Embed Size (px)
Citation preview

F’sychiotry Research, 29: 123-l 36 Elsevier
123
Eye Movements in Cocaine Abusers
Joseph L. Demer, Nora D. Volkow, lmogen Ulrich, Kenneth Krajewski, Chester M. Davis, and Franklin I. Porter
Received November 14, 1988; revised version received March 2.5, 1989; accepted June II, 1989.
Abstract. Using electro-oculography, we quantitatively investigated eye move- ments in nine heavy cocaine abusers and three groups of controls. Plasma levels of 3-methoxy-4-hydroxyphenylglycol (MHPG), a major metabolite of brain nor- epinephrine, in cocaine abusers were reduced to an average of 53% of normal. Cocaine abusers had normal smooth pursuit, optokinetic nystagmus, vestibulo- ocular reflex, visual suppression of the vestibulo-ocular reflex, and caloric nystagmus. Data were suggestive of a significant reduction in the degree of immediate enhancement of visual-vestibulo-ocular reflex gain by magnified vision in the cocaine abusers. However, adaptive plasticity of the vestibulo-ocular reflex was normal in the cocaine abusers. These results suggest that, despite animal data linking vestibulo-ocular reflex plasticity to central norepinephrine, this neurotransmitter may not be important to plasticity in human beings.
Key Words. Catecholamines, 3-methoxy-4-hydroxyphenylglycol, cocaine, eye movements. visual-vestibular interaction.
Eye movement recording is a powerful technique for the quantitative evaluation of brain function since eye movement control is mediated by extensive brainstem, cerebellar, and cortical regions. It is thus not surprising that numerous neurological and psychiatric conditions have been associated with abnormal eye movements. Smooth pursuit tracking of small targets is reportedly abnormal in a high percentage of patients with schizophrenia (Lipton et al., 1983). Rapid refixational eye move- ments, called saccades, have been found to have prolonged latency, hypometria, and occasionally reduced velocity in Parkinson’s disease (Leigh and Zee, 1983). Saccadic initiation is markedly abnormal in Huntington’s disease (Leigh and Zee, 1983). The vestibulo-ocular reflex (VOR) and its interaction with visual tracking have been found to be deranged in psychotic schizophrenics (Jones and Pivik, 1985).
Joseph L. Demer, M.D., Ph.D., is Assistant Professor of Ophthalmology and Neurology, Jules Stein Eye Institute and Department of Neurology, University of California at Los Angeles, CA. Nora D. Volkow, M.D., is Director. Department of Medicine, Brookhaven National Laboratory, Upton, NY. Imogen Ulrich. B.A., is Low Vision Coordinator, Cullen Eye Institute, Baylor College of Medicine, Houston, TX. Kenneth Krajewsky, M.D., is Associate Professor, Department of Psychiatry, University of Texas Health Science Center at Houston, TX. Chester M. Davis, Ph.D., is Associate Professor, University of Texas Mental Sciences Institute, University of Texas Health Science Center at Houston, TX. Franklin I. Porter, M.S., O.D., is Assistant Professor of Ophthalmology, Cullen Eye institute, Baylor College of Medicine, Houston, TX. (Reprint requests to Dr. J.L. Demer. Jules Stein Eye Institute, UCLA, 10833 Le Conte Ave., Los Angeles, CA 90024-1771, USA.)
0165-1781 /X9/$03.50 @ 1989 Elsevier Scientific Publishers Ireland Ltd

124
One ocular motor reflex that is well understood in a quantitative sense is the vestibulo-ocular reflex (VOR), which uses head velocity information transduced by the vestibular apparatus to produce compensatory eye movements that stabilize eye
position in space. Visually perceived motion is also tracked by eye movements in cooperation with the VOR to produce visual-vestibular interaction (VVI). The VOR produces eye movements within 12 ms of head movement (Lanman et al., 1978) making it the body’s fastest reflex. As a consequence of this delayed visual information, the VOR must act in a preprogrammed manner, making compensatory eye movements before visual feedback is available. The VOR functions in complete darkness. VOR gain is defined to be the ratio of slow-phase eye velocity to head velocity.
Ideally, VOR gain would be 1.0 to stabilize retinal images perfectly, since this means that a given head velocity evokes an equal eye velocity in the opposite direction. Measured VOR gain is usually slightly lower than this, but in the light the VOR is enhanced by visual-vestibular interaction to increase the gain to 1.0 (Demer et al., 1987). The visually enhanced VOR is called the visual-vestibulo-ocular reflex (VVOR). VVOR gain is equal to the ratio of slow-phase eye velocity to head velocity during measurement in light. Although visual enhancement of VOR gain is physiologically useful, it has only a limited capability to instantaneously influence gain of compensatory eye movements. The brain apparently uses cumulative retinal slip or efference copy of corrective visual pursuit commands, correlated with head movement, to gradually adjust VOR gain, a motor learning process known as VOR
gain plasticity (Miles and Lisberger, 198 1). To stabilize retinal images when telescopic spectacles are worn requires that
VVOR gain increase to equal the magnification of the telescopes (Demer et al., 1987). Such a large gain increase, by a factor of 2 to 8, is physiologically challenging, and both VOR gain plasticity and VVI usually contribute toward achieving it. Significant, although usually incomplete, VOR gain plasticity can readily be demonstrated in normal humans and many animal species (Demer et al., 1987).
Two studies have demonstrated that catecholamine depletion produced by intracisternal injection of 6-hydroxydopamine is associated with impaired VOR gain plasticity (Keller and Smith, 1983; McElligott and Freedman, 1986). This pharmacological manipulation has been demonstrated to result in significant depletion of cerebellar norepinephrine (NE) content (McElligott and Freedman,
1986). Another agent that depletes central catecholamines is cocaine. Animal ex-
periments have demonstrated that cocaine blocks presynaptic reuptake of NE (Had- field et al., 1980) and dopamine (DA) (Koe, 1976). Decreased levels of NE and DA have been found in the brains of chronically treated animals (Ho et al., 1977) and decreased noradrenergic activity in chronic cocaine abusers has been reported in studies measuring 3-methoxy-4-hydroxyphenylglycol (MHPG), a major metabolite of brain NE (Tennant, 1985; Krajewski and Davis, 1987) and an indicator of central noradrenergic activity when measured in plasma (Elsworth et al., 1982). Taken together, such evidence suggests that chronic cocaine abusers might exhibit abnormal VOR gain plasticity, and perhaps other ocular motor abnormalities.

