Embed Size (px)
Citation preview

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1Introduction to Freshwater Algae
1.1 General introduction
Algae are widely present in freshwater environments,such as lakes and rivers, where they are typicallypresent as micro-organisms – visible only with the aidof a light microscope. Although relatively inconspic-uous, they have a major importance in the freshwaterenvironment, both in terms of fundamental ecologyand in relation to human use of natural resources.
This book considers the diversity of algae in fresh-water environments and gives a general overview ofthe major groups of these organisms (Chapter 1),methods of collection and enumeration (Chapter 2)and keys to algal groups and major genera (Chap-ter 4). Algae are considered as indicators of en-vironmental conditions (bioindicators) in terms ofindividual species (Chapter 1) and as communities(Chapter 3).
1.1.1 Algae – an overview
The word ‘algae’ originates from the Latin word forseaweed and is now applied to a broad assemblageof organisms that can be defined both in terms ofmorphology and general physiology. They are simpleorganisms, without differentiation into roots, stemsand leaves – and their sexual organs are not en-closed within protective coverings. In terms of physi-ology, they are fundamentally autotrophic (obtainingall their materials from inorganic sources) and pho-
tosynthetic – generating complex carbon compoundsfrom carbon dioxide and light energy. Some algaehave become secondarily heterotrophic, taking upcomplex organic molecules by organotrophy or het-erotrophy (Tuchman, 1996), but still retaining fun-damental genetic affinities with their photosyntheticrelatives (Pfandl et al., 2009).
The term ‘algae’ (singular alga) is not strictly ataxonomic term but is used as an inclusive label fora number of different phyla that fit the broad de-scription noted above. These organisms include bothprokaryotes (Section 1.3, cells lacking a membrane-bound nucleus) and eukaryotes (cells with a nucleusplus typical membrane-bound organelles).
Humans have long made use of algal species, bothliving and dead. Fossil algal diatomite deposits, forexample, in the form of light but strong rocks, havebeen used as building materials and filtration media inwater purification and swimming pools. Some fossilalgae, such as Botryococcus, can give rise to oil-richdeposits. Certain species of green algae are cultivatedfor the purpose of extracting key biochemicals foruse in medicine and cosmetics. Even blue-green al-gae, often regarded as nuisance organisms, may havebeneficial uses. This is particularly the case for Spir-ulina, which was harvested by the Aztecs of Mexicoand is still used by the people around Lake Chad asa dietary supplement. Spirulina tablets may still beobtained in some health food shops. Blue-green al-gae are, however, are better known in the freshwaterenvironment as nuisance organisms, forming denseblooms having adverse effects on human activities
Freshwater Algae: Identification and Use as Bioindicators Edward G. Bellinger and David C. SigeeC© 2010 John Wiley & Sons, Ltd
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
2 1 INTRODUCTION TO FRESHWATER ALGAE
by producing toxins, clogging water courses and im-pairing recreational activities.
1.1.2 Algae as primary producers
As fixers of carbon and generators of biomass, algaeare one of three major groups of photosynthetic or-ganism within the freshwater environment. They aredistinguished from higher plants (macrophytes) interms of size and taxonomy, and from photosyntheticbacteria in terms of their biochemistry. Unlike algae,photosynthetic bacteria are strict anaerobes and donot evolve oxygen as part of the photosyntheticprocess.
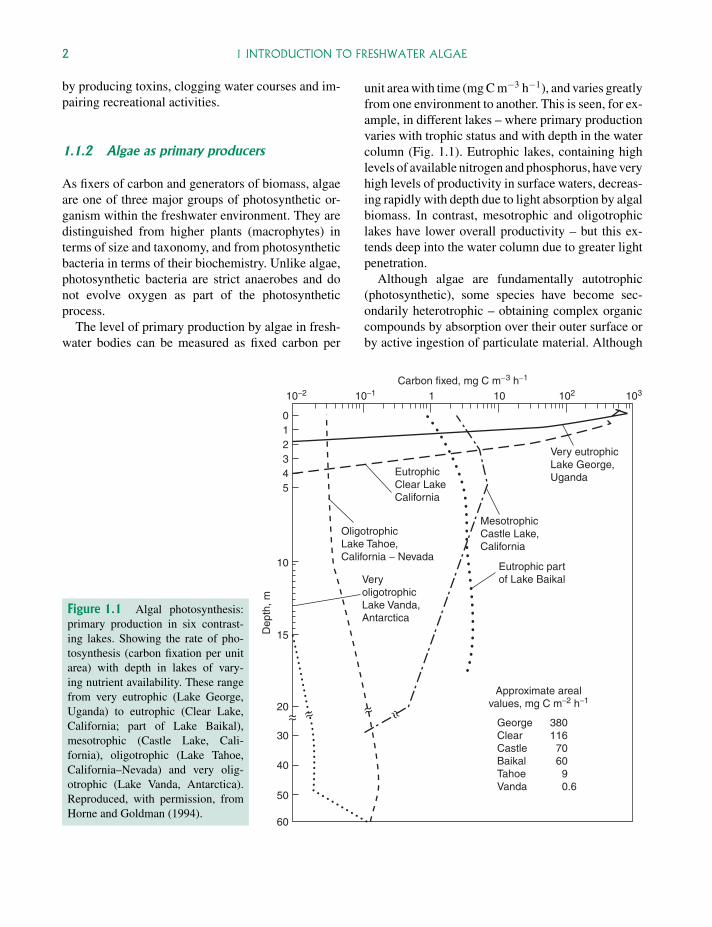
The level of primary production by algae in fresh-water bodies can be measured as fixed carbon per
unit area with time (mg C m−3 h−1), and varies greatlyfrom one environment to another. This is seen, for ex-ample, in different lakes – where primary productionvaries with trophic status and with depth in the watercolumn (Fig. 1.1). Eutrophic lakes, containing highlevels of available nitrogen and phosphorus, have veryhigh levels of productivity in surface waters, decreas-ing rapidly with depth due to light absorption by algalbiomass. In contrast, mesotrophic and oligotrophiclakes have lower overall productivity – but this ex-tends deep into the water column due to greater lightpenetration.
