Embed Size (px)
Citation preview

Oecologia (1988) 77 : 343-349 Oecologia �9 Springer-Verlag 1988
Food habits and the basal rate of metabolism in birds
Brian K. McNab Department of Zoology, University of Florida, Gainesville, FL 32611, USA
Summary. The correlation of basal rate of metabolism with various factors is examined in birds. Chief among these is body mass. As in mammals, much of the remaining varia- tion in basal rate among birds is associated with food ha- bits. Birds other than passerines that feed on grass, nectar, flying insects, or vertebrates generally have basal rates that are similar to mammals of the same mass and food habits. In contrast, most invertebrate-eating birds that weigh over 100 g have higher basal rates than equally-sized, inverte- brate-eating mammals. The high basal rates of small passer- ines equal those of small mammals that do not enter torpor and represent the minimal cost of continuous endothermy. Large passerines and small procellariiforms, charadrii- forms, and psittaciforms generally have higher basal rates than mammals with the same mass and food habits. The high basal rates of passerines (in combination with altricial habits) may have significance in permitting high post-natal growth rates and the exploitation of seasonally abundant resources. These interrelations may contribute to the pre- dominance of passerines in temperate land environments.
Key words: Birds - Food habits - Basal rate of metabolism - Post-natal growth rates - Temperate passerines
Basal rate of metabolism in birds has been reviewed repea- tedly (e.g., Brody and Procter 1932; Benedict 1938; La- siewski and Dawson 1967; Zar 1968; Aschoff and Pohl 1970). These studies concluded that: 1) basal rate in birds scales with respect to body mass as it does in mammals, i.e., that total rates are proportional to ca. m~ 2) birds, collectively, have higher basal rates than mammals at the same mass; and 3) among birds, passerines have the highest basal rates. Prinzinger and H/inssler (1980), however, ar- gued that the difference in basal rate between passerines and other birds was exaggerated by the usual difference in body size. When these groups are compared at a common mass the difference in basal rate disappears. Latitude (= cli- mate), body color, and their interaction also have been im- plied or suggested by various authors to influence basal rate in birds (Scholander et al. 1950; Weathers 1979; Ellis 1981; Hails 1983; Ellis 1984; Bennett and Harvey 1987).
This paper examines whether an additional factor may influence basal rate in birds: food habits. Bennett and Har- vey (1987) have examined this possibility with an elaborate statistical analysis. They concluded that the principal source
of variation in avian basal rates, other than body mass, is taxonomic affiliation. Contrary to a similar analysis by Elgar and Harvey (1987), basal rate is associated with food habits in mammals (McNab 1969, 1978, 1983, 1986a, 1987). Given the high basal rates of birds, their rates might also be expected to reflect diet. Consequently, the claim that birds have higher basal rates than mammals should be re- examined when body mass and food habits are considered simultaneously. Any difference in basal rate existing be- tween birds and mammals, when body mass alone is used as a variable, may be related to the differential occurrence of food habits in mammals and birds, or to a differential response by these endotherms to food habits. The question whether a difference in basal rate between mammals and birds can be accounted for by the generally higher body temperature of birds, or whether this difference in body temperature reflects the difference in rate of metabolism and/or insulation between birds and mammals is unclear.
The principal impediment to an analysis of the factors reponsible for setting basal rate in birds is the narrow eco- logical and phylogenetic spectra of birds that have been studied. A disproportionate emphasis has existed on tem- perate members of the Order Passeriformes and the Family Fringillidae. This emphasis is reduced by the use of exten- sive measurements by Soviet biologists, which were summa- rized by Kendeigh et al. (1977), although individual mea- surements were not themselves presented; these data are used here with some concern whether they were measured under standard conditions. At least, the measurements were made at night, and in the few cases where measurements were made on species measured by others, the data are in substantive agreement. Some measurements on tropical birds have been provided by Vleck and Vleck (1981), Barth- olomew etal. (1983), Hails (1983), Ellis 0984), Bucher (1985), and Prinzinger and Jackel (1986).
The influence of food habits on the basal rate of birds
To compensate for the modest data base in birds, the fol- lowing analysis, which is summarized in Table 1 and Figs. I 3, compares the basal rate of birds having a particu- lar food habit with eutherian mammals of the same mass and food habit. At small masses food habits have relatively little influence on eutherian basal rates, but at masses greater than 100 g, basal rate is correlated with food habits. The eating of seeds, grass, or vertebrates is generally asso-

344
Table 1. A comparison of basal rates in small and large birds with eutherian mammals having the same mass and diet"
Diet Eutherian Birds mammals
small large
Passerines Others
small large small large
Herbivore
seeds h h? h h h/1 h fruit h 1 h ? 1 1 nectar h 1? h - h - grass h h - - h leaves h 1 - - 1 h/1 ?