125
Methods
Nine right-handed cocaine abusers (average age = 29.2 + 4.2 years, range 25-34) were studied (Table I). Patients were obtained from a 6-week inpatient program for drug detoxification, and were tested between 6 and 27 (mean IS. I + 7.5) days after admission. Duration of cocaine abuse ranged from 3 to 120 (mean 44 + 40) months. Inclusion criteria were: (I) I)SM-III criteria (Committee on Nomenclature and Statistics, 1980) as assessed by two independent psychiatrists, N.D.V. and K.J.K.; (2) at least 3 months of cocaine abuse before the current hospitalization; and (3) consumption of an average of I .4 5 I.8 grams (range 0.3-5.0) of cocaine per day administered via snorting or freebase routes. Patients with another concurrent mental disorder or with a history of past psychiatric illnesses apart from cocaine abuse were excluded. Absence of cocaine consumption during hospitalization was monitored with periodic evaluations of cocaine metabolites in urine (Ambre et al., 1982).
Patients underwent an eye examination to verify that they were visually normal. Ma.jor medical illnesses were ruled out for all patients. Most of the patients abused other drugs such as marijuana and alcohol sporadically, but none of them were addicted to any of these drugs. and by history had not used them in the 30 days preceding the study. A minority of subjects used prescription medications, but were instructed to abstain from their USC on the day of testing.
Three control groups were employed, taken retrospectively from subjects undergoing eye movement testing batteries for other studies. Data in these subjects arc valid for control purposes, since testing and data analysis were according to automated protocols unchanged
Table 1. Clinical characteristics of cocaine abusers
Cocaine Abuse amount Other
Patient Age duration abused drugs No. Sex (vr) (mo) Route Wdav) abused Medications
1 M 26 120
2 F 33 3
3 M 26 60
4 M 27 12
Inhalation,
Smoking
Smoking
Smoking
Smoking
M 34 6 Smoking
F 25 76 Smoking
7 F 26 10 Smoking 10 Marijuana
6 M 27 72 Inhalation
9 M 37 36 Inhalation
Nofe. M = male. F = female.
7 Marijuana
LSD, PCP
1.5
3
5
Marijuana
Phenobarbital
Diazepam
Alcohol
Marijuana
Mandrex
0.75 Alcohol Marijuana
Tobacco
5 Marijuana
Alcohol
Tobacco
2.5 Alcohol
2 Marijuana
Alcohol
Desipramine
Acetominophen
Acetominophen
Chlordiazepoxide
Haloperidol
Acetominophen
Desipramine
Acetominophen
Acetominophen
Ibuprofen
Acetominophen
Desipramine
Acetominophen

126
during testing of all subjects and patients. The largest was the age- and sex-matched control group, consisting of IX healthy volunteers; while there was no reason to suspect that these subjects were using illicit drugs, they had been questioned only regarding medication ingestion and had not specifically denied other drug use. The average age of the age- and sex-matched control group was 2X.4 -t 4. I years.
To confirm that the age- and sex-matched control group’s data were not significantly influenced by unrecognized drug abuse. we identified a smaller control group of nine previously tested normal subjects who were recontacted and denied any history of drug abuse. These subjects were considered the normal, drug-free control group, and had an average age of 32.2 + 16.3 years. A separate plasticity control group was identified because this specialized type of testing is lengthy and was rarely performed in subjects undergoing other studies. The plasticity control group was another age-matched group of nine healthy volunteers. who also specifically denied any history of drug abuse and had an average age of 29.3 * 4.5 years.
Catecholamine depletion in cocaine abusers was determined by plasma M HfYi levels measured using liquid chromatography and electrochemical detection. All blood samples were drawn at approximately IO:00 a.m. An internal standard (4-methoxy-3-hydroxyphenylglycol) was added to plasma which was then extracted in ethyl acetate after acid and base partitioning. A standard curve prepared by addition of known amounts of M H PG was used to quantitate the M H PC; in plasma. The interassay coefficient of variation of repeated runs (n = 9) for a pooled standard over a 2-month period was 4.7% I.ogistical considerations related to date of hospital admission and laboratory operation made it impossible to obtain M HPG levels in some cocaine abusers. Since eye movement control sub.jects were asccrtaincd retrospectively, M H PC levels were not measured in these sub.jects.
Horirontal eye movements were recorded using DC-coupled, digitally sampled (200 HI) electro-oculography with bandwidth 40 H7, as described elsewhere (Demer et al., 19X7). Stimulus presentation and data acquisition were under the control of a digital computer. To study saccades, we had subjects track a projected spot of light deflected by computer- controlled mirror galvanometers to produce a pseudo-random pattern of target drsplacements of varying si7es up to 40”. To study smooth pursuit. we had subjects track a spot of light moving sinusoidally at 0.2 H7, velocity amplitude 3O”/sec (half the peak-to-peak value). Pursuit gain was computed as the ratio of response velocity to stimulus velocity. To study optokinetic nystagmus (OKN), we surrounded subjects by a full-field, cylindrrcal curtain having vertical black stripes sinusoidally rotated at 0.2 H7, velocity amplitude 30”;‘scc.
