Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, June 2009, p. 2488–2498 Vol. 77, No. 60019-9567/09/$08.00�0 doi:10.1128/IAI.00919-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
The Fasciola hepatica Tegumental Antigen Suppresses Dendritic CellMaturation and Function�
Clare M. Hamilton,1 David J. Dowling,1 Christine E. Loscher,2 Russell M. Morphew,3Peter M. Brophy,3 and Sandra M. O’Neill1*
Parasite Immune Modulation Group, School of Nursing,1 and Immunomodulation Group, School of Biotechnology,2
Dublin City University, Glasnevin, Dublin 9, Ireland, and Institute of Biological Sciences, Aberystwyth University,Aberystwyth, Ceredigion, Wales, United Kingdom3
Received 24 July 2008/Returned for modification 28 August 2008/Accepted 23 March 2009
Parasitic worms and molecules derived from them have powerful anti-inflammatory properties and are shown tohave therapeutic effects on inflammatory diseases. The helminth Fasciola hepatica has been reported to suppressantigen-specific Th1 responses in concurrent bacterial infections, thus demonstrating its anti-inflammatory abilityin vivo. Here, F. hepatica tegumental antigen (Teg) was shown to significantly suppress serum levels of gammainterferon (IFN-�) and interleukin-12p70 (IL-12p70) in a model of septic shock. Since dendritic cells (DCs) are agood source of IL-12p70 and critical in driving adaptive immunity, we investigated the effects of F. hepatica Teg onthe activation and function of murine DCs. While Teg alone did not induce cytokine production or cell surfacemarker expression on DCs, it significantly suppressed cytokine production (IL-12p70, IL-6, IL-10, tumor necrosisfactor alpha, and nitrite) and cell surface marker expression (CD80, CD86, and CD40) in DCs matured with a rangeof Toll-like receptor (TLR) and non-TLR ligands. Teg works independently of the TLR4 pathway, since it stillfunctioned in DCs generated from TLR4 mutant and knockout mice. It impaired DC function by inhibiting theirphagocytic capacity and their ability to prime T cells. It does not appear to target the common components(extracellular signal-regulated kinase, Jun N-terminal protein kinase, or p38) of the TLR pathways; however, itsuppressed the active p65 subunit of the transcription factor NF-�B in mature DCs, which could explain theimpairment of proinflammatory cytokine production. Overall, our results demonstrate the potent anti-inflamma-tory properties of F. hepatica Teg and its therapeutic potential as an anti-inflammatory agent.
Dendritic cells (DCs) are highly specialized antigen-present-ing cells, important in stimulating the primary immune re-sponse and driving inflammatory responses associated withimmunopathology. Immature DCs reside in peripheral tissueswhere they are well situated to recognize foreign pathogensthrough Toll-like receptors (TLRs)—pattern recognition re-ceptors which directly recognize conserved microbial mole-cules known as pathogen-associated molecular patterns (30).To date, 13 known mammalian TLRs have been characterized,10 in humans and 13 in mice, with Toll-like receptor 1 (TLR1)to TLR9 conserved in humans and mice (27). Each receptorcan recognize a panel of endogenous and exogenous stimulithat initiates the “classical” maturation of DCs. Lipopolysac-charide (LPS), which binds and signals through TLR4, is themost characterized TLR ligand and has shaped much of ourknowledge of DC biology. LPS activation of TLR4 triggers asignaling cascade including mitogen-activated protein kinases(MAPKs) and NF-�B, culminating in the production of proin-flammatory cytokines and the differentiation of Th1 cells (27).The activation of these signaling cascades is common to mostTLR pathways. This activation process is also accompanied byenhanced expression of costimulatory molecules, such asCD40, CD80, and CD86, as well as major histocompatibilitycomplex (MHC) class II (4). TLR activation is associated with
inflammatory disorders, such as inflammatory bowel diseaseand septic shock (1); thus, mechanisms of suppressing TLRmaturation of DCs may lead to the development of potentialtherapeutics (11, 31).
Parasitic worms (helminths) classically induce highly polar-ized Th2 and T-regulatory immune responses, correlating withchronic infection but with little associated immune pathology(35). Controlling immune pathology, in particular inflamma-tory responses, prolongs the worm’s survival within the hostand thus increases its chances of completing the life cycle. Incontrast to bacteria and viruses, little is known about howhelminths induce DC maturation; however, studies demon-strate that they differ predominantly by their DC activationstatus which lacks many of the classical maturation markers,such as costimulatory molecules (CD80, CD86, MHC class II[MHC-II], and CD40) and proinflammatory cytokines (inter-leukin-12p70 [IL-12p70] and tumor necrosis factor alpha[TNF-�]) (33). Although helminth-matured DCs are classed asbeing less phenotypically mature, they are still capable of prim-ing naïve T cells to activate Th2/tolerogenic T cells associatedwith helminth infection (28, 52).
Helminths have also evolved means of evading or suppress-ing inflammatory responses (17, 34), rendering a host moresusceptible to secondary bystander infections, such as malaria,tuberculosis, and human immunodeficiency virus infection,that require Th1 immunity for protection (6, 14, 40). In linewith this, helminth antigens have been shown to suppress DCmaturation and function when stimulated with TLR ligands.For example, the filarial nematode-secreted product, ES-62,has been shown to significantly suppress LPS-induced produc-
* Corresponding author. Mailing address: Parasite Immune Modu-lation Group, School of Nursing, Faculty of Science and Health, Dub-lin City University, Glasnevin, Dublin 9, Ireland. Phone: 353-1-700-5455. Fax: 353-1-700-7919. E-mail: [email protected].
� Published ahead of print on 30 March 2009.
2488
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

tion of IL-12p40 and IL-12p70 from DCs and IL-12p40, IL-12p70, TNF-�, and IL-6 production from macrophages (20).Similarly, Schistosoma mansoni egg antigen significantly sup-pressed TLR ligand-induced production of IL-12p40 andIL-12p70 by DCs in vitro, while simultaneously enhancingLPS-induced IL-10 production (9, 29). With such potent sup-pression of proinflammatory responses by DCs, helminth anti-gens have been hypothesized to antagonize the Th1 immunityrequired to control intracellular pathogens (5), as well as ame-liorate certain proinflammatory autoimmune diseases (15, 16,32, 49).
Fasciola hepatica (liver fluke) is a parasitic helminth and thecausative agent of fasciolosis, an emerging important disease inhumans (36). F. hepatica induces a dominant Th2/T-regulatorytype immune response (10, 18) and suppresses Th1 responsesto bystander infections, such as Bordetella pertussis and Myco-bacterium tuberculosis during coinfection (7, 19, 44). Fasciola-derived antigens have been shown to mimic this effect by sup-pressing antigen-specific Th1 immune responses both in vitroand in vivo (45). They also modulate the functions of innateimmune cells, inducing alternative activation of macrophageswhich are thought to inhibit the production of gamma inter-feron (IFN-�) by anti-CD3-stimulated naïve CD4� T cells(12). However, the effect of Fasciola antigen on DC functionhas yet to be determined. In this study, we have isolated thetegumental coat from F. hepatica and are the first to investigatethe effects of F. hepatica tegumental antigen on DC maturationand function.
MATERIALS AND METHODS
Animals and antigens. C57BL/6j and BALB/c mice, 6 to 8 weeks old, werepurchased from Harlan UK Ltd. (Bicester, United Kingdom) and DO11.10 TCRtransgenic mice (on a BALB/c background) were either a kind gift from PadraicFallon (Trinity College Dublin, Ireland) or purchased from Jackson Laboratory(Bar Harbor, ME). All mice were maintained according to the Irish Departmentof Health guidelines. C3H/HeJ and C3H/HeN bone marrow cells were a giftfrom Christine Loscher (Dublin City University, Ireland) and TLR4�/� (on aC57BL/6j background) bone marrow cells were a gift from Padraic Fallon. F.hepatica tegumental antigen (Teg) was prepared by adapting a previously pub-lished method (26). In brief, F. hepatica adult worms were washed in sterilephosphate-buffered saline (PBS) and incubated in 1% Nonidet P-40 (NP-40[Sigma Aldrich] in PBS) for 30 min, and supernatant was collected. NP-40 wasremoved using Extracti-Gel D detergent-removing gel (Pierce), and the remain-ing supernatant was centrifuged at 14,000 � g for 30 min at 4°C prior to beingfiltered/concentrated using compressed air, and then stored at �20°C. Proteinconcentrations were measured using the bicinchoninic acid protein assay kit(Pierce), and endotoxin levels were assessed using the Pyrogene endotoxin de-tection system (Cambrex) which utilizes recombinant factor C, an endotoxin-sensitive protein. F. hepatica Teg gave endotoxin levels similar to backgroundlevels and to complete RPMI 1640 medium (supplemented with 5% heat-inac-tivated fetal calf serum, 100 U/ml penicillin, 100 �g/ml streptomycin, 2 mML-glutamine, and 50 �M 2-mercaptoethanol), so the medium was taken to beendotoxin-free.
Septic shock. Groups of four C57BL/6j mice were injected intraperitoneally(i.p.) with F. hepatica Teg (15 �g for each mouse), either 2.5 h before or after i.p.injection with LPS (Escherichia coli 0111:B4, Alexis; 1 �g for each mouse).Control mice received PBS, F. hepatica Teg, or LPS only (i.p.). Mice weresacrificed by cervical dislocation 6 h later, and blood samples were taken bycardiac puncture. Serum concentrations of IFN-�, IL-12p40, and IL-12p70 weremeasured by enzyme-linked immunosorbent assays (ELISAs) (BD OptEIAELISA sets; BD Biosciences).