Carnivore
vertebrates h h - - h h/1 invertebrates h 1 h h h h/1 flying insects 1 - h - h/1 1
a The level of basal rate is compared to the Kleiber mammal curve, the values being above the Kleiber curve (h), below (1), both (h/l), unmeasured or uncertain (?), or no species in the category ( ) . Small species weigh less than 100 g
ciated with high basal rates, while feeding on fruit, the leaves of woody plants, or invertebrates is associated with low basal rates (McNab 1986a). When folivory or frugivory is found in sedentary, arboreal mammals , basal rate is especially low, reflecting small muscle masses (McNab 1978). Mixed food habits in mammals are associated with basal rates intermediate to those found in species that spec- ialize on the components of a mixed diet (McNab 1986a). F o o d habits have their greatest impact on basal rate at large body masses because total rate of metabol ism in- creases with mass and, thus, a species with a large mass is more likely to encounter a resource l imitat ion based on the availability, quality, or digestibility of the food used.
Herbivory in birds other than passerines
The correlat ion of basal rate with herbivorous food habits in birds other than passerines is shown in Fig. 1, as well as the general pat tern found in mammals with similar ha-
bits. Herbivory, broadly defined, can be conveniently bro- ken into five catagories: grass-, seed-, fruit-, and nectar- eating, and the consumpt ion of the leaves of woody plants.
Birds that feed selectively on grass (e.g., " r a t i t e s " and Anseriformes), or on seeds (most Gall iformes, Psittaci- formes, and many Columbiformes), usually have the same basal rates as mammals of the same mass and food habits (Fig. 1). Thus, the ostrich (Struthio camelus) has a minimal rate (Crawford and Schmidt-Nielsen 1967) equal to that (Taylor et al. 1969) found in the waterbuck (Kobus defassa); both were measured in the dayt ime and weighed 100 kg. Among small seed-eaters, however, birds have higher basal rates than mammals (Fig. 1).
The general similarity in basal rates between seed-eating birds and mammals extends to an association with climate. The only seed-eating mammals known to have low basal rates live in deserts, especially members of the Heteromyi- dae (Fig. 1). Mas t crops in the desert are ephemeral. Thus, a reduction in basal rate, coupled with hoarding behavior and torpor , reduces the sensitivity of heteromyids to a vari- able food resource. Desert-dwelling, seed-eating birds have basal rates that are nearly identical to those found in equally-sized heteromyids (Fig. 1): they include the Inca dove (Scardafella inca) from Nor th America (MacMil len and Trost 1967 a), p lumed pigeon (Lophophaps ferruginea) from Austra l ia (Dawson and Bennett 1973), and least seed- snipe (Thinocorus rumicivorus) in South America (Ehlers and Mor ton 1982). The California quail (Lophortyx califor- nicus), which is found near desert water holes, also has a somewhat low basal rate (Brush 1965), only slightly greater than is typical of large heteromyids (Fig. 1). Like many equally-sized heteromyids, Scardafella uses to rpor (MacMil len and Trost 1967 a, b).