Normal peripheral vestibular function was verified in cocaine abusers using warm and cold water caloric testing, with data analysis as conventionally described (Coats, 1975). Caloric testing evokes nystagmus by stimulating the vestibular apparatus using thermal gradients produced by irrigating the external auditory canals with warm and cool water. The VOR was studied in darkness during vertical axis, whole-body, sinusoidal rotation in a servomotor- driven chair. Rotations were performed at 0.0125, 0.05, 0. I. and 0.2 HI, with velocity amplitude 60’/sec. VOR fixation suppression was studied by instructing subjects to visually fixate a target light attached to the chair during rotation at 0.05 Hz, velocity amplitude hO”/sec, in an otherwise darkened environment. The VVOR was studied by recording eye movements as subjects were sinusoidally rotated without magnifying spectacles in a lighted environment at 0.05 H7, velocity amplitude 30”/sec. VOR and VVOR gains were computed as described elsewhere (Demer et al., 1989h).
We used binocular, 4X telescopic spectacles having a visual field diameter of 10.3” to study VOR gain plasticity and VVI with magnified vision. The peripheral field outside the telescope eyepieces was occluded. After initial measurement of VOR and VVOR gains, subjects underwent adapative training for I5 min during sinusoidal rotation at 0.2 H7. amplitude 20”/sec. In the training, which commonly produces an adaptive VOR gain increase (Demer et al., 1989h), subjects viewed with telescopic spectacles a distant video monitor displaying entertainment programming. After adaptation, VOR and VVOR measurements were
repeated.

127
Results
At least one measurement of plasma MHPG level was made in each of five subjects during hospital admission. Mean MHPG level was 12.6 * 3.5 pmol/ml (mean * SD, n = 5). The comparable value for 22 normal laboratory staff members was 23.7 * 6.4 pmol/ml (n = 22) (Krajewski and Davis, 1987). This difference, a reduction to 53% of control, is significant at the p < 0.0005 level; in two patients, MHPG levels were more than 2 SD below the mean value for controls. Repeated MHPG measurements were made in three patients. In patient 4, the MHPG level 1 week before eye movement testing was 15.5 pM/ ml, and was 12.7 pM/ ml on the first of 2 days of eye movement testing. In patient 5, the MHPG level was 14.6 PM/ml 3 weeks before eye movement testing, and was 12.7 pM/ml 2 weeks before eye movement testing. In patient 6, the MHPG level was 17.5 pM/ml five days before eye movement testing, and was 9.5 pM/ml4 weeks after testing. While it was not possible to verify the exact onset of abstinence from cocaine in these subjects, these data indicate such a prolonged reduction that levels would not be sensitive to the exact date of
measurement. Cocaine abusers and two groups of controls were subjected to a comprehensive
battery of quantitative eye movement tests. During eye movement testing, cocaine abusers performed according to instructions, but often became distracted from ocular motor tasks and expressed irritability. Both cocaine abusers and normal control subjects tended to become sleepy during data collection, but were alerted by mental exercise tasks.
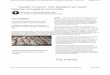
Representative saccadic eye movement data are shown in Fig. 1 for patient 6, and illustrate normal performance 9 days after initiation of cocaine detoxification for
daily consumption of 5 g. A quantitative comparison of saccadic eye movements of cocaine users and controls is seen in Table 2. Saccadic latency is defined as the time delay between movement of the target and initiation of the eye movement. Saccadic
accuracy is defined as the size (“) of the initial saccadic eye movement divided by the size (“) of the target movement, expressed as a percentage. It may be seen that there was no significant difference in saccadic latency, velocity, or accuracy between cocaine users and either group of controls.
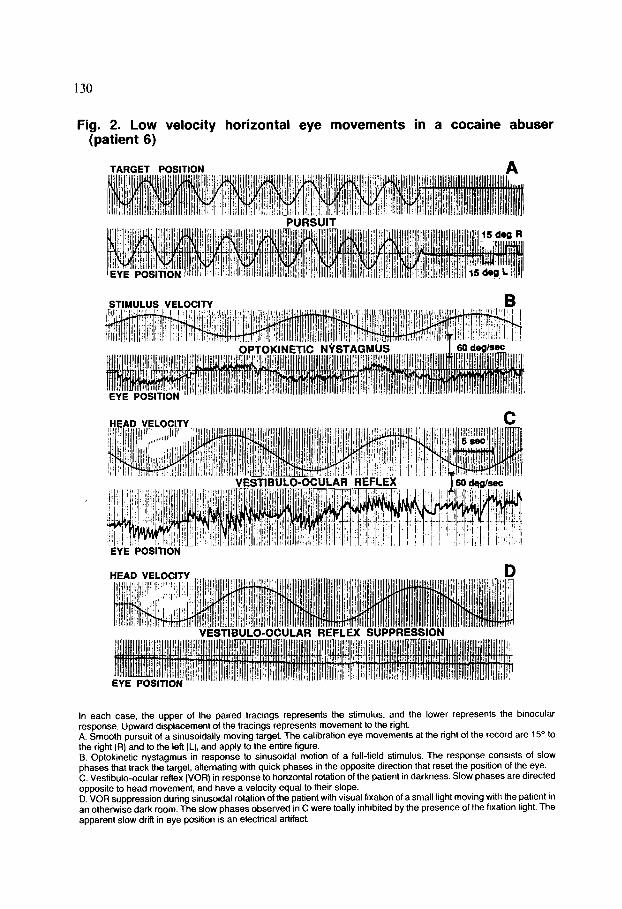
Representative, unprocessed slow eye movement data are illustrated in Fig. 2 for patient 6. Performance is strikingly normal. The digitally sampled, slow-phase eye velocity data illustrated in Fig. 3 are similar to data produced in analysis of all the types of eye movements involving sinusoidal stimuli. Summary data on charac- teristics of tracking eye movements and caloric nystagmus in cocaine abusers and control subjects are provided in Table 3. There was no significant difference between cocaine users and controls in gain of smooth pursuit, as well as OKN, for a sinusoidally moving stimulus. VOR gain and phase were not significantly different for cocaine users and controls at any frequency tested. Gain during VOR fixation suppression was not significantly different for the cocaine users and control group. VVOR gain with unmagnified vision was essentially equal to the ideal value of 1.0 for all groups. Caloric responses were normal in both groups. There was a difference in cumulative eye position between the cocaine abusers and the matched controls that was statistically significant @ = 0.02), but since it was not confirmed in the
Fig
. 1 I
Sac
cad
ic e
ye m
ove
men
ts i
n a
coca
ine
abu
ser
(pat
ien
t 6)
TSQ
dea
LE
FT
EY
E
b 4
c S
OS
?m
SO
S
ym
coo
SW
so0
100
0 10
$0
S
o 10
0
10
co
30
\ 40
LCrr
<
--
Nt?
