Embed Size (px)
Citation preview

This article was downloaded by: [University of Guelph]On: 07 September 2012, At: 18:26Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Studies on Neotropical Fauna and EnvironmentPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/nnfe20
Feeding habits of the maned wolf, Chrysocyonbrachyurus (Carnivora: Canidae), in southeast BrazilAdriana de Arruda Bueno a & José Carlos Motta-Junior aa Departamento de Ecologia, Instituto de Biociências, Universidade de São Paulo, São Paulo,Brazil
Version of record first published: 23 Jul 2009
To cite this article: Adriana de Arruda Bueno & José Carlos Motta-Junior (2009): Feeding habits of the maned wolf,Chrysocyon brachyurus (Carnivora: Canidae), in southeast Brazil, Studies on Neotropical Fauna and Environment, 44:2, 67-75
To link to this article: http://dx.doi.org/10.1080/01650520902891413
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

Studies on Neotropical Fauna and EnvironmentVol. 44, No. 2, August 2009, 67–75
ISSN 0165-0521 print/ISSN 1744-5140 online© 2009 Taylor & FrancisDOI: 10.1080/01650520902891413http://www.informaworld.com
NNFEORIGINAL ARTICLE
Feeding habits of the maned wolf, Chrysocyon brachyurus (Carnivora: Canidae), in southeast BrazilStudies on Neotropical Fauna and EnvironmentAdriana de Arruda Bueno* & José Carlos Motta-Junior
Departamento de Ecologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, Brazil
(Received 23 May 2005; accepted 13 March 2009)
The feeding habits of the maned wolf were studied in southeast Brazil to understand its response towardschanges in the environment and in relation to its prey. By occurrence, miscellaneous fruits, small mammalsand wolf’s fruit were the most consumed items. Armadillos, small mammals and wolf’s fruit (Solanum lycocar-pum) provided most of the ingested biomass. While wolf’s fruit and small mammals were mainly consumed inthe dry season, other miscellaneous fruits were taken mostly in the wet season. There was selectivity in thepredation on some small mammal species. The maned wolf’s diet followed patterns similar to those found inmore pristine areas.
O hábito alimentar do lobo guará foi estudado no sudeste do Brasil para descrever suas respostas as alterações doambiente e em relação as suas presas. Por frequência de ocorrência, as frutas silvestres, os pequenos mamíferos ea fruta-do-lobo foram os itens mais consumidos. Tatus, pequenos mamíferos e a frutas-do-lobo (Solanumlycocarpum) constituíram os itens mais importantes em relação à biomassa ingerida. A fruta-do-lobo e os peque-nos mamíferos foram consumidos principalmente na estação seca, enquanto os demais frutos silvestres na épocadas chuvas. Houve seleção de algumas espécies de pequenos mamíferos. A dieta do lobo guará apresentoupadrões semelhantes aqueles encontrados em áreas mais preservadas.
Keywords: Brazil; Chrysocyon brachyurus; diet; prey selection; savannah; seasonality
Introduction
The maned wolf (Chrysocyon brachyurus), a large SouthAmerican grassland canid, is included in the IUCN RedList (International Union for Conservation of Natureand Natural Resources) as a “lower risk – near threat-ened” species (IUCN 2008) and in the Brazilian list ofanimals threatened with extinction as “vulnerable”(Ministério do Meio Ambiente 2008). Although its ori-ginal range is wide, including open vegetation all overParaguay, northeast Argentina, northwest Uruguay,southeast Peru, large parts of central and east Bolivia,and central and south Brazil, it was severely reducedor replaced by pasture and agriculture (Paula et al.2008). The maned wolf’s diet is well-known (Dietz1984; Motta-Junior et al. 1996; Aragona & Setz 2001;Bueno et al. 2002; Juarez & Marinho-Filho 2002;Bueno & Motta-Junior 2004; Jácomo et al. 2004;Queirolo & Motta-Junior 2007; Rodrigues et al.2007); however, studies on prey selection were seldomconducted (except for Motta-Junior 2000; Bueno &Motta-Junior 2006; Queirolo & Motta-Junior 2007;Rodrigues et al. 2007). As studies of prey selectivityby neotropical carnivores are necessary to elucidate
patterns of predation among South American grass-land carnivores (Jaksic et al. 1981; Jaksic & Simonetti1987), our main goals were to learn seasonality andselectivity of small mammals in the diet of the manedwolf to better understand its response towards changesin the environment and the relation with its animal prey.
Material and methods
Study site
The research took place in the Estação Experimentalde Itapetininga (EEXI), Itapetininga, state of SãoPaulo, Brazil (23º42′S, 47º57′W, 6706 ha). The climateis Cfa, hot with a dry winter (Köppen 1948), withmarked wet (September to March) and dry (April toOctober) seasons. The EEXI contains mostly Pinusspp. and Eucalyptus spp. plantations, as well as areaswith native vegetation, savannah and grasslands(locally named “cerrado” and “campo-cerrado”,respectively), marshes and gallery forests. The naturalareas have been invaded by exotic grass and pines andthey are disturbed by human settlement and extractionof pine resin.
*Corresponding author. Email: [email protected]
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012

68 A. de Arruda Bueno & J. C. Motta-Junior
Collection and analysis of fecal samples
From March 1998 to February 2000 we walked anddrove vehicles to search for fecal samples monthly. Inview of the fact that no visit in the study area waspossible in September 1998 and February 1999, datawere gathered for a total of 22 months. According toour estimates, by direct observation or by listening tovocalizations of different individuals at the sametime, there were three to five wolves at least partiallyactive in the area during this study period. Manedwolf’s feces were identified by shape, diameter, smell,wolf fur (allogrooming) and presence of tracks (Dietz1984; Becker & Dalponte 1991). Feces with uncertainidentification were not included in this study. Wewashed feces in a washing machine (Springer & Smith1981; Uresk & Sharps 1986) in the laboratory, andthen dried them at 50ºC for 24–48 h in an oven-drier.
We identified remains of teeth, bones, fur, bills,feathers, scales and seeds by comparing diagnosticcharacteristics (patterns of shape, size and color) with areference collection obtained simultaneously in thestudy site. In case of doubt, the remains were analyzedby specialists. This material, such as pairs of mandiblesor bills, was used to count the minimum number of ani-mal individuals consumed (Emmons 1987; Motta-Junior et al. 1996). The estimates of prey biomassingested were calculated by counting the minimumnumber of individuals in feces and then multiplying thisnumber by the mean body mass of each species at thestudy site. When direct assessment was not possible, theliterature (Eisenberg & Redford 1999) and museumspecialists were consulted to obtain other prey meanweights. Whenever prey age was identified in feces bymeans of teeth or bone conditions, the respective meanweights for each age class were applied to estimate bio-mass ingestion by the wolves. We assumed completeingestion of prey when teeth, claws and bones of allparts of the skeleton were found in the feces, includinglarger prey as armadillos (Motta-Junior et al. 1996).Following Emmons (1987), weight was corrected tothree-quarters of original mean adult weight for preyabove 5 kg. Whenever it was possible to identify the sizebut not the species, the estimated biomass consumed ofunidentified prey was based on the average body massof known species of similar size. The mean number ofseeds/fruit and mean mass of one fruit for each specieswere obtained in the field to estimate the number offruit and biomass ingested by the maned wolf (accord-ing to Castro et al. 1994; Motta-Junior et al. 1996).