Few fruit-eating birds have had their rates of metabo- lism measured. Frui t -eat ing as a specialty, often coupled with insect-eating, is pr incipal ly found in the tropics. The basal rates of mousebirds (Colius), African fruit- and leaf- eaters, are low and similar to those of fruit-eating bats (Bartholomew and Trost 1970; Prinzinger e t a l . 1981) (Fig. 1). Trogon rufus, a Neot ropica l t ropical fruit- and in- sect-eater, has a basal rate similar to those of mousebirds and fruit-eating bats ( l ? o 2 / m = l . 2 5 c m 3 / g . h , mass 53 g; personal observations). Trogons are sedentary, which raises the possibili ty that, as in some arboreal mammals , an
.•4.0- ~ 3 . 0 0.5
E 2.0-
o
~o.e %
-~ 0.4
m~ -0.5
0.2
X | GRASS FRUIT | NECTAR | Rt:tites X Coliiformes
+ D X~ ~ X \ / ,Anserif . . . . . =FTrogonif . . . . . X ~ ~ "" /~ SEEDS ~ N U T S NECTAR
X O GoHiforrnes X Trochil[dae /k Columbiformes + Drepanididae [ ] Psittaciformes ~ Meliphagidoe
~ . , (~. Char adii f . . . . . . Nectariniidae �9 �9 ~ LEAVES
SCARDAFELLA ~ �9 Galllformes
DESERT SEEDS - GRASS LOPHOPHAPS /
LOPHORTYX SEEDS 8~ NUTS
. . . . 5 .0
LOgLoM
;g ,'o ;o ,60 ',kg 5 ,b ~o ,oo Body Moss
Fig. 1. Mass-specific basal rate of metabolism as a function of body mass in herbivorous birds. Polygons enclose basal rates in eutherian mammals having specified food habits (McNab 1986a). The Kleiber (1932) and boundary curves (McNab 1983) are indicated by K and B, respectively. Data from Benedict (1938), Lasiewski (1963), Lasiewski and Dawson (1967), Lasiewski et al. (1967), MacMillen and Trost (1967a, b), Bartholomew and Trost (1970), Hainesworth and Wolf (1970), Dawson and Bennett (1973), MacMillen (1974), Hainsworth et al. (1977), Kendeigh et al. (1977), Beuchat et al. (1979), Collins et al. (1980), Prinzinger and H~inssler (1980), Prinzinger et al. (1981), Ehlers and Morton (1982), Bartholomew et al. (1983), Bucher (1985), and Prinzinger and Jackel (1986)
345
INVERTEBRATES I I ~ c ~ C E ~: APTERYGIFORMES VERTEBRATES ' ~ ,~ CICONIIFORMES �9 SPBENISCIFORMES
�9 40 0 CHARADRIrFORMES �9 PELECANIFORMES
0,5 ~1 ~ CUCULIFORMES ~1 PROCELLARIFORMES E] PICIFORMES �9 CICONIIFORMES o FALCONIFORMES I~ FALCONIFORMES
FLYING INSECTS �9 CHARADRIIFORMES DO 2.0 + CAPRIMULGIFORMES I~1 CUCULIFORMES
.~ L0
~ o 8
0.6
2 i~ 04
i 0.2
LO 2.0 3.0 4.0 5.0
Log iO M
i i i ~ I ~ ~ ~0 5~0 i ~og 50 ~00 500 kg 5 ~ ~00 B o d y M a s s
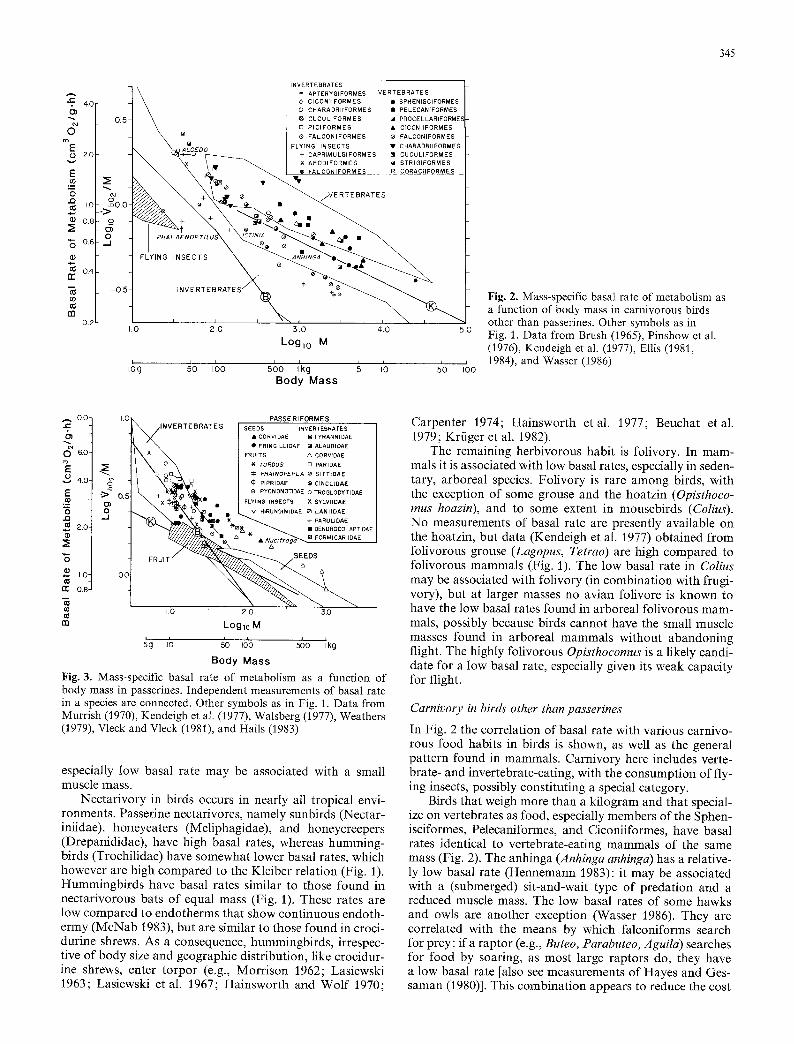
Fig. 2. Mass-specific basal rate of metabolism as a function of body mass in carnivorous birds other than passerines. Other symbols as in Fig. 1. Data from Brush (1965), Pinshow et al. (1976), Kendeigh et al. (1977), Ellis (1981, 1984), and Wasser (1986)
~.~ I0.0~ I ,O l~ PASSERIFORMES .1=: j~ ~ / INVERTEBRATES SEEDS ~ R A T E S
I~ "~" & CORVIDAE ~I TYRANNIBAE
~" ~ N x ~ �9 FRINGILLIDAE [] ALAUDIDAE ( ~ ~C FRUITS ~ CORV~DAE
TURDUS [] PARIDAE ~ . P..,NOPEPLA ~ s, moAE
2 4.0 ~ 1 / ~ . ~ i V * . . . . . . . E "O'RELI~ E "> 0 51 \ + * % ~iL * .YCNONOT, DAE 0 T.OGLOOYT,O.E
.-- " �9 �9 FLYING INSECTS X SYLVIIDAE "0 ~ �9 �9 V HiRUNDINIDAE ~ LANIIDAE r =..I �9 ~ 4- PARULIDAE
2,C 4)( �9 DENDROCOLAPTIDAE / ~ , B FORMICARIIDAE
�9 ~ i.o: o.
r r 0.8-
i.D ' i 3.o
m Log~o M i I i i i i
5g Io 50 Ioo 50o Ikg
Body Mass
Fig. 3. Mass-specific basal rate of metabolism as a function of body mass in passerines. Independent measurements of basal rate in a species are connected. Other symbols as in Fig. 1. Data from Murrish (1970), Kendeigh et al. (1977), Walsberg (1977), Weathers (1979), Vleck and Vleck (1981), and Hails (1983)
especially low basal rate may be associated with a small muscle mass.
