Embed Size (px)
Citation preview

B. Wiedemann und I? Heisig
Fluorierte Chinolon-Carbonsauren: ~~ ~
Wird der Fortschritt durch die Resistenz-
entwicklung aufgeho ben ?
Zusammenfassung
Mit Nalidixinsaure wurde 1962 der erste Ver- treter der 4-Chinoloncarbonsauren (+Chi- nolone) in die antibakterielle Therapie einge- fuhrt. Begrenzte pharmakokinetische Verfiig- barkeit, schmales Wirkungsspektrum und rasche Resistenzentwicklung schrankten den klinischen Einsatz schnell ein. Nach wenig er- folgreichen Variationen des Heterocyclus brachte die Einfuhrung zweier Substituenten an Pos. 6 (Fluor) und 7 (Piperazinrest) mit Substanzen wie Norfloxacin oder Ciproflo- xacin einen entscheidenden Fortschritt so- wohl fur die Pharmakokinetik als auch die antibakterielle Wirksamkeit. Als molekulare Zielstruktur fur die Wirkung der 4-Chinolo- ne wurde ein Komplex aus DNS und dem bakteriellen Enzym Gyrase identifiziert, das an der Kontrolle der raumlichen Struktur der DNS beteiligt ist. Veranderungen in einem be- stimmten Bereich des Enzyms durch Muta- tionen sind die wichtigste Ursache fur Chino- lonresistenz. Daneben finden sich Verande- rungen in der Zellwand als weitere Ursache, wodurch der Zugang der 4-Chinolone zur Gyrase innerhalb der Zellen eingeschrankt wird. Klinische Bedeutung haben derartige Mutationen wohl bei naturlicherweise weni- ger empfindlichen Bakterien wie Pseudomo- naden, Staphylokokken oder Enterokokken. Das hat in den letzten Jahren zu einer leichten Zunahme der Resistenz ( 4 0 %) bei diesen Organismen gefuhrt. Sehr empfindliche Spe- zies wie z. B. Escherichia coli zeigen dagegen nach Ergebnissen iiberregionaler Studien kei- ne generelle Zunahme der Resistenz. Den- noch finden sich vereinzelt hochresistente Isolate, bei denen Doppelmutationen in einem Bereich des Gyraseenzyms auszumachen sind. Offensichtlich gehen mit solchen Dop- pelmutationen Beeintrachtigungen der nor- malen Enzymfunktion einher, die erst durch
weitere Mutationen kompensiert werden miissen.
Ausgehend von Untersuchungen zur Wech- selwirkung zwischen 4-Chinolon und En- zym-DNS-Komplex sollten Erkenntnisse iiber die Rezeptorstruktur zu erhalten sein, die eine wichtige Grundlage fur die weitere Entwicklung von 4-Chinolonderivaten dar- stellen. Ein Beispiel fur solche Weiterentwick- lungen stellt Bay Y 3118 dar, das bei in vitro
Untersuchungen eine Affinitat zum Rezeptor gezeigt hat, die allen bisherigen Derivaten uberlegen ist. Dadurch konnte nicht nur das bisherige Wirkungsdefizit im grampositiven Bereich kompensiert werden, sondern auch die Resistenzentwicklung verlangsamt wer- den.
Von besonderer Bedeutung fur die Klinik sind vor allem Hygienemahahmen, um eine Ausbreitung resistenter Clone zu verhindern und engmaschige Resistenzepidemiologie, um rechtzeitig auf die Selektion resistenter Klone durch Anderung des Therapieschemas reagieren zu konnen.
Einleitung
Die Pyridon-P-Carbonsauren oder 4-Chino- lone sind synthetische, antibakteriell wirken- de Substanzen, die bereits Ende der Vierziger Jahre beschrieben worden sind. Ca. 80 Mo- lekule sind bislang aus dieser Substanzgruppe patentiert worden. Nalidixinsaure (Abb. 1) war die erste Substanz dieser Gruppe, die 1962 als Harnwegstherapeutikum eingefiihrt wurde [9]. Nalidixinsaure hat den Nachteil, dai3 sie schlecht resorbiert wird, geringe Se- rumkonzentrationen ausweist und ein nur auf gramnegative Bakterien, hauptsachlich Esche- richia coli, beschranktes Wirkungsspektrum hat. Dariiber hinaus kommt es zu einer ra- schen Resistenzentwicklung gegenuber dieser Substanz.
Tabelle 1. MHK-Medianwexte als Ausdruck der Wirksamkeit von Chinolonen
NAL NOR OFLCIP E N 0 FLE LOM TMA SPA BAY
E.coli K.pneumoniae E.cloacae Cfreundii K.oxytoca M.morganii P.mirabilis P.vulgaris Salmonella spp Serratia spp P.aeruginosa S.aureus Staph.koag.0 E.faecalis
2 <0,03 0,03 <0,008 4 <0,03 0,06 <0,008 4 <0,03 0,03 <0,008 4 <0,03 0,03 <0,008 4 <0,03 0,03 <0,008 2 <0,03 0,06 <0,008 4 <0,03 0,06 <0,008 2 <0,03 0,06 <0,008 4 <0,03 0,06 10,008 2 0,12 0,12 0,03 >128 0,25 1 0,06 >128 1 0,25 0,12 >128 0,25 0,12 >128 2 2 0,5
0,03 0,06 0,06 0,03 0,03 0,004 0,06 0,06 0,06 0,06 0,06 0,01 0,03 0,06 0,06 0,06 0,06 0,008 0,03 0,06 0,06 0,03 0,03 0,008 0,06 0,06 0,06 0,03 0,06 0,008 0,06 0,06 0,06 0,25 0,5 0,03 0,125 0,125 0,125 0,25 0,5 0,06 0,06 0,06 0,06 0,5 0,5 0,5 0,06 0,06 0,06 0,06 0,03 0,Ol 0,125 0,25 0,125 0,5 0,6 0,06 0,25 1 0,5 0,25 1 0,25 0,5 0,5 0,5 0,06 0,06 0,03 0,25 0,5 0,5 0,25 0,25 0,03 4 4 4 1 0,5 0,25
Tab. 1. MHK-Medianwerte als Ausdruck der Wirksamkeit von Chinolonen auf verschie- dene grampositive und gramnegative Bakterien
Die MHK-Werte sind definitionsgemafl in mg/L angegeben. Der Medianwert wird an- hand der Verteilung der MHK-Werte fur eine Vielzahl von Einzelvertretern (Stammen) einer Spezies als Scheitelpunkt der Verteilungskurve graphisch ermittelt. (Staph. koag. 0, koagulase-negative Staphylokokken; Salmonella spp., verschiedene Spezies der Gattung Salmonella)