Isolation and maturation of bone marrow-derived DCs. Bone marrow-derivedimmature DCs were prepared by culturing bone marrow cells, isolated from thefemurs and tibia of mice, in complete RPMI 1640 medium with recombinantmouse granulocyte-macrophage colony-stimulating factor (GM-CSF) (20 ng/ml;R&D Systems Ltd., United Kingdom) at 37°C. On days 3 and 6 of culture, fresh
medium with GM-CSF (20 ng/ml) was added to the cells. On day 8, cells wereharvested, counted, and used for experimental assays.
DCs were seeded into 24-well plates (Nunc) at 106/ml in complete RPMI 1640medium plus 5 ng/ml GM-CSF. The cells were treated with F. hepatica Teg (15�g/ml) for 2.5 h prior to stimulation with LPS (Escherichia coli 0111:B4; Alexisand Sigma Aldrich; 100 ng/ml), zymosan A (Saccharomyces cerevisiae; 5 �g/ml),peptidoglycan (PGN) (Staphylococcus aureus peptidoglycan; 5 �g/ml), poly(I:C)(polyinosinic acid � polycytidylic acid, a synthetic analogue of double-strandedRNA; 2.5 �g/ml), flagellin (Salmonella enterica serovar Typhimurium; 0.5 �g/ml), CpG (stimulatory oligonucleotide 1826 [ODN1826]; 5 �g/ml) (all fromInvivoGen). or phorbol myristate acetate (PMA; Sigma Aldrich; 50 ng/ml) for18 h. In time course experiments, DCs were treated with F. hepatica Teg 2.5 hbefore, at the same time as, or 2.5 h after LPS stimulation. Control cells weretreated with medium, Teg, or TLR ligand alone. In the MAPK inhibitor studies,cells were incubated with extracellular signal-regulated kinase (ERK) (U0126;Sigma Aldrich), p38 (SB 202190; Sigma Aldrich), or Jun N-terminal proteinkinase (JNK) inhibitors [JNK inhibitor I (L)-form; Calbiochem] (all 5 �M) 1hour prior to TLR ligand stimulation. Control cells were treated with medium,Teg, or LPS alone and had inhibitors added.
Characterization of matured DCs. Expression of cell surface markers on DCswas quantified by three-color flow cytometry using phycoerythrin-, fluoresceinisothiocyanate (FITC)-, or phycoerythrin-Cy5.5-conjugated antibodies specificfor CD80, CD86, CD40, MHC-II (BD Biosciences), and CD11c (Caltag Labo-ratories). Appropriately labeled isotype-matched antibodies were used as con-trols. DC purity was �90% positive for expression of CD11c. Acquisition wasperformed using a FACSCalibur flow cytometer (BD Biosciences), and analysisof results was performed using FlowJo software (Tree Star). Supernatants fromcultured DCs were tested for the production of IL-12p70, IL-10, TNF-�, IL-12p40, IL-6 (BD OptEIA ELISA sets; BD Biosciences), and IL-23 (eBioscienceELISA set) by ELISAs, and nitrite was measured using Griess reagent (Promega,Madison, WI).
RNA extraction and RT-PCR. RNA was recovered from cultured cells by thedirect addition of Tri-reagent (Sigma Aldrich) to the wells following aspiration ofsupernatants. Total cellular RNA was subsequently extracted from the prepara-tion according to the manufacturers’ specifications. For reverse transcription-PCR (RT-PCR), first-strand cDNA was produced with oligo(dT) primers from 1�g total RNA using avian myeloblastosis virus reverse transcriptase (reversetranscription system; Promega) at 42°C for 15 min, followed by 95°C for 5 minand 4°C for 5 min. A 1-�l aliquot of the resultant cDNA was amplified usingprimers specific for IL-12p35, IL-12p40, and -actin (42) under the followingconditions: 35 cycles, with 1 cycle consisting of 45 s of denaturation at 95°C, 45 sof annealing of primers at 56°C (-actin and p40) or 51°C (p35), and 60 s ofelongation at 72°C; followed by a step of 10 min at 72°C. All PCR products wereelectrophoresed on 1% agarose gel and visualized by ethidium bromide staining.
Cell viability and apoptosis detection. Cell viability and growth during exper-imental conditions was first assessed by seeding DCs into 96-well plates (5 � 104
cells/ml and 1 � 106 cells/ml, respectively) and treating the cells with F. hepaticaTeg (15 �g/ml) for 2.5 h prior to stimulation with LPS for 18 h. Control cells weretreated with medium, Teg, or LPS alone. Following incubation, cell viability wasassessed by adding 40 �l MTS solution [3-(4,5-dimethylthiazol-2-yl)-5-(3-car-boxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS), plus phenazineethosulfate] to each well. Following an incubation of 3 h at 37°C, the absorbanceof each well was read at 450 nm. Cell growth was assessed by adding a volume ofMTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide] solutionequivalent to 10% of culture medium to each well and incubating for 3 h at 37°C.The culture medium was then removed, the resulting MTT formazan crystalswere dissolved by adding MTT solvent (equal to the original culture volume),and absorbance was read at 562 nm.
Cell viability was also measured using the annexin V-FITC apoptosis detectionkit I (BD Biosciences). Briefly, 1 � 106 cells/ml were seeded into a 24-well plateand treated with F. hepatica Teg (15 �g/ml) and each TLR ligand, either sepa-rately or together, or medium alone. Cells were harvested 24 h later, washedtwice in cold PBS and incubated in binding buffer (0.1 M HEPES-NaOH, 1.4 MNaCl, 25 mM CaCl2) with annexin V-FITC and propidium iodide for 15 min.Following the addition of more binding buffer, cells were immediately analyzedby flow cytometry.
TLR screening. F. hepatica Teg-induced stimulation of TLR2 to TLR9 wastested in HEK-293 cells expressing a given TLR protein as well as a reportergene driven by the NF-�B promoter (InvivoGen). Each 293-TLR cell line wasinduced with a known specific ligand as a positive control, and a recombinantHEK-293 cell line for the reporter gene only was used as a negative control.
Protein extraction and Western blot analysis. Total protein was extractedfrom cell lysates using radioimmunoprecipitation assay (RIPA) buffer containing
VOL. 77, 2009 F. HEPATICA Teg SUPPRESSES DC MATURATION AND FUNCTION 2489
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1.0% NP-40, 0.5% sodium deoxy-cholate, 0.1% sodium dodecyl sulfate, and protease and phosphatase inhibitorcocktails (Sigma Aldrich). Cells were incubated in the extraction buffer on ice for5 min before being centrifuged at 8,000 � g for 10 min at 4°C. Supernatants weretransferred to clean tubes, and protein concentrations were determined using thebicinchoninic acid protein assay kit (Pierce). Protein samples (10 �g) andprestained protein markers (Precision Plus protein standards; Bio-Rad) wereseparated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis andblotted onto 0.45-�m Immobilon-P polyvinylidene difluoride membrane (SigmaAldrich). Membranes were blocked for 1 h at room temperature in 5% nonfatdried milk in PBS and incubated overnight at 4°C with anti-NF-�Bp65 (CellSignaling Technology; 1:1,000). Membranes were washed in PBS with 0.05%Tween 20 and incubated for 1 h at room temperature with peroxide-conjugatedanti-rabbit immunoglobulin G (IgG) (Sigma Aldrich; 1:2,000). After furtherwashing, proteins were visualized with supersignal (Pierce), exposed to film for10 to 30 min, and processed using an FPM 100A processor (Fujifilm). Proteinbands were quantified using the GeneSnap acquisition and GeneTools analysissoftware (GeneGenius gel documentation and analysis system; Syngene).
Phagocytosis assay. The phagocytic ability of DCs was measured using theCytoSelect 96-well phagocytosis assay (Cell Biolabs Inc.). Briefly, DCs fromC57BL/6 mice were plated at 0.5 � 106/ml in complete RPMI 1640 medium andincubated overnight at 37°C to allow the cells to adhere to the plate. DCs werethen treated with F. hepatica Teg (15 �g/ml), LPS (100 ng/ml), or zymosan A (5�g/ml) alone or with Teg and LPS or Teg and zymosan at the same time for 2.5 hbefore the addition of sheep erythrocytes opsonized by IgG at a ratio of 50:1.Supernatants were aspirated after 1 h, and adherent cells were washed withsterile PBS to remove nonphagocytosed erythrocytes. Adherent DCs were thenlysed, substrate solution was added, and the amount of engulfed erythrocytes wasdetermined by colorimetric assay using an absorbance of 610 nm. Negative-control cells were treated with 2 �M cytochalasin D to block phagocytosis.
In vivo T-cell assays, cytokine measurements, and proliferation. For assays ofin vivo T-cell priming, DCs isolated from DO11.10 mice were stimulated withmedium or F. hepatica Teg (15 �g/ml) in the presence of ovalbumin (OVA)peptide (323-ISQAVHAAHAEINEAGR-329; 100 nM; GenScript Corp.) for24 h. After the DCs were washed with sterile endotoxin-free PBS, the cells (3 �105) were delivered into the sternum of naïve DO11.10 mice by subcutaneousinjection. After 7 days, skin draining lymph nodes (sdLNs) were removed and asingle-cell suspension of cells was plated with medium and OVA peptide (500nM) or with PMA (25 ng/ml; Sigma Aldrich) and anti-CD3 monoclonal antibody(1 �g/ml; clone 145-2C11; BD Biosciences). After 72 h, supernatants wereremoved for measurement of IL-4, IFN-�, and IL-10 by commercial assay (R&DSystems). Lymph node cells as described above were also plated on a 96-wellplate, and after 72 h, 1 �Ci of [3H]thymidine was added per well. After 4 hours,cells were harvested onto filter plates, and [3H]thymidine uptake was determinedvia a liquid scintillation counter. Flow cytometric analysis of CD4, CD28, andcytotoxic-T-lymphocyte-associated antigen 4 (CTLA-4) (BD Biosciences) ex-pression on sdLN cells was performed as described above.