Nectarivory in birds occurs in nearly all tropical envi- ronments. Passerine nectarivores, namely sunbirds (Nectar- iniidae), honeyeaters (Meliphagidae), and honeycreepers (Drepanididae), have high basal rates, whereas humming- birds (Trochilidae) have somewhat lower basal rates, which however are high compared to the Kleiber relation (Fig. 1). Hummingbirds have basal rates similar to those found in nectarivorous bats of equal mass (Fig. 1). These rates are low compared to endotherms that show continuous endoth- ermy (McNab 1983), but are similar to those found in croci- durine shrews. As a consequence, hummingbirds, irrespec- tive of body size and geographic distribution, like crocidur- ine shrews, enter torpor (e.g., Morrison 1962; Lasiewski 1963; Lasiewski et al. 1967; Hainsworth and Wolf 1970;
Carpenter 1974; Hainsworth etal. 1977; Beuchat etal . 1979; Krfiger et al. 1982).
The remaining herbivorous habit is folivory. In mam- mals it is associated with low basal rates, especially in seden- tary, arboreal species. Folivory is rare among birds, with the exception of some grouse and the hoatzin (Opisthoco- mus hoazin), and to some extent in mousebirds (Colius). No measurements of basal rate are presently available on the hoatzin, but data (Kendeigh et al. 1977) obtained from folivorous grouse (Lagopus, Tetrao) are high compared to folivorous mammals (Fig. 1). The low basal rate in Colius may be associated with folivory (in combination with frugi- vory), but at larger masses no avian folivore is known to have the low basal rates found in arboreal folivorous mam- mals, possibly because birds cannot have the small muscle masses found in arboreal mammals without abandoning flight. The highly folivorous Opisthocomus is a likely candi- date for a low basal rate, especially given its weak capacity for flight.
Carnivory in birds' other than passerines
In Fig. 2 the correlation of basal rate with various carnivo- rous food habits in birds is shown, as well as the general pattern found in mammals. Carnivory here includes verte- brate- and invertebrate-eating, with the consumption of fly- ing insects, possibly constituting a special category.
Birds that weigh more than a kilogram and that special- ize on vertebrates as food, especially members of the Sphen- isciformes, Pelecaniformes, and Ciconiiformes, have basal rates identical to vertebrate-eating mammals of the same mass (Fig. 2). The anhinga (Anhinga anhinga) has a relative- ly low basal rate (Hennemann 1983): it may be associated with a (submerged) sit-and-wait type of predation and a reduced muscle mass. The low basal rates of some hawks and owls are another exception (Wasser 1986). They are correlated with the means by which falconiforms search for prey: if a raptor (e.g., Buteo, Parabuteo, Aguila) searches for food by soaring, as most large raptors do, they have a low basal rate [also see measurements of Hayes and Ges- saman (1980)]. This combination appears to reduce the cost

346
of flight and maintenance to balance the energy budgets of species that cannot be assured of finding prey. In con- trast, species belonging to the genera Accipiter and Falco have high basal rates in association with the use of powered flight and the directed pursuit of prey. A similar pattern between hunting style and basal rate is found in fish-eating sea birds (Ellis 1984).
At masses less than a kilogram, vertebrate-eating birds generally have high basal rates of metabolism (Fig. 2). In- cluded are small owls (some of which are insectivorous), small hawks (many of which are insectivorous), procellarii- forms, and the kingfisher (Alcedo).
Smaller invertebrate-eating birds, including wood- peckers and cuckoos, like small invertebrate-eating mam- mals that do not enter torpor, have the high basal rates that describe the curve in species that maintain the integrity of endothermy (Fig. 2). At a mass of 40 to 50 g, petrels belonging to the genus Oceanodroma, which feed on inverte- brates and fish, appear to have even higher basal rates (Iver- sen and Krog 1972). At larger masses, invertebrate-eating birds generally have basal rates that are greater than those found in invertebrate-eating mammals; included are the fla- mingo (Phoenicopterus antiquorum), ibis (Guara alba), and woodcock (Scolopax rusticola) (Benedict and Fox 1927; Kendeigh et al. 1977). These rates are similar to those of birds and mammals that feed on vertebrates. In contrast, the kiwis (Apterygiformes), which feed on soil invertebrates, have basal rates (Calder and Dawson 1978) equivalent to those found in mammals that feed on soil invertebrates (Fig. 2).