286 Pbarmazie in unserer Zeit / 22. Jahrg. 1993 / Nr. fi-6 0 VCH Verlagsgesellschaft mbH, 69469 Weinbeim, 1993 0048-3664/93/054611-0286 $5.00 + .2;i/O
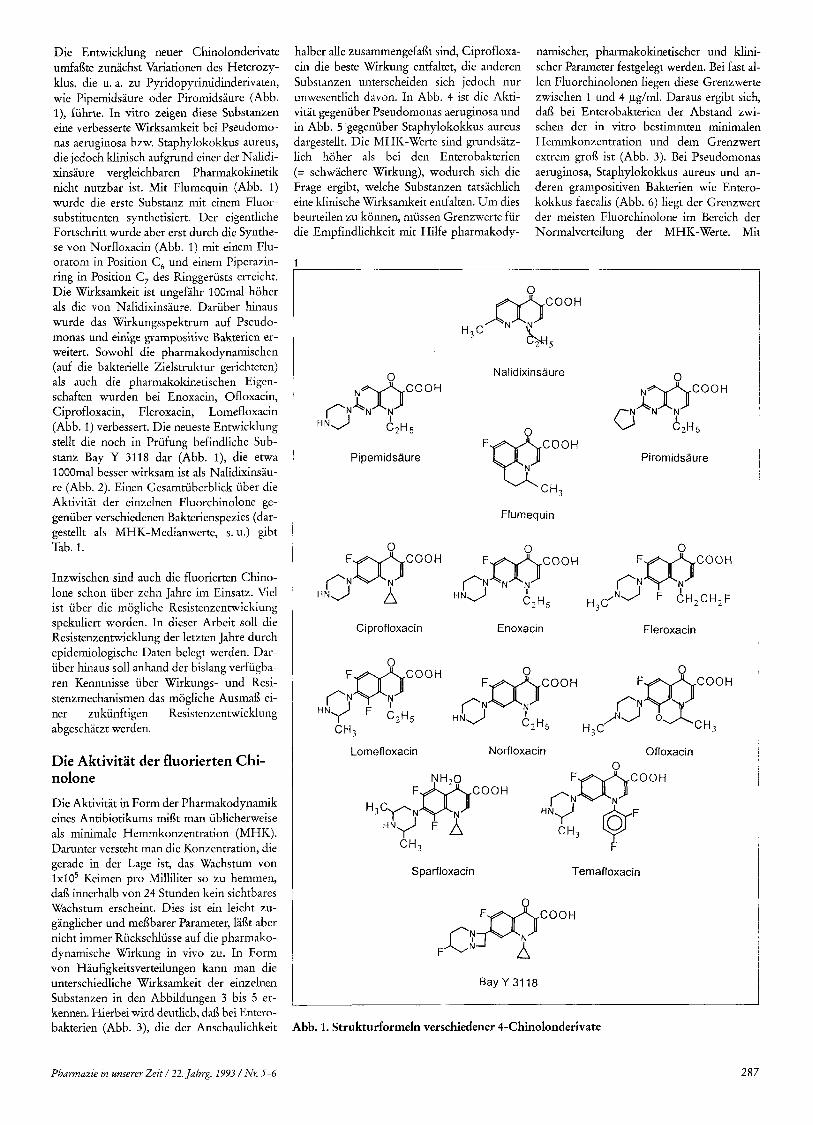
Die Entwicklung neuer Chinolonderivate umfai3te zunachst Variationen des Heterozy- klus, die u. a. zu Pyridopyrimidinderivaten, wie Pipemidsaure oder Piromidsaure (Abb. I), fiihrte. In vitro zeigen diese Substanzen eine verbesserte Wirksamkeit bei Pseudomo- nas aeruginosa bzw. Staphylokokkus aureus, die jedoch klinisch aufgrund einer der Nalidi- xinsaure vergleichbaren Pharmakokinetik nicht nutzbar ist. Mit Flumequin (Abb. 1) wurde die erste Substanz mit einem Fluor- substituenten synthetisiert. Der eigentliche Fortschritt wurde aber erst durch die Synthe- se von Norfloxacin (Abb. 1) mit einem Flu- oratom in Position C , und einem Piperazin- ring in Position C, des Ringgeriists erreicht. Die Wirksamkeit ist ungefahr 100mal hoher als die von Nalidixinsaure. Dariiber hinaus wurde das Wirkungsspektrum auf Pseudo- monas und einige grampositive Bakterien er- weitert. Sowohl die pharmakodynamischen (auf die bakterielle Zielstruktur gerichteten) als auch die pharmakokinetischen Eigen- schaften wurden bei Enoxacin, Ofloxacin, Ciprofloxacin, Fleroxacin, Lomefloxacin (Abb. 1) verbessert. Die neueste Entwicklung stellt die noch in Priifung befindliche Sub- stanz Bay Y 3118 dar (Abb. I), d' ie etwa 1000mal besser wirksam ist als Nalidixinsau- re (Abb. 2). Einen Gesamtiiberblick iiber die Aktivitat der einzelnen Fluorchinolone ge- geniiber verschiedenen Bakterienspezies (dar- gestellt als MHK-Medianwerte, s . u.) gibt Tab. 1 .
Inzwischen sind auch die fluorierten Chino- lone schon iiber zehn Jahre im Einsatz. Vie1 ist iiber die mogliche Resistenzentwicklung spekuliert worden. In dieser Arbeit soll die Resistenzentwicklung der letzten Jahre durch epidemiologische Daten belegt werden. Dar- iiber hinaus soll anhand der bislang verfiigba- ren Kenntnisse iiber Wirkungs- und Resi- stenzmechanismen das mogliche Ausmafl ei- ner zukiinftigen Resistenzentwicklung abgeschatzt werden.
Die Aktivitat der fluorierten Chi- nolone
Die Aktivitat in Form der Pharmakodynamik eines Antibiotikums miflt man iiblicherweise als minimale Hemmkonzentration (MHK). Darunter versteht man die Konzentration, die gerade in der Lage ist, das Wachstum von lx105 Keimen pro Milliliter so zu hemmen, dafl innerhalb von 24 Stunden kein sichtbares Wachstum erscheint. Dies ist ein leicht zu- ganglicher und meflbarer Parameter, laat aber nicht immer Riickschliisse auf die pharmako- dynamische Wirkung in vivo zu. In Form von Haufigkeitsverteilungen kann man die unterschiedliche Wirksamkeit der einzelnen Substanzen in den Abbildungen 3 bis 5 er- kennen. Hierbei wird deutlich, dai3 bei Entero- bakterien (Abb. 3) , die der Anschaulichkeit
halber alle zusammengefai3t sind, Ciprofloxa- cin die beste Wirkung entfaltet, die anderen Substanzen unterscheiden sich jedoch nur unwesentlich davon. In Abb. 4 ist die Akti- vitat gegeniiber Pseudomonas aeruginosa und in Abb. 5 gegeniiber Staphylokokkus aureus dargestellt. Die MHK-Werte sind grundsatz- lich hoher als bei den Enterobakterien (= schwachere Wirkung), wodurch sich die Frage ergibt, welche Substanzen tatsachlich eine klinische Wirksamkeit entfalten. Um dies beurteilen zu konnen, miissen Grenzwerte fur
namischer, pharmakokinetischer und klini- scher Parameter festgelegt werden. Bei fast al- len Fluorchinolonen liegen diese Grenzwerte zwischen 1 und 4 pg/ml. Daraus ergibt sich, dai3 bei Enterobakterien der Abstand zwi- schen der in vitro bestimmten minimalen Hemmkonzentration und dem Grenzwert extrem grofi ist (Abb. 3). Bei Pseudomonas aeruginosa, Staphylokokkus aureus und an- deren grampositiven Bakterien wie Entero- kokkus faecalis (Abb. 6) liegt der Grenzwert der meisten Fluorchinolone im Bereich der
die Empfindlichkeit n i t Hilfe pharmakody- Normalverteilung der MHK-Werte. Mit
Pipemidsaure
Ciprofloxacin
Lomefloxacin
Nalidixinsaure
gAN N ~ c O O H 62H5
Piromidsaure
Flumequin
0
PN FmCooH N I
H , t N J P N F&cooH F CH,CH,F C2H5
H A &
Enoxacin Fleroxacin
Norfloxacin Ofloxacin 0
Sparfloxacin Temafloxacin
F ct&$ooH A
BayY 3118
Abb. 1. Strukturformeln verschiedener 4-Chinolonderivate
Pbamazie in unserer Zeit / 22. Jabrg. 1993 / Nx 5-6 287
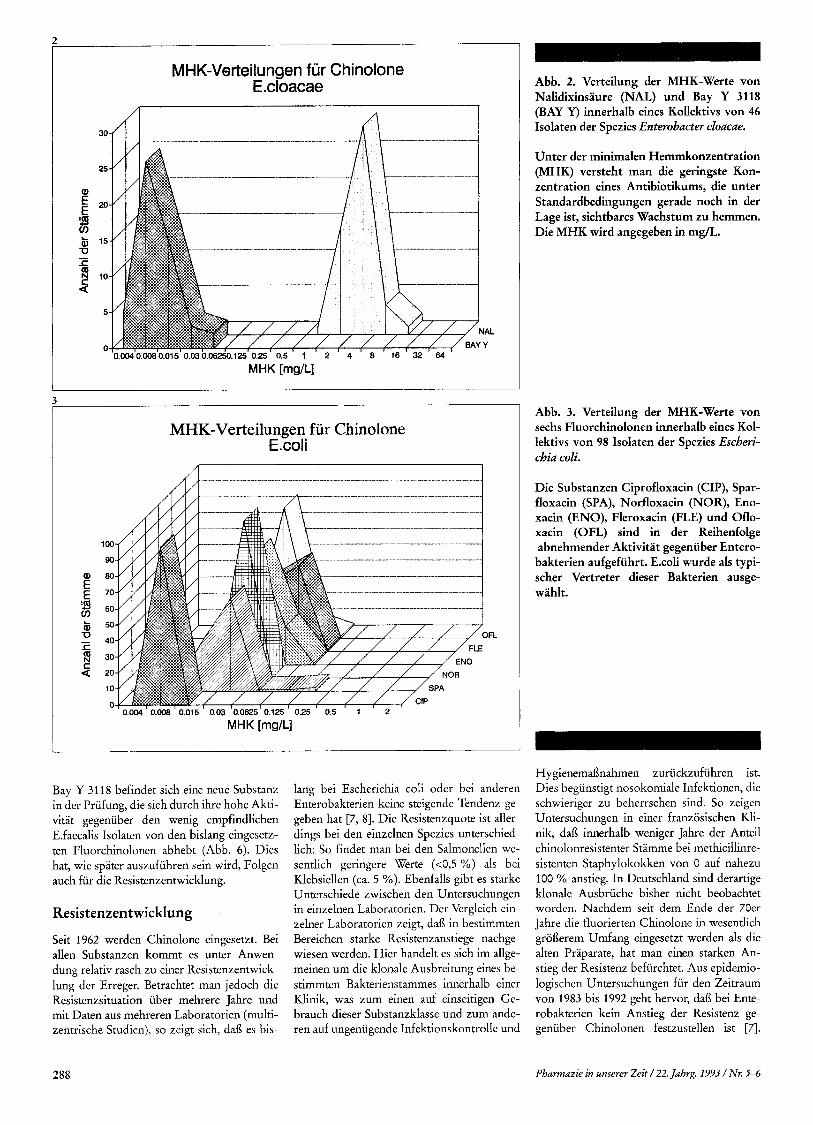
MHK-Verteilungen fur Chinolone E .cloacae
AL
MHK-Verteilungen fur Chinolone E.coli
MHK [mg/L]
Bay Y 3118 befindet sich eine neue Substanz in der Prufung, die sich durch ihre hohe Akti- vitat gegeniiber den wenig empfindlichen E.faecalis Isolaten von den bislang eingesetz- ten Fluorchinolonen abhebt (Abb. 6). Dies hat, wie spater auszufuhren sein wird, Folgen auch fur die Resistenzentwicklung.