Although algae are fundamentally autotrophic(photosynthetic), some species have become sec-ondarily heterotrophic – obtaining complex organiccompounds by absorption over their outer surface orby active ingestion of particulate material. Although
Approximate arealvalues, mg C m−2 h−1
GeorgeClearCastleBaikalTahoeVanda
380116706090.6
Eutrophic partof Lake Baikal
MesotrophicCastle Lake,California
EutrophicClear LakeCalifornia
VeryoligotrophicLake Vanda,Antarctica
OligotrophicLake Tahoe,California − Nevada
Very eutrophicLake George,Uganda
Dep
th, m
Carbon fixed, mg C m−3 h−1
10−2 10−1 102 1031 10
012345
10
15
20
30
40
50
60
Figure 1.1 Algal photosynthesis:primary production in six contrast-ing lakes. Showing the rate of pho-tosynthesis (carbon fixation per unitarea) with depth in lakes of vary-ing nutrient availability. These rangefrom very eutrophic (Lake George,Uganda) to eutrophic (Clear Lake,California; part of Lake Baikal),mesotrophic (Castle Lake, Cali-fornia), oligotrophic (Lake Tahoe,California–Nevada) and very olig-otrophic (Lake Vanda, Antarctica).Reproduced, with permission, fromHorne and Goldman (1994).

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.1 GENERAL INTRODUCTION 3
such organisms often superficially resemble proto-zoa in terms of their lack of chlorophyll, vigorousmotility and active ingestion of organic material, theymay still be regarded as algae due to their phyloge-netic affinities.
1.1.3 Freshwater environments
Aquatic biology can be divided into two major disci-plines – limnology (water bodies within continentalboundaries) and oceanography (dealing with oceansand seas, occurring between continents). This bookfocuses on aquatic algae present within continen-tal boundaries, where water is typically fresh (non-saline) and where water bodies are of two main types:
� standing (lentic) waters – particularly lakes andwetlands
� running (lotic) waters – including streams andrivers.
The distinction between lentic and lotic systemsis not absolute, since many ‘standing waters’ suchas lakes have a small but continuous flow-through ofwater, and many large rivers have a relatively lowrate of flow at certain times of year. Although thedifference between standing and running waters isnot absolute, it is an important distinction in relationto the algae present, since lentic systems are typicallydominated by planktonic algae and lotic systems bybenthic organisms.
Although this volume deals primarily with al-gae present within ‘conventional freshwater systems’such as lakes and rivers, it also considers algae presentwithin more extreme freshwater environments suchas hot springs, algae present in semi-saline (brackish)and saline conditions (e.g. estuaries and saline lakes)and algae present within snow (where the water isin a frozen state for most of the year).
1.1.4 Planktonic and benthic algae
Within freshwater ecosystems, algae occur eitheras free-floating (planktonic) or substrate-associated
(largely benthic) organisms. Planktonic algae driftfreely within the main body of water (with somespecies able to regulate their position within the watercolumn), while substrate-associated organisms are ei-ther fixed in position (attached) or have limited move-ment in relation to their substrate. These substrate-associated algae are in dynamic equilibrium withplanktonic organisms (see Fig. 2.1), with the balancedepending on two main factors – the depth of waterand the rate of water flow. Build-up of phytoplank-ton populations requires a low rate of flow (otherwisethey flush out of the system) and adequate light levels,so they tend to predominate at the surface of lakes andslow moving rivers. Benthic algae require adequatelight (shallow waters) and can tolerate high rates ofwater flow, so predominate over phytoplankton infast flowing rivers and streams. Benthic algae alsorequire adequate attachment sites – which include in-organic substrate, submerged water plants and emer-gent water plants at the edge of the water body. Thedistinction between planktonic and non-planktonicalgae is ecologically important and is also rele-vant to algal sampling and enumeration procedures(Chapter 2).
Planktonic algae
Planktonic algae dominate the main water body ofstanding waters, occurring as a defined seasonal suc-cession of species in temperate lakes. The tempo-ral sequence depends on lake trophic status (Sec-tion 3.2.3; Table 3.3) with algae forming denseblooms (see Glossary) in eutrophic lakes of diatoms(Fig. 1.16), colonial blue-green algae (Fig. 1.5) andlate populations of dinoflagellates (Fig. 1.10). Duringthe annual cycle, phytoplankton blooms correspondto peaks in algal biovolume and chlorophyll-a con-centration, and troughs in turbidity (see Fig. 2.8).
Benthic algae
Benthic algae occur at the bottom of the water col-umn in lakes and rivers, and are directly associatedwith sediments – including rocks, mud and organic

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
4 1 INTRODUCTION TO FRESHWATER ALGAE
Table 1.1 Size Range of Phytoplankton
CategoryLinear Size (Cell orColony Diameter, µm) Biovolume* (µm3) Unicellular Organisms Colonial Organisms
Picoplankton 0.2–2 4.2 × 10−3–4.2 Photosynthetic bacteriaBlue-green algae
SynechococcusSynechocystis
–
Nanoplankton 2–20 4.2–4.2 × 103 Blue-green algaeCryptophytes
CryptomonasRhodomonas
Microplankton 20–200 4.2 × 103–4.2 × 106 DinoflagellatesCeratiumPeridinium
DiatomsAsterionella
Macroplankton >200 >4.2 × 106 – Blue-green algaeAnabaenaMicrocystis
Biovolume values are based on a sphere (volume = 4/3π r3).Table reproduced from Sigee, 2004.
debris. These attached algae may form majorgrowths on inorganic surfaces or on organic debris,where they are frequently present in mixed biofilms(with bacteria, fungi and invertebrates also present).Under high light conditions, the biofilm may becomedominated by extensive growths of filamentous al-gae – forming a periphyton community (Fig. 2.23).Attached algae may also be fixed to living organismsas epiphytes – including higher plants (Fig. 2.29),larger attached algae (Fig. 2.28) and large plank-tonic colonial algae. Some substrate-associated al-gae are not attached, but are able to move acrosssubstrate surfaces (e.g. pennate diatoms), are looselyretained with gelatinous biofilms or are held withinthe tangled filamentous threads of mature periphytonbiofilms.
