rodriguez, P. J. Green, M. Betlach, H. L.
Heyneker, H. W. Boyer, J. Crosa, and S. Falkow. 1977. Con-struction and characterisation of new cloning vehicles. II. Amultipurpose cloning system. Gene 2:95-113.
2. Brenner, D. J., A. C. McWhorter, J. K. L. Knutson, and A. G.Steigerwalt. 1982. Escherichia vulneris: a new species of Entero-bacteriaceae associated with human wounds. J. Clin. Micro-biol. 15:1133-1140.
3. Brenner, D. J., A. G. Steigerwalt, G. W. Gorman, H. W.Wilkinson, W. F. Bibb, M. Hackel, R. L. Tyndall, J. Campbell,J. C. Feeley, W. L. Thacker, P. Skaliy, W. T. Martin, B. J.Brake, B. S. Fields, H. V. McEachern, and L. K. Corcoran.1985. Ten new species of Legionella. Int. J. Syst. Bacteriol.35:50-59.
4. Brewin, N. J., E. A. Wood, and J. P. W. Young. 1983. Contri-bution of the symbiotic plasmid to the competitiveness ofRhizobium leguminosarum. J. Gen. Microbiol. 129:2973-2977.
5. Clewell, D. B. 1972. Nature of ColEl plasmid replication inEscherichia coli in the presence of chloramphenicol. J. Bacte-riol. 110:667-676.
6. Crow, V. L., B. D. W. Jarvis, and R. M. Greenwood. 1981.Deoxyribonucleic acid homologies among acid-producingstrains of Rhizobium. Int. J. Syst. Bacteriol. 31:152-172.
7. De Jong, T. M., N. J. Brewin, A. W. B. Johnston, and D. A.Phillips. 1982. Improvement of symbiotic properties in Rhizo-biumn leguminosarum by plasmid transfer. J. Gen. Microbiol.128:1829-1838.
8. Ditta, G., S. Stanfield, D. Corbin, and D. R. Helsinki. 1980.Broad host range DNA cloning system for gram-negative bac-teria: construction of a gene bank of Rhizobiumn ineliloti. Proc.Natl. Acad. Sci. USA 77:7347-7351.
9. Djordjevic, M. A., D. W. Gabriel, and B. G. Rolfe. 1987.Rliizobium-the refined parasite of legumes. Annu. Rev. Phy-topathol. 25:145-168.
10. Djordjevic, M. A., W. Zurkowski, J. Shine, and B. G. Rolfe.1983. Sym plasmid transfer to various symbiotic mutants ofRhizobium trifolii. R. leguiminosarum, and R. meliloti. J. Bac-teriol. 156:1035-1045.
11. Eckhardt, T. 1978. A rapid method for the identification ofplasmid deoxyribonucleic acid in bacteria. Plasmid 1:584-588.
12. Espuny, M. R., F. J. Ollero, R. A. Bellogin, J. E. Ruiz-Sainz,and J. Perez-Silva. 1987. Transfer of the Rhizobiutn leguminosa-rum biovar trifolii symbiotic plasmid pRtr5a to a strain ofRhizobium sp. that nodulates on Hedysaruin coronarium. J.Appl. Bacteriol. 63:13-20.
13. Fischer, S. G., and L. S. Lerman. 1979. Length-independentseparation of DNA restriction fragments in two dimensional gelelectrophoresis. Cell 16:191-200.
14. Friedman, A. M., S. R. Long, S. E. Brown, W. J. Buikema, andF. M. Ausubel. 1982. Construction of a broad host range cosmidcloning vector and its use in the genetic analysis of Rhizobiumnmutants. Gene 18:289-296.
15. Grunstein, M., and D. Hogness. 1975. Colony hybridisation: a
VOL. 55, 1989
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
method for the isolation of cloned DNAs that contain a specificgene. Proc. Natl. Acad. Sci. USA 72:3961-3965.
16. Haas, D., and B. W. Holloway. 1976. R factor variants withenhanced sex factor activity in Pseuidot1ontias (riegincg>osa. Mol.Gen. Genet. 144:243-251.
17. Hoagland, D. R., and D. T. Arnon. 1938. The water culturemethod for growing plants without soil. University of CaliforniaAgriculture Experiment Station Circulation no. 347.