Resistenzentwicklung
Seit 1962 werden Chinolone eingesetzt. Bei allen Substanzen kommt es unter Anwen- dung relativ rasch zu einer Resistenzentwick- lung der Erreger. Betrachtet man jedoch die Resistenzsituation uber mehrere Jahre und mit Daten aus mehreren Laboratorien (multi- zentrische Studien), so zeigt sich, dai3 es bis-
lang bei Escherichia coli oder bei anderen Enterobakterien keine steigende Tendenz ge- geben hat [7, 81. Die Resistenzquote ist aller- dings bei den einzelnen Spezies unterschied- lich: So findet man bei den Salmonellen we- sentlich geringere Werte (<0,5 %) als bei Klebsiellen (ca. 5 %). Ebenfalls gibt es starke Unterschiede zwischen den Untersuchungen in einzelnen Laboratorien. Der Vergleich ein- zelner Laboratorien zeigt, dai3 in bestimmten Bereichen starke Resistenzanstiege nachge- wiesen werden. Hier handelt es sich im allge- meinen urn die klonale Ausbreitung eines be- stimmten Bakterienstammes innerhalb einer Klinik, was zum einen auf einseitigen Ge- brauch dieser Substanzklasse und zum ande- ren auf ungenugende Infektionskontrolle und
288
Abb. 2. Verteilung der MHK-Werte von Nalidixinsaure (NAL) und Bay Y 3118 (BAY Y) innerhalb eines Kollektivs von 46 Isolaten der Spezies Enterobacter cloacae.
Unter der minimalen Hemmkonzentration (MHK) versteht man die geringste Kon- zentration eines Antibiotikurns, die unter Standardbedingungen gerade noch in der Lage ist, sichtbares Wachstum zu hemmen. Die MHK wird angegeben in mg/L.
Abb. 3. Verteilung der MHK-Werte von sechs Fluorchinolonen innerhalb eines Kol- lektivs von 98 Isolaten der Spezies Escheri- chia coli.
Die Substanzen Ciprofloxacin (CIP), Spar- floxacin (SPA), Norfloxacin (NOR), Eno- xacin (ENO), Fleroxacin (FLE) und Oflo- xacin (OFL) sind in der Reihenfolge abnehmender Aktivitat gegeniiber Entero- bakterien aufgefuhrt. E.coli wurde als typi- scher Vertreter dieser Bakterien ausge- wahlt.
Hygienemdnahmen zuriickzufiihren ist. Dies begunstigt nosokomiale Infektionen, die schwieriger zu beherrschen sind. So zeigen Untersuchungen in einer franzosischen Kli- nik, dai3 innerhalb weniger Jahre der Anteil chinolonresistenter Stamme bei methicillinre- sistenten Staphylokokken von 0 auf nahezu 100 % anstieg. In Deutschland sind derartige klonale Ausbriiche bisher nicht beobachtet worden. Nachdem seit dem Ende der 70er Jahre die fluorierten Chinolone in wesentlich groi3erem Umfang eingesetzt werden als die alten Praparate, hat man einen starken An- stieg der Resistenz befurchtet. Aus epidemio- logischen Untersuchungen fur den Zeitraum von 1983 bis 1992 geht hervor, dai3 bei Ente- robakterien kein Anstieg der Resistenz ge- genuber Chinolonen festzustellen ist [7].
P b a m z i e in wnserer Zeit / 22. Jabrg. 1993 / Nx 5 4
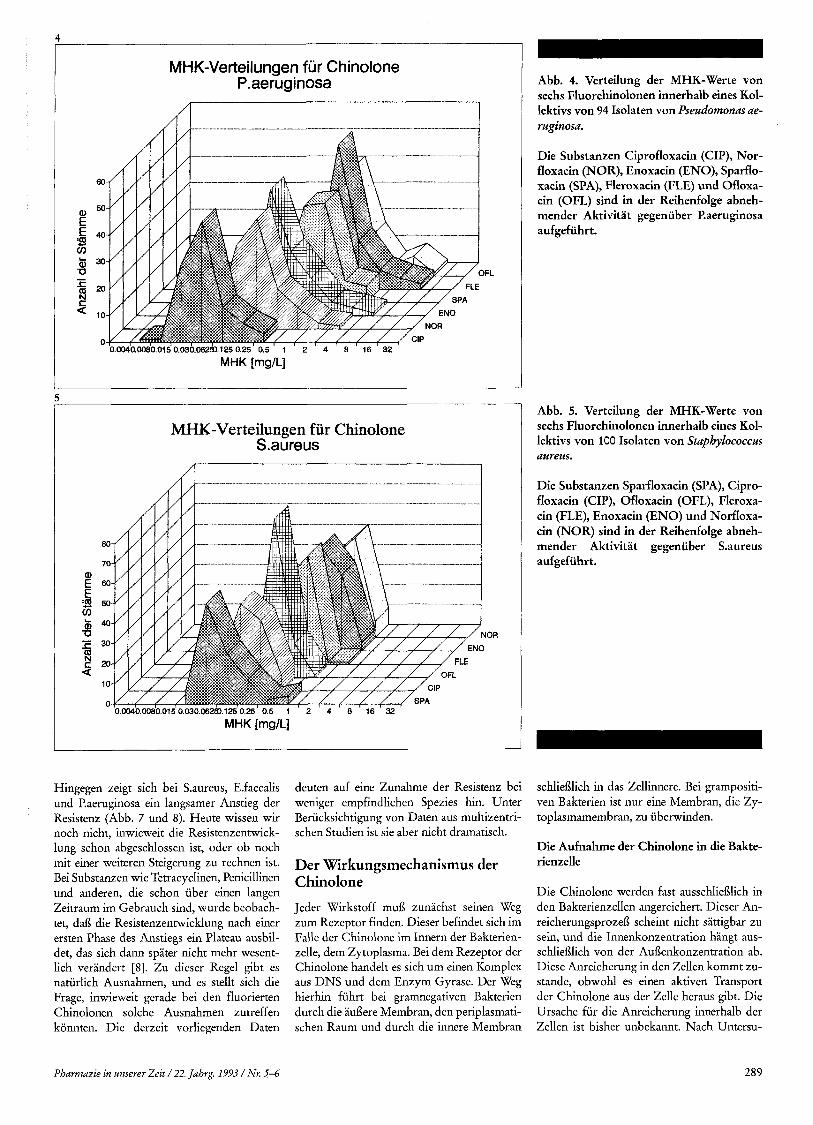
MHK-Vetteilungen fur Chinolone P. aerug inosa
.............. .. ............ _. _I_.__ .- I -- --
. ...
50
40
30 FL
20
10
0 0.0040.0080.015 0.030.0629).125 0.25 0.5 1 2 4 8 16 32
MHK-Verteilungen fur Chinolone S.aureus
MHK [mg/L] -
Hingegen zeigt sich bei S.aureus, E.faecalis und P.aeruginosa ein langsamer Anstieg der Resistenz (Abb. 7 und 8). Heute wissen wir noch nicht, inwieweit die Resistenzentwick- lung schon abgeschlossen ist, oder ob noch mit einer weiteren Steigerung zu rechnen ist. Bei Substanzen wie Tetracyclinen, Penicillinen und anderen, die schon iiber einen langen Zeitraum im Gebrauch sind, wurde beobach- tet, dafl die Resistenzentwicklung nach einer ersten Phase des Anstiegs ein Plateau ausbil- det, das sich dann spater nicht mehr wesent- lich verandert [8]. Zu dieser Regel gibt es natiirlich Ausnahmen, und es stellt sich die Frage, inwieweit gerade bei den fluorierten Chinolonen solche Ausnahmen zutreffen konnten. Die derzeit vorliegenden Daten
deuten auf eine Zunahme der Resistenz bei weniger empfindlichen Spezies hin. Unter Berucksichtigung von Daten aus multizentri- schen Studien ist sie aber nicht dramatisch.
Der Wirkungsmechanismus der Chinolone
Jeder Wirkstoff mu6 zunachst seinen Weg zum Rezeptor finden. Dieser befindet sich im Falle der Chinolone im Innern der Bakterien- zelle, dem Zytoplasma. Bei dem Rezeptor der Chinolone handelt es sich um einen Komplex aus DNS und dem Enzym Gyrase. Der Weg hierhin fiihrt bei gramnegativen Bakterien durch die aufiere Membran, den periplasmati- schen Raum und durch die innere Membran
Abb. 4. Verteilung der MHK-Werte von sechs Fluorchinolonen innerhalb eines Kol- lektivs von 94 Isolaten von Pseudomonas ae- ruginosa.