Sampling of small mammals
We sampled small mammals from June 1999 toFebruary 2000, conducting mark–recapture procedures.
We established two transects, one in grassland savannahand one in savannah, with 35 Tomahawk traps(40 × 18 × 17 cm) each, set 10 m apart from eachother. The EEXI harbors 19.2% of open native vege-tation (JP Metzger, personal communication), whichwere assumed to be the most frequently visited habitatby the wolves (see Dietz 1984; Motta-Junior & Martins2002; Jácomo et al. 2004; Queirolo & Motta-Junior2007). Moreover, large portions of the study area(> 60%) were covered with exotic Pinus and Eucalyptusplantations without any understory. The traps wereopened four days per month, totalling 2520 trap-nights.We used a bait mixture of banana, sardine, peanutbutter, corn flour and dog food (Frolic®) to avoidselective trapping due to diet preferences. Each smallmammal captured was marked with perforations inthe ears for individual identification, weighed, sexedand released. We compared abundance of smallmammals in the diet and in the study area with con-tingency table chi-square (χ2) tests (Jaksic 1989; Zar1999) and Bonferroni confidence intervals accordingto Byers & Steinhorst (1984) and Plumpton & Lutz(1993). In the prey selectivity analysis, we used onlyfecal samples collected from June 1999 to February2000, since live-trap procedures were performed inthis period.
The relative importance of food items in the sam-pled feces was calculated using frequency of occur-rence (number of feces containing a certain item as afunction of the total number of occurrences of allitems, following Dietz 1984) and estimated biomassingested. To test for seasonal variation in the diet, weused the modified Morisita’s coefficient (Wolda 1981)to compare the frequency of occurrence of prey itemsamong months. Also goodness of fit chi-square testswere employed to verify whether some groups ofitems were consumed according to the season of theyear (e.g., Dietz 1984; Motta-Junior et al. 1996). Thefood-niche breadth using the standardized Levinsmeasure (Bst) was employed considering the wholeperiod of study and the two seasons separately (Krebs1999). In all tests statistical significance was P < 0.05.
Results
The study was based on the analysis of 614 feces,yielding 2070 occurrences, distributed among 70 iden-tified prey and fruit items. Eighteen were vegetal and52 were animal food items (Table 1). By occurrence,the wolves consumed both plant (57.0%) and animal(43.0%) food in similar proportions, exhibiting anomnivorous diet. The most frequently consumed itemswere miscellaneous fruits (24.2%), small mammals(21.4%) and wolf’s fruits (Solanum lycocarpum, 17.2%).By biomass, armadillos (28.2%), small mammals
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012
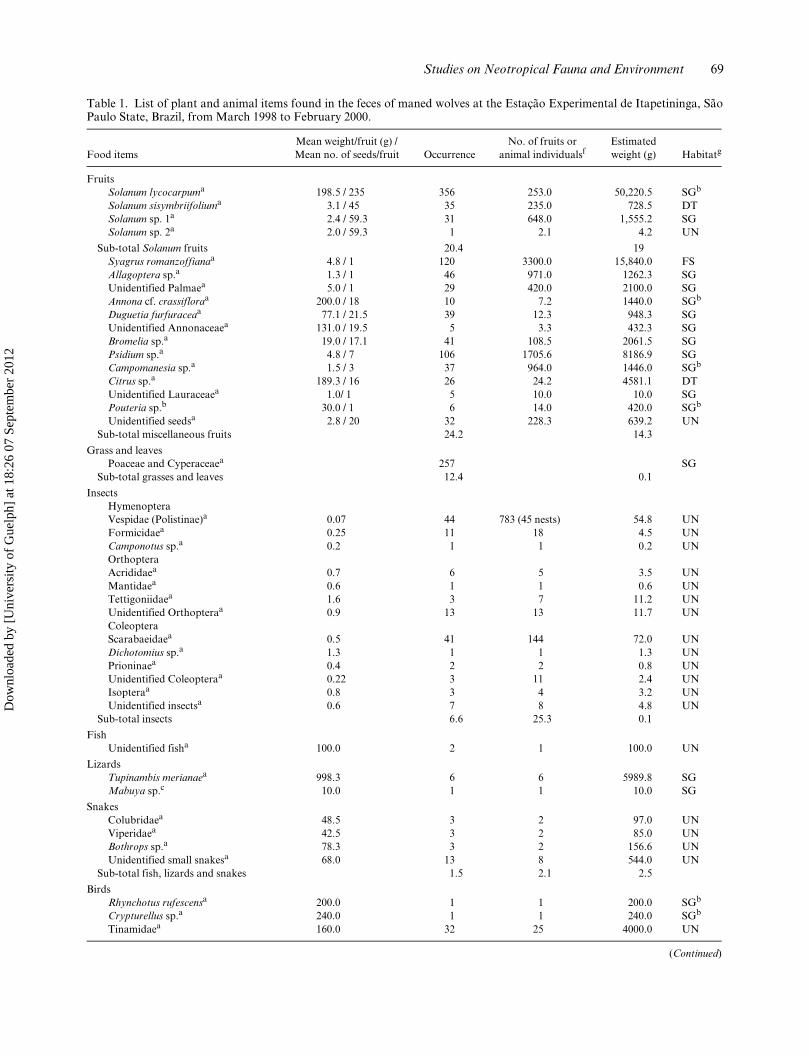
Studies on Neotropical Fauna and Environment 69
Table 1. List of plant and animal items found in the feces of maned wolves at the Estação Experimental de Itapetininga, SãoPaulo State, Brazil, from March 1998 to February 2000.