Many algal species have both planktonic and ben-thic stages in their life cycle. In some cases theydevelop as actively photosynthetic benthic organ-isms, which subsequently detach and become plank-tonic. In other cases the alga spends most of its ac-tively photosynthetic growth phase in the planktonicenvironment, but overwinters as a dormant metabol-ically inactive phase. Light micrographs of the dis-tinctive overwintering phases of two major bloom-forming algae (Ceratium and Anabaena) are shownin Fig. 2.7.
1.1.5 Size and shape
Size range
The microscopic nature of freshwater algae tendsto give the impression that they all occur within abroadly similar size range. This is not the case witheither free floating or attached algae.
In the planktonic environment (Table 1.1), algaerange from small prokaryotic unicells (diameter<1 µm) to large globular colonies of blue-green algaesuch as Microcystis (diameter reaching 2000 µm) –just visible to the naked eye. This enormous sizerange represents four orders of magnitude on a linearbasis (×12 as volume) and is similar to that seenfor higher plants in terrestrial ecosystems such astropical rainforest.
Planktonic algae are frequently characterized inrelation to discrete size bands – picoplankton(<2 µm), nanoplankton (2–20 µm), microplankton(20–200 µm) and macroplankton (>200 µm). Eachsize band is characterized by particular groups ofalgae (Table 1.1).
In the benthic environment, the size range ofattached algae is even greater – ranging from smallunicells (which colonize freshly exposed surfaces) toextended filamentous algae of the mature periphyton

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.2 TAXONOMIC VARIATION – THE MAJOR GROUPS OF ALGAE 5
community. Filaments of attached algae such asCladophora, for example, can extend several cen-timetres into the surrounding aquatic medium. Thesemacroscopic algae frequently have small colonialalgae and unicells attached as epiphytes (Fig. 2.28),so there is a wide spectrum of sizes within thelocalized microenvironment.
Diversity of shape
The shape of algal cells ranges from simple singlenon-motile spheres to complex multicellular struc-tures (Fig. 1.2). The simplest structure is a unicellu-lar non-motile sphere (Fig. 1.2b), which may becomeelaborated by the acquisition of flagella (Fig. 1.2c),by a change of body shape (Fig. 1.2a) or by the devel-opment of elongate spines and processes (Fig. 1.2d).Cells may come together in groups without definednumber or shape (Fig. 1.2e) or may form globularcolonies that have a defined morphology (Fig. 1.2f,g).Cells may also join together to form linear colonies(filaments) which may be unbranched or branched(Fig. 1.2h, i).
Although motility is normally associated with thepossession of flagella, some algae (e.g. the diatomNavicula and the blue-green Oscillatoria) can movewithout the aid of flagellae by the secretion of sur-face mucilage. In many algae, the presence of sur-face mucilage is also important in increasing overallcell/colony size and influencing shape.
Size and shape, along with other major phenotypiccharacteristics, are clearly important in the classifica-tion and identification of algal species. At a functionaland ecological level, size and shape are also impor-tant in terms of solute and gas exchange, absorptionof light, rates of growth and cell division, sedimen-tation in the water column, cell/colony motility andgrazing by zooplankton and (Sigee, 2004).
1.2 Taxonomic variation – the majorgroups of algae
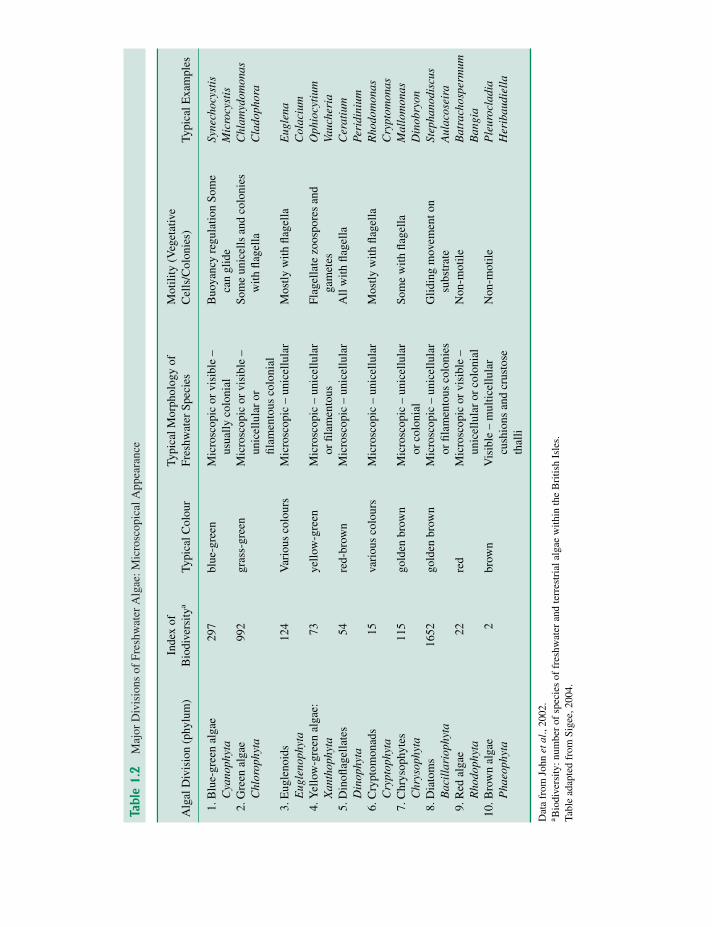
Freshwater algae can be grouped into 10 major divi-sions (phyla) in relation to microscopical appearance(Table 1.2) and biochemical/cytological characteris-tics (Table 1.3). Some indication of the ecological andtaxonomic diversity of these groups is given by the
Figure 1.2 General shapes of algae. Non-motile uni-cells: (a) Selenastrum; (b) Chlorella. Motile unicell: (c)Chlamydomonas. Non-motile colony: (d) Scenedesmus(e) Asterionella. Motile colony: (f) Pandorina; (g)Volvox. Unbranched filament (h) Spirogyra. Branchedfilament (i) Cladophora. Reproduced, with permission,from Bellinger (1992).