Statistics. All data were analyzed for normality prior to statistical testing.When comparisons of the values for multiple groups were made, data wereanalyzed using one-way analysis of variance. For comparisons of values for twogroups, the Student’s t test was used. For all tests, a P value of 0.05 was deemedsignificant.
RESULTS
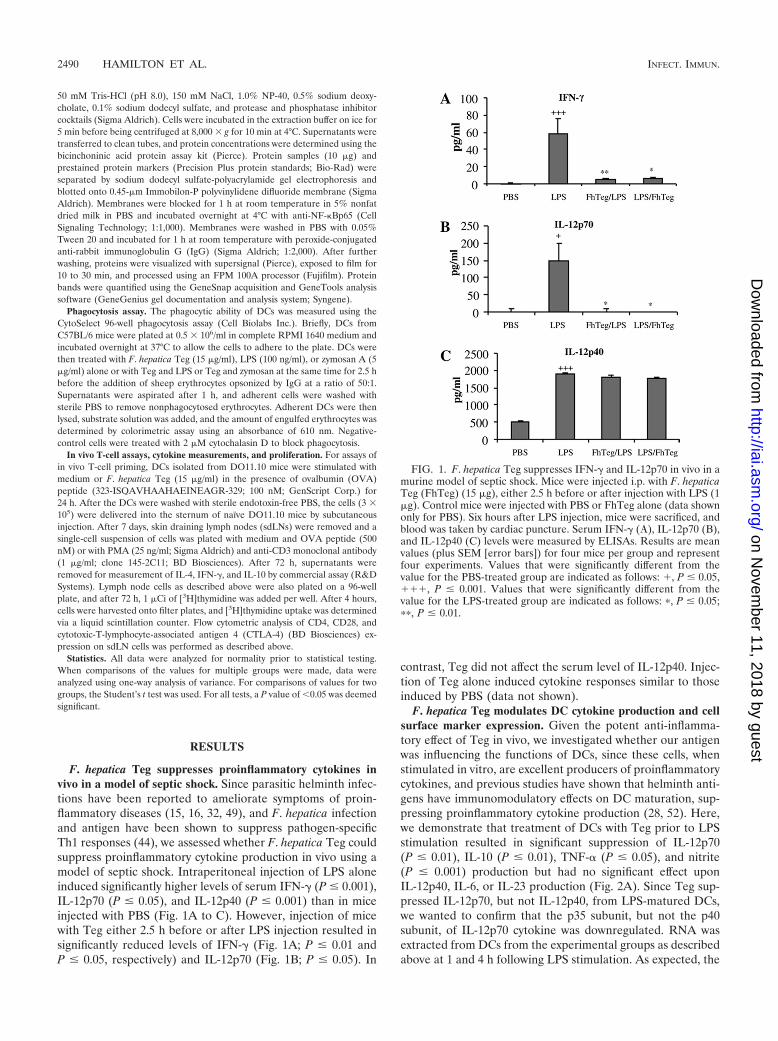
F. hepatica Teg suppresses proinflammatory cytokines invivo in a model of septic shock. Since parasitic helminth infec-tions have been reported to ameliorate symptoms of proin-flammatory diseases (15, 16, 32, 49), and F. hepatica infectionand antigen have been shown to suppress pathogen-specificTh1 responses (44), we assessed whether F. hepatica Teg couldsuppress proinflammatory cytokine production in vivo using amodel of septic shock. Intraperitoneal injection of LPS aloneinduced significantly higher levels of serum IFN-� (P � 0.001),IL-12p70 (P � 0.05), and IL-12p40 (P � 0.001) than in miceinjected with PBS (Fig. 1A to C). However, injection of micewith Teg either 2.5 h before or after LPS injection resulted insignificantly reduced levels of IFN-� (Fig. 1A; P � 0.01 andP � 0.05, respectively) and IL-12p70 (Fig. 1B; P � 0.05). In
contrast, Teg did not affect the serum level of IL-12p40. Injec-tion of Teg alone induced cytokine responses similar to thoseinduced by PBS (data not shown).
F. hepatica Teg modulates DC cytokine production and cellsurface marker expression. Given the potent anti-inflamma-tory effect of Teg in vivo, we investigated whether our antigenwas influencing the functions of DCs, since these cells, whenstimulated in vitro, are excellent producers of proinflammatorycytokines, and previous studies have shown that helminth anti-gens have immunomodulatory effects on DC maturation, sup-pressing proinflammatory cytokine production (28, 52). Here,we demonstrate that treatment of DCs with Teg prior to LPSstimulation resulted in significant suppression of IL-12p70(P � 0.01), IL-10 (P � 0.01), TNF-� (P � 0.05), and nitrite(P � 0.001) production but had no significant effect uponIL-12p40, IL-6, or IL-23 production (Fig. 2A). Since Teg sup-pressed IL-12p70, but not IL-12p40, from LPS-matured DCs,we wanted to confirm that the p35 subunit, but not the p40subunit, of IL-12p70 cytokine was downregulated. RNA wasextracted from DCs from the experimental groups as describedabove at 1 and 4 h following LPS stimulation. As expected, the
FIG. 1. F. hepatica Teg suppresses IFN-� and IL-12p70 in vivo in amurine model of septic shock. Mice were injected i.p. with F. hepaticaTeg (FhTeg) (15 �g), either 2.5 h before or after injection with LPS (1�g). Control mice were injected with PBS or FhTeg alone (data shownonly for PBS). Six hours after LPS injection, mice were sacrificed, andblood was taken by cardiac puncture. Serum IFN-� (�), IL-12p70 (B),and IL-12p40 (C) levels were measured by ELISAs. Results are meanvalues (plus SEM [error bars]) for four mice per group and representfour experiments. Values that were significantly different from thevalue for the PBS-treated group are indicated as follows: �, P � 0.05,���, P � 0.001. Values that were significantly different from thevalue for the LPS-treated group are indicated as follows: �, P � 0.05;��, P � 0.01.
2490 HAMILTON ET AL. INFECT. IMMUN.
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
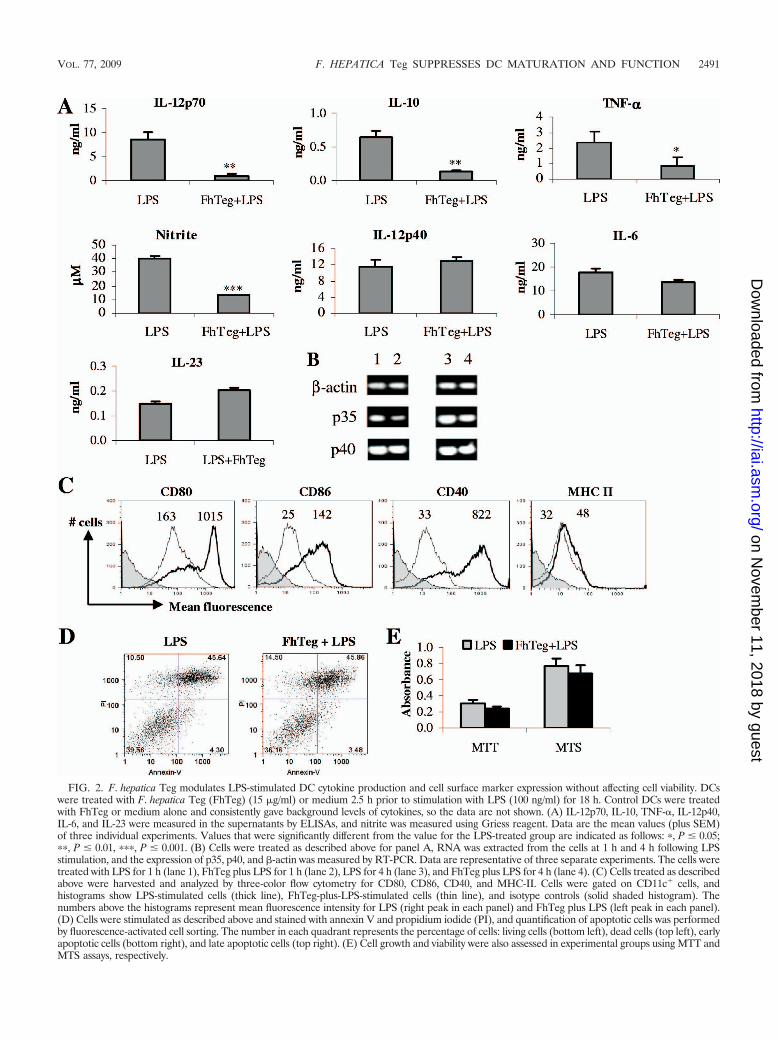
FIG. 2. F. hepatica Teg modulates LPS-stimulated DC cytokine production and cell surface marker expression without affecting cell viability. DCswere treated with F. hepatica Teg (FhTeg) (15 �g/ml) or medium 2.5 h prior to stimulation with LPS (100 ng/ml) for 18 h. Control DCs were treatedwith FhTeg or medium alone and consistently gave background levels of cytokines, so the data are not shown. (A) IL-12p70, IL-10, TNF-�, IL-12p40,IL-6, and IL-23 were measured in the supernatants by ELISAs, and nitrite was measured using Griess reagent. Data are the mean values (plus SEM)of three individual experiments. Values that were significantly different from the value for the LPS-treated group are indicated as follows: �, P � 0.05;��, P � 0.01, ���, P � 0.001. (B) Cells were treated as described above for panel A, RNA was extracted from the cells at 1 h and 4 h following LPSstimulation, and the expression of p35, p40, and -actin was measured by RT-PCR. Data are representative of three separate experiments. The cells weretreated with LPS for 1 h (lane 1), FhTeg plus LPS for 1 h (lane 2), LPS for 4 h (lane 3), and FhTeg plus LPS for 4 h (lane 4). (C) Cells treated as describedabove were harvested and analyzed by three-color flow cytometry for CD80, CD86, CD40, and MHC-II. Cells were gated on CD11c� cells, andhistograms show LPS-stimulated cells (thick line), FhTeg-plus-LPS-stimulated cells (thin line), and isotype controls (solid shaded histogram). Thenumbers above the histograms represent mean fluorescence intensity for LPS (right peak in each panel) and FhTeg plus LPS (left peak in each panel).(D) Cells were stimulated as described above and stained with annexin V and propidium iodide (PI), and quantification of apoptotic cells was performedby fluorescence-activated cell sorting. The number in each quadrant represents the percentage of cells: living cells (bottom left), dead cells (top left), earlyapoptotic cells (bottom right), and late apoptotic cells (top right). (E) Cell growth and viability were also assessed in experimental groups using MTT andMTS assays, respectively.