18. Holmes, D. S., and M. Quigley. 1981. A rapid boiling method forthe preparation of bacterial plasmids. Anal. Biochem. 114:193-197.
19. Hooykaas, P. J. J., A. A. N. van Brussel, H. den Dulk-Ras,G. M. S. van Slogteren, and R. A. Schilperoort. 1981. Symplasmid of Rhihobimn t-ifolii expressed in different rhizobialspecies and Agrobacteiiin tiumefaeiens. Nature (London) 291:351-353.
20. Ish-Horowicz, D., and J. F. Burke. 1981. Rapid and efficientcosmid cloning. Nucleic Acids Res. 9:2989-2998.
21. Jarvis, B. D. W., A. G. Dick, and R. M. Greenwood. 1980.Deoxyribonucleic acid homology among strains of Rlizobiomtrifolii and related species. Int. J. Syst. Bacteriol. 30:42-52.
22. Jarvis, B. D. W., M. Gillis, and J. DeLey. 1986. Intra- andintergeneric similarities between the ribosomal ribonucleic acidcistrons of Rlhizobium and Bradwrhizobium species and somerelated bacteria. Int. J. Syst. Bacteriol. 36:129-138.
23. Johnson, J. L. 1984. Nucleic acids in bacterial classification p.8-11. In N. R. Krieg and J. G. Holt (ed.). Bergey's manual ofsystematic bacteriology. vol. 1. The Williams & Wilkins Co.,Baltimore.
24. Johnston, A. W. B., J. L. Beynon, A. V. Buchanan-Wollaston,S. M. Setchell, P. R. Hirsch, and J. E. Beringer. 1978. Highfrequency transfer of nodulating ability between strains andspecies of Rhlizobiiiin. Nature (London) 276:634-636.
25. Jordan, D. C. 1984. Family III Rhizohiaceae Conn. 1938, p.234-256. In N. R. Krieg and J. G. Holt (ed.). Bergey's manualof systematic bacteriology, vol. 1. The Williams & Wilkins Co.,Baltimore.
26. Knudsen, G. R., M. V. Walter, L. A. Porteous, V. J. Prince,J. L. Armstrong, and R. J. Seidler. 1988. Predictive model ofconjugative plasmid transfer in the rhizosphere and phyllo-sphere. Appl. Environ. Microbiol. 54:343-347.
27. Kondorosi, A., E. Kondorosi, C. E. Pankhurst, W. J. Broughton,and Z. Banfalvi. 1982. Mobilisation of a Rhizohiim imelilotimegaplasmid carrying nodulation and nitrogen fixation genesinto other rhizobia and Agrobacteriuln. Mol. Gen. Genet.188:433-439.
28. Lamb, J. W., G. Hombrecher, and A. W. B. Johnston. 1982.Plasmid-determined nodulation and nitrogen-fixation abilities inRhizohiumn plia seoli. Mol. Gen. Genet. 186:449-452.
29. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning, a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
30. McDonell, M. W., M. N. Simon, and F. W. Studier. 1977.Analysis of restriction fragments of T4 DNA and determinationof molecular weights by electrophoresis in neutral and alkalinegels. J. Mol. Biql. 110:119-146.
31. Mozzo, T., E. Cabrera, and T. Ruiz-Argueso. 1988. Diversity ofplasmid profiles and conservation of symbiotic nitrogen fixationgenes in newly isolated Rliizohiini strains nodulating sulla(Hedvsai-rim coronariuln L.) Appl. Environ. Microbiol. 54:1262-1267.
32. O'Connell, M. P., M. F. Hynes, and A. Puhler. 1987. Incompat-ability between a Rliizobiu,m Sym plasmid and a Ri plasmid ofAgrobacteriuln. Plasmid 18:156-163.
33. Plazinski, J., and B. G. Rolfe. 1985. Sym plasmid genes ofRliizobilmn trifolii expressed in Lignobacter and Pseuidolinonas
strains. J. Bacteriol. 162:1261-1269.34. Pramer, D., and E. L. Schmidt. 1965. Experimental soil micro-
biology. Burgess Publishing Co., Minneapolis.35. Ronson, C. W., and D. B. Scott. 1983. Identification, broad host
range mobilisation and mutagenesis of R. trifolii sym: R68.45co-integrate plasmid, p. 177-187. In A. Puhler (ed.), Moleculargenetics of the bacteria-plant interaction. Springer-Verlag, Ber-lin.