number of constituent species (Table 1.2) for fresh-water and terrestrial algae in the British Isles (takenfrom John et al., 2002), with green algae and diatomsfar outnumbering other groups – reflecting theirwidespread occurrence and ability to live in diversehabitats. Diatoms in particular (over 1600 species)are ecologically successful, both as planktonic andbenthic organisms. In addition to the above groups,John et al. (2002) also list other phyla – Raphidophyta(two species), Haptophyta (five species), Eustigmato-phyta (three species), Prasinophyta (13 species) andGlaucophyta (two species). Although these minorphyla have taxonomic and phylogenetic interest, theyhave less impact in the freshwater environment.
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
Tabl
e1
.2M
ajor
Div
isio
nsof
Fres
hwat
erA
lgae
:Mic
rosc
opic
alA
ppea
ranc
e
Alg
alD
ivis
ion
(phy
lum
)In
dex
ofB
iodi
vers
itya
Typi
calC
olou
rTy
pica
lMor
phol
ogy
ofFr
eshw
ater
Spec
ies
Mot
ility
(Veg
etat
ive
Cel
ls/C
olon
ies)
Typi
calE
xam
ples
1.B
lue-
gree
nal
gae
Cya
noph
yta
297
blue
-gre
enM
icro
scop
icor
visi
ble
–us
ually
colo
nial
Buo
yanc
yre
gula
tion
Som
eca
ngl
ide
Syne
choc
ysti
sM
icro
cyst
is2.
Gre
enal
gae
Chl
orop
hyta
992
gras
s-gr
een
Mic
rosc
opic
orvi
sibl
e–
unic
ellu
lar
orfil
amen
tous
colo
nial
Som
eun
icel
lsan
dco
loni
esw
ithfla
gella
Chl
amyd
omon
asC
lado
phor
a
3.E
ugle
noid
sE
ugle
noph
yta
124
Var
ious
colo
urs
Mic
rosc
opic
–un
icel
lula
rM
ostly
with
flage
llaE
ugle
naC
olac
ium
4.Y
ello
w-g
reen
alga
e:X
anth
ophy
ta73
yello
w-g
reen
Mic
rosc
opic
–un
icel
lula
ror
filam
ento
usFl
agel
late
zoos
pore
san
dga
met
esO
phio
cyti
umVa
uche
ria
5.D
inofl
agel
late
sD
inop
hyta
54re
d-br
own
Mic
rosc
opic
–un
icel
lula
rA
llw
ithfla
gella
Cer
atiu
mPe
ridi
nium
6.C
rypt
omon
ads
Cry
ptop
hyta
15va
riou
sco
lour
sM
icro
scop
ic–
unic
ellu
lar
Mos
tlyw
ithfla
gella
Rho
dom
onas
Cry
ptom
onas
7.C
hrys
ophy
tes
Chr
ysop
hyta
115
gold
enbr
own
Mic
rosc
opic
–un
icel
lula
ror
colo
nial
Som
ew
ithfla
gella
Mal
lom
onas
Din
obry
on8.
Dia
tom
sB
acil
lari
ophy
ta16
52go
lden
brow
nM
icro
scop
ic–
unic
ellu
lar
orfil
amen
tous
colo
nies
Glid
ing
mov
emen
ton
subs
trat
eSt
epha
nodi
scus
Aul
acos
eira
9.R
edal
gae
Rho
doph
yta
22re
dM
icro
scop
icor
visi
ble
–un
icel
lula
ror
colo
nial
Non
-mot
ileB
atra
chos
perm
umB
angi
a10
.Bro
wn
alga
eP
haeo
phyt
a2
brow
nV
isib
le–
mul
ticel
lula
rcu
shio
nsan
dcr
usto
seth
alli
Non
-mot
ileP
leur
ocla
dia
Her
ibau
diel
la
Dat
afr
omJo
hnet
al.,
2002
.a B
iodi
vers
ity:n
umbe
rof
spec
ies
offr
eshw
ater
and
terr
estr
iala
lgae
with
inth
eB
ritis
hIs
les.
Tabl
ead
apte
dfr
omSi
gee,
2004
.
6
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
Tabl
e1
.3M
ajor
Div
isio
nsof
Fres
hwat
erA
lgae
:Bio
chem
ical
and
Cyt
olog
ical
Cha
ract
eris
tics
Pigm
enta
tion#
Chl
orop
last
Fine
-Str
uctu
reFl
agel
laA
lgal
Div
isio
nD
iag.