The remaining carnivorous habit, feeding on flying in- sects, is correlated with low basal rates in Caprimulgi- formes, as it is in bats. In fact, the poorwill (Phalaenoptilus nuttalli), which has a very low basal rate, is the only bird known to hibernate (e.g., Jaeger 1949). The Mississippi kite (Ictinia mississippiensis), which obtains much of its insect diet (e.g., dragonflies and cicadas) in flight, also has a low basal rate (Wasser 1986). At small masses, swifts (Apodi- formes), which also feed on insects in flight, appear to have high basal rates (Fig. 2). Note, however, that only one swift (Apus apus) has had its basal rate measured (Kendeigh et al. 1977). Like many insectivorous bats, swifts are known to enter torpor (Koskimies 1950; Bartholomew et al. 1957).
The influence of food habits in passerines
Passerines that weigh less than 20 g have basal rates equal to those found in mammals of the same mass that do not enter torpor (Fig. 3). That is, in both small mammals and passerines, basal rate is principally set by the cost of contin- uous thermoregulation, even though basal rate is measured in thermoneutrality, i.e., without any increment reflecting the direct cost of temperature regulation. Nevertheless, at small masses some modest differentiation in the level of energy expenditure of passerines occurs in relation to food habits: the lowest basal rates are in New World warblers (Parulidae), Old World warblers (Sylviidae), and tyrant fly- catchers (Tyrannidae), all of which generally feed on active adult or larval insects, or in the tropical bulbuls (Pycnonoti- dae), which feed on fruit and insects. Passerines that feed on nectar (Nectariniidae, Meliphagidae, and Drepanididae) and on flying insects (Hirundinidae) have much higher basal rates than do other birds with these habits (Figs. 1 and 3). The highest basal rates among small passerines are
found in temperate wrens (Troglodytidae), titmice (Pari- dae), and finches (Fringillidae). Wrens selectively feed on adult and larval insects; the fringillids that have been mea- sured, especially crossbills and grosbeaks, eat seeds almost exclusively; and titmice feed both on seeds and larval in- sects.
These modest differences in basal rate may reflect differ- ences in the seasonal availability of food, active insects be- ing unavailable during cold weather, and fruit production often being limited to one season. The principal difference between temperate passerines that eat seasonal foods and those that eat foods that are continuously available, how- ever, is not in the level of the basal rate, but in the tendency of the former to evade the worst winter conditions by long- distance migration.
At masses greater than 20 g, passerines have higher bas- al rates than are found in other mammals of the same mass and food habits. For example, mixed-diet, facultatively-fru- givorous thrushes belonging to the genus Turdus have high basal rates, as do large seed-eating fringillids, the seed- eating nutcracker (Nucifraga), and omnivorous corvids (Fig. 3). Some other birds of the same mass, especially small Procellariiformes, Charadriiformes, and Psittaciformes, have basal rates equal to large passerines (Figs. 2 and 3). The one moderately-sized, invertebrate-eating passerine that has a (comparatively) low basal rate is the dipper (Cin- clus mexicanus) (Murrish 1970).
Discussion
A definitive analysis of the factors influencing basal rate of metabolism in endotherms cannot be accomplished by the examination of one family, one order, or even one class. Those correlations that reflect fundamental ecological rela- tions are, in one example, difficult to separate from correla- tions produced by the coincidences of history, or by the idiosyncratic associations peculiar to a particular group of organisms (see Bennett and Harvey 1987). Thus, the use of flight by most birds and the use of torpor by many small mammals influences the relationship of energetics to food habits and to body mass (McNab 1988).
This analysis of avian energetics, especially relative to the pattern seen in mammals, gives insight into the factors that may influence energy expenditure (see Table 1).
1) Birds often have basal rates that vary in relation to body mass and food habits as they do in mammals. This conclusion is most striking in birds other than passer- ines that feed on grass, seeds, nectar, and to a lesser extent on vertebrates and flying insects.
2) Most large birds known to have basal rates as low as some large invertebrate-eating mammals are selected hawks, eagles, and owls that feed on cryptic and widely- dispersed vertebrates.
3) Invertebrate-eating birds that weight over 100 g, un- like equally-sized invertebrate-eating mammals, generally have basal rates equal to those of vertebrate-eating birds and mammals. Most large invertebrate-eating birds en- counter their food suspended in water, a medium that is easy to filter, whereas most equally-sized mammals that feed on invertebrates extract them from soil. Such mammals ingest large quantities of soil and detritus (McNab 1984), thereby reducing the energy density of the ingested volume.
4) Relatively few birds feed on the leaves of woody plants. Those folivorous grouse that have had their basal

347
E 13
(.3
o CO o
_J
0.6
0.4
a ./ 0.2
/ O0 v , , ,
" o.o LO
Basal Rate of
,o 2000 _// 7 / KL=O.17V o /m
_ ~ , o o - . . . . ~ g / 2 o ,o-2og ~I �9 ~ / / / A / El 40-50g
/ ~ / / * k ~ /x 200 50Og OTHER BIRDS
so-sog @ �9 IOO-200g
--P �9 400-600g �9 750-900g �9 I-2kg
o ' i o ' 4 . 0
Metabolism (cm 30z/g.h)
Fig. 4. Logistic growth constant in birds as a function of mass-specific basal rate of metabolism. Data on logistic growth constants were obtained from Ricklefs (1968, 1973, 1976). The two species with different estimates for the growth constant are indicated by a range and a mean. Species within a given range of body masses and form of development are enclosed. The form of development is either altricial (a) or precocial (p). The dashed curve is derived in the text
rates measured have rates that are much greater than those found in arboreal folivorous mammals, as is to be expected because no grouse is as sedentary as these mammals. In contrast, mousebirds, which have a mixed diet of fruit and leaves, have low basal rates (Fig. 1); the extent to which these low rates are associated with the intake of leaves is unclear.