VOL. 77, 2009 F. HEPATICA Teg SUPPRESSES DC MATURATION AND FUNCTION 2491
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

expression of p35, but not p40, was suppressed in DCs in thepresence of Teg (Fig. 2B). Both medium and Teg alone wereincluded as controls and gave similar levels of expression (datanot shown).
Stimulation of DCs with LPS resulted in a marked increaseof CD80, CD86, and CD40 expression (Fig. 2C), which istypical of LPS-induced DC maturation, and a slight increase inMHC-II expression. Treatment of DCs with F. hepatica Tegprior to stimulation with LPS resulted in a dramatic decreaseof CD80, CD86, and CD40 expression and a slight decrease inMHC-II expression. DCs treated with medium or Teg aloneconsistently gave very low to undetectable background levels ofcytokines and comparable levels of cell surface marker expres-sion, so these data are not shown.
With such potent inhibitory effects on DC maturation, wemeasured the influence of F. hepatica Teg on cell viability usingan annexin V assay. Cells incubated with LPS and Teg plusLPS exhibited similar profiles of live, early apoptotic, and lateapoptotic cells (Fig. 2D), indicating that the difference in cy-tokine and surface marker expression between the experimen-tal groups may not be attributed to cell death. This was furtherconfirmed with the MTT and MTS assays, which demonstratedthat treatment with Teg did not inhibit cell growth or viability(Fig. 2E). Treatment of DCs with medium or Teg alone gavesimilar results in all experiments (data not shown).
Suppressive effects of F. hepatica Teg are not influenced bythe time of exposure to DCs and are not TLR4 dependent. Toinvestigate whether the suppressive effects of F. hepatica Tegwere due to competition with LPS for TLR4 binding, we in-
cubated DCs with Teg 2.5 h prior to, at the same time as, or2.5 h after LPS stimulation, and the effects on cytokine pro-duction and cell surface marker expression were examined.The timing of exposure made no difference to the suppressionof IL-12p70, IL-10, and nitrite (Fig. 3A), with levels beingsignificantly reduced at all time points (P � 0.001), thus sug-gesting that it is unlikely that Teg is interfering with LPSbinding. The timing of exposure also made no difference to thesuppression of TNF-� and did not alter the lack of suppressionof IL-6, IL-12p40, or IL-23 (data not shown). There was alsono difference in the suppression of CD80, CD86, and CD40expression (data not shown).
To demonstrate that F. hepatica Teg does not exert its sup-pressive effects through TLR4, the experiment was repeated inDCs from C3H/HeJ and C3H/HeN mice and TLR4 knockout(TLR4�/�) mice. C3H/HeJ mice exhibit a point mutation inthe TIR domain of TLR4 (47), and although they express thereceptor, they fail to produce a response to LPS. As suspectedand in support of the timing study, the suppressive effects ofTeg were not lost in DCs from HeJ or TLR4�/� mice (Fig. 3Band C). DCs from control HeN and TLR4�/� mice producedsignificant levels of IL-12p70, IL-10, and nitrite in response toLPS and PGN (TLR2) compared to the levels produced inresponse to medium alone (P � 0.001; medium data notshown) and demonstrated significant reductions when treatedwith Teg prior to stimulation (P � 0.001 and P � 0.01, respec-tively). DCs from HeJ and TLR4�/� mice failed to respond toLPS, but the Teg-mediated suppression of PGN-induced IL-12p70, IL-10, and nitrite was still intact. There was also no
FIG. 3. Suppression of LPS-induced DC cytokine production by F. hepatica Teg does not depend upon time of exposure and is not mediatedthrough TLR4. (A) DCs were treated with F. hepatica Teg (15 �g/ml) or medium 2.5 h before (�2.5 h), at the same time as (0 h), or 2.5 h after(�2.5 h) LPS stimulation (100 ng/ml). After 18 h, IL-12p70 and IL-10 were measured in the supernatants by ELISAs, and nitrite was measuredusing Griess reagent. (B and C) DCs from C3H/HeN (control) and C3H/HeJ (TLR4 mutant) mice (B) and DCs from TLR4 knockout mice(TLR4�/�) and controls (TLR4�/�) (C) were treated with F. hepatica Teg (FhTeg) (15 �g/ml) or medium 2.5 h prior to stimulation with LPS (100ng/ml) or PGN (5 �g/ml). IL-12p70, IL-10, and nitrite were measured as described above for panel A. Data are the mean values (plus SEM [errorbars]) of three individual experiments. Values that were significantly different from the value for the LPS- or PGN-treated group are indicated asfollows: ��, P � 0.01; ���, P � 0.001.
2492 HAMILTON ET AL. INFECT. IMMUN.
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
difference in the suppression of PGN-induced CD80, CD86,and CD40 expression (data not shown). These results indicatethat not only does Teg not require TLR4 to exert its suppres-sive effects but it may have a broad suppressive effect on DCfunction, rather than targeting a single TLR pathway.
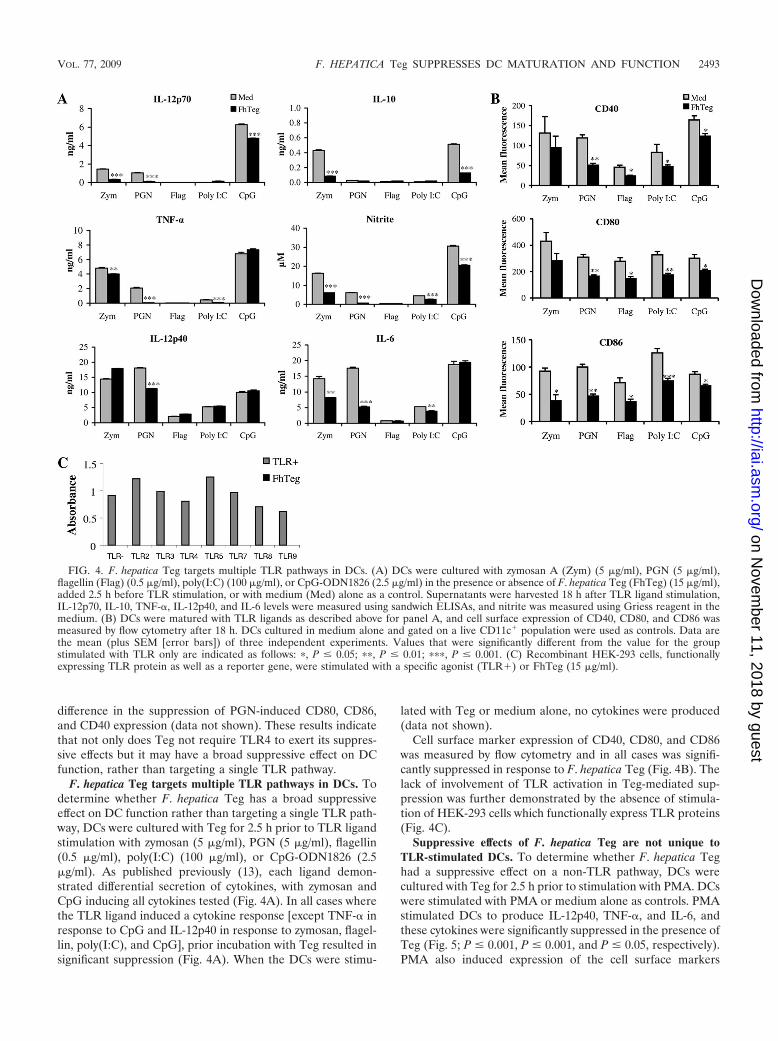
F. hepatica Teg targets multiple TLR pathways in DCs. Todetermine whether F. hepatica Teg has a broad suppressiveeffect on DC function rather than targeting a single TLR path-way, DCs were cultured with Teg for 2.5 h prior to TLR ligandstimulation with zymosan (5 �g/ml), PGN (5 �g/ml), flagellin(0.5 �g/ml), poly(I:C) (100 �g/ml), or CpG-ODN1826 (2.5�g/ml). As published previously (13), each ligand demon-strated differential secretion of cytokines, with zymosan andCpG inducing all cytokines tested (Fig. 4A). In all cases wherethe TLR ligand induced a cytokine response [except TNF-� inresponse to CpG and IL-12p40 in response to zymosan, flagel-lin, poly(I:C), and CpG], prior incubation with Teg resulted insignificant suppression (Fig. 4A). When the DCs were stimu-
lated with Teg or medium alone, no cytokines were produced(data not shown).