LlT
tJO
C(D
CE
) ->
o*
onl
A. H
oriz
onta
l eye
pos
lion
durin
g tr
acki
ng o
f a ra
ndom
ly m
ovin
g ta
rget
. Onl
y a
port
ion
of th
e en
tire
reco
rd is
sho
wn.
R =
rig
ht; L
= l
eft.
Tim
e is
sho
wn
from
left
to r
ight
. 6.
His
togr
ams o
f sac
cadi
c la
tenc
y W
ay
time)
and
acc
urac
y fo
r bo
th e
yes
obta
ined
by
digi
tal p
roce
ssin
g of
sac
cade
s in
clud
ing
thos
e sh
own
in A
. MS
= m
illis
econ
d.
C. R
elat
ions
hip
betw
een
sacc
adic
am
plitu
de a
nd v
eloc
ity fo
r the
left
eye,
for s
acca
des
incl
udin
g th
ose
in A
. The
nor
mal
rang
e is
bou
nded
by
the
dotte
d cu
rve.
Sac
cadi
c ve
loci
ties
for t
his
patie
nt a
re a
t the
upp
er li
mits
of t
hose
see
n in
nor
mal
sub
ject
s.
D. S
imila
r to
C, b
ut fo
r the
rig
ht e
ye.
129
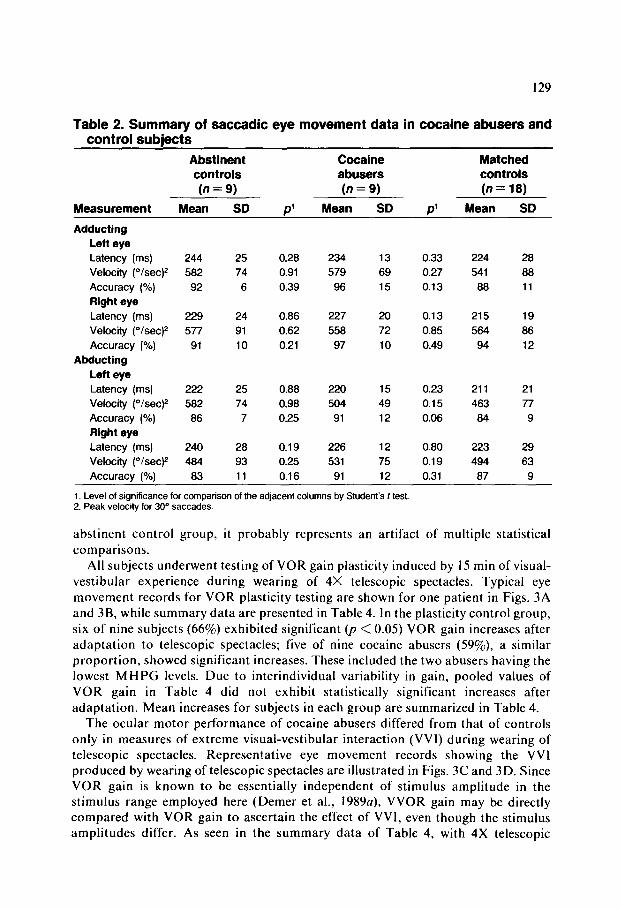
Table 2. Summary of saccadic eye movement data in cocaine abusers and control subjects
Cocaine abusers (n = 9)
Mean SD Measurement
Abstinent controls (n = 9)
Mean SD
Matched controls (n = 18)
Mean SD p’ P’
Adducting
Lefl eye
Latency (ms)
Velocity (O/set)* Accuracy (%) Right eye
Latency (ms) Velocity ("lsec)*
Accuracy (%)
Abducting
Left eye
Latency (ms)
Velocity ("/sec)2
Accuracy (%)
Right eye
Latency (ms)
Velocity (O/set)*
244 25 0.28 234 13 0.33 224 28
582 74 0.91 579 69 0.27 541 88
92 6 0.39 96 15 0.13 88 11
229 24 0.86 227 20 0.13 215 19
577 91 0.62 558 72 0.85 564 86
91 10 0.21 97 10 0.49 94 12
222 25 0.88 220 15 0.23 211 21
582 74 0.98 504 49 0.15 463 77
86 7 0.25 91 12 0.06 84 9
240 28 0.19 226 12 0.80 223 29
484 93 0.25 531 75 0.19 494 63
Accuracy (%) 83 11 0.16 91 12 0.31 87 9
1. Level of significance for comparison of the adjacent columns by Student’s t test. 2. Peak velocity for 30“ saccades.
abstinent control group, it probably represents an artifact of multiple statistical comparisons.