5) As in mammals, a small muscle mass in association with sedentary habits or flightlessness may be correlated with a low basal rate of metabolism. For example, kiwis, which feed on soil invertebrates, have low basal rates. The fact that the woodcock, which also feeds on soil inverte- brates, has a higher basal rate suggests that the kiwi's low rate is associated with flightlessness. That is, birds do not have basal rates as low as some invertebrate-eating mam- mals because of the nearly universal use by birds of flight with its requirement of a proportionally large muscle mass.
6) At masses less than 20 g, passerines tend to have the same basal rates as are found in equally-sized mammals that do not enter torpor. This similarity is dictated by the minimal cost of continuous endothermy at a small mass (McNab 1983). Consequently, the high basal rate in small passerines is not a property of passerines per se, but reflects the absence of torpor in passerines.
7) The conclusion that birds have higher basal rates of metabolism than mammals is correct principally when body mass is used as the only variable. When other factors in addition to mass are considered, the difference between birds and mammals is reduced. More mammals than birds have low basal rates in association with a reduced muscle mass. The general use of flight by birds permits the evasion of seasonally harsh environments, but prevents a marked reduction of muscle mass. Flightless birds, thus, may gener- ally have " m a m m a l i a n " rates of metabolism. Some of the apparent difference in basal rate between birds and mam- mals may also reflect a differential occurrence of food ha- bits between these classes (e.g., fewer folivores among birds), and to a limited extent a different consequence for birds of having a similar habit (e.g., invertebrativory). Note- worthy among birds having higher basal rates than expected from mass and food habits are intermediate-to-large passer- ines, small procellariiforms, and charadriiforms.
What is the significance of these high basal rates? As with eutherians (McNab 1980, 1986b; Lillegraven et al. 1987), a high basal rate may be translated into a high rate
of reproduction. The question posed here is: Does the suc- cess of passerines in temperate and polar terrestrial environ- ments (Klopfer and MacArthur 1960; Faaborg 1977) reflect a coupling of reproduction to maintenance energetics and the use of food resources that permit high basal rates? Spe- cifically, passerines may predominate in seasonal environ- ments because, coupled to high basal rates, they have high post-natal growth rates. High growth rates permit young to grow and to store sufficient amounts of fat to overwinter or migrate and permit adults to have more than one brood during a short summer.
Is growth rate correlated with basal rate in birds? At first glance, the answer appears to be yes in those (few) species for which both sets of data are available (Fig. 4). Much of this correlation, however, actually reflects the scal- ing of each parameter with body mass. Thus, Ricklefs (1976) described the scaling function for the logistic growth constant in temperate altricial land birds (most o f which were passerines) as K L = l . 1 4 m -~ where m is body mass. The Aschoff-Pohl (1970) quiescent-phase passerine equation (converted to mass-specific oxygen consumption) is i?qjm=6.60m 0.27. Because m-~176 KL = 0.17 Vojm, in which 0.17 = 1.14/6.60. Clearly, much of the increase in KL with an increase in l/o2/m simply reflects a decrease in body mass. Yet, some variation in both basal rate and growth constant occurs at all masses. Within cer- tain groups of birds, namely altricial species that weigh between 30 and 50 g or between 100 and 200 g, and preco- cial species that weigh between 400 and 600 g, the growth constant increases with basal rate independent of body size (Fig. 4). A positive correlation consequently appears to ex- ist between KL and basal rate when birds of a developmental class are grouped by body mass. Yet, high basal rates are associated with low growth rates in some small, altricial procellariiforms (e.g., petrels).
The data available conform to the suggestion that growth rates are positively correlated, at least among pas- serines, with basal rate at a given body mass. Tropical pas- serines tend to have lower post-natal growth constants than temperate passerines (Ricklefs 1968, 1976), which may re- flect an increase of basal rate with latitude. Of special inter- est are the low growth rates of specialist frugivores (see Mor ton 1973). High growth rates facilitate repeated repro- duction at high latitudes where abundant food supplies are available for only short periods each year. Whether the

348
combinat ion of small size, altriciality, food habits, and a high basal rate can completely account for the differential exploitation by passerines of high latitudes is unclear.
Acknowledgements. I thank Lawrence Alexander, Holly Daniels, Laurie Eberhardt, Hugh Ellis, Peter Feinsinger, Alejandro Grajal, Willard Hennemann, Douglas Levey, David Ligon, Philip Long, Carlos Martinez del Rio, Steven Thompson, and Charles Woods for critically examining one or more versions of this manuscript. This manuscript has also greatly profited from following most of the suggestions of an anonymous reviewer.