Cell surface marker expression of CD40, CD80, and CD86was measured by flow cytometry and in all cases was signifi-cantly suppressed in response to F. hepatica Teg (Fig. 4B). Thelack of involvement of TLR activation in Teg-mediated sup-pression was further demonstrated by the absence of stimula-tion of HEK-293 cells which functionally express TLR proteins(Fig. 4C).
Suppressive effects of F. hepatica Teg are not unique toTLR-stimulated DCs. To determine whether F. hepatica Teghad a suppressive effect on a non-TLR pathway, DCs werecultured with Teg for 2.5 h prior to stimulation with PMA. DCswere stimulated with PMA or medium alone as controls. PMAstimulated DCs to produce IL-12p40, TNF-�, and IL-6, andthese cytokines were significantly suppressed in the presence ofTeg (Fig. 5; P � 0.001, P � 0.001, and P � 0.05, respectively).PMA also induced expression of the cell surface markers
FIG. 4. F. hepatica Teg targets multiple TLR pathways in DCs. (A) DCs were cultured with zymosan A (Zym) (5 �g/ml), PGN (5 �g/ml),flagellin (Flag) (0.5 �g/ml), poly(I:C) (100 �g/ml), or CpG-ODN1826 (2.5 �g/ml) in the presence or absence of F. hepatica Teg (FhTeg) (15 �g/ml),added 2.5 h before TLR stimulation, or with medium (Med) alone as a control. Supernatants were harvested 18 h after TLR ligand stimulation,IL-12p70, IL-10, TNF-�, IL-12p40, and IL-6 levels were measured using sandwich ELISAs, and nitrite was measured using Griess reagent in themedium. (B) DCs were matured with TLR ligands as described above for panel A, and cell surface expression of CD40, CD80, and CD86 wasmeasured by flow cytometry after 18 h. DCs cultured in medium alone and gated on a live CD11c� population were used as controls. Data arethe mean (plus SEM [error bars]) of three independent experiments. Values that were significantly different from the value for the groupstimulated with TLR only are indicated as follows: �, P � 0.05; ��, P � 0.01; ���, P � 0.001. (C) Recombinant HEK-293 cells, functionallyexpressing TLR protein as well as a reporter gene, were stimulated with a specific agonist (TLR�) or FhTeg (15 �g/ml).
VOL. 77, 2009 F. HEPATICA Teg SUPPRESSES DC MATURATION AND FUNCTION 2493
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
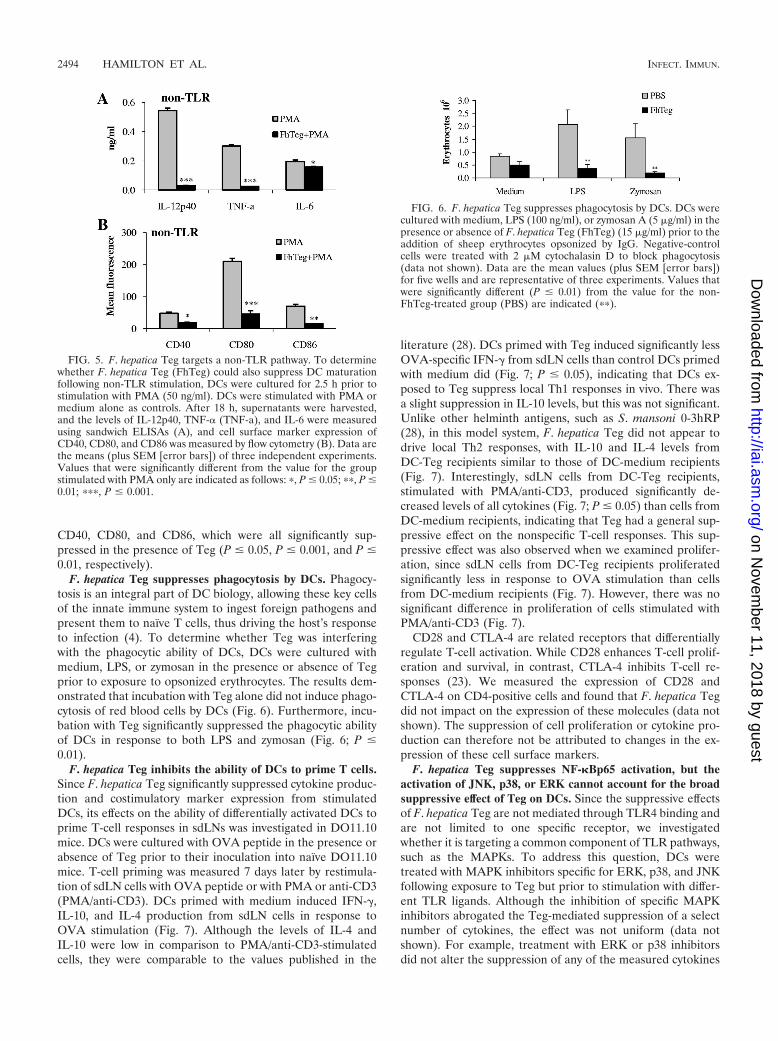
CD40, CD80, and CD86, which were all significantly sup-pressed in the presence of Teg (P � 0.05, P � 0.001, and P �0.01, respectively).
F. hepatica Teg suppresses phagocytosis by DCs. Phagocy-tosis is an integral part of DC biology, allowing these key cellsof the innate immune system to ingest foreign pathogens andpresent them to naïve T cells, thus driving the host’s responseto infection (4). To determine whether Teg was interferingwith the phagocytic ability of DCs, DCs were cultured withmedium, LPS, or zymosan in the presence or absence of Tegprior to exposure to opsonized erythrocytes. The results dem-onstrated that incubation with Teg alone did not induce phago-cytosis of red blood cells by DCs (Fig. 6). Furthermore, incu-bation with Teg significantly suppressed the phagocytic abilityof DCs in response to both LPS and zymosan (Fig. 6; P �0.01).
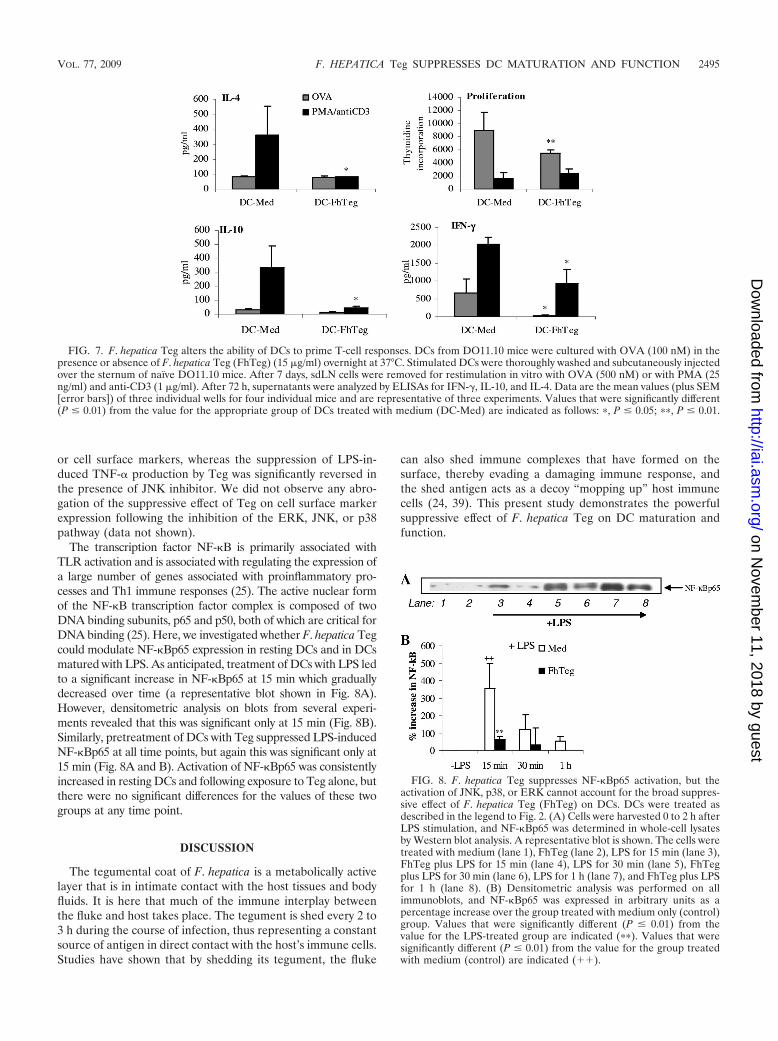
F. hepatica Teg inhibits the ability of DCs to prime T cells.Since F. hepatica Teg significantly suppressed cytokine produc-tion and costimulatory marker expression from stimulatedDCs, its effects on the ability of differentially activated DCs toprime T-cell responses in sdLNs was investigated in DO11.10mice. DCs were cultured with OVA peptide in the presence orabsence of Teg prior to their inoculation into naïve DO11.10mice. T-cell priming was measured 7 days later by restimula-tion of sdLN cells with OVA peptide or with PMA or anti-CD3(PMA/anti-CD3). DCs primed with medium induced IFN-�,IL-10, and IL-4 production from sdLN cells in response toOVA stimulation (Fig. 7). Although the levels of IL-4 andIL-10 were low in comparison to PMA/anti-CD3-stimulatedcells, they were comparable to the values published in the
literature (28). DCs primed with Teg induced significantly lessOVA-specific IFN-� from sdLN cells than control DCs primedwith medium did (Fig. 7; P � 0.05), indicating that DCs ex-posed to Teg suppress local Th1 responses in vivo. There wasa slight suppression in IL-10 levels, but this was not significant.Unlike other helminth antigens, such as S. mansoni 0-3hRP(28), in this model system, F. hepatica Teg did not appear todrive local Th2 responses, with IL-10 and IL-4 levels fromDC-Teg recipients similar to those of DC-medium recipients(Fig. 7). Interestingly, sdLN cells from DC-Teg recipients,stimulated with PMA/anti-CD3, produced significantly de-creased levels of all cytokines (Fig. 7; P � 0.05) than cells fromDC-medium recipients, indicating that Teg had a general sup-pressive effect on the nonspecific T-cell responses. This sup-pressive effect was also observed when we examined prolifer-ation, since sdLN cells from DC-Teg recipients proliferatedsignificantly less in response to OVA stimulation than cellsfrom DC-medium recipients (Fig. 7). However, there was nosignificant difference in proliferation of cells stimulated withPMA/anti-CD3 (Fig. 7).