*St
arch
-lik
eO
uter
Thy
lako
id(V
eget
ativ
eC
ells
(phy
lum
)C
hlor
ophy
llsC
arot
enes
Car
oten
oids
Res
erve
Ext
erna
lCov
erin
gM
embr
anes
Gro
ups
&G
amet
es)
1.B
lue-
gree
nal
gae
Cya
noph
yta
aβ
zea-
Cya
no-p
hyce
anst
arch
α
Pept
idog
lyca
nm
atri
ces
orw
alls
00
0
2.G
reen
alga
eC
hlor
ophy
taa,
bα,β,γ
viol
a-T
rue
star
chα
Cel
lulo
sew
alls
,sc
ales
22–
60–
man
y.Si
mila
r(i
soko
nt)
3.E
ugle
noid
sE
ugle
noph
yta
a,b
β,γ
Para
myl
onβ
Prot
ein
pelli
cle
33
1–2
emer
gent
4.Y
ello
w-g
reen
alga
e:X
anth
ophy
taa,
c 1,c
2α,β
Chr
ysol
amin
arin
βPe
ctin
orpe
ctic
acid
wal
l4
32
uneq
ual
(het
erok
ont)
5.D
inofl
agel
late
sD
inop
hyta
a,c 2
βpe
ri-
Tru
est
arch
αC
ellu
lose
thec
a(o
rna
ked)
33
2un
equa
l(h
eter
okon
t)6.
Cry
ptom
onad
sC
rypt
ophy
taa,
c 2α,β
allo
-T
rue
star
chα
Cel
lulo
sepe
ripl
ast
42
2eq
ual(
isok
ont)
7.C
hrys
ophy
tes
Chr
ysop
hyta
a,c 1
,c2,c
3α,β,ε
Chr
ysol
amin
arin
βPe
ctin
,plu
sm
iner
als
and
silic
a
43
2un
equa
l(h
eter
okon
t)
8.D
iato
ms
Bac
illa
rio-
phyt
aa,
c 1,c
2,c
3β,ε
fuco
-C
hrys
olam
inar
inβ
Opa
line
silic
afr
ustu
le4
41,
repr
oduc
tive
cells
only
9.R
edal
gae
Rho
doph
yta
aα,β
Flor
idea
nst
arch
αW
alls
with
gala
ctos
epo
lym
erm
atri
x
20
0
10.B
row
nal
gae
Pha
eoph
yta
a,c 1
,c2,c
3β,ε
Lam
inar
inβ
Wal
lsw
ithal
gina
tem
atri
x4
32
uneq
ual
(het
erok
ont)
repr
oduc
tive
cells
only
Dat
afr
omL
ee(1
997)
,van
den
Hoe
ket
al.(
1995
),Jo
hnet
al.(
2002
)an
dW
ehr
and
Shea
th(2
003)
.#M
ajor
pigm
ents
are
show
nin
bold
type
.*D
iagn
ostic
caro
teno
ids,
used
forH
PLC
anal
ysis
(Fig
.2.1
1):z
ea-(
Zea
xant
hin:
also
pres
enti
nch
loro
phyt
es,c
rypt
ophy
tes)
,vio
la-(
viol
axan
thin
),pe
ri-(
peri
dini
n),a
llo-(
allo
xant
hin)
,fu
co-
(fuc
oxan
thin
,als
opr
esen
tin
chry
soph
ytes
).St
arch
-lik
ere
serv
esα
:α-1
,4gl
ucan
;β:β
-1,3
gluc
an.
Tabl
ead
apte
dfr
omSi
gee,
2004
.
7
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
8 1 INTRODUCTION TO FRESHWATER ALGAE
In terms of diversity, freshwater algae also havea major division into prokaryotes (blue-green algae)and eukaryotes (remaining groups) based on cell size,ultrastructure, antibiotic resistance and general phys-iology. Even within the eukaryote groups, fundamen-tal differences in phenotype and molecular character-istics indicate evolutionary derivation from a rangeof ancestral types (polyphyletic origins).
1.2.1 Microscopical appearance
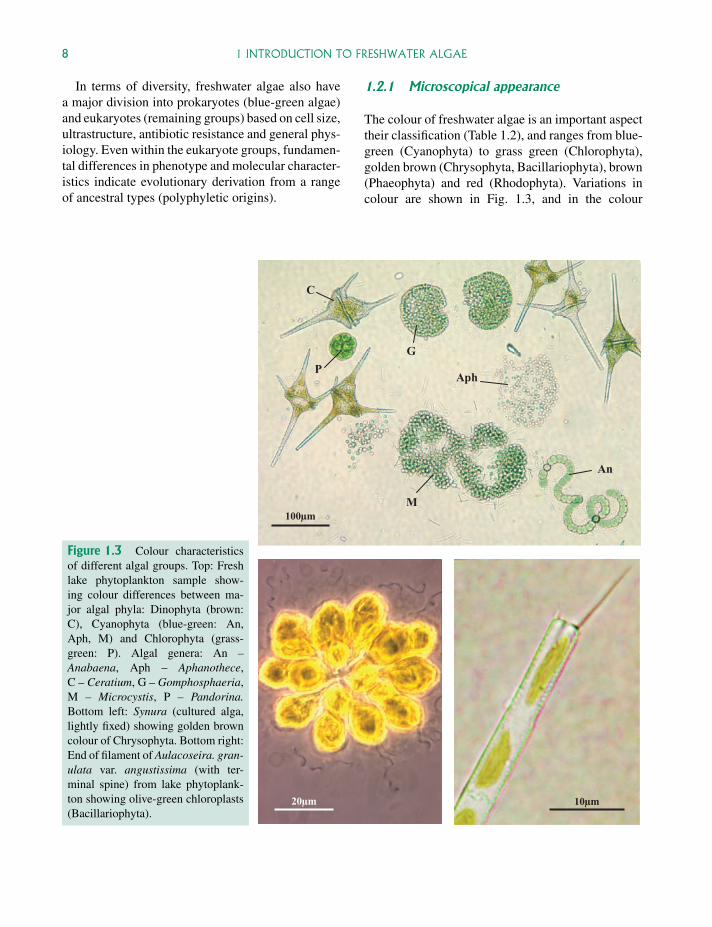
The colour of freshwater algae is an important aspecttheir classification (Table 1.2), and ranges from blue-green (Cyanophyta) to grass green (Chlorophyta),golden brown (Chrysophyta, Bacillariophyta), brown(Phaeophyta) and red (Rhodophyta). Variations incolour are shown in Fig. 1.3, and in the colour
100µm
20µm 10µm
C
M
An
Aph P
G
Figure 1.3 Colour characteristicsof different algal groups. Top: Freshlake phytoplankton sample show-ing colour differences between ma-jor algal phyla: Dinophyta (brown:C), Cyanophyta (blue-green: An,Aph, M) and Chlorophyta (grass-green: P). Algal genera: An –Anabaena, Aph – Aphanothece,C – Ceratium, G – Gomphosphaeria,M – Microcystis, P – Pandorina.Bottom left: Synura (cultured alga,lightly fixed) showing golden browncolour of Chrysophyta. Bottom right:End of filament of Aulacoseira. gran-ulata var. angustissima (with ter-minal spine) from lake phytoplank-ton showing olive-green chloroplasts(Bacillariophyta).