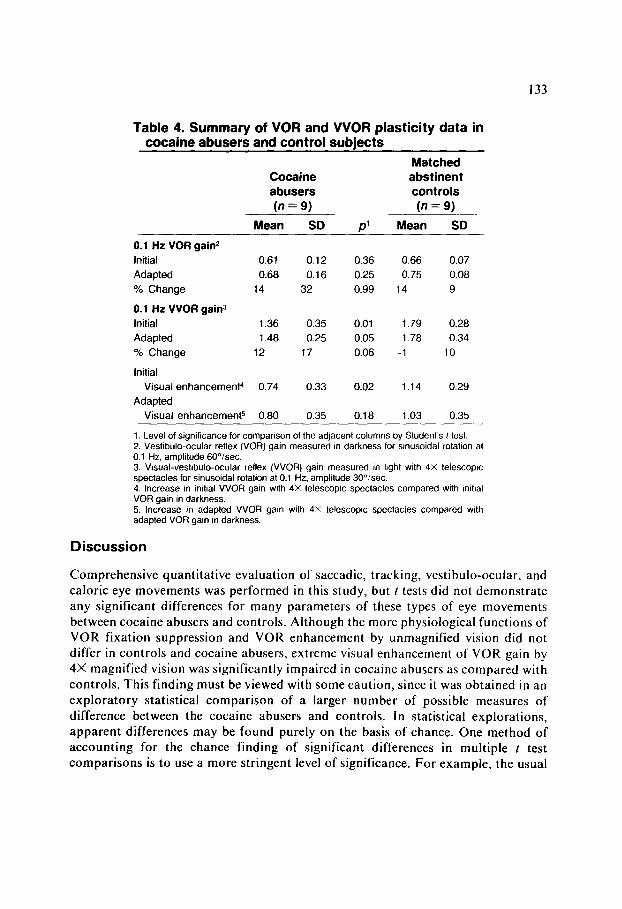
All subjects underwent testing of VOR gain plasticity induced by 15 min of visual- vestibular experience during wearing of 4X telescopic spectacles. Typical eye movement records for VOR plasticity testing are shown for one patient in Figs. 3A and 3B, while summary data are presented in Table 4. In the plasticity control group, six of nine subjects (66%) exhibited significant (p < 0.05) VOR gain increases after adaptation to telescopic spectacles; five of nine cocaine abusers (59%), a similar proportion, showed significant increases. These included the two abusers having the lowest M HPG levels. Due to interindividual variability in gain, pooled values of VOR gain in Table 4 did not exhibit statistically significant increases after adaptation. Mean increases for subjects in each group are summarized in Table 4.
The ocular motor performance of cocaine abusers differed from that of controls only in measures of extreme visual-vestibular interaction (VVI) during wearing of telescopic spectacles. Representative eye movement records showing the VVI produced by wearing of telescopic spectacles are illustrated in Figs. 3C and 3D. Since VOR gain is known to be essentially independent of stimulus amplitude in the stimulus range employed here (Demer et al., 1989a), VVOR gain may be directly compared with VOR gain to ascertain the effect of VVI, even though the stimulus amplitudes differ. As seen in the summary data of Table 4, with 4X telescopic
130
Fig. 2. Low velocity horizontal eye movements in a cocaine abuser (patient 6)
STIMULUS VELOCITY B
HEAD VELOCITY.
EYE POSfTlON
In each case, the upper of the paired tracings represents the stimulus, and the lower represents the binocular response. Upward displacement of the tracings represents movement to the right. A. Smooth pursuit of a sinusoidally moving target. The calibration eye movements at the right of the record are 15” to the right (R) and to the left (L), and apply to the entrre figure. B. Optokinetic nystagmus in response to sinusoidal motion of a full-field stimulus. The response consrsts of slow phases that track the target, alternating with quick phases in the opposite direction that reset the position of the eye. C. Vestibulo-ocular reflex (VOR) in response to horizontal rotation of the patient in darkness. Slow phases are directed opposite to head movement, and have a velocity equal to their slope. D. VOR suppression during sinusoidal rotation of the patient with visual fixation of a small light moving with the patient In an otherwise dark room. The slow phases observed in C were toally inhibited by the presence of the fixation light. The apparent slow drift in eye position is an electrical artifact.
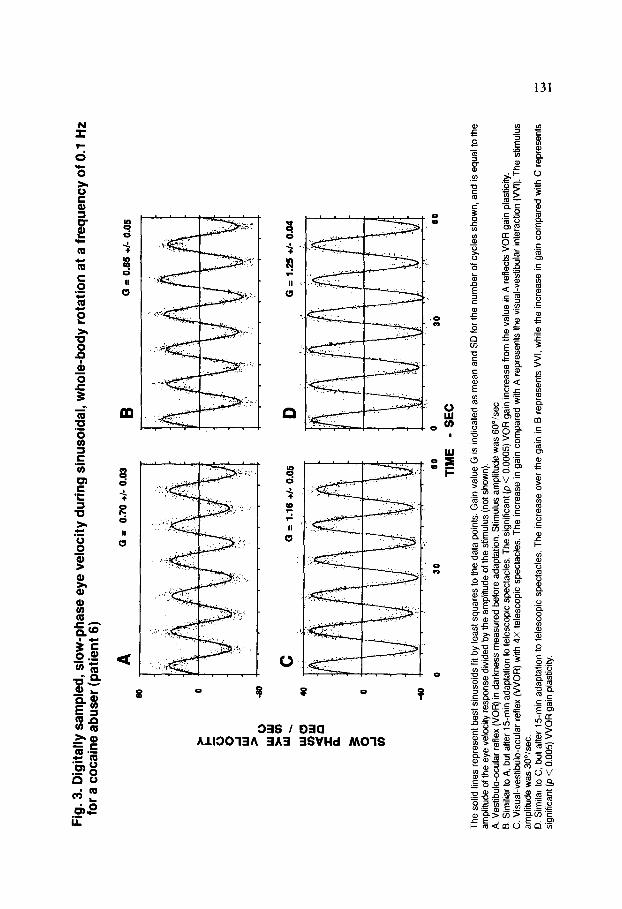
Fig
. 3.
Dig
ital
ly s
amp
led
, sl
ow
-ph
ase
eye
velo
city
du
rin
g s
inu
soid
al,
wh
ole
-bo
dy
rota
tion
at
a fr
equ
ency
of
0.1
Hz
for
a co
cain
e ab
use
r (p
atie
nt
6)
B
a =
0.65
+I
- 0.
05
C
G
= 1.
16
+I-
0.06
D
0
= 1.
25
+I-
0.04
0 30
60
TIM
E
0 3b
60
- S
EC
The
sol
id l
ines
re
pres
ent
best
sin
usoi
ds
fit b
y le
ast
squa
res
to t
he d
ata
poin
ts.