References
Aschoff J, Pohl H (1970) Rhythmic variations in energy metabo- lism. Fed Proc 29:1541-1552
Bartholomew GA, Trost CH (1970) Temperature regulation in the speckled mousebird, Colius striatus. Condor 72:141-146
Bartholomew GA, Howell TA, Cade TJ (1957) Torpidity in the white-throated swift, Anna hummingbird and poor-will. Con- dor 59 : 145-155
Bartholomew GA, Vleck CM, Bucher TL (1983) Energy metabo- lism and nocturnal hypothermia in two tropical passerine frugi- vores, Manacus vitellinus and Pipra mentalis. Physiol Zool 56:370-379
Benedict FG (1938) Vital energetics: a study in comparative basal metabolism. Carnegie Inst Wash Publ 503:1-215
Benedict FG, Fox EL (1927) The gaseous metabolism of large large wild birds under aviary life. Proc Am Philos Soc 66:511-534
Bennett PM, Harvey PH (1987) Active and resting metabolism in birds : allometry, phylogeny and ecology. J Zoo1213 : 327-363
Beuchat CA, Chaplin SB, Morton ML (1979) Ambient temperature and the daily energetics of two species of hummingbirds, Ca- lypte anna and Selasphorus rufus. Physiol Zool 52:280-295
Brody S, Procter RC (1932) Relation between basal metabolism and mature body weight in different species of mammals and birds. Univ Mo Agr Exp Stn Res Bull 166:89-101
Brush AH (1965) Energetics, temperature regulation and circula- tion in resting, active and defeathered California quail, Lophor- tyx californicus. Comp Biochem Physiol 15:399-421
Bucher TL (1985) Ventilation and oxygen consumption in Ama- zona viridigenalis. J Comp Physiol B 155:269-276
Calder WA III, Dawson TJ (1978) Resting metabolic rates of ratite birds : the kiwis and the emu. Comp Biochem Physiol A 60: 479- 481
Carpenter FL (1974) Torpor in an Andean hummingbird: its eco- logical significance. Science 183:545-547
Collins BG, Cary G, Payne S (1980) Metabolism, thermoregulation and evaporative water loss in two species of Australian nectar- feeding birds (Family Melaphagidae). Comp Biochem Physiol A67 : 629-635
Crawford EC, Schmidt-Nielsen K (1967) Temperature regulation and evaporative cooling in the ostrich. Am J Physiol 212:347-353
Dawson WR, Bennett AF (1973) Roles of metabolic level and temperature regulation in the adjustment of western plumed pigeons (Lophophaps ferruginea) to desert conditions. Comp Biochem Physiol A44:249-266
Ehlers R, Morton ML (1982) Metabolic rate and evaporative water loss in the least seed-snipe, Thinocorus rumicivorus. Comp Bio- chem Physiol A 73 : 233-235
Elgar MA, Harvey PH (1987) Basal metabolic rates in mammals: allometry, phylogeny and ecology. Funct Ecol 1:2~36
Ellis HI (1981) Metabolism and solar radiation in dark and white herons in hot climates. Physiol Zool 53 : 358-372
Ellis HI (1984) Energetics of free-ranging seabirds. In: Whittow GC, Rahn H (eds) Seabird energetics. Plenum Press, New York, pp 203-234
Faaborg J (1977) Metabolic rates, resources, and the occurrence
of non-passerines in terrestrial avian environments. Am Nat 111:903-916
Hails CJ (1983) The metabolic rate of tropical birds. Condor 85 : 61-65
Hainsworth FR, Wolf LL (1970) Regulation of oxygen consump- tion and body temperature during torpor in a hummingbird, Eulampisjugularis. Science 168:368-369
Hainsworth FR, Collins BG, Wolf LL (1977) The function of tor- por in hummingbirds. Physiol Zool 50:215-222
Hayes SR, Gessaman JA (1980) The combined effects of air tem- perature, wind and radiation on the resting metabolism of avian predators. J Therm Biol 5:119-125
Hennemann WW III (1983) Environmental influences on the ener- getics and behavior of anhingas and double-crested cormorants. Physiol Zool 56:201-216
Iversen JA, Krog J (1972) Body temperatures and resting metabolic rates in small petrels. Norw J Zool 20:141-144
Jaeger EC (1949) Further observations on the hibernation of the poor-will. Condor 51 : 105-109
Kendeigh SC, Dol'nik VR, Gavrilov VM (1977) Avian energetics. In: Pinowski J, Kendeigh SC (eds) Granivorous birds in eco- systems, I.B.P. 12. Cambridge University Press, Cambridge, pp 127 204
Kleiber M (1932) Body size and metabolism. Hilgardia 6:315-353 Klopfer PH, MacArthur RH (1960) Niche size and faunal diversity.