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.2 TAXONOMIC VARIATION – THE MAJOR GROUPS OF ALGAE 9
photographs of Chapter 4. The use of colour as a tax-onomic marker can be deceptive, however, since thenormal balance of pigments may vary. Green algaeliving on snow, for example, may have a preponder-ance of carotenoid pigments – forming a ‘red bloom’(Hoham and Duval, 2001). Heterotrophic algae takeup complex organic molecules by surface absorption(organotrophy) or ingestion (phagotrophy), and haveeither retained photosynthetic pigments (photo-organotrophs, mixotrophs) or lost pigmentationcompletely (obligate heterotrophs) (Sigee, 2004).Even within a ‘normal’ ecological situation, thecolour of a particular alga can show considerablevariation (see, for example, Anabaena, Fig. 4.24).
Apart from colour, the other obvious character-istics under the light microscope are overall size,whether the organism is unicellular or colonial andwhether it is motile (actively moving) or non-motile.Within different groups, algae may be largely uni-cellular (euglenoids, dinoflagellates, cryptophytes),multicellular (brown algae) or a mixture of thetwo (other groups). Motility (single cells or entirecolonies) is also an important feature, with some al-gal groups being entirely flagellate (dinoflagellates,cryptophytes) while others are a mixture of flagellateand non-flagellate organisms (green algae, xantho-phytes). Other groups of algae are entirely withoutflagella, but are able to move by buoyancy regula-tion (blue-greens), gliding movements on substratum(blue-greens, diatoms) or are entirely non-motile (redand brown algae).
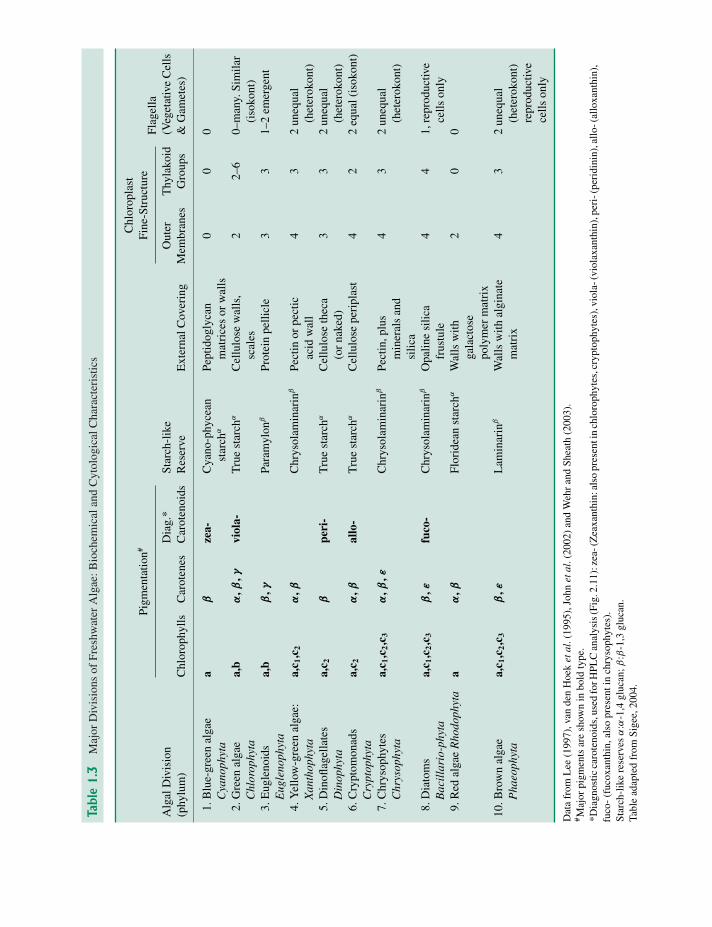
1.2.2 Biochemistry and cell structure
Major biochemical features of freshwater algae in-clude pigmentation, food reserves and external cov-ering (Table 1.3). Different groups have distinc-tive combinations of chlorophylls and carotenes,while only three groups (blue-greens, cryptomon-ads, red algae) have phycobilins. All pigmented al-gae have chlorophyll-a, which can therefore be usedfor the estimation of total biomass (Chapter 2).Diagnostic carotenoids have been particularly use-ful in high-performance liquid chromatography(HPLC) identification and quantitation of major algalgroups within mixed phytoplankton samples (Sec-
tion 2.3.3, Fig 2.11), and have been particularlyuseful in the analysis of estuarine eutrophication(Section 3.5.2).