Gai
n va
lue
G i
s In
dica
ted
as m
ean
and
SD
for
the
num
ber
of c
ycle
s sh
own,
an
d is
equ
al
to t
he
ampl
itude
of
the
eye
velo
city
re
spon
se
divi
ded
by t
he a
mpl
itude
of
the
stim
ulus
(n
ot s
how
n).
A.
Ves
tibul
o-oc
ular
re
flex
(VO
R)
in d
arkn
ess
mea
sure
d be
fore
ada
ptat
ion.
S
timul
us
ampl
itude
w
as 6
0”/s
ec.
B. S
imili
ar t
o A
, bu
t af
ter
15m
in
adap
tatio
n to
tel
esco
pic
spec
tacl
es.
The
sig
nific
ant
(p <
0.
0005
) V
OR
gai
n in
crea
se
from
the
val
ue
in A
ref
lect
s V
OR
gai
n pl
astic
ity.
C.
Vis
ual-v
estib
ule-
ocul
ar
refte
x (V
VO
R)
with
4X
te
lesc
opic
sp
ecta
cles
. T
he
incr
ease
in
gai
n co
mpa
red
with
A r
epre
sent
s th
e vi
sual
-ves
tibul
ar
inte
ract
ion
(VV
I).
The
st
imul
us
ampl
itude
w
as 3
0°/s
ec.
D.
Sim
ilar
to C
. bu
t af
ter
15m
in
adap
tatio
n to
tel
esco
pic
spec
tacl
es.
The
in
crea
se
over
the
gai
n in
B r
epre
sent
s V
VI,
whi
le
the
incr
ease
in
gai
n co
mpa
red
with
C r
epre
sent
s si
gnrf
ican
t @
< 0
.005
) V
VO
R g
ain
plas
ticity
.
__
~._
--
_ ~
..__
___-
-

132
spectacles, both initial and adapted VVOR gains were significantly lower in the cocaine abusers than in the controls. When compared within individual subjects, the amount of gain enhancement initially produced by the wearing of 4X telescopic spectacles (that is, the difference between initial VOR gain and initial VVOR gain) was significantly lower in the cocaine abusers than in the controls. Although the average amount of immediate gain enhancement in cocaine users was lower than in controls, after adaptation to telescopic spectacles, four of nine cocaine abusers and two of nine controls exhibited statistically significant increases in VVOR gain (as compared with values for the same subjects before adaptation).
Table 3. Summary of tracking and vestibular eye movement data in cocaine abusers and control subjects
Abstinent Cocaine Matched controls abusers controls (n = 9) (n=9) (n = 18)
Measurem~t Mean SD p1 Mean SD p1 Mean SD
Pursuit gain* 0.0125 Hz VOR3
Gain Phase (O)
0.05 Hz VOR3 Gain Phase (“)
0.2 Hz VOFP Gain Phase(O)
VOR suppression4 VVOR gain5
OKN gains
Caloric nystagmus Unilateral weak7
Directional preponderance’
Cumulative
0.90 0.04 0.57
0.57 0.12 0.77
32 5 0.64
0.70 0.10 0.59
9 2 0.97
0.77 0.13 0.97
-2 3 0.19
0.03 0.01 0.57
7.03 0.03 0.33
0.91 0.10 0.64
5.90 7.12 0.83
6.43 6.11 0.69
0.91 0.07 0.21 0.94 0.05
0.59 0.13 0.49 0.56 0.12
31 6 0.70 31 8
0.72 0.09 0.23 0.67 0.10
9 6 0.93 9 4
0.68 0.16 0.40 0.72 0.11
0 2 0.87 0 4
0.03 0.02 0.33 0.02 0.01
1 .oo 0.08 0.56 1.02 0.09
0.88 0.14 0.39 0.93 0.15
6.65 6.06 0.50 8.46 6.22
7.91 7.91 0.79 8.81 7.33
position (0) 8450 3105 0.94 8588 3336 0.02 5772 2203
1. Level of significance for comparison of the adjacent columns by Student’s t test. 2. For tracking of a target moving sinusoidally at 0.2 Hz, ampfiiude 30°/sec. 3. Vestibulo-ocular reflex (VOR) measured for sinusoidal rotation with amplitude 60°/sec. 4. Gain during VOR suppression. measured for sinusoidal rotation at 0.05 Hz, amplitude 60”lsec. with a fixation light moving with the subject, in a dark room. 5. Visual-vestibulo-ocular reflex (VVOR) gain measured for sinusoidal rotation in light (without magnifying spectacles). at 0.02 Hz, amplitude 3Pisec. 6. Optokinetic nystagmus (OKN) gain measured for sinusoidal rotation of optokinetic drum at 0.05 Hz. amplitude 3fY1sec. 7. In percent.