Die Substanzen Ciprofloxacin (CIP), Nor- floxacin (NOR), Enoxacin (ENO), Sparflo- xacin (SPA), Fleroxacin (FLE) und Ofloxa- cin (OFL) sind in der Reihenfolge abneh- mender Aktivitat gegeniiber P.aeruginosa aufgefiihrt.
Abb. 5. Verteilung der MHK-Werte von sechs Fluorchinolonen innerhalb eines Kol- lektivs von IOO Isolaten von Staphylococcus aureus.
Die Substanzen Sparfloxacin (SPA), Cipro- floxacin (CIP), Ofloxacin (OFL), Fleroxa- cin (FLE), Enoxacin (ENO) und Norfloxa- cin (NOR) sind in der Reihenfolge abneh- mender Aktivitat gegeniiber S.aureus aufgefiihrt.
schliefilich in das Zellinnere. Bei grampositi- ven Bakterien ist nur eine Membran, die Zy- toplasmamembran, zu uberwinden.
Die Aufnahme der Chinolone in die Bakte- rienzelle
Die Chinolone werden fast ausschliefilich in den Bakterienzellen angereichert. Dieser An- reicherungsprozefi scheint nicht sattigbar zu sein, und die Innenkonzentration hangt aus- schliefilich von der Aufienkonzentration ab. Diese Anreicherung in den Zellen kommt zu- stande, obwohl es einen aktiven Transport der Chinolone aus der Zelle heraus gibt. Die Ursache fur die Anreicherung innerhalb der Zellen ist bisher unbekannt. Nach Untersu-
Pharmazie in unserer Zeit / 22. Jahrg. 1993 / Nx 5.4 289
6
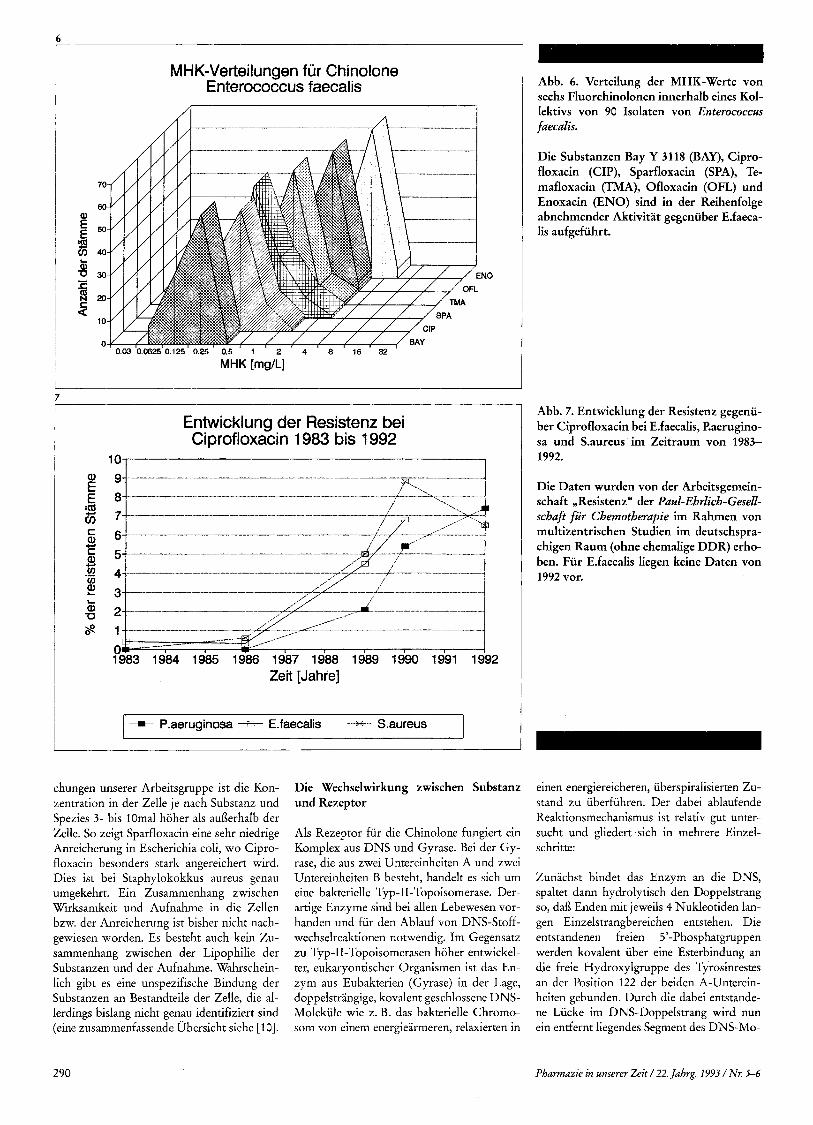
MHK-Verteilungen fur Chinolone En terococcus faecal is
j MHK [mg/L]
7
Entwicklung der Resistenz bei Ciprofloxacin 1983 bis 1992
1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 Zeit [Jahre]
P.aeruginosa - E.faecalis Saureus i chungen unserer Arbeitsgruppe ist die Kon- zentration in der Zelle je nach Substanz und Spezies 3- bis 10mal hoher als auflerhalb der Zelle. So zeigt Sparfloxacin eine sehr niedrige Anreicherung in Escherichia coli, wo Cipro- floxacin besonders stark angereichert wird. Dies ist bei Staphylokokkus aureus genau umgekehrt. Ein Zusammenhang zwischen Wirksamkeit und Aufnahme in die Zellen bzw. der Anreicherung ist bisher nicht nach- gewiesen worden. Es besteht auch kein Zu- sammenhang zwischen der Lipophilie der Substanzen und der Aufnahme. Wahrschein- lich gibt es eine unspezifische Bindung der Substanzen an Bestandteile der Zelle, die al- lerdings bislang nicht genau identifiziert sind (eine zusammenfassende Ubersicht siehe [lo].
Abb. 6. Verteilung der MHK-Werte von sechs Fluorchinolonen innerhalb eines Kol- lektivs von 90 Isolaten von Enterococcm faecalis.
Die Substanzen Bay Y 3118 (BAY), Cipro- floxacin (CIP), Sparfloxacin (SPA), Te- mafloxacin (TMA), Ofloxacin (OFL) und Enoxacin (ENO) sind in der Reihenfolge abnehmender Aktivitat gegeniiber EAaeca- lis aufgefiihrt.
Abb. 7. Entwicklung der Resistenz gegenii- ber Ciprofloxacin bei E.faecalis, P.aerugino- sa und S.aureus im Zeitraum von 1983- 1992.
Die Daten wurden von der Arbeitsgemein- schaft ,,Resistenz" der Paul-Ehrlich-Gesell- schaji fiir Chemotherapie im Rahmen von multizentrischen Studien im deutschspra- chigen Raum (ohne ehemalige DDR) erho- ben. Fur E.faecalis liegen keine Daten von 1992 vor.
Die Wechselwirkung zwischen Substanz und Rezeptor
Als Rezeptor fur die Chinolone fungiert ein Komplex aus DNS und Gyrase. Bei der Gy- rase, die aus zwei Untereinheiten A und zwei Untereinheiten B besteht, handelt es sich um eine bakterielle Typ-11-Topoisomerase. Der- artige Enzyme sind bei allen Lebewesen vor- handen und fur den Ablauf von DNS-Stoff- wechselreaktionen notwendig. Im. Gegensatz zu Typ-11-Topoisomerasen hoher entwickel- ter, eukaryontischer Organismen ist das En- zym aus Eubakterien (Gyrase) in der Lage, doppelstrangige, kovalent geschlossene DNS- Molekule wie z. B. das bakterielle Chromo- som von einem energiearmeren, relaxierten in
einen energiereicheren, uberspiralisierten Zu- stand zu uberfiihren. Der dabei ablaufende Reaktionsmechanismus ist relativ gut unter- sucht und gliedert .sich in mehrere Einzel- schritte:
Zunachst bindet das Enzym an die DNS, spaltet dann hydrolytisch den Doppelstrang so, dai3 Enden mit jeweils 4 Nukleotiden lan- gen Einzelstrangbereichen entstehen. Die entstandenen freien 5'-Phosphatgruppen werden kovalent uber eine Esterbindung an die freie Hydroxylgruppe des Tyrosinrestes an der Position 122 der beiden A-Unterein- heiten gebunden. Durch die dabei entstande- ne Lucke im DNS-Doppelstrang wird nun ein entfernt liegendes Segment des DNS-Mo-
290 P h a m z i e in unserer Zeit / 22. Jahrg. 1993 / N,: J-6
9
M H K-Vertei I u ng vo n Ci prof loxaci n P. aerug i nosa
... " I .. ..