Visualization of key differences in cell structurenormally requires the higher resolution of oil im-mersion (light microscopy), transmission or scanningelectron microscopy (TEM or SEM) and includesboth internal (e.g. chloroplast fine structure) and ex-ternal (e.g. location/number of flagella, cell surfaceornamentation) features. Comparisons of light andelectron microscope images are shown in Fig. 1.4(light/TEM) and Figs. 4.56 and 4.57 (light/SEM).
1.2.3 Molecular characterizationand identification
Although identification of algal taxa is normallybased on microscopic characteristics (particularlymorphology and colour), there are a number of sit-uations in which molecular techniques have takenprecedence, such as the following.
� No clear morphological characteristics are avail-able. This has particularly been the case for unicel-lular blue-green algae.
� Algae are relatively inaccessible and difficult tovisualize. This is the case for biofilms, where algaeare enclosed in a gelatinous matrix, and in manycases are a relatively small component of a veryheterogeneous community of organisms.
� Diversity is being studied within species, wherestrains are often distinguished in biochemical andgenetic terms.
Ultimately, species definition and identification inboth prokaryote and eukaryote algae may depend onmolecular analysis, with determination of unique anddefining DNA sequences followed by developmentof species-specific nucleotide probes from these (seebelow) .This approach would be particularly relevantin the case of blue-green algae, but there are a numberof problems in relation to species-specificity in thisgroup (Castenholz, 1992):
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
10 1 INTRODUCTION TO FRESHWATER ALGAE
Ch
Ca
Ch
P
Th
Gy
V
1µm3µm
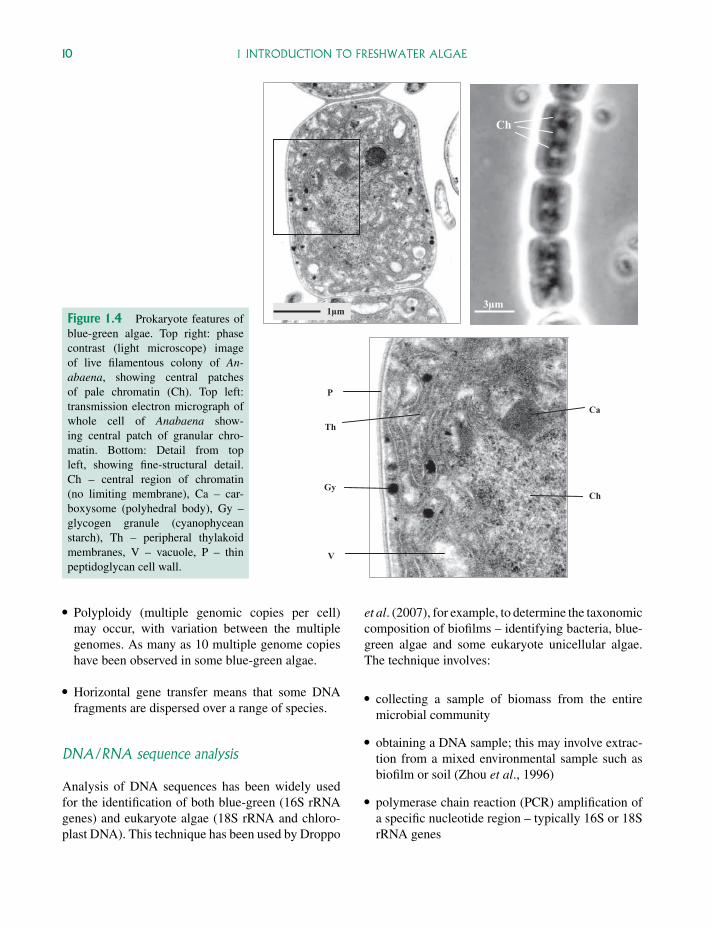
Figure 1.4 Prokaryote features ofblue-green algae. Top right: phasecontrast (light microscope) imageof live filamentous colony of An-abaena, showing central patchesof pale chromatin (Ch). Top left:transmission electron micrograph ofwhole cell of Anabaena show-ing central patch of granular chro-matin. Bottom: Detail from topleft, showing fine-structural detail.Ch – central region of chromatin(no limiting membrane), Ca – car-boxysome (polyhedral body), Gy –glycogen granule (cyanophyceanstarch), Th – peripheral thylakoidmembranes, V – vacuole, P – thinpeptidoglycan cell wall.
� Polyploidy (multiple genomic copies per cell)may occur, with variation between the multiplegenomes. As many as 10 multiple genome copieshave been observed in some blue-green algae.
� Horizontal gene transfer means that some DNAfragments are dispersed over a range of species.
DNA/RNA sequence analysis
Analysis of DNA sequences has been widely usedfor the identification of both blue-green (16S rRNAgenes) and eukaryote algae (18S rRNA and chloro-plast DNA). This technique has been used by Droppo
et al. (2007), for example, to determine the taxonomiccomposition of biofilms – identifying bacteria, blue-green algae and some eukaryote unicellular algae.The technique involves:
� collecting a sample of biomass from the entiremicrobial community
� obtaining a DNA sample; this may involve extrac-tion from a mixed environmental sample such asbiofilm or soil (Zhou et al., 1996)
� polymerase chain reaction (PCR) amplification ofa specific nucleotide region – typically 16S or 18SrRNA genes

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.2 TAXONOMIC VARIATION – THE MAJOR GROUPS OF ALGAE 11
� separation of the amplified strands by denatur-ing gradient gel electrophoresis (DGGE), or pu-rification of the PCR products using a rapid puri-fication kit
� sequence analysis with comparison to a standarddatabase; sequence identification is normally basedon a match of at least 90%.