Food itemsMean weight/fruit (g) / Mean no. of seeds/fruit Occurrence
No. of fruits or animal individualsf
Estimated weight (g) Habitatg
FruitsSolanum lycocarpuma 198.5 / 235 356 253.0 50,220.5 SGb
Solanum sisymbriifoliuma 3.1 / 45 35 235.0 728.5 DTSolanum sp. 1a 2.4 / 59.3 31 648.0 1,555.2 SGSolanum sp. 2a 2.0 / 59.3 1 2.1 4.2 UN
Sub-total Solanum fruits 20.4 19Syagrus romanzoffianaa 4.8 / 1 120 3300.0 15,840.0 FSAllagoptera sp.a 1.3 / 1 46 971.0 1262.3 SGUnidentified Palmaea 5.0 / 1 29 420.0 2100.0 SGAnnona cf. crassifloraa 200.0 / 18 10 7.2 1440.0 SGb
Duguetia furfuraceaa 77.1 / 21.5 39 12.3 948.3 SGUnidentified Annonaceaea 131.0 / 19.5 5 3.3 432.3 SGBromelia sp.a 19.0 / 17.1 41 108.5 2061.5 SGPsidium sp.a 4.8 / 7 106 1705.6 8186.9 SGCampomanesia sp.a 1.5 / 3 37 964.0 1446.0 SGb
Citrus sp.a 189.3 / 16 26 24.2 4581.1 DTUnidentified Lauraceaea 1.0/ 1 5 10.0 10.0 SGPouteria sp.b 30.0 / 1 6 14.0 420.0 SGb
Unidentified seedsa 2.8 / 20 32 228.3 639.2 UNSub-total miscellaneous fruits 24.2 14.3
Grass and leavesPoaceae and Cyperaceaea 257 SG
Sub-total grasses and leaves 12.4 0.1
InsectsHymenopteraVespidae (Polistinae)a 0.07 44 783 (45 nests) 54.8 UNFormicidaea 0.25 11 18 4.5 UNCamponotus sp.a 0.2 1 1 0.2 UNOrthopteraAcrididaea 0.7 6 5 3.5 UNMantidaea 0.6 1 1 0.6 UNTettigoniidaea 1.6 3 7 11.2 UNUnidentified Orthopteraa 0.9 13 13 11.7 UNColeopteraScarabaeidaea 0.5 41 144 72.0 UNDichotomius sp.a 1.3 1 1 1.3 UNPrioninaea 0.4 2 2 0.8 UNUnidentified Coleopteraa 0.22 3 11 2.4 UNIsopteraa 0.8 3 4 3.2 UNUnidentified insectsa 0.6 7 8 4.8 UN
Sub-total insects 6.6 25.3 0.1
FishUnidentified fisha 100.0 2 1 100.0 UN
LizardsTupinambis merianaea 998.3 6 6 5989.8 SGMabuya sp.c 10.0 1 1 10.0 SG
SnakesColubridaea 48.5 3 2 97.0 UNViperidaea 42.5 3 2 85.0 UNBothrops sp.a 78.3 3 2 156.6 UNUnidentified small snakesa 68.0 13 8 544.0 UN
Sub-total fish, lizards and snakes 1.5 2.1 2.5
BirdsRhynchotus rufescensa 200.0 1 1 200.0 SGb
Crypturellus sp.a 240.0 1 1 240.0 SGb
Tinamidaea 160.0 32 25 4000.0 UN
(Continued)
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012
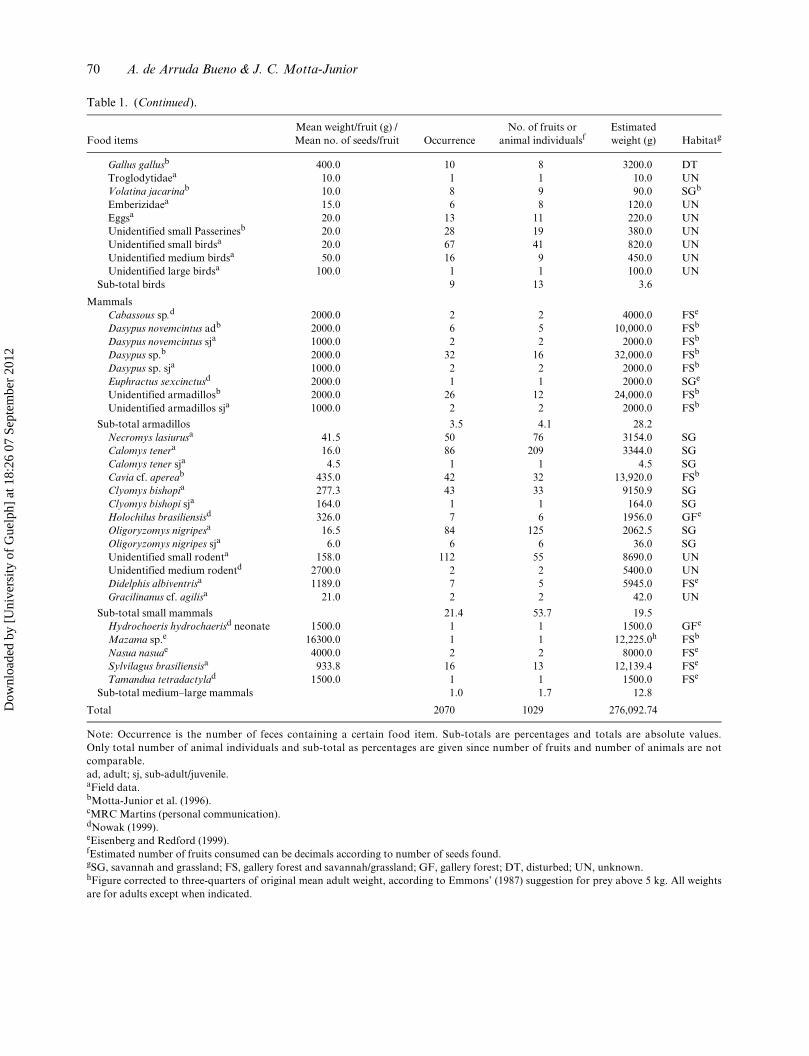
70 A. de Arruda Bueno & J. C. Motta-Junior
Table 1. (Continued).