0 0,008 0,03 0,12 0,5 2 8 32
0,016 0,06 0,25 1 4 16 MHK [mg/l]
Abb. 8. Prozentuale Verteilung der MHK-Werte von Ciprofloxacin fur Isolate von l? aeru- ginosa aus den Jahren 1983,1986,1989 und 1990.
Fur die Jahre 1983 und 1986 ergibt sich eine einfache Gauss'sche Verteilung mit einem Me- dianwert bei 0,06-0,125 mg/L und einem relativ geringen Anteil weniger empfindlicher Isolate, der sich nicht von der Gesamtpopulation abhebt. In den Jahren 1989 und 1990 fin- det sich dagegen eine bimodale Verteilung, in der ein groi3er Anteil empfindlicher Isolate (Medianwert 0,125 mg/I+) sich von einem kleinen Anteil resistenter Isolate (durchschnittli- che MHK bei 16 mg/L) abhebt. Aus der Zunahme des zweiten Teils der Population ergibt sich eine Verbreiterung des MHK-Verteilungsdiagramms.
lekuls durchgeschleust. Dieser Vorgang ent- spricht topologisch der Entwindung der bei- den umeinandergewundenen DNS-Strange.
Zum Abschlufi werden die urspriinglichen Phosphodiesterbindungen der DNS regene- riert. Bei diesen Reaktionsschritten bleibt die in der Phosphodiesterbindung der DNS kon- servierte Energie erhalten. Jedoch wird durch eine in der B-Untereinheit vorhandene ATPa- se ATP hydrolysiert, um das Enzym wieder in die Ausgangskonformation zu iiberfiihren.
Durch die Reaktionsfolge wird erreicht, dafi die Anzahl der Windungen der beiden DNS- Strange umeinander, die strukturbedingt nor- malerweise bei einem Wert von 10,4 Basen- paaren pro Helixwindung liegt, schrittweise verringert wird. Die dabei in dem DNS- Ringmolekiil erzeugte Torsionsspannung fiihrt letztendlich dam, dafi sich die Helixachse, die sich im relaxierten Zustand in einer Ebene befindet, im Raum verdreht und das Molekul in den energiereicheren iiberspi- ralisierten Zustand ubergeht (Abb. 9) (aus- fiihrliche Darstellung siehe [l] und [17]).
Damit wird - unter Beteiligung histonahnli-
cher DNS-Bindeproteine - ermoglicht, d d ein bakterielles Chromosom, das im ausge- streckten Zustand etwa 1000mal langer ist als eine Bakterienzelle, in dieser untergebracht werden kann.
Die Chinolone binden nun nach Untersu- chungen von Shen und Pernet [13] an den DNS-Gyrase-Komplex, jedoch weder an DNS noch an Gyrase alleine. Es wird ange- nommen, dafi der genaue Bindungsort der Chinolone im Komplex eine sogenannte Chi- nolontasche ist, an deren Ausbildung die Un- tereinheiten A sowie Bereiche der DNS betei- ligt sind (Abb. l la) . Vermutlich tragen die kurzen einzelstrangigen DNS-Enden zur Chinolonbindung an den Komplex bei.
Nach Untersuchungen unserer Arbeitsgrup- pe lassen sich die Affinitaten der einzelnen Chinolone an den Enzym-DNS-Komplex gut mit der Wirksamkeit der Substanzen auf die gesamte Zelle korrelieren (Abb. 10). Ein ternarer Komplex aus DNS, Gyrase und Ci- profloxacin besitzt demnach eine Halbwerts- zeit in vitro von ungefahr zweieinhalb Stun- den [18]. Legt man die gleiche Stabilitat in vivo zugrunde, so wird verstandlich, dafi
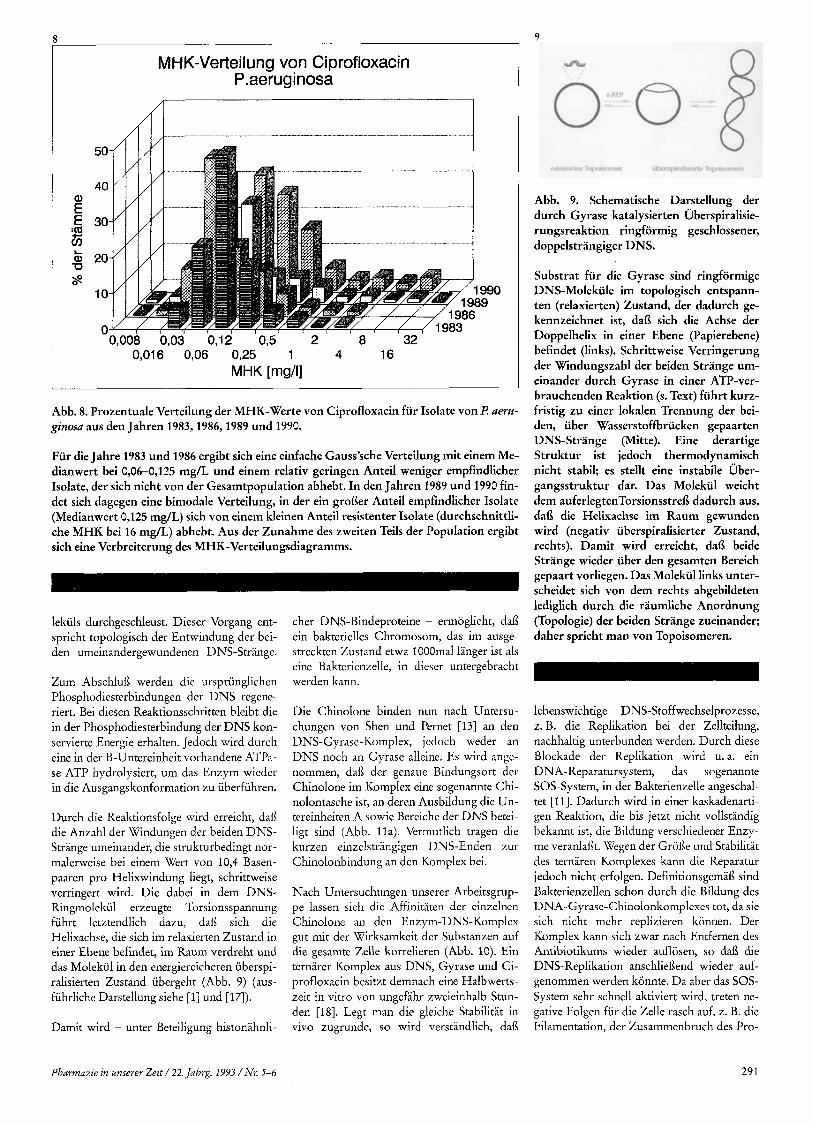
Abb. 9. Schematische Darstellung der durch Gyrase katalysierten Iliberspiralisie- rungsreaktion ringformig geschlossener, doppelstrangiger DNS.
Substrat fur die Gyrase sind ringformige DNS-Molekiile im topologisch entspann- ten (relaxierten) Zustand, der dadurch ge- kennzeichnet ist, dai3 sich die Achse der Doppelhelix in einer Ebene (Papierebene) befindet (links). Schrittweise Verringerung der Windungszahl der beiden Strange um- einander durch Gyrase in einer ATP-ver- brauchenden Reaktion (s. Text) fuhrt kurz- fristig zu einer lokalen Trennung der bei- den, iiber Wasserstoffbriicken gepaarten DNS-Strange (Mitte). Eine derartige Struktur ist jedoch thermodynamisch nicht stabil; es stellt eine instabile Uber- gangsstruktur dar. Das Molekul weicht dem auferlegtenTorsionsstrei3 dadurch aus, dai3 die Helixachse im Raum gewunden wird (negativ uberspiralisierter Zustand, rechts). Damit wird erreicht, dal3 beide Strange wieder uber den gesamten Bereich gepaart vorliegen. Das Molekul links unter- scheidet sich von dem rechts abgebildeten lediglich durch die raumliche Anordnung (Topologie) der beiden Strange zueinander; daher spricht man von Topoisomeren.
lebenswichtige DNS-Stoffwechselprozesse, z. B. die Replikation bei der Zellteilung, nachhaltig unterbunden werden. Durch diese Blockade der Replikation wird u. a. ein DNA-Reparatursystem, das sogenannte SOS-System, in der Bakterienzelle angeschal- tet [I l l . Dadurch wird in einer kaskadenarti- gen Reaktion, die bis jetzt nicht vollstandig bekannt ist, die Bildung verschiedener Enzy- me veranlai3t. Wegen der Grofie und Stabilitat des ternaren Komplexes kann die Reparatur jedoch nicht erfolgen. Definitionsgemafi sind Bakterienzellen schon durch die Bildung des DNA-Gyrase-Chinolonkomplexes tot, da sie sich nicht mehr replizieren konnen. Der Komplex kann sich zwar nach Entfernen des Antibiotikums wieder auflosen, so dai3 die DNS-Replikation anschliei3end wieder auf- genommen werden konnte. Da aber das SOS- System sehr schnell aktiviert wird, treten ne- gative Folgen fur die Zelle rasch auf, z. B. die Filamentation, der Zusammenbruch des Pro-
Phamzazie in unserer Zeit / 22. Jahrg. 1993 / N,: 5-6 291

Affinitat von Chinolonen zu DNA-Gyrase
'I ... ............................................ ........................... ........
tonengradienten und schliei3lich die Lyse der Bakterienzelle. Bei vielen Chinolonen sterben die Zellen innerhalb von einer Stunde schon ab.