Am Nat 94:293-300 Koskimies J (1950) The life of the swift, Micropus apus (L.), in
relation to the weather. Ann Acad Sci Fenn 15:1-151 Kriiger K, Prinzinger R, Schuchmann K-L (1982) Torpor and me-
tabolism in hummingbirds. Comp Biochem Physiol A73:679- 689
Lasiewski RC (1963) Oxygen consumption of torpid, resting, ac- tive, and flying hummingbirds. Physiol Zool 36:122-140
Lasiewski RC, Dawson WR (1967) A re-examination of the rela- tion between standard metabolic rate and body weight in birds. Condor 69:13-23
Lasiewski RC, Weathers WW, Bernstein MH (1967) Physiological responses of the giant hummingbird, Patagona gigas. Comp Biochem Physiol 23 : 797-813
Lillegraven JA, Thompson SD, McNab BK, Patton JL (1987) The origin of eutherian mammals. Biol J Linn Soc 32:281-336
MacMillen RE (1974) Bioenergetics of Hawaiian honeycreepers: the amakihi (Loxops virens) and the anianiau (L. parva). Con- dor 76: 62-69
MacMillen RE, Trost CH (1967a) Thermoregulation and water loss in the Inca dove. Comp Biochem Physiol 20:263-273
MacMillen RE, Trost CH (1967b) Nocturnal hypothermia in the Inca dove, Scardafella inca. J Comp Physiol 23:243-253
McNab BK (1969) The economics of temperature regulation in neotropical bats. Comp Biochem Physiol 31:227-268
McNab BK (1978) Energetics of arboreal folivores: physiological problems and ecological consequences of feeding on an ubiquit- ous food supply. In: Montgomery GG (ed) The ecology of arboreal folivores. Smithsonian Inst. Press, Washington, D.C., pp 153-162
McNab BK (1980) Food habits, energetics, and the population biology of mammals. Am Nat 116:106-124
McNab BK (1983) Energetics, body size, and the limits of endoth- ermy. J Zool 199:1-29
McNab BK (1984) Physiological convergence amongst ant-eating and termite-eating mammals. J Zool 203:485-510
McNab BK (1986 a) The influence of food habits on the energetics of eutherian mammals. Ecol Monogr 56:1-19
McNab BK (1986b) Food habits, energetics, and the reproduction of marsupials. J Zool 208 : 595-614
McNab BK (1987) Basal rate and phylogeny. Funct Ecol 1 : 159-160
McNab BK (1988) Complications inherent in scaling basal rate of metabolism in mammals. Q Rev Biol 63 : 25-54
Morrison PR (1962) Modifications of body temperature by activity in Brazilian hummingbirds. Condor 64:315-323

349
Morton ES (1973) On the evolutionary advantages and disadvan- tages of fruit eating in tropical birds. Am Nat 107 : 8-22
Murrish DE (1970) Responses to temperature in the dipper, Cinclus mexicanus. Comp Biochem Physiol 34:859-869
Pinshow B, Fedak MA, Battles DR, Schmidt-Nielsen K (1976) Energy expenditure for thermoregulation and locomotion in emperor penguins. Am J Physiol 231:903-912
Prinzinger R, H/inssler I (1980) Metabolism-weight relationship in some small nonpasserine birds. Experientia 36:1299-1300
Prinzinger R, Jackel S (1986) Energy metabolism, respiration fre- quency and O2-consumption per breathing act in 11 different sunbird species during day and night. Experientia 42:1002-1003
Prinzinger R, G6ppel R, Lorenz A, Kulzer E (1981) Body tempera- ture and metabolism in the red-backed mousebird (Colius cas- tanotus) during fasting and torpor. Comp Biochem Physiol A69:689 692
Ricklefs RE (1968) Patterns of growth in birds. Ibis 110:419-451 Ricklefs RE (1973) Patterns of growth in birds. II. Growth rate
and mode of development. Ibis 115 : 177-201 Ricklefs RE (1976) Growth rates of birds in the humid New World
tropics. Ibis 118:179 207
Scholander PF, Hock R, Walters V, Irving L (1950) Adaptation to cold in arctic and tropical mammals and birds in relation to body temperature, insulation, and basal metabolic rate. Biol Bull 99 : 259-271
Taylor CR, Spinage CA, Lyman CP (1969) Water relations of the waterbuck, an East African antelope. Am J Physiol 217:630-634
Vleck CM, Vleck D (1981) Metabolic rate in five tropical bird species. Condor 81 : 89-91
Wasser JS (1986) The relationship of energetics of falconiform birds to body mass and climate. Condor 88 : 57-62
Walsberg GE (1977) Ecology and energetics of contrasting social systems in Phainopepla nitens (Aves: Ptilogonatidae). Univ Ca- lif Publ Zool 108:1-63
Weathers WW (1979) Climatic adaptation in avian standard meta- bolic rate. Oecologia (Berlin) 42:81-89
Zar JH (1968) Standard metabolism comparisons between orders of birds. Condor 70:278
Received June 4, 1988
![HEAT STROKE - ksacpr.org.saksacpr.org.sa/GSSHYD-DT5381/UploadData/CourseContent/73f30ce1-e8bc...Sources of body heat ? Normal basal metabolism [BMR] Activity of different organs [](https://img.pdfslide.net/doc/110x75/5cc4974e88c993d8658c49ca/heat-stroke-of-body-heat-normal-basal-metabolism-bmr-activity-of-different.jpg)




