133
Table 4. Summary of VOR and VVOR plasticity data in cocaine abusers and control subiects
Cocaine abusers (n = 9)
Mean SD
0.1 Hz VOR gain2
Initial
Adapted
% Change
0.1 Hz VVOR gain3
Initial
Adapted
% Change
Initial
Visual enhanceme&
Adapted
Visual enhancemeW
0.61 0.12 0.36 0.66 0.07
0.68 0.16 0.25 0.75 0.08
14 32 0.99 14 9
1.36 0.35 0.01 1.79 0.28
1.48 0.25 0.05 1.78 0.34
12 17 0.06 -1 10
0.74
0.80 0.35 0.18 1.03 0.35
0.33
P’
0.02
Matched abstinent controls
(n = 9)
Mean SD
1.14 0.29
1. Level of significance for comparison of the adjacent columns by Student’s t test. 2. Vestibulo-ocular reflex (VOR) gain measured In darkness for sinusoidal rotation at 0.1 Hz, amplitude 60”/sec. 3. Visual-vestibulo-ocular reflex (VVOR) gain measured in light with 4X telescopic spectacles for sinusoidal rotation at 0.1 Hz. amplitude 30”/sec. 4. Increase in irMaI VVOR gain with 4X telescoptc spectacles compared with mihal VOR gain in darkness. 5. Increase in adapted VVOR gain with 4X telescopic spectacles compared with adapted VOR gam in darkness
Discussion
Comprehensive quantitative evaluation of saccadic, tracking, vestibulo-ocular, and caloric eye movements was performed in this study, but I tests did not demonstrate any significant differences for many parameters of these types of eye movements between cocaine abusers and controls. Although the more physiological functions of VOR fixation suppression and VOR enhancement by unmagnified vision did not differ in controls and cocaine abusers, extreme visual enhancement of VOR gain by 4X magnified vision was significantly impaired in cocaine abusers as compared with controls. This finding must be viewed with some caution, since it was obtained in an exploratory statistical comparison of a larger number of possible measures of difference between the cocaine abusers and controls. In statistical explorations, apparent differences may be found purely on the basis of chance. One method of accounting for the chance finding of significant differences in multiple t test comparisons is to use a more stringent level of significance. For example, the usual

134
level of 0.05 could be divided by the number of t test comparisons made. However, this approach is not entirely satisfactory since it cannot account for the possible large number of z tests that were or might have been made, but are not reported. We prefer to regard the current finding as suggestive, but not conclusive, and to examine corroborating data. Of the total of 58 I-test results reported here, the great majority showed no significant differences. However, two significant differences and two nearly significant differences (Table 4) all represent measures of one physiological variable: visual-vestibular interaction with magnified vision. While chance might be expected to produce occasional significant t-test values for random physiological variables, the impairment in visual-vestibular interaction was found both before and after adaptation to telescopic spectacles, making it likely to be a real deficit rather than a chance artifact of data sampling. Although additional study of this issue seems warranted, confirmation of this abnormality would imply that cocaine abusers have a functional eye movement deficit only when wearing telescopic spectacles; the observed measures of visual-vestibular interaction under normal, unmagnified viewing conditions were intact. Telescopic spectacles are much more challenging to visual-vestibular interaction than unmagnified vision, so this deficit does not imply functional disability under normal conditions.
Since cocaine has been known to induce schizophrenic-like psychosis, and since marked eye movement abnormalities have been described in schizophrenic patients, it is notable that most eye movements were normal in chronic cocaine abusers. Though none of the cocaine abusers tested were psychotic at the time of the study, their lack of eye movement abnormalities may reflect more widespread dysfunction in the brain in schizophrenia as opposed to more focal dysfunction induced by chronic cocaine consumption.
The cocaine abusers studied here exhibited plasma levels of MHPG, a major metabolite of brain NE, averaging only 53% of normal. Serial M H PG measurements suggested that levels continued to decline during inpatient detoxification despite presumed abstinence from cocaine. Although some fluctuations in M HPG levels are to be expected due to diurnal variations (Wehr et al., 1980; Baker et al., 1988), all blood samples were collected at midmorning to minimize this effect. Although NE levels increase with upright posture and wakefulness (Linsell et al., 1985) patient activity in the detoxification unit remained ad lib throughout hospitalization, minimizing activity as a potential source of systemic variation in this MHPG precursor. Three cocaine abusers were under treatment with desipramine, an antidepressant known to reduce MHPG in the cerebrospinal fluid (Potter et al., 1985). Desipramine, a relatively specific inhibitor of NE uptake, would be expected to produce a reduction similar to cocaine in central NE.
It is thus likely that these cocaine abusers were to some degree depleted of brain NE. However, despite the finding that NE depletion in cats severely decreases VOR gain plasticity (Keller and Smith, 1983; McElligott and Freedman, 1986), the cocaine abusers exhibited normal VOR gain plasticity in a 15-min adaptation paradigm. It is possible that the depletion of brain NE in cocaine abusers was insufficient to inhibit plasticity, since NE levels in the cerebellums of cats showing impaired plasticity averaged only 24% of control levels (McElligott and Freedman, 1986). Plasma

135
M HPG levels may not necessarily reflect local NE concentration in individual brain structures, and these may be influenced differently by cocaine in humans than by 6-hydroxydopamine in cats. A third explanation of this discrepancy may lie in possible differences in the methods of demonstration of VOR gain plasticity. The cat experiments involved measurement of VOR gain plasticity induced over a period of at least 4 hours, while in the present study plasticity was induced over a period of 15 min. It is possible that different mechanisms having different sensitivities to NE depletion operate to modify VOR gain over short and long time periods. Since the cocaine abusers had frequently been treated with psychotropic medications until the day before eye movement testing, a role of such medications in maintaining plasticity despite NE depletion might be hypothesized. Nevertheless, the present results suggest that NE may not be important in VOR gain plasticity in human beings.
The only eye movement deficit in cocaine abusers identified in the exploratory analysis was in VW with magnified vision. The visual enhancement of VOR gain is a complex phenomenon known to be deranged by lesions of the vestibulocerebellum (Baloh et al., 1981). Although the neurotransmitters involved in VVI are unknown, the present findings suggest that catecholamines may be important. It is important to note that, although immediate VVI was impaired in the cocaine abusers, VOR gain plasticity, a long-term adaptive modification, was not measurably impaired. This confirms the dissociation of these two phenomena reported after visual cortical lesions in cats (Tusa et al., 1989) suggesting that different structures underlie VVI
than underlie VOR gain plasticity. These findings should be interpreted cautiously because they are based upon a
relatively small number of individuals who abused cocaine in an uncontrolled environment. The unreliability of drug abusers with respect to history and quantity of drug abused, as well as consumption of impure material obtained from the streets, complicates historical assessment of substances consumed. However, depressed levels ol‘ MHPG provide objective corroboration of catecholamine depletion, and the clearly normal VOR plasticity and other eye movement responses in the cocaine abusers are striking. Further investigations into the neuropharmacological effects of cocaine consumption are warranted.
Acknowledgments. This study was supported by NE1 grant EY-06394 and the Clayton Foundation for Research. Computer programming was done by Margaret Kallsen, Bradford Daniels, and Philip Szeto. Joseph L. Demer received support from the Karl Kirchgessner Foundation, Research to Prevent Blindness, and the Permanent Charities Committee of the Entertainment Industry.