Die Aktivierung des SOS-Systems ist aller- dings keine notwendige Bedingung fur die bakterizide Wirkung der Chinolone, da auch solche Zellen unter Chinoloneinwirkung ab- sterben, die aufgrund eines genetischen De- fekts das SOS-System nicht aktivieren kon- nen. Die gesamten Mechanismen, die zum Zelltod fiihren, sind derzeit noch nicht ver- standen und daher Gegenstand intensiver Forschung.
Resistenz - eine Begriffsbestimmung
Die Resistenz eines Krankheitserregers ge- geniiber einem Antibiotikum ist klinisch so definiert, dai3 die Substanzen eine experimen- tell ermittelbare minimale Hemmkonzentra- tion (MHK) in einer Hohe aufweisen, die ei- nen Therapieerfolg unwahrscheinlich erschei- nen lai3t. Um die Entscheidung, ob ein Erreger resistent ist oder nicht, nicht zu einer Ermessensfrage werden zu lassen, werden in der DIN 58 940 Grenzwerte definiert, die der Beurteilung zugrunde gelegt werden. Dieser rein klinische Resistenzbegriff ist fur die Bio- logie nicht ausschlaggebend. Vielmehr mufi sich der Begriff ,,Resistenz" daran orientie- ren, ob die Erreger eine Eigenschaft hinzuge- wonnen haben, die es ihnen ermoglicht, hohere Konzentrationen einer Substanz zu tolerieren als ihre Elternzellen (erworbene oder sekundare Resistenz). Die MHK-Werte einer Substanz fur einzelne Vertreter einer Spezies liegen ublicherweise sehr nah beiein-
ander. Tragt man die MHK-Werte gegen die Haufigkeit bei den einzelnen Stammen auf, ergibt sich eine Gauss'sche Verteilung (vgl. Abb. 8). Bakterien, deren MHK auflerhalb dieser Verteilung liegt, sind damit aus biologi- scher Sicht als resistent zu bezeichnen. Solche Starnme mussen aber noch nicht klinisch resi- stent sein. Im Falle der Fluorchinolone liegen die MHK-Werte fur Enterobakterien derartig niedrig, dafi die durch eine oder unter Um- standen sogar zwei Mutationen erreichte Stei- gerung der MHK klinisch nicht bedeutsam ist. Staphylokokken und Pseudomonaden hingegen konnen aufgrund der naturlicher- weise geringeren Empfindlichkeit bereits durch einen einzigen Mutationsschritt klini- sche Resistenz erlangen (vgl. Tab. 1 und Abb. 8).
Der Resistenzmechanismus
Grundsatzlich gibt es drei Mechanismen, die zur Resistenz von Bakterien gegenuber Che- motherapeutika fiihren:
1. die Substanz wird durch Enzyme der Bak- terien inaktiviert;
2. der Zugang der Substanz zum Rezeptor ist beeintrachtigt, z. B. weil die Penetration durch die aui3ere Membran der Bakterien ver- hindert wird;
3. die Rezeptorstruktur ist so verandert, dai3 die Substanz nicht mehr bindet.
Bei den Chinolonen ist ein enzymatischer Abbau der Substanzen bisher nicht beobach- tet worden. Alle bisher identifizierten Resi- stenzmechanismen umfassen entweder eine
Abb. 10. Vergleichende Gegeniiberstellung der pharmakodynamischen Aktivitat ver- schiedener Chinolonderivate.
Die relative Affinitat von zehn verschiede- nen Chinolonderivaten - Bay Y 3118 (BAY), Ciprofloxacin (CIP), Sparfloxacin (SPA), Norfloxacin (NOR), Fleroxacin (FLE), Ofloxacin (OFL), Lomefloxacin (LOM), Enoxacin (ENO), Nalidixinsaure (NAL) und Pipemidsaure (PIP) - zu einem Komplex aus DNS (Plasmid pBR322) und Gyrase (aus E.coli K-12) wurde als das Aus- mai3 der Verdrangung eines radioaktiv markierten Fluorchinolonderivates (Enro- floxacin) aus der Komplexbindung durch die jeweilige Substanz bestimmt [18]. Zwi- schen fluorierten und unfluorierten Sub- stanzen (NAL, PIP) ist ein deutlicher Un- terschied erkennbar.
Veranderung des Rezeptors oder eine Beein- trachtigung des Zugangs.
Kreuzresistenz
Von wenigen Ausnahmen abgesehen gibt es eine absolute Kreuzresistenz zwischen allen Chinolonen. Darunter versteht man den gleichzeitigen Anstieg der Resistenz gegenu- berallen Chinolonen. Das bedeutet nicht im- mer, dai3 es damit auch zu einer klinischen Kreuzresistenz kommt, da diese ja strikt durch Grenzwerte bemessen wird. Zu dieser Regel gibt es einige Ausnahmen, die aber kli- nisch keine Rolle spielen.
Die Veranderung der bakteriellen Mem- bran
Fur die Aufnahme von Chinolonen in gram- negative Bakterien sind die Porine, porenbil- dende Proteine, die in der aufleren Membran liegen, von groi3er Bedeutung. Wenn durch Mutation die Struktur der Porine oder deren Expression so verandert ist, dai3 sie den Durchtritt der Chinolone durch die aufiere Mernbran einschranken, bedeutet das fur die minimale Hemmkonzentration der Chinolo- nen eine Erhohung um ein bis drei Stufen. Zwar hat eine solche Mutation alleine damit nur einen geringen, klinisch meist unbedeu- tenden Effekt, im Zusammenwirken mit an- deren Mutationen kann ihnen jedoch groflere Bedeutung zukommen [6]. Da diese Porine gleichzeitig auch fur die Aufnahme anderer Substanzen wichtig sind, reduziert sich die Empfindlichkeit gegenuber manchen P-Lak- tamantibiotika, Tetrazyklinen und Chloram- phenicol (zusammenfassende Ubersicht uber Penetrationsmutanten bei [12]). Bei Staphylo-
292 Pharmazie in unserer Zeit / 22. Jahrg. 1993 / Nr. 5-6

l l a l l c
Abb. Ila. Modell der Bindung von Chino- lonmolekiilen an einen Komplex aus DNS und empfindlicher Gyrase von E.coli.
Dargestellt ist ein 4 Nukleotide langes, ein- zelstrangiges Ende des von Gyrase gespal- tenen DNS Doppelstranges, das durch eine Phosphatesterbindung mit der freien 5'- Phosphatgruppe an die Hydroxylgruppe des Tyrosinrestes 122 in der einen der bei- den A-Untereinheiten der Gyrase gebun- den ist. Ein benachbarter Bereich derselben Untereinheit formt eine sogenannte ,,Chi- nolontasche", in die Chinolonmolekiile hineinschliipfen konnen. Man geht davon aus, dai3 die Chinolonbindung durch Wechselwirkungen sowohl mit Teilen des einzelstrangigen DNS-Bereiches als auch mit mehreren Aminosauren in der Bin- dungstasche stabilisiert wird. Wahrscheinli- che Kandidaten sind Serin an Pos. 83 und Aspartat an Pos. 87 (s. Text). AuSerdem ist nach eigenen Daten eine Beteiligung der B- Untereinheit dabei nicht auszuschliei3en [5]. Die hohe Affinitat der Chinolonmo- lekiile zu dieser Bindungstasche driickt sich in einer sehr hohen Empfindlichkeit von Zellen mit einer derartigen (normalemp- findlichen) Gyrase aus (MHK von 0,008 mg/L).
Abb. Ilb. Modell der Bindung von Chino- lonmolekiilen an einen Komplex aus DNS und Gyrase von E.coli, die durch eine Ein- Schritt-Mutation an Aminosaure 83 der A- Untereinheiten verandert ist.
Gegeniiber der Abb. l l a hat eine Mutation zu einem Austausch der Aminosaure Serin an Pos. 83 zu Leucin gefiihrt. Nach der von uns entwickelten Modellvorstellung fiihrt dies zu einer Schwachung der Chinolon- bindung an die Bindungstasche. Fur die Ausbildung eines ternaren Komplexes aus DNS, Gyrase und Chinolon ist eine hohere Ligandenkonzentration erforderlich. Fur E.coli Zellen rnit einer derartigen Mutation werden daher um einen Faktor von 16 er- hohte MHK-Werte gefunden (0,125 mg/L).
Abb. llc. Modell der Bindung von Chino- lonmolekiilen an einen Komplex aus DNS und Gyrase von E.coli, die durch eine Dop- pelmutation an den Aminosauren 83 und 87 der A-Untereinheiten verandert ist.