Food itemsMean weight/fruit (g) / Mean no. of seeds/fruit Occurrence
No. of fruits or animal individualsf
Estimated weight (g) Habitatg
Gallus gallusb 400.0 10 8 3200.0 DTTroglodytidaea 10.0 1 1 10.0 UNVolatina jacarinab 10.0 8 9 90.0 SGb
Emberizidaea 15.0 6 8 120.0 UNEggsa 20.0 13 11 220.0 UNUnidentified small Passerinesb 20.0 28 19 380.0 UNUnidentified small birdsa 20.0 67 41 820.0 UNUnidentified medium birdsa 50.0 16 9 450.0 UNUnidentified large birdsa 100.0 1 1 100.0 UN
Sub-total birds 9 13 3.6
MammalsCabassous sp.d 2000.0 2 2 4000.0 FSe
Dasypus novemcintus adb 2000.0 6 5 10,000.0 FSb
Dasypus novemcintus sja 1000.0 2 2 2000.0 FSb
Dasypus sp.b 2000.0 32 16 32,000.0 FSb
Dasypus sp. sja 1000.0 2 2 2000.0 FSb
Euphractus sexcinctusd 2000.0 1 1 2000.0 SGe
Unidentified armadillosb 2000.0 26 12 24,000.0 FSb
Unidentified armadillos sja 1000.0 2 2 2000.0 FSb
Sub-total armadillos 3.5 4.1 28.2Necromys lasiurusa 41.5 50 76 3154.0 SGCalomys tenera 16.0 86 209 3344.0 SGCalomys tener sja 4.5 1 1 4.5 SGCavia cf. apereab 435.0 42 32 13,920.0 FSb
Clyomys bishopia 277.3 43 33 9150.9 SGClyomys bishopi sja 164.0 1 1 164.0 SGHolochilus brasiliensisd 326.0 7 6 1956.0 GFe
Oligoryzomys nigripesa 16.5 84 125 2062.5 SGOligoryzomys nigripes sja 6.0 6 6 36.0 SGUnidentified small rodenta 158.0 112 55 8690.0 UNUnidentified medium rodentd 2700.0 2 2 5400.0 UNDidelphis albiventrisa 1189.0 7 5 5945.0 FSe
Gracilinanus cf. agilisa 21.0 2 2 42.0 UN
Sub-total small mammals 21.4 53.7 19.5Hydrochoeris hydrochaerisd neonate 1500.0 1 1 1500.0 GFe
Mazama sp.e 16300.0 1 1 12,225.0h FSb
Nasua nasuae 4000.0 2 2 8000.0 FSe
Sylvilagus brasiliensisa 933.8 16 13 12,139.4 FSe
Tamandua tetradactylad 1500.0 1 1 1500.0 FSe
Sub-total medium–large mammals 1.0 1.7 12.8
Total 2070 1029 276,092.74
Note: Occurrence is the number of feces containing a certain food item. Sub-totals are percentages and totals are absolute values.Only total number of animal individuals and sub-total as percentages are given since number of fruits and number of animals are notcomparable.ad, adult; sj, sub-adult/juvenile.aField data.bMotta-Junior et al. (1996).cMRC Martins (personal communication).dNowak (1999).eEisenberg and Redford (1999).fEstimated number of fruits consumed can be decimals according to number of seeds found.gSG, savannah and grassland; FS, gallery forest and savannah/grassland; GF, gallery forest; DT, disturbed; UN, unknown.hFigure corrected to three-quarters of original mean adult weight, according to Emmons’ (1987) suggestion for prey above 5 kg. All weightsare for adults except when indicated.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012
Studies on Neotropical Fauna and Environment 71
(19.5%) and wolf’s fruits (18.2%) were the bulk of thediet. Most of the diet (60.7% of occurrences) wasbased on items found in physiognomies of nativesavannah (Table 1).
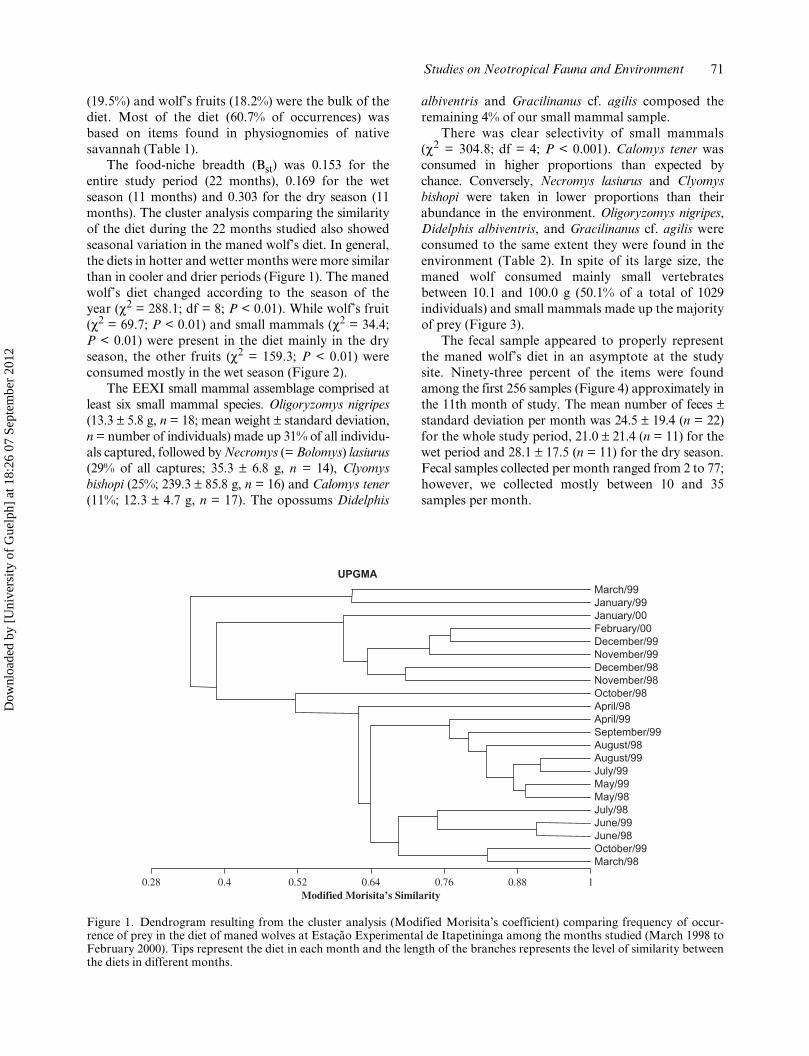
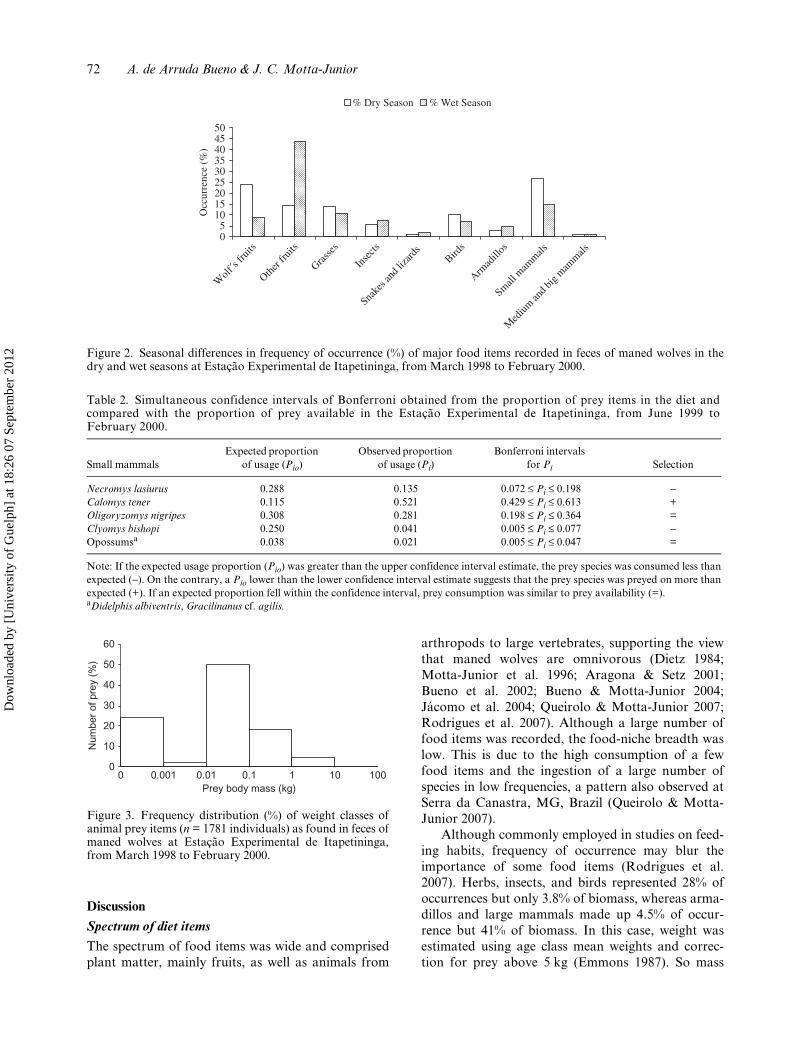
The food-niche breadth (Bst) was 0.153 for theentire study period (22 months), 0.169 for the wetseason (11 months) and 0.303 for the dry season (11months). The cluster analysis comparing the similarityof the diet during the 22 months studied also showedseasonal variation in the maned wolf’s diet. In general,the diets in hotter and wetter months were more similarthan in cooler and drier periods (Figure 1). The manedwolf’s diet changed according to the season of theyear (χ2 = 288.1; df = 8; P < 0.01). While wolf’s fruit(χ2 = 69.7; P < 0.01) and small mammals (χ2 = 34.4;P < 0.01) were present in the diet mainly in the dryseason, the other fruits (χ2 = 159.3; P < 0.01) wereconsumed mostly in the wet season (Figure 2).