CD28 and CTLA-4 are related receptors that differentiallyregulate T-cell activation. While CD28 enhances T-cell prolif-eration and survival, in contrast, CTLA-4 inhibits T-cell re-sponses (23). We measured the expression of CD28 andCTLA-4 on CD4-positive cells and found that F. hepatica Tegdid not impact on the expression of these molecules (data notshown). The suppression of cell proliferation or cytokine pro-duction can therefore not be attributed to changes in the ex-pression of these cell surface markers.
F. hepatica Teg suppresses NF-�Bp65 activation, but theactivation of JNK, p38, or ERK cannot account for the broadsuppressive effect of Teg on DCs. Since the suppressive effectsof F. hepatica Teg are not mediated through TLR4 binding andare not limited to one specific receptor, we investigatedwhether it is targeting a common component of TLR pathways,such as the MAPKs. To address this question, DCs weretreated with MAPK inhibitors specific for ERK, p38, and JNKfollowing exposure to Teg but prior to stimulation with differ-ent TLR ligands. Although the inhibition of specific MAPKinhibitors abrogated the Teg-mediated suppression of a selectnumber of cytokines, the effect was not uniform (data notshown). For example, treatment with ERK or p38 inhibitorsdid not alter the suppression of any of the measured cytokines
FIG. 5. F. hepatica Teg targets a non-TLR pathway. To determinewhether F. hepatica Teg (FhTeg) could also suppress DC maturationfollowing non-TLR stimulation, DCs were cultured for 2.5 h prior tostimulation with PMA (50 ng/ml). DCs were stimulated with PMA ormedium alone as controls. After 18 h, supernatants were harvested,and the levels of IL-12p40, TNF-� (TNF-a), and IL-6 were measuredusing sandwich ELISAs (A), and cell surface marker expression ofCD40, CD80, and CD86 was measured by flow cytometry (B). Data arethe means (plus SEM [error bars]) of three independent experiments.Values that were significantly different from the value for the groupstimulated with PMA only are indicated as follows: �, P � 0.05; ��, P �0.01; ���, P � 0.001.
FIG. 6. F. hepatica Teg suppresses phagocytosis by DCs. DCs werecultured with medium, LPS (100 ng/ml), or zymosan A (5 �g/ml) in thepresence or absence of F. hepatica Teg (FhTeg) (15 �g/ml) prior to theaddition of sheep erythrocytes opsonized by IgG. Negative-controlcells were treated with 2 �M cytochalasin D to block phagocytosis(data not shown). Data are the mean values (plus SEM [error bars])for five wells and are representative of three experiments. Values thatwere significantly different (P � 0.01) from the value for the non-FhTeg-treated group (PBS) are indicated (��).
2494 HAMILTON ET AL. INFECT. IMMUN.
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
or cell surface markers, whereas the suppression of LPS-in-duced TNF-� production by Teg was significantly reversed inthe presence of JNK inhibitor. We did not observe any abro-gation of the suppressive effect of Teg on cell surface markerexpression following the inhibition of the ERK, JNK, or p38pathway (data not shown).
The transcription factor NF-�B is primarily associated withTLR activation and is associated with regulating the expression ofa large number of genes associated with proinflammatory pro-cesses and Th1 immune responses (25). The active nuclear formof the NF-�B transcription factor complex is composed of twoDNA binding subunits, p65 and p50, both of which are critical forDNA binding (25). Here, we investigated whether F. hepatica Tegcould modulate NF-�Bp65 expression in resting DCs and in DCsmatured with LPS. As anticipated, treatment of DCs with LPS ledto a significant increase in NF-�Bp65 at 15 min which graduallydecreased over time (a representative blot shown in Fig. 8A).However, densitometric analysis on blots from several experi-ments revealed that this was significant only at 15 min (Fig. 8B).Similarly, pretreatment of DCs with Teg suppressed LPS-inducedNF-�Bp65 at all time points, but again this was significant only at15 min (Fig. 8A and B). Activation of NF-�Bp65 was consistentlyincreased in resting DCs and following exposure to Teg alone, butthere were no significant differences for the values of these twogroups at any time point.
DISCUSSION
The tegumental coat of F. hepatica is a metabolically activelayer that is in intimate contact with the host tissues and bodyfluids. It is here that much of the immune interplay betweenthe fluke and host takes place. The tegument is shed every 2 to3 h during the course of infection, thus representing a constantsource of antigen in direct contact with the host’s immune cells.Studies have shown that by shedding its tegument, the fluke
can also shed immune complexes that have formed on thesurface, thereby evading a damaging immune response, andthe shed antigen acts as a decoy “mopping up” host immunecells (24, 39). This present study demonstrates the powerfulsuppressive effect of F. hepatica Teg on DC maturation andfunction.
FIG. 7. F. hepatica Teg alters the ability of DCs to prime T-cell responses. DCs from DO11.10 mice were cultured with OVA (100 nM) in thepresence or absence of F. hepatica Teg (FhTeg) (15 �g/ml) overnight at 37°C. Stimulated DCs were thoroughly washed and subcutaneously injectedover the sternum of naïve DO11.10 mice. After 7 days, sdLN cells were removed for restimulation in vitro with OVA (500 nM) or with PMA (25ng/ml) and anti-CD3 (1 �g/ml). After 72 h, supernatants were analyzed by ELISAs for IFN-�, IL-10, and IL-4. Data are the mean values (plus SEM[error bars]) of three individual wells for four individual mice and are representative of three experiments. Values that were significantly different(P � 0.01) from the value for the appropriate group of DCs treated with medium (DC-Med) are indicated as follows: �, P � 0.05; ��, P � 0.01.
FIG. 8. F. hepatica Teg suppresses NF-�Bp65 activation, but theactivation of JNK, p38, or ERK cannot account for the broad suppres-sive effect of F. hepatica Teg (FhTeg) on DCs. DCs were treated asdescribed in the legend to Fig. 2. (A) Cells were harvested 0 to 2 h afterLPS stimulation, and NF-�Bp65 was determined in whole-cell lysatesby Western blot analysis. A representative blot is shown. The cells weretreated with medium (lane 1), FhTeg (lane 2), LPS for 15 min (lane 3),FhTeg plus LPS for 15 min (lane 4), LPS for 30 min (lane 5), FhTegplus LPS for 30 min (lane 6), LPS for 1 h (lane 7), and FhTeg plus LPSfor 1 h (lane 8). (B) Densitometric analysis was performed on allimmunoblots, and NF-�Bp65 was expressed in arbitrary units as apercentage increase over the group treated with medium only (control)group. Values that were significantly different (P � 0.01) from thevalue for the LPS-treated group are indicated (��). Values that weresignificantly different (P � 0.01) from the value for the group treatedwith medium (control) are indicated (��).
VOL. 77, 2009 F. HEPATICA Teg SUPPRESSES DC MATURATION AND FUNCTION 2495
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

In initial experiments, the modulatory role of F. hepatica Tegin vivo in a model of septic shock was examined. Mice injectedwith F. hepatica Teg either prior to or following LPS exposuredemonstrated significantly suppressed levels of serum IL-12p70 and IFN-�, but not IL-12p40. The suppression of proin-flammatory cytokines in vivo carries implications for the con-trol of concurrent infections with F. hepatica, as studies haveshown that it suppresses IFN-�, altering the immune responseto tuberculosis (45) and delaying bacterial clearance in con-current infections with Salmonella enterica serovar Dublin orBordetella pertussis (2, 7). Since DCs are key players in drivingadaptive immunity, we went on to examine the effects of Tegon these cells.
First, we investigated the effect of F. hepatica Teg on DCmaturation alone, and our results supported other studieswhich demonstrate that exposure of immature DCs to hel-minth-derived antigens failed to induce DC maturation, asthese cells did not secrete cytokines or increase costimulatorymarker expression in response to Teg (3, 46). However, otherstudies have shown that helminth-derived antigens, such as thefilaria-derived antigen, ES-62, which induces IL-12p40, IL-6,and TNF-� production from DCs, can partially induce DCmaturation (21) and Schistosoma mansoni larva antigen, whichcan induce low, yet significant, increases in IL-12p40 and IL-6from DCs (28). Despite the inactivation or partial activation ofDCs, many helminth-matured DCs are still capable of drivingpotent Th2 immune responses (28, 52). We have yet to deter-mine whether Teg drives Th2 or T-regulatory immune re-sponses.