Durch eine zusatzliche Mutation, die zum Austausch der Aminosaure an Pos. 87 von Aspartat zu Glycin fiihrt, wird die Chino- lonbindung an die Bindungstasche weiter geschwacht. In vitro sind ca. mehr als 2000fach hohere Konzentrationen an Chi- nolon erforderlich, um das Enzym sichtbar in seiner Aktivitat zu hemmen. Ein Isolat von E.coli mit einer solchen Doppelmuta- tion weist eine gegeniiber einem normal- empfindlichen Stamm um das 16 OOOfach erhohte MHK (128 mg/L) auf [4]. Untersu- chungen an in vitro selektierten Mutanten mit derselben Doppelmutation in der Gyrase A-Untereinheit deuten allerdings auch auf Veranderungen der Funktions- fahigkeit des Enzyms hin.
Tabelle 2. Ergebnisse eines Tests auf Dominanz des gyrA-Gens fur eine chinolonempfind- liche Gyrase A-Untereinheit iiber das resistente Allel.
Angegeben sind die Empfindlichkeiten (als MHK-Werte in mg/L) fur Ciprofloxacin (CIP) und Nalidixinsaure (NAL) eines normal chinolonempfindlichen Stammes von E.coli K-12 (C600S) und seines durch Mutation im gyrA-Gen chinolonresistenten Abkommlings (C600SN) in Ab- und Anwesenheit des Plasmides pBP517. Dieses Plasmid - ein ringformi- ger DNS-Doppelstrang, der selbstandig replizieren kann, tragt eine Kopie des gyrA-Gens fur eine chinolonempfindliche Gyrase A-Untereinheit. Die Anwesenheit des Plasmides pBP517 wirkt sich nicht auf die Chinolonempfindlichkeit von C600S Zellen aus. Dagegen wird die Chinolonempfindlichkeit von C600SN Zellen, die durch eine gyrA-Mutation un- empfindlicher gegen Chinolone geworden sind, in Anwesenheit des chinolonempfindli- chen gyrA-Allels auf Plasmid pBP517 wieder so empfindlich wie der Ausgangsstamm C600S: Das empfindliche gyrA-Allel ist dominant iiber das resistente [2].
P b a m z i e in unserer Zeit / 22. Jabrg. 1993 / Nr. 5-6 293

kokken wurde vor kurzem eine Mutation (norA) identifiziert, die zu einem erhohten Efflux der Chinolone aus der Zelle fiihrt und damit ebenfalls zu einer Verringerung des Netto-Einstroms von Chinolonen in die Zel- len fiihrt. Vermutlich ist dabei ein naturlicher- weise vorhandenes Effluxsystem verandert U61.
Die Modifikation des Rezeptors
Als Rezeptor fur die Chinolone wurden die DNA-Gyrase-Komplexe bereits oben be- schrieben, wobei friihzeitig klar war, dai3 Ver- anderungen in den Gyrase-Untereinheiten A oder B fur die Resistenz gegeniiber Chinolo- nen verantwortlich sind [14, 191.
Andere Autoren beschrieben bereits vor die- ser Zeit das Dominanzphanomen des geneti- schen Merkmals fur Chinolonempfindlich- keit iiber das der Chinolonresistenz [2]. Dies bedeutet, dai3 ein Bakterienstamm, der durch eine Veranderung in einem der Gene fur die Gyrase-Untereinheiten A oder B (gyrA oder gyrB) resistent gegeniiber Chinolonen ist, wieder empfindlich wird, wenn eine zweite - fur Chinolonempfindlichkeit codierende - Genkopie der jeweiligen Untereinheit z. B. mittels eines Plasmids in die Zelle einge- schleust wird (Tab. 2). Dies 1a8t sich leicht verstehen, wenn man bedenkt, dai3 nur ein einziges chinolonempfindliches Gyrasemo- lekul, das in Anwesenheit von Chinolon in ei- nem stabilen, ternaren Komplex an die DNS gebunden bleibt, ausreicht, um die Replikati- on der DNA zu verhindern und damit den Zelltod einzuleiten (s. 0.). Mittlerweile sind eine Vielzahl von Veranderungen in den Gy- rasegenen gyrA und gyrB bei resistenten Bak- terien identifiziert worden (Tab. 3). Die hier- bei auftretenden Veranderungen im Protein werden fur die verringerte Wirksamkeit der Chinolone verantwortlich gemacht.
Untersuchungen unserer Arbeitsgruppe zei- gen, da8 bei hochresistenten Mutanten der normalerweise sehr empfindlichen Entero- bakterien (E.coli und Salmonella typhimuri- um) gleichzeitig zwei Aminosauren, Serin-83 (zu Leucin) und Aspartat-87 (zu Glycin bzw. Asparagin), ausgetauscht sind, was mit einem Anstieg der MHK um mehr als das 10 000fa- che verbunden ist [3, 4, 51. Schematisch ist diese Veranderung in den Abbildungen 1 la-c dargestellt: Vermutlich miissen die Chinolon- molekiile in die Chinolontasche innerhalb des Gyrase-DNS-Komplexes hineinschlup- fen und hier gleichzeitig an die einstrangige DNA und Bereiche des Enzyms (z. B. Serin- 83 und/oder Aspartat-87) binden (Abb. 1 la). Durch eine einzelne Mutation wird die Affi- nitat geringfugig verandert (Abb. Ilb), durch zwei Mutationen wird sie so stark verandert, dai3 die Wirkung iiber 10 OOOmal schlechter ist als beim Wildtypstamm (Abb. llc).
294
Die Mutationsrate
Um festzustellen, wie haufig Mutationen bei Bakterienstammen in Richtung einer be- stimmten Eigenschaft auftreten, kann man die Mutationsraten bestimmen, indem man die Zahl der Bakterien innerhalb einer Population ermittelt, die diese gesuchte Eigenschaft ent- wickeln. Hierbei konnte unsere Arbeitsgrup- pe feststellen, da8 bei E.coli die Mutation der Aminosaure 83 vom Serin zum Leucin mit ei- ner Frequenz von lo-'' auftritt. Die Mutation der Aminosaure 87, Aspartat zu Glycin, tritt ebenfalls mit dieser Haufigkeit auf. Daher sollte man annehmen, dai3 mit einer Frequenz von hochresistente Mutanten entstehen konnen. In vitro ist es nicht moglich, Mutan- ten in zwei aufeinanderfolgenden Schritten zu isolieren, die beide Mutationen im gyrA Gen besitzen. Nach unseren Untersuchungen (noch unveroffentlicht) trat bei der Selektion solcher gyrA Doppelmutanten zusatzlich eine Mutation auf, die zu Veranderungen in der Zellwand fiihrt. Diese Dreifach-Mutanten zeigen ein gegeniiber dem chinolonempfindli- chen Ausgangsstamm deutlich verlangsamtes Wachstum, was sich in vivo als nachteilig er- weisen wiirde. Offensichtlich ist mindestens eine zusatzliche Mutation notwendig, um die Lebensfahigkeit der Bakterien herzustellen.
Das bedeutet, dai3 in vitro die Veranderung der Bakterienzelle zur hochgradigen Resi- stem gegeniiber Fluorchinolonen mit einer Frequenz von hochstens auftreten kann.
Da derartig groi3e Bakterienpopulationen gar nicht existieren, miissen die Mutationen nacheinander in kleineren Populationen auf- treten. Schliei3t man von diesen in vitro Un- tersuchungen auf die Situation in vivo, so be- deutet das, daf3 erst eine konsequente Selek- tion zu dieser Resistenz fiihren kann. Eine derartige Resistenzentwicklung sollte aber zu verhindern sein, wenn das therapeutische Re- gime geandert wird, bevor der letzte Mutati- onsschritt erreicht wurde.
Dagegen kann bei Bakterien, deren MHK nahe am Grenzwert liegt, eine Mutation be- reits in einem Schritt zu einer klinisch bedeut- samen Resistenz fiihren. Dies ist bei Entero- kokken, Staphylokokken und Paeruginosa der Fall ( s . 0.). Hier kann es unter einer nor- malen Therapie zur Entwicklung klinisch re- sistenter Bakterienstamme kommen, die sich aus dem Infektionsherd rekrutieren. Dies ist jedoch selten, da iiblicherweise durch die Chinolontherapie die Zahl der lebenden Zel- len derartig reduziert wird, da8 die Popula- tionsstarke niedriger ist als die fur die Selekti- on solcher Mutanten notwendigen Zellzah- len.
Ausblick
Die Resistenz gegeniiber Chinolonen ist nicht plasmidkodiert. Damit ist eine horizontale Ausbreitung von einer Bakterienzelle auf eine andere derselben Generation oder sogar von einer Spezies auf eine andere nicht gegeben.