The EEXI small mammal assemblage comprised atleast six small mammal species. Oligoryzomys nigripes(13.3 ± 5.8 g, n = 18; mean weight ± standard deviation,n = number of individuals) made up 31% of all individu-als captured, followed by Necromys (= Bolomys) lasiurus(29% of all captures; 35.3 ± 6.8 g, n = 14), Clyomysbishopi (25%; 239.3 ± 85.8 g, n = 16) and Calomys tener(11%; 12.3 ± 4.7 g, n = 17). The opossums Didelphis
albiventris and Gracilinanus cf. agilis composed theremaining 4% of our small mammal sample.
There was clear selectivity of small mammals(χ2 = 304.8; df = 4; P < 0.001). Calomys tener wasconsumed in higher proportions than expected bychance. Conversely, Necromys lasiurus and Clyomysbishopi were taken in lower proportions than theirabundance in the environment. Oligoryzomys nigripes,Didelphis albiventris, and Gracilinanus cf. agilis wereconsumed to the same extent they were found in theenvironment (Table 2). In spite of its large size, themaned wolf consumed mainly small vertebratesbetween 10.1 and 100.0 g (50.1% of a total of 1029individuals) and small mammals made up the majorityof prey (Figure 3).
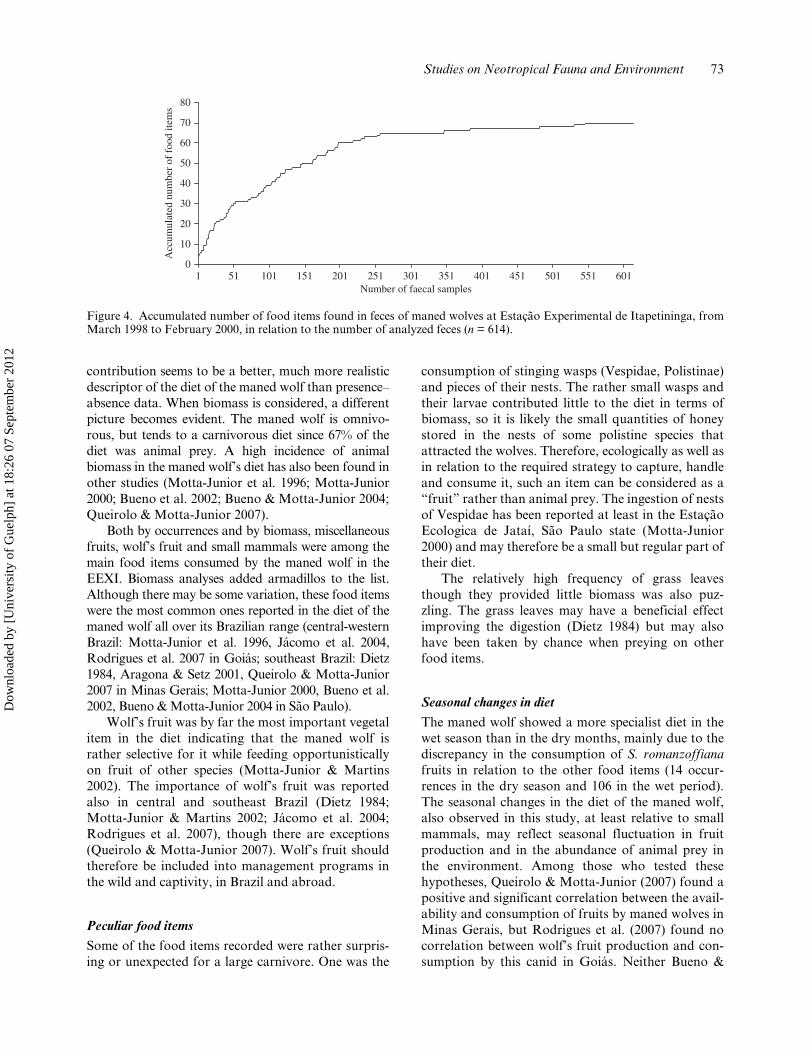
The fecal sample appeared to properly representthe maned wolf’s diet in an asymptote at the studysite. Ninety-three percent of the items were foundamong the first 256 samples (Figure 4) approximately inthe 11th month of study. The mean number of feces ±standard deviation per month was 24.5 ± 19.4 (n = 22)for the whole study period, 21.0 ± 21.4 (n = 11) for thewet period and 28.1 ± 17.5 (n = 11) for the dry season.Fecal samples collected per month ranged from 2 to 77;however, we collected mostly between 10 and 35samples per month.
Figure 1. Dendrogram resulting from the cluster analysis (Modified Morisita’s coefficient) comparing frequency of occur-rence of prey in the diet of maned wolves at Estação Experimental de Itapetininga among the months studied (March 1998 toFebruary 2000). Tips represent the diet in each month and the length of the branches represents the level of similarity betweenthe diets in different months.