Although F. hepatica Teg failed to induce cytokine produc-tion and cell surface marker expression, treatment of DCs withTeg rendered them hyporesponsive to a range of TLR ligandswith significant decreases in cytokine production (IL-12p70,IL-10, IL-6, TNF-�, and nitrite) and costimulatory markerexpression (CD80, CD86, and CD40). A decrease in proin-flammatory cytokines is in keeping with a helminth’s need tosuppress inflammatory processes (21), and since IL-12p70 is animportant polarizing cytokine known to drive Th1 differentia-tion (50), this suppression may result in the Th1 suppressionobserved during F. hepatica coinfections. The decrease inIL-10 observed was not anticipated, since this cytokine is tra-ditionally associated with an anti-inflammatory or regulatoryresponse (37). However, recent studies show IL-10 productionby Th1 cells, emphasizing the versatile role that IL-10 playsduring infection (43). Cytokine suppression by Teg was selec-tive, given that in general IL-12p40 and IL-23 production werenot affected. This is of interest given that the p40 subunit isshared by IL-12 and IL-23 and both cytokines belong to thesame family of proinflammatory cytokines (51).
In addition to cytokine production, costimulation is essentialfor the successful activation and differentiation of naïve CD4�
T cells, and similarly, the suppression of costimulatory markerscould influence Th1 cell differentiation. This was confirmed inthe T-cell priming studies, which demonstrated that F. hepaticaTeg-primed DCs significantly suppressed local Th1 immuneresponses. These results indicate that exposure to Teg, and thesubsequent lack of cytokine production and costimulatory mol-ecule expression, can interfere with the function of DCs andtheir ability to prime naïve T cells. The effect of our antigen onDC function was further confirmed by the demonstration of
reduced phagocytic ability by Teg-primed DCs which wouldalso impact on the development of the adaptive immune re-sponse. We hypothesize, therefore, that Teg maintains the DCsin an immature state, impairing their function and ultimatelymodulating the development of adaptive T-cell responses. Ourdata support previous findings which demonstrate that F. he-patica infection and antigens suppress Th1 immune responsesin vivo (7, 19). Furthermore, the suppression of Th1 responsesis likely to be a survival mechanism for the worm, since vaccinetrials in domestic livestock have shown the importance of in-ducing strong Th1 immune responses in the host to protectagainst challenge infection (38).
While the exact mechanism of F. hepatica Teg-mediatedsuppression remains to be elucidated, the timing of exposure ofDCs to our antigen did not affect the suppressive effects, sug-gesting that it does not compete with LPS for binding and thatit is not exerting its effects through TLR4 itself. This wasconfirmed by the fact that Teg is still effective in DCs fromTLR4 mutant and knockout mice. Furthermore, Teg was ableto suppress the effects of all TLR ligands tested (which in-cluded both MyD88-dependent and -independent pathways),in addition to non-TLR ligands. While we would not directlyrule out the involvement of other TLRs, it seems likely thatTeg utilizes a pathway common to these receptors, particularlysince it did not activate HEK-296 cells functionally expressinga range of TLRs.
The MAPK pathway is a highly conserved pathway involvedin the initiation of DC maturation through all known TLRligands (13). Activation of the three main mammalian groups,JNK, ERK, and p38, culminates in the release of cytokinesfrom DCs following the downstream activation of a signalingcascade involving adaptor proteins, such as MyD88 and Mal(41). Helminth-mediated suppression of cytokine productionby antigen-presenting cells has been shown to be modulated byMAPK pathways. For example, ES-62-mediated suppressionof LPS-induced IL-12p40 by macrophages can be reversed withthe addition of the ERK inhibitor (22), and S. mansoni eggantigen dramatically reduces LPS-stimulated phosphorylationof p38 (29). However, in the present study, we have shown thatF. hepatica Teg does not target these pathways, since MAPKpathways are involved in the suppressive effect of only a smallnumber of select cytokines.
Instead, we found that F. hepatica Teg targeted the tran-scription factor NF-�Bp65, one of the active subunits of theNF-�B complex involved in NF-�B binding to DNA. Thiseffect on downstream signaling events could explain the ob-served decrease in proinflammatory cytokines demonstrated inthis study. Activation of NF-�B is also inhibited by other par-asites, such as Brugia malayi (48) and Toxoplasma gondii (8),where NF-�B translocation is blocked from entering the nu-cleus. We have yet to determine whether Teg can suppressother members of the NF-�B family (such as the p50 subunit)or other transcription factors, such as interferon regulatoryfactor 3, which are also activated following LPS stimulation.
In summary, this is the first study to report that F. hepaticategumental antigen modulates DC maturation and function,which may go some way to explaining its reported immuno-modulatory properties in vivo. F. hepatica tegumental antigenis a heterogeneous group of molecules, and preliminary pro-teomic studies have already pointed to a number of potential
2496 HAMILTON ET AL. INFECT. IMMUN.
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

molecules that may be involved in its anti-inflammatory capac-ity (R. M. Morphew and P. M. Brophy, unpublished data).These proteomic studies also show that the profile observed forthe tegument differs from that of its F. hepatica excretory-secretory products, and preliminary studies using excretory-secretory products demonstrate that it has a different modula-tory effect on DC maturation (C. M. Hamilton, D. J. Dowling,and S. M. O’Neill, unpublished data). Although the preciseimmunological scenario is not completely understood, it isclear that Teg maintains the DCs in an immature state, im-pairing their function and the subsequent development ofadaptive immunity. Given the powerful modulatory effect thatTeg exhibits, understanding its exact mechanisms may lead tothe development of novel immune therapeutics for the treat-ment of Th1-mediated inflammatory diseases (15, 49).
ACKNOWLEDGMENTS
This work was supported by the Dublin City University Faculty ofScience and Health Targeted Research Development Fund, EuropeanUnion (DELIVER) (grant FOOD-CT-2005-023025), and BBSRC(grant BB/C503638/2).
We thank Carolyn Wilson (Dublin City University [DCU]) for tech-nical support. We also thank Anthony Ryan and Eve Draper (DCU)and Bernie Mahon and Ciaran Skerry (National University of Ireland,Maynooth) for assistance with experiments and Cariosa Noone (DCU)for critical reading of the manuscript.
REFERENCES
1. Abreu, M. T., and M. Arditi. 2004. Innate immunity and Toll-like receptors:clinical implications of basic science research. J. Pediatr. 144:421–429.
2. Aitken, M. M., P. W. Jones, G. A. Hall, D. L. Hughes, and G. T. Brown. 1981.Responses of fluke-infected and fluke-free cattle to experimental reinfectionwith Salmonella dublin. Res. Vet. Sci. 31:120–126.
3. Balic, A., Y. Harcus, M. J. Holland, and R. M. Maizels. 2004. Selectivematuration of dendritic cells by Nippostrongylus brasiliensis-secreted proteinsdrives Th2 immune responses. Eur. J. Immunol. 34:3047–3059.
4. Banchereau, J., and R. M. Steinman. 1998. Dendritic cells and the control ofimmunity. Nature 392:245–252.
5. Beutler, B. 2004. Inferences, questions and possibilities in Toll-like receptorsignalling. Nature 430:257–263.
6. Borkow, G., and Z. Bentwich. 2006. HIV and helminth co-infection: is de-worming necessary? Parasite Immunol. 28:605–612.
7. Brady, M. T., S. M. O’Neill, J. P. Dalton, and K. H. G. Mills. 1999. Fasciolahepatica suppresses a protective Th1 response against Bordetella pertussis.Infect. Immun. 67:5372–5378.
8. Butcher, B. A., L. Kim, P. E. Johnson, and E. Y. Denkers. 2001. Toxoplasmagondii tachyzoites inhibit proinflammatory cytokine induction in infectedmacrophages by preventing nuclear translocation of the transcription factorNF-kappa B. J. Immunol. 167:2193–2201.
9. Cervi, L., A. S. MacDonald, C. Kane, F. Dzierszinski, and E. J. Pearce. 2004.Dendritic cells copulsed with microbial and helminth antigens undergo mod-ified maturation, segregate the antigens to distinct intracellular compart-ments, and concurrently induce microbe-specific Th1 and helminth-specificTh2 responses. J. Immunol. 172:2016–2020.
10. Clery, D., P. Torgerson, and G. Mulcahy. 1996. Immune responses of chron-ically infected adult cattle to Fasciola hepatica. Vet. Parasitol. 62:71–82.
11. Daubeuf, B., J. Mathison, S. Spiller, S. Hugues, F. Herren, S. Ferlin, and M.Kosco-Vilbois. 2007. TLR4/MD-2 monoclonal antibody therapy affords pro-tection in experimental models of septic shock. J. Immunol. 179:6107–6114.
12. Donnelly, S., S. M. O’Neill, M. Sekiya, G. Mulcahy, and J. P. Dalton. 2005.Thioredoxin peroxidase secreted by Fasciola hepatica induces the alternativeactivation of macrophages. Infect. Immun. 73:166–173.
13. Dowling, D. J., C. M. Hamilton, and S. M. O’Neill. 2008. A comparativeanalysis of cytokine responses, cell surface marker expression and MAPKs inDCs matured with LPS compared with a panel of TLR ligands. Cytokine41:254–262.