36. Schleifer, K. H., and E. Stackebrandt. 1983. Molecular system-atics of procaryotes. Annu. Rev. Microbiol. 37:143-187.
37. Schofield, P. R., M. A. Djordjevic, B. G. Rolfe, J. Shine, andJ. M. Watson. 1983. A molecular linkage map of nitrogenaseand nodulation genes in Rhizobiumn trifoiji. Mol. Gen. Genet.192:459-465.
38. Schofield, P. R., A. H. Gibson, W. F. Dudman, and J. M.Watson. 1987. Evidence for genetic exchange and recombina-tion of Rlhizobiii'i symbiotic plasmids in a soil population. AppI.Environ. Microbiol. 53:2942-2947.
39. Schurter, W., and B. W. Holloway. 1987. Interactions betweenthe transposable element IS21 on R68.45 and Tn7 in Pseiudoino-nicis aeCrulgitiosa PAO. Plasmid 17:61-64.
40. Scott, D. B., C. B. Court, C. W. Ronson, K. F. Scott, J. M.Watson, P. R. Schofield, and J. Shine. 1984. Organisation ofnodulation and nitrogen fixation genes on a Rhizohiimn trifoliisymbiotic plasmid. Arch. Microbiol. 139:151-157.
41. Scott, D. B., and C. W. Ronson. 1982. Identification andmobilization by cointegrate formation of a nodulation plasmid inRhizobiiin truifolii. J. Bacteriol. 151:36-43.
42. Scott, D. B., K.-Y. Chua, B. D. W. Jarvis, and C. E. Pankhurst.1985. Molecular cloning of a nodulation gene from fast andslow-growing strains of Lotius rhizobia. Mol. Gen. Genet. 201:43-50.
43. Scott, K. F., J. E. Hughes, P. M. Gresshoff, J. E. Beringer, B. G.Rolfe, and J. Shine. 1982. Molecular cloning of Rhizobiumntrifoii genes involved in symbiotic nitrogen fixation. J. Mol.AppI. Genet. 1:315-326.
44. Selander, R. K., R. M. McKinney, T. S. Whittam, W. F. Bibb,D. J. Brenner, F. S. Nolte, and P. E. Pattison. 1985. Geneticstructure of populations of Legionella pueiuinophila. J. Bacte-riol. 163:1021-1037.
45. Skot, L., P. R. Hirsch, and J. F. Witty. 1986. Genetic factors inRhizobiuln affecting the symbiotic carbon costs of N, fixationand host plant biomass production. J. AppI. Bacteriol. 61:239-246.
46. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by electrophoresis. J. Mol. Biol.98:503-517.
47. Vincent, J. M. 1970. A manual for the practical study of the rootnodule bacteria, p. 3-4. Blackwell Scientific Publications, Ox-ford.
48. Watson, J. M., and P. R. Schofield. 1985. Species specificsymbiotic plasmid-located repeated DNA sequences in Rhizo-biunm trifolii. Mol. Gen. Genet. 199:279-289.
49. Whitfield, P. L., P. H. Seeburg, and J. Shine. 1982. The humanpro-opiomelanocortin gene: organisation, sequence and inter-spersion with repetitive DNA. DNA 1:133-143.
50. Woomer, P., P. W. Singleton, and B. B. Bohlool. 1988. Ecolog-ical indicators of native rhizobia in tropical soils. Appl. Environ.Microbiol. 54:1112-1116.
51. Young, J. P. W. 1985. Rlhizobiiuuni population genetics: enzymepolymorphism in isolates from peas, clover, beans and lucernegrown at the same site. J. Gen. Microbiol. 131:2399-2408.
52. Young, J. P. W., L. Demetriou, and R. G. Apte. 1987. Rhizo-biion population genetics: enzyme polymorphism in Rhizobiiumleguoninosariutn from plants and soil in a pea crop. Appl.Environ. Microbiol. 53:397-402.
1434 JARVIS ET AL.
on February 13, 2020 by guest
http://aem.asm
.org/D
ownloaded from





