UPGMA
Modified Morisita’s Similarity
March/98October/99June/98June/99July/98May/98May/99July/99August/99August/98September/99April/99April/98October/98November/98December/98November/99December/99February/00January/00January/99March/99
0.28 0.4 0.52 0.64 0.76 0.88 1
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012
72 A. de Arruda Bueno & J. C. Motta-Junior
Discussion
Spectrum of diet items
The spectrum of food items was wide and comprisedplant matter, mainly fruits, as well as animals from
arthropods to large vertebrates, supporting the viewthat maned wolves are omnivorous (Dietz 1984;Motta-Junior et al. 1996; Aragona & Setz 2001;Bueno et al. 2002; Bueno & Motta-Junior 2004;Jácomo et al. 2004; Queirolo & Motta-Junior 2007;Rodrigues et al. 2007). Although a large number offood items was recorded, the food-niche breadth waslow. This is due to the high consumption of a fewfood items and the ingestion of a large number ofspecies in low frequencies, a pattern also observed atSerra da Canastra, MG, Brazil (Queirolo & Motta-Junior 2007).
Although commonly employed in studies on feed-ing habits, frequency of occurrence may blur theimportance of some food items (Rodrigues et al.2007). Herbs, insects, and birds represented 28% ofoccurrences but only 3.8% of biomass, whereas arma-dillos and large mammals made up 4.5% of occur-rence but 41% of biomass. In this case, weight wasestimated using age class mean weights and correc-tion for prey above 5 kg (Emmons 1987). So mass
Figure 2. Seasonal differences in frequency of occurrence (%) of major food items recorded in feces of maned wolves in thedry and wet seasons at Estação Experimental de Itapetininga, from March 1998 to February 2000.
05
101520253035404550
Wolf
´s fru
its
Other f
ruits
Grasse
s
Inse
cts
Snake
s and
liza
rds
Birds
Armad
illos
Small m
ammals
Med
ium an
d big
mammals
Occ
urre
nce
(%)
% Dry Season % Wet Season
Table 2. Simultaneous confidence intervals of Bonferroni obtained from the proportion of prey items in the diet andcompared with the proportion of prey available in the Estação Experimental de Itapetininga, from June 1999 toFebruary 2000.
Small mammalsExpected proportion
of usage (Pio)Observed proportion
of usage (Pi)Bonferroni intervals
for Pi Selection
Necromys lasiurus 0.288 0.135 0.072 ≤ Pi ≤ 0.198 −Calomys tener 0.115 0.521 0.429 ≤ Pi ≤ 0.613 +Oligoryzomys nigripes 0.308 0.281 0.198 ≤ Pi ≤ 0.364 =Clyomys bishopi 0.250 0.041 0.005 ≤ Pi ≤ 0.077 −Opossumsa 0.038 0.021 0.005 ≤ Pi ≤ 0.047 =
Note: If the expected usage proportion (Pio) was greater than the upper confidence interval estimate, the prey species was consumed less thanexpected (−). On the contrary, a Pio lower than the lower confidence interval estimate suggests that the prey species was preyed on more thanexpected (+). If an expected proportion fell within the confidence interval, prey consumption was similar to prey availability (=).aDidelphis albiventris, Gracilinanus cf. agilis.
Figure 3. Frequency distribution (%) of weight classes ofanimal prey items (n = 1781 individuals) as found in feces ofmaned wolves at Estação Experimental de Itapetininga,from March 1998 to February 2000.
0
10
20
30
40
50
60
Prey body mass (kg)
Num
ber
of p
rey
(%)
0 0.001 0.01 0.1 1 10 100
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012
Studies on Neotropical Fauna and Environment 73
contribution seems to be a better, much more realisticdescriptor of the diet of the maned wolf than presence–absence data. When biomass is considered, a differentpicture becomes evident. The maned wolf is omnivo-rous, but tends to a carnivorous diet since 67% of thediet was animal prey. A high incidence of animalbiomass in the maned wolf’s diet has also been found inother studies (Motta-Junior et al. 1996; Motta-Junior2000; Bueno et al. 2002; Bueno & Motta-Junior 2004;Queirolo & Motta-Junior 2007).
Both by occurrences and by biomass, miscellaneousfruits, wolf’s fruit and small mammals were among themain food items consumed by the maned wolf in theEEXI. Biomass analyses added armadillos to the list.Although there may be some variation, these food itemswere the most common ones reported in the diet of themaned wolf all over its Brazilian range (central-westernBrazil: Motta-Junior et al. 1996, Jácomo et al. 2004,Rodrigues et al. 2007 in Goiás; southeast Brazil: Dietz1984, Aragona & Setz 2001, Queirolo & Motta-Junior2007 in Minas Gerais; Motta-Junior 2000, Bueno et al.2002, Bueno & Motta-Junior 2004 in São Paulo).
Wolf’s fruit was by far the most important vegetalitem in the diet indicating that the maned wolf israther selective for it while feeding opportunisticallyon fruit of other species (Motta-Junior & Martins2002). The importance of wolf’s fruit was reportedalso in central and southeast Brazil (Dietz 1984;Motta-Junior & Martins 2002; Jácomo et al. 2004;Rodrigues et al. 2007), though there are exceptions(Queirolo & Motta-Junior 2007). Wolf’s fruit shouldtherefore be included into management programs inthe wild and captivity, in Brazil and abroad.
Peculiar food items
Some of the food items recorded were rather surpris-ing or unexpected for a large carnivore. One was the
consumption of stinging wasps (Vespidae, Polistinae)and pieces of their nests. The rather small wasps andtheir larvae contributed little to the diet in terms ofbiomass, so it is likely the small quantities of honeystored in the nests of some polistine species thatattracted the wolves. Therefore, ecologically as well asin relation to the required strategy to capture, handleand consume it, such an item can be considered as a“fruit” rather than animal prey. The ingestion of nestsof Vespidae has been reported at least in the EstaçãoEcologica de Jataí, São Paulo state (Motta-Junior2000) and may therefore be a small but regular part oftheir diet.
The relatively high frequency of grass leavesthough they provided little biomass was also puz-zling. The grass leaves may have a beneficial effectimproving the digestion (Dietz 1984) but may alsohave been taken by chance when preying on otherfood items.
Seasonal changes in diet
The maned wolf showed a more specialist diet in thewet season than in the dry months, mainly due to thediscrepancy in the consumption of S. romanzoffianafruits in relation to the other food items (14 occur-rences in the dry season and 106 in the wet period).The seasonal changes in the diet of the maned wolf,also observed in this study, at least relative to smallmammals, may reflect seasonal fluctuation in fruitproduction and in the abundance of animal prey inthe environment. Among those who tested thesehypotheses, Queirolo & Motta-Junior (2007) found apositive and significant correlation between the avail-ability and consumption of fruits by maned wolves inMinas Gerais, but Rodrigues et al. (2007) found nocorrelation between wolf’s fruit production and con-sumption by this canid in Goiás. Neither Bueno &
Figure 4. Accumulated number of food items found in feces of maned wolves at Estação Experimental de Itapetininga, fromMarch 1998 to February 2000, in relation to the number of analyzed feces (n = 614).