References
Ambre, J.J.; Ruo, T.H.; Smith, G.L.; Backes, D.; and Smith, C.M. Ecgonine methyl ester, a major metabolite of cocaine. Journal of’Anafytical Tuxicofogy. 6:26-29, 1982.
Baker, N.J.; Adler, L.E.; Waldo, M.; Gerhardt, G.; Drebing, C.; Cox, B.; Berry, S.; Phillips, W.; and Freedman, R. Reproducibility of the measurement of plasma noradrenergic and dopaminergic metabolites in normal subjects. Psychiatry Research, 23: I 19- 130, 1988.
Baloh, R.W.; Yee, R.D.; Kimm, J.; and Honrubia, V. Vestibulo-ocular reflex in patients with lesions involving the vestibulocerebellum. Experimental Neurology, 72:141-152, 1981.

136
Coats, A.C. Electronystagmography. In: Bradford, L.J., ed. Physiological Measures of the Audio-vestibular System. New York: Academic Press, 1975.
Committee on Nomenclature and Statistics. DSM-III: Diagnostic and Statistical Manual of Mental Disorders. 3rd ed. Washington, DC: American Psychiatric Association, 1980.
Demer, J.L.; Goldberg, J.; Jenkins, H.A.; and Porter, F.I. Vestibulo-ocular reflex during magnified vision: Adaptation to reduce visual-vestibular conflict. Aviation, Space, and Environmental Medicine, 58(Suppl. 9):Al75-Al79, 1987.
Demer, J.L.; Goldberg, J.; Porter, F.1.; Jenkins, H.A.; and Schmidt, K. Visual-vestibular interaction with telescopic spectacles. Submitted for publication, 1989a.
Demer, J.L.; Porter, F.1.; Goldberg, J.; Jenkins, H.A.; and Schmidt, K. Adaptation to telescopic spectacles: Vestibulo-ocular reflex plasticity. Investigative Ophthalmology and Visual Science, 30: l59- 170, 1989b.
Elsworth, J.D.; Redmond, D.E., Jr., and Roth, H.R. Plasma and cerebrospinal fluid 3- methoxy-4-hydroxyphenethylene glycol (M HPG) as indices of brain norepinephrine metabolism in primates. Brain Research, 235: I I5 124, 1982.
Hadfield, M.G.; Mott, D.E.W.; and Ismay, J.A. Cocaine: Effect of in vivo administration on synaptosomal uptake of norepinephrine. Biochemical Pharmacology, 29: l86l- 1863, 1980.
Ho, B.T.; Taylor, D.L.; Estevez, V.S.; Englert, L.F.; and McKenna, M.L. Behavioral effects of cocaine-Metabolic and neurochemical approach. In: Ellinwood, E.H., and Kilbey, M.M., eds. Cocaine and Other Stimulants. New York: Plenum Press, 1977.
Jones, A.M., and Pivik, R.T. Vestibular activation, smooth pursuit tracking, and psychosis. Psychiatry Research, I4:29 l-308, 1985.
Keller, E.L., and Smith, M.J. Suppressed visual adaptation of the vestibulo-ocular reflex in catecholamine-depleted cats. Brain Research, 258:323-327, 1983.
Koe, B.K. Molecular geometry of inhibition of the uptake of catecholamines and serotonin in synaptosomal preparations of the rat brain. Journal of Pharmacology and Experimental Therapeutics, 199:649-661, 1976.
Krajewski, K.J., and Davis, CM. Cocaine abstinence symptomatology and MHPG. Proceedings of the American Psychiatric Association, 1987. p. 236.
Lanman, J.; Bizzi, E.; and Allum, J. The coordination of eye and head movement during smooth pursuit. Brain Research, 153:39-53, 1978.
Leigh, R.J., and Zee, D.S. The Neurology of Eye Movements. Philadelphia: F.A. Davis, 1983.
Linsell, C.R.; Lightman, S.L.; Mullen, P.E.; Brown, M.J.; and Causon, R.C. Circadian rhythms of epinephrine and norepinephrine in man. Journal of Clinical Endocrinology and Metabolism. 60:1210-1215, 1985.
Lipton, R.B.; Levy, D.L.; Holzman, P.S.; and Levin, S. Eye movement dysfunctions in psychiatric patients: A review. Schizophrenia Bulletin. 9: 13-32, 1983.
McElligott, J.G., and Freedman, W. Pharmacological manipulation of vestibular plasticity. In: Keller, E.L., and Zee, D.S., eds. Adaptive Processes in Visual and Oculomotor Systems. Oxford: Pergamon Press, 1986.
Miles, F.A., and Lisberger, S.G. Plasticity in the vestibulo-ocular reflex: A new hypothesis. Annual Review of Neuroscience, 41273-299, 198 I.
Potter, W.Z.; Scheinin, M.; Golden, R.N.; Rudorfer, M.V.; Cowdry, R.W.; Cal& H.M.; Ross, R.J.; and Linnoila, M. Selective antidepressants and cerebrospinal fluid: Lack of specificity on norepinephrine and serotonin metabolites. Archives of General Psychiatry, 42:1171-l 177, 1985.
Tennant, F.S. Effect of cocaine dependence on plasma phenylalanine and tyrosine levels and on urinary M H PG excretion. American Journal of Psychiatry, 142: 1200-l 20 I, 1985.
Tusa, R.J.; Demer, J.L.; and Herdman, S.J. Cortical areas involved in OKN and VOR in cats: Cortical lesions. Journal of Neuroscience, 9: I l63- I 178, 1989.
Wehr, T.A.; Muscettola, G.; and Goodwin, F.K. Urinary 3-methoxy-4-hydroxyphenyl- glycol circadian rhythm: Early timing (phase advance) in manic-depressives compared with normal subjects. Archives of General Psychiatry, 37:257-263, 1980.