14. Elias, D., S. Britton, A. Kassu, and H. Akuffo. 2007. Chronic helminthinfections may negatively influence immunity against tuberculosis and otherdiseases of public health importance. Expert Rev. Anti. Infect. Ther. 5:475–484.
15. Elliott, D. E., J. Li, A. Blum, A. Metwali, K. Qadir, J. F. Urban, Jr., and J. V.Weinstock. 2003. Exposure to schistosome eggs protects mice from TNBS-induced colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 284:G385–G391.
16. Elliott, D. E., T. Setiawan, A. Metwali, A. Blum, J. F. Urban, Jr., and J. V.
Weinstock. 2004. Heligmosomoides polygyrus inhibits established colitis inIL-10-deficient mice. Eur. J. Immunol. 34:2690–2698.
17. Fallon, P. G., E. J. Richardson, P. Smith, and D. W. Dunne. 2000. Elevatedtype 1, diminished type 2 cytokines and impaired antibody response areassociated with hepatotoxicity and mortalities during Schistosoma mansoniinfection of CD4-depleted mice. Eur. J. Immunol. 30:470–480.
18. Flynn, R. J., and G. Mulcahy. 2008. The roles of IL-10 and TGF- incontrolling IL-4 and IFN-� production during experimental Fasciola hepaticainfection. Int. J. Parasitol. 38:1673–1680.
19. Flynn, R. J., C. Mannion, O. Golden, O. Hacariz, and G. Mulcahy. 2007.Experimental Fasciola hepatica infection alters responses to tests used fordiagnosis of bovine tuberculosis. Infect. Immun. 75:1373–1381.
20. Goodridge, H. S., E. H. Wilson, W. Harnett, C. C. Campbell, M. M. Harnett,and F. Y. Liew. 2001. Modulation of macrophage cytokine production byES-62, a secreted product of the filarial nematode Acanthocheilonema viteae.J. Immunol. 167:940–945.
21. Goodridge, H. S., F. A. Marshall, K. J. Else, K. M. Houston, C. Egan, L.Al-Riyami, and F. Y. Liew. 2005. Immunomodulation via novel use of TLR4by the filarial nematode phosphorylcholine-containing secreted product, ES-62. J. Immunol. 174:284–293.
22. Goodridge, H. S., W. Harnett, F. Y. Liew, and M. M. Harnett. 2003. Differ-ential regulation of interleukin-12 p40 and p35 induction via Erk mitogen-activated protein kinase-dependent and -independent mechanisms and theimplications for bioactive IL-12 and IL-23 responses. Immunology 109:415–425.
23. Green, J. M. 2000. The B7/CD28/CTLA-4 T cell activation pathway: impli-cations for inflammatory lung disease. Am. J. Respir. Cell Mol. Biol. 22:261–264.
24. Halton, D. 2004. Microscopy and the helminth parasite. Micron 35:361–390.25. Hayden, M. S., A. P. West, and S. Ghosh. 2006. NF-�B and the immune
response. Oncogene 25:6758–6780.26. Hillyer, G. V. 1980. Isolation of Fasciola hepatica tegument antigens. J. Clin.
Microbiol. 12:695–699.27. Janeway, C. A., and R. Medzhitov. 2002. Innate immune recognition. Annu.
Rev. Immunol. 20:197–216.28. Jenkins, S. J., and A. P. Mountford. 2005. Dendritic cells activated with
products released by schistosome larvae drive Th2-type immune responses,which can be inhibited by manipulation of CD40 costimulation. Infect. Im-mun. 73:395–402.
29. Kane, C. M., L. Cervi, J. Sun, A. S. McKee, K. S. Masek, S. Shapira, C. A.Hunter, and E. J. Pearce. 2004. Helminth antigens modulate TLR-initiateddendritic cell activation. J. Immunol. 173:7454–7461.
30. Kapsenberg, M. 2003. Dendritic-cell control of pathogen-driven T-cell po-larization. Nat. Rev. Immunol. 3:984–993.
31. Kobayashi, M., M. N. Kweon, H. Kuwata, R. D. Schreiber, H. Kiyono, K.Takeda, and S. Akira. 2003. Toll-like receptor-dependent production ofIL-12p40 causes chronic enterocolitis in myeloid cell-specific Stat3-deficientmice. J. Clin. Investig. 111:1297–1308.
32. La Flamme, A. C., K. Ruddenklau, and B. T. Backstrom. 2003. Schistoso-miasis decreases central nervous system inflammation and alters the pro-gression of experimental autoimmune encephalomyelitis. Infect. Immun.71:4996–5004.
33. MacDonald, A. S., and R. M. Maizels. 2008. Alarming dendritic cells for Th2induction. J. Exp. Med. 205:13–17.
34. Maizels, R. M., A. Balic, N. Gomez-Escobar, M. Nair, M. D. Taylor, and J. D.Allen. 2004. Helminth parasites—masters of immune regulation. Immunol.Rev. 201:89–116.
35. Maizels, R. M., and M. Yazdanbakhsh. 2003. Immune regulation by hel-minth parasites: cellular and molecular mechanisms. Nat. Rev. Immunol.3:733–744.
36. Mas-Coma, S., M. D. Barques, and J. G. Esteban. 1999. Human fasciolosis,p. 411–447. In J. P. Dalton (ed.), Fasciolosis. CAB International, Oxon,United Kingdom.
37. Moore, K. W., R. L. de Waal Malefyt, R. Coffman, and A. O’Garra. 2001.Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 19:683–765.
38. Mulcahy, G., F. O’Connor, S. McGonigle, A. Dowd, D. G. Clery, S. J.Andrews, and J. P. Dalton. 1998. Correlation of specific antibody titre andavidity with protection in cattle immunized against Fasciola hepatica. Vac-cine 16:932–939.
39. Mulcahy, G., P. Joyce, and J. P. Dalton. 1999. Immunology of Fasciolahepatica infection, p. 341–375. In J. P. Dalton (ed.), Fasciolosis. CAB Inter-national, Oxon, United Kingdom.
40. Mwangi, T. W., J. M. Bethony, and S. Brooker. 2006. Malaria and helminthinteractions in humans: an epidemiological viewpoint. Ann. Trop. Med.Parasitol. 100:551–570.
41. Nakahara, T., Y. Moroi, H. Uchi, and M. Furue. 2006. Differential role ofMAPK signaling in human dendritic cell maturation and Th1/Th2 engage-ment. J. Dermatol. Sci. 42:1–11.
42. Noone, C. M., E. A. Lewis, A. B. Frawely, R. W. Newman, B. P. Mahon, K. H.Mills, and P. A. Johnson. 2005. Novel mechanism of immunosuppression byinfluenza virus haemagglutinin: selective suppression of interleukin 12 p35
VOL. 77, 2009 F. HEPATICA Teg SUPPRESSES DC MATURATION AND FUNCTION 2497
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

transcription in murine bone marrow-derived dendritic cells. J. Gen. Virol.86:1885–1890.
43. O’Garra, A., and P. Vieira. 2007. TH1 cells control themselves by producinginterleukin-10. Nat. Rev. Immunol. 7:425–428.
44. O’Neill, S. M., M. T. Brady, J. J. Callanan, G. Mulcahy, P. Joyce, K. H. G.Mills, and J. P. Dalton. 2000. Fasciola hepatica infection downregulates Th1responses in mice. Parasite Immunol. 22:147–155.
45. O’Neill, S. M., K. H. Mills, and J. P. Dalton. 2001. Fasciola hepatica cathep-sin L cysteine proteinase suppresses Bordetella pertussis-specific interferon-gamma production in vivo. Parasite Immunol. 23:541–547.
46. Perona-Wright, G., S. J. Jenkins, and A. S. MacDonald. 2006. Dendritic cellactivation and function in response to Schistosoma mansoni. Int. J. Parasitol.36:711–721.
47. Poltorak, A., X. He, I. Smirnova, M. Y. Liu, C. Van Huffel, X. Du, and D.Birdwell. 1998. Defective LPS signaling in C3H/HeJ and C57BL/10ScCrmice: mutations in Tlr4 gene. Science 282:2085–2088.
48. Semnani, R. T., P. G. Venugopal, C. A. Leifer, S. Mostbock, H. Sabzevari,and T. B. Nutman. 2008. Inhibition of TLR3 and TLR4 function and ex-pression in human dendritic cells by helminth parasites. Blood 112:1290–1298.
49. Summers, R. W., D. E. Elliot, J. F. Urban, R. Thompson, and J. V. Wein-stock. 2005. Trichuris suis therapy in Crohn’s disease. Gut 54:87–90.
50. Trinchieri, G. 2003. Interleukin-12 and the regulation of innate resistanceand adaptive immunity. Nat. Rev. Immunol. 3:133–146.
51. Waibler, Z., U. Kalinke, J. Will, M. H. Juan, J. M. Pfeilschifter, and H. H.Radeke. 2007. TLR-ligand stimulated interleukin-23 subunit expression andassembly is regulated differentially in murine plasmacytoid and myeloiddendritic cells. Mol. Immunol. 44:1483–1489.
52. Whelan, M., M. M. Harnett, K. M. Houston, V. Patel, W. Harnett, and K. P.Rigley. 2000. A filarial nematode-secreted product signals dendritic cells toacquire a phenotype that drives development of Th2 cells. J. Immunol.164:6453–6460.
Editor: J. F. Urban, Jr.
2498 HAMILTON ET AL. INFECT. IMMUN.
on Novem
ber 11, 2018 by guesthttp://iai.asm
.org/D
ownloaded from





