Tabelle 3. Mutationen im gyrA-Gen von Escherichia coli mit Auswirkung auf die Chino- lonempfindlichkeit
~
Pas. (Nt) Basenaustausch POS. (As) Aminosaureaustausch MHK Cip [mg/L]
__
199 241 247 248 248 250 259 260 317 318
G - T G + T T + G C + T C + G G + C G + A A - T A + G G - T
- 67 81 83 83 83 84 87 87
106 106
A L A + S E R G L Y + C Y S S E R + A L A S E R + L E U S E R + T R P A L A + P R O A S P + A S N A S P + V A L G L N + A R G G L N + H I S
0,0125 0,05 091 0,055 0,39
071 0,2 0,06 0,05 0.05
0,5
Tab. 3. Mutationen im gyrA-Gen von E.coli mit Auswirkung auf die Chinolonempfind- lichkeit
Die Angaben zur Position des Nukleotids (Nt), das bei einer Mutante zu einem Austausch einer Aminosaure (As) fiihrt, beziehen sich auf die DNS-Sequenz der codierenden Region des gyrA-Gens von E.coli K-12 [15]. Nukleotid 1 ist das Adenin irn ersten Codon (ATG) des Gens. Die Empfindlichkeit der jeweiligen Mutanten gegeniiber Ciprofloxacin (CIP) ist als MHK angegeben. Die Daten sind teilweise entnommen aus [20].
P h a m z i e in unserer Zeit / 22. Jahrg. 1993 / Nx >4

Die Resistenz gegenuber Chinolonen entsteht ausschliefllich aufgrund von Mutationen und gleichzeitiger Selektion. Bei Enterobakterien ist eine Selektion klinisch resistenter Bakteri- en unter giinstigen Bedingungen vermeidbar, weil wahrscheinlich mindestens vier Mutati- onsschritte bei gleichzeitiger Selektion auf- einanderfolgen miissen. Bei Pseudomonaden, Enterokokken und Staphylokokken ist die Resistenzentwicklung nicht vermeidbar.
Nach bisher vorliegenden Daten scheint diese Resistenzquote sich auf ein Niveau einzupen- deln, das fur alle drei Bakterienspezies noch unterhalb von 10 % liegt. Man mui3 aber in Kliniken oder einzelnen Abteilungen mit der klonalen Ausbreitung resistenter Bakterien- stamme rechnen. In diesen Fallen sind hospi- talhygienische Mafinahmen notwendig, um derartige Stamme wieder zu eliminieren.
Literatur
[l] Gellert M. (1981) DNA topoisomerases. Annual Rev. Biochem. 50,879-910.
[2] Hane M. W., Wood, T. H. (1969) Escheri- chia coli K-12 mutants resistant to nalidixic acid: genetic mapping and dominance studies. J. Bacteriol. 99,238-241.
[3] Heisig P., Wiedemann B. (1991) Use of a broad-host-range gyrA plasmid for the cha- racterization of fluoroquinolone-resistant gram-negative bacteria. Antimicrob. Agents Chemother. 35,2031-2036.
[4] Heisig P., Schedletzky H., Falkenstein- Paul H. (1993) Mutations in the gyrA of a highly fluoroquinolone-resistant clinical iso- late of Escherichia coli. Antimicrob. Agents Chemother. 37,696-701.
[5] Heisig P. (1993) Fluoroquinolone resis- tance in a Salmonella typhimurium isolate is due to alterations in both gyrA and gyrB ge- nes. J. Antimicrob. Chemother., im Druck.
[6] Hooper D. C., Wolfson J. S., Ng E. Y., Swartz M. N. (1987) Mechanisms of action of and resistance to ciprofloxacin. Amer. J. Med. 82, suppl. 4A, 12-20.
[7] Kresken M., Wiedemann B. (1988) Deve- lopment of resistance to nalidixic acid and the fluoroquinolones after the introduction of norfloxacin and ofloxacin. Antimicrob. Agents Chemother. 32,1285-1288.
[8] Kresken M., Wiedemann B. (1987) Die Epidemiologie der Resistenz bei Bakterien und ihre Bedeutung fur die Wirksamkeit von Chemotherapeutika. Fortschritte der Anti- mirobiellen, antineoplastischen Chemothera- pie 6,869-1063.
[9] Lesher G. Y., Froehlich E. J., Gruett M. D., BaleyJ. H.,. Brundage R. P (1962) 1,8- Naphthyridine derivates: a new class of che- motherapeutic agents. J. Med. Pharm. 5, 1063-1 065.
[lo] Nikaido H., Thanassi D. G. (1993) Pene- tration of lipophilic agents with multiple pro- tonation sites into bacterial cells: tetracyclines and fluoroquinolones as examples. Anti- microb. Agents Chemother. 37,1393-1399.
[ l l ] Piddock L. J. V., Wise R. (1987) Induc- tion of the SOS-response by 4-quinolone an- timicrobial agents. FEMS Microbiol. Letters 41,288-294.
[12] Piddock L. J. V., Wise R. (1989) Mecha- nisms of resistance to quinolones and clinical perspectives. J. Antimicrob. Chemother. 23,
[13] Shen L. L., Pernet A. G. (1985) Mecha- nism of inhibition of DNA gyrase by ana- logues of nalidixic acid: The target of the drugs is DNA. Proc. Natl. Acad. Sci. USA
475-478.
82.307-311.
[14] Sugino A., Peebles C. L., Kreuzer K. N., Cozzarelli N. R. (1977) Mechanism of action of nalidixic acid: Purification of Escherichia coli nalA gene product and its relationship to DNA gyrase and a novel nicking-closing enzyme. Proc. Natl. Acad. Sci. USA 74, 4767-71.
[15] Swanberg S. L., Wang J. C. (1987) Clo- ning and sequencing of the Escherichia coli gyrA gene coding for the A subunit of DNA gyrase. J. Mol. Biol. 197,729-736.
[16] Ubukata K., Sugiura M., Konno M. (199 1) The norA gene conferring new quino- lone resistance in Staphylococcus epidermi- dis. Chemotherapy (Tokyo) 39,1001-1013.
[17] J. C. Wang (1985) DNA topoisomerases. Annual Rev. Biochem. 54,665-697.
[18] Schedletzky H., Wiedemann B. (1992) Die Wirkung von Norfloxacin, einem neuen Wirkstoff in der Augenheilkunde. Der Augenspiegel 1, ".
[19] Yamagishi J. I., Yoshida H., Yamayoshi H., Nakamura S. (1986) Nalidixic acid-resi- stant mutations of the gyrB gene of Escheri- chia coli. Mol. Gen. Genet. 211, 1-7.
[20] H. Yoshida, M. Bogaki, M. Nakamura, S. Nakamura (1990) Quinolone resistance-de- termining region in the DNA gyrase gyrA gene of Escherichia coli. Antimicrob. Agents Chemother. 34,1271-1272.
Weitere Literatur bei den Autoren auf Anfra- ge erhaltlich.
Prof. Dr. B. Wiedemann (Tg. 1939). Studium der Natunvissenschaft in Karlsruhe, Tubin- gen und Kiel ab SS 1959, 1966 Promotion in Kiel mit Hauptfach Mikrobiologie. 1965- 1966 wiss. Assistent am Hygiene-Institut der Universitat Kiel, 1966 Universitat Frankfurt, 1972 Habilitation fur Medizinische Mikro- biologie bei Prof. Dr. H. Knothe, 1972-1973 Visiting Lecturer, Universitat Bristol, 1974 Professor fur Mikrobiologie Universitat Bonn, Leiter der Abteilung Pharmazeutische Mikrobiologie, Vorsitzender der Paul-Ehr- lich-Gesellschaft von 1987-1992.
Dr. Peter Heisig, Apotheker, geb. 1958 in Ber- lin. Nach Abschlui3 des Pharmaziestudiums an der Freien Universitat Berlin 1981 und Praktischem Jahr in der Curtius-Apotheke, Berlin-Lichterfelde, Approbation als Apothe- ker 1982. Von 1983-1987 Anfertigung einer Dissertation iiber ein molekulargenetisches Grundlagenthema in der Arbeitsgruppe Dr. Kahmann am Max-Planck-Institut fiir Mole- kulargenetik, Berlin. Promotion zum Dr. rer. nat. an der Universitat Bonn 1987. Seit Okto- ber 1987 Wissenschaftlicher Mitarbeiter bei Prof. Dr. Wiedemann, Pharmazeutische Mi- krobiologie der Universitat Bonn. Seit 1992 Habilitand ebendort auf dem Gebiet moleku- larer Mechanismen der bakteriellen Resistenz gegenuber Antibiotika. Fur Arbeiten iiber 4- Chinolonresistenz Preistrager des Wissen- schaftspreises der Paul-Ehrlich-Gesellschaft fur Chemotherapie e.V. des Jahres 1992.
Anschrift:
Prof. Dr. B. Wiedemann, Dr. P. Heisig, Phar- mazeutische Mikrobiologie, Meckenheimer Allee 168,53115 Bonn.
Pharmuzie in unserer Zeit / 22. Jahrg. 1993 / Nx 5-6 295




