0
10
20
30
40
50
60
70
80
1 51 101 151 201 251 301 351 401 451 501 551 601Number of faecal samples
Acc
umul
ated
num
ber
of f
ood
item
s
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012

74 A. de Arruda Bueno & J. C. Motta-Junior
Motta-Junior (2006) nor Queirolo & Motta-Junior(2007) detected a correlation between small mammalabundance in the environment and in the wolf’s dietin the central state of São Paulo or in Minas Gerais,respectively.
The seasonal consumption of wolf’s fruit and mis-cellaneous fruits by the maned wolf is frequentlyreported (Motta-Junior et al. 1996; Aragona & Setz2001; Bueno et al. 2002; Silva & Talamoni 2003;Santos et al. 2003; Queirolo & Motta-Junior 2007;Rodrigues et al. 2007), also suggesting an innate char-acteristic of the species. Particularly Motta-Junior(2000), Motta-Junior & Martins (2002), and Queirolo& Motta-Junior (2007) found a significantly negativeassociation between the frequency in the diet of wolf’sfruits and other fruits. When wolves consumed fewwolf’s fruits they ingested more other fruits and viceversa. Likewise, consumption of wolf’s fruits was neg-atively correlated with the abundance of other fruits.These relationships suggest that wolves use wolf’sfruits to maintain an approximately constant rate offruit intake in the face of seasonal variation in fruitavailability (Motta-Junior & Martins 2002; Queirolo& Motta-Junior 2007).
Prey selectivity
Ecological characteristics of the maned wolf mayexplain the consumption of relatively small prey since itis a solitary canid and preys on sparsely distributed fooditems, when compared to similar-sized canids, whichprey cooperatively on larger prey herds (Moehlman1986). Though not common, active predation on largeprey by the maned wolf was already observed elsewhere(e.g., Bestelmeyer & Westbrook 1998), but it is not clearif the capybara and deer here reported were activelypreyed on or scavenged. However, no traces of necro-phagous arthropods were found, suggesting predation.
The frequently consumed rodent C. tener is mostlyterrestrial (Nowak 1999). Much to the contrary, thelow predation on C. bishopi could be understood by itssemi-fossorial habits (Avila-Pires & Wutke 1981;Vieira 1997), being possibly less vulnerable to theattack of the maned wolf. Although N. lasiurus ismainly terrestrial, it can build burrow systems (Nowak1999), which may offer some protection against thewolves. On the other hand, O. nigripes and opossums(Gracilinanus cf. agilis), all of them with some scanso-rial ability (Nowak 1999), were consumed in the sameproportions they were found in the field.
However, Bueno & Motta-Junior (2006) and Que-irolo & Motta-Junior (2007) found opposite resultsrelative to prey selection by the maned wolf inEstação Ecologica de Itirapina (EEI), state of SãoPaulo, and in the Parque Nacional da Serra da
Canastra (PNSC), state of Minas Gerais, respectively.The former revealed lower consumption of C. tenerand O. nigripes and higher predation on C. bishopithan expected by chance, sampling small mammalswith large pitfall traps (bucket of 100 liters) in the EEI,whereas the latter reported selection of N. lasiurus inSCNP, sampling with traditional wire-cage live-traps(45 × 20 × 20 cm, Tomahawk). Comparisons amongstudies with different sampling methods, study areaand small mammal assemblage prevent capturing ageneral pattern relative to prey selection by themaned wolf. Additional concern should be taken inrelation to the prey selection studies. We assume preyabundance in the environment assessed with traps is asubstitute for prey availability for the predator. Inthis case, traps should be as efficient as the predatorsin capturing different prey species and the methodol-ogy should reveal the real prey composition and abun-dance in the environment. However, to date, there isno other method to assess availability (Jaksic 1989), soprey abundance in patches where and when predatorshunt is the only practical substitute for prey assess-ment (Jaksic 1989). Future studies to evaluate thevulnerability of these species should also be developedto clarify the reasons for selectivity by this predator,both by in-depth studies on ecomorphological traits(cf. Corley et al. 1995) and by taking into account thepeculiarities of the study sites.
Diet pattern
Although the study area was extremely disturbed,with exotic vegetation and presence of human activi-ties, the maned wolf’s diet followed similar patternsfound by other studies in more pristine reserves(Dietz 1984; Motta-Junior et al. 1996; Aragona &Setz 2001; Jácomo et al. 2004; Queirolo & Motta-Jun-ior 2007). Except for Citrus fruits and a few poultry(1.2 and 0.5%, respectively), no other exotic item wasfound in the diet at the study site. Main food itemswere those found in physiognomies of savannah, show-ing the importance of this environment for this canid, asreported by Motta-Junior et al. (1996), Motta-Junior &Martins (2002), and Queirolo & Motta-Junior (2007).Although maned wolves, by the ability to include newexotic items in their diet, show some plasticitytowards environmental changes, this should be seenwith caution. Consumption of items such as poultryor crops may increase conflicts with human interests,leading to decrease of wolf populations.
Acknowledgements
We thank Javier A. Simonetti for reviewing an earlierversion of the manuscript. Two anonymous referees
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012

Studies on Neotropical Fauna and Environment 75
and Anne Zillikens gave valuable suggestions to thefinal version. Maria Helena P. Machado helped withthe English text. Antônio C. Dias and Luís C. Sousaprovided logistical support and assistance to collectionsin the field. Alexandre Percequillo, Luis F. Silveira andMárcio R. C. Martins helped, respectively, with mam-mal, bird and reptile identifications. Financial supportwas provided by a grant from FAPESP – Fundação deAmparo à Pesquisa do Estado de São Paulo.
References
Aragona M, Setz EZF. 2001. Diet of the maned wolf, Chrysocyonbrachyurus (Mammalia: Canidae), during wet and dry seasons atIbitipoca State Park, Brazil. J Zool. 254:131–136.
Avila-Pires FD, Wutke MRC. 1981. Taxonomia e evolução deClyomys Thomas 1916 (Rodentia, Echimyidae). Rev Bras Biol.41:529–534.
Becker M, Dalponte JC. 1991. Rastros de mamíferos silvestresbrasileiros: um guia de campo. 2nd ed. Brasília: Editora daUniversidade de Brasília. 180 p.
Bestelmeyer SV, Westbrook C. 1998. Maned wolf (Chrysocyonbrachyurus) predation on Pampas deer (Ozotoceros bezoarticus)in central Brazil. Mammalia. 62:591–595.
Bueno AA, Belentani SCS, Motta-Junior JC. 2002. Feeding ecol-ogy of the maned wolf, Chrysocyon brachyurus (Illiger, 1815)(Mammalia: Canidae), in the Ecological Station of Itirapina, SãoPaulo State, Brazil. Biota Neotrop. [Internet] 2(2):1–9. Availablefrom: http://www.biotaneotropica.org.br/v2n2/en/fullpaper?bn01802022002+en
Bueno AA, Motta-Junior JC. 2004. Feeding ecology of two syn-topic canids, the maned wolf (Chrysocyon brachyurus) and thecrab-eating fox (Cerdocyon thous), in Southeastern Brazil. RevChil Hist Nat. 77:5–14.
Bueno AA, Motta-Junior JC. 2006. Small mammal selection andfunctional response in the diet of the maned wolf, Chrysocyonbrachyurus (Mammalia: Canidae), in southeast Brazil. MastozoolNeotrop. 13(1):11–19.
Byers CR, Steinhorst RK. 1984. Clarification of a technique foranalysis of utilization-availability data. J Wildl Manage. 48(3):1050–1053.
Castro SA, Silva SI, Meserve PL, Gutierrez JR, Contreras LC,Jaksic FM. 1994. Frugivoría y dispersión de semillas de pimiento(Schinus molle) por el zorro culpeo (Pseudalopex culpaeus) en elParque Nacional Fray Jorge (IV Región, Chile). Rev Chil HistNat. 67:169–176.
Corley JC, Fernadez GJ, Capurro AF, Novaro AJ, Funes MC,Travaini A. 1995. Selection of cricetine prey by the culpeo fox inPatagonia: a differential prey vulnerability hypothesis. Mammalia.59(3):315–325.
Dietz JM. 1984. Ecology and social organization of the maned wolf(Chrysocyon brachyurus). Smithson Contrib Zool. 392:1–51.
Eisenberg JF, Redford KH. 1999. Mammals of the Neotropics: thecentral Neotropics – Ecuador, Peru, Bolivia, Brazil. Vol. III.Chicago: University of Chigaco Press. 609 p.
Emmons LH. 1987. Comparative feeding ecology of felids in aneotropical rainforest. Behav Ecol Sociobiol. 20:271–283.
IUCN. 2008. IUCN red list of threatened species. Available from:www.iucnredlist.org
Jácomo ATA, Silveira L, Diniz-Filho JAF. 2004. Niche separationbetween the maned wolf (Chrysocyon brachyurus), the crab-eatingfox (Dusicyon thous) and the hoary fox (Dusicyon vetulus) incentral Brazil. J Zool. 262:99–106.
Jaksic FM. 1989. Opportunism vs. selectivity among carnivorouspredators that eat mammalian prey: a statistical test of hypotheses.Oikos. 56:427–430.
Jaksic FM, Greene HW, Yanez JL. 1981. The guild structure of acommunity of predatory vertebrates in central Chile. Oecologia.49:21–28.
Jaksic FM, Simonetti JA. 1987. Predator/prey relationships amongterrestrial vertebrates: an exhaustive review of studies conductedin Southern South America. Rev Chil Hist Nat. 60:221–244.
Juarez KM, Marinho-Filho J. 2002. Diet, habitat use, and homeranges of sympatric canids in central. Brazil J Mammal. 83:925–933.
Köppen W. 1948. Climatologia: con un estudio de los climas de latierra. Mexico: Fondo de Cultura Economica. 479 p.
Krebs CJ. 1999. Ecological methodology. 2nd ed. Menlo Park(CA): Addison Wesley Longman. 620 p.
Ministério do Meio Ambiente. 2008. Lista nacional das espécies dafauna brasileira ameaçadas de extinção. Available from: http://www.mma.gov.br/port/sbf/fauna
Moehlman P. 1986. Ecology of cooperation in canids. In: RubensteinDI, Wrangham RW, editors. Ecological aspects of social evolu-tion: birds and mammals. Princeton (NJ): Princeton UniversityPress. p. 64–86.
Motta-Junior JC. 2000. Variação temporal e seleção de presas na dietado lobo guará, Chrysocyon brachyurus (Mammalia: Canidae), naEstação ecológica de Jataí, Luiz Antônio, SP. In: Santos JE, RivesJSR, editors. Estudos integrados em ecossistemas. Estação Ecológicade Jataí. Vol. I. São Carlos (Brazil): Rima Editora. p. 331–346.
Motta-Junior JC, Martins K. 2002. The frugivorous diet of themaned wolf, Chrysocyon brachyurus, in Brazil: ecology andconservation. In: Levey DJ, Silva WR, Galetti M, editors. Seeddispersal and frugivory: ecology, evolution and conservation.Wallingford (UK): CABI Publishing. p. 291–303.
Motta-Junior JC, Talamoni SA, Lombardi JA, Simokomaki K.1996. Diet of the maned wolf, Chrysocyon brachyurus, in centralBrazil. J Zool. 240:277–284.
Nowak RM. 1999. Walker’s mammals of the world. Vol. II. 6th ed.Baltimore (MD): Johns Hopkins University Press. 936 p.
Paula RC, Medici P, Morato RG. 2008. Plano de ação para aConservação do Lobo-Guará. Analise de viabilidade populacionale de habitat. Brasília: IBAMA. 160 p.
Plumpton DI, Lutz RS. 1993. Prey selection and food habits ofBurrowing owls in Colorado. Great Basin Nat. 53(3):299–304.
Queirolo D, Motta-Junior JC. 2007. Prey availability and diet ofmaned wolf in Serra da Canastra National Park, southeasternBrazil. Acta Theriol. 52(4):391–402.
Rodrigues FHG, Hass A, Lacerda ACR, Grando RLSC, BagnoMA, Bezerra AMR, Silva WR. 2007. Feeding habits of themaned wolf (Chrysocyon brachyurus) in the Brazilian cerrado.Mastozool Neotrop. 14(1):37–51.
Santos E, Setz EZF, Gobbi N. 2003. Diet of the maned wolf(Chrysocyon brachyurus) and its role in seed dispersal on a cattleranch in Brazil. J Zool. 260:203–208.
Silva AJ, Talamoni AA. 2003. Diet adjustments of the maned wolf,Chrysocyon brachyurus (Illiger) (Mammalia, Canidae), subjectedto supplemental feeding in a private natural reserve, SoutheasternBrazil. Rev Bras Zool. 20(2):339–345.
Springer JT, Smith JS. 1981. Summer food habits of coyotes inCentral Wyoming. Great Basin Nat. 41(4):449–456.
Uresk D, Sharps JC. 1986. Denning habitat and diet of the swiftfox in Western South Dakota. Great Basin Nat. 46(2):249–253.
Vieira MV. 1997. Dynamics of a rodent assemblage in a cerrado ofsoutheast Brazil. Rev Bras Biol. 57(1):99–107.
Wolda H. 1981. Similarity indices, sample size and diversity.Oecologia. 50:296–302.
Zar JH. 1999. Biostatistical analysis. Upper Saddle River (NJ):Prentice Hall. 663 p.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
uelp
h] a
t 18:
26 0
7 Se
ptem
ber
2012





