The pattern of bands in the DGGE gel gives anestimate of community complexity, and the intensityof individual bands (derived from individual species)a measure of population size. In addition to providingtaxonomic information where classical morphologi-cal characteristics do not apply, molecular identifica-tion also has the advantage that the whole of themicrobial community (communal DNA sample)is being analysed in an objective way. Non-photosynthetic bacteria, Archaea, and protozoaare also identified in addition to prokaryote and
eukaryote algae (Droppo et al., 2007; Galandet al., 2008).
Potential limitations of molecular analysis are thatidentification may be tentative (often only to genuslevel) and taxonomic quantitation (relative numbersof different algae) is difficult. The technique has beenparticularly useful in relation to the biodiversity ofmarine blue-green unicellular algae, but has also beenused in a number of freshwater systems (Table 1.4)and at the freshwater/marine interface (nif H genes).
nifH genes Foster et al. (2007) used DNA quanti-tative PCR (QPCR) technology to amplify and detectthe presence of species-specific nif H genes in blue-green algae. This gene encodes the iron-containingprotein nitrogenase (the key enzyme involved innitrogen fixation), and provides a marker for ni-trogen fixing (diazotrophic) algae in the freshwaterenvironment. The technique was used to demon-strate that the blue-green algal symbiont Richelia
Table 1.4 Molecular Identification of Algal Species in Aquatic Environmental Samples
Environmental Sample Technique Reference
Picoplankton in Lake Baikal Direct sequencing Semenova andKuznedelov (1998)
Colonial blue-greens: Anabaena,Microcystis, Nodularia
PCR-RFLP analysis of the cpcB-A intergenicspacer & flanking regions
Bolch et al. (1996)
Flagellate nanoplankton SSU rRNA probes for Paraphysomonas(Chrysophyte)
Caron et al. (1999)
Mixed diatom populations andlaboratory cultures
Large subunit rRNA probe for Pseudo-Nitzschia(Diatom)
Scholin et al. (1997)
Estuarine river samples Use of ITS-specific PCR assays for Pfiesteria(Dinoflagellate)
Litaker et al. (2003)
Laboratory biofilm samples:blue-greens and unicell eukaryotes
Amplification of 16sRNA genes, with denaturinggel electrophoresis (DGGE)
Droppo et al. (2007)
Diazotrophic blue-green algae withinthe Amazon river freshwater plume
Quantitative PCR (QPCR) analysis of the nif Hgene (encodes part of the nitrogenize enzyme)
Foster et al. (2007)
Arctic ecosystems: Freshwaterstamukhi lake and inflow river
Sequencing of 18s rRNA genes to identify majorcryptophyte river populations, plus minor lakepopulations of diatoms
Galand et al. (2008)
Lake and river flagellate samples Population heterogeneity in Spumella(Chrysophyte) – SSU rRNA sequences
Pfandl et al. (2009)
Pond samples Population heterogeneity in Desmodesmus(Chlorophyte) – ITS2rRNA sequences
Vanormelingen et al.(2009)
SSU rRNA – small subunit ribosomal RNA.ITS – internal transcribed spacer.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
12 1 INTRODUCTION TO FRESHWATER ALGAE
(associated with the diatom Hemiaulus hauckii) wasspecifically linked to the Amazon freshwater outflow(river plume) in the western tropical North Atlantic(WTNA) ocean, and that the H. hauckii–Richeliacomplex could be used as a bio-indicator for pocketsof freshwater within the WTNA ocean.
Molecular probes The development of species-specific oligonucleotide probes from DNA sequencedata, followed by in situ hybridization, has consider-able potential for the identification and counting ofalgae in environmental samples.
As with direct sequencing, this technique has par-ticular advantages with small unicellular algae, wherethere are often relatively few morphological featuresavailable for identification. Caron et al. (1999) se-quenced the small-subunit ribosomal genes of fourspecies of the colourless chrysophyte genus Para-physomonas, leading to the development of oligonu-cleotide probes for P. imperforata and P. bandaiensis.
Molecular probes have major potential for the de-tection of nuisance algae, particularly those that pro-duce toxins. They have been used, for example, to dis-tinguish toxic from non-toxic diatom species (Scholinet al., 1997), where differentiation would otherwiserequire the time-consuming application of scanningand transmission electron microscopy. They have alsobeen used for the rapid identification of Pfiesteria pis-cicida, a potentially toxic dinoflagellate that has beenthe cause of extensive fish mortalities in coastal riversof the eastern United States. Litaker et al. (2003) usedunique sequences in the internal transcribed spacer(ITS) regions ITS1 and ITS2 to develop PCR assayscapable of detecting Pfiesteria in natural river assem-blages. These have been successfully used to detectthe potentially harmful organism in the St Johns Riversystem, Florida (USA).
1.3 Blue-green algae
Blue-green algae (Cyanophyta) are widely-occurringthroughout freshwater environments, ranging in sizefrom unicellar forms such as Synechococcus (barelyvisible under the light microscope – Figs. 2.16 and4.31) to large colonial algae such as Microcystis (Fig.4.34) and Anabaena (Fig. 4.24a). Large colonies ofthe latter can be readily seen with the naked eye, and
show a simple globular or filamentous form withcopious mucilage. The balance of photosyntheticpigments present in blue-green algae (Table 1.3)varies with light spectrum and intensity, resulting ina range of colours from brown to blue-green (Fig.4.24a). The phycobilin pigments are particularlyprominent, with phycocyanin tending to predominateover phycoerythrin at low light levels, giving the cellsthe blue-green colour typical of these algae. It is thusan advantage to observe material from shaded as wellas better illuminated situations wherever possible.
1.3.1 Cytology
Prokaryote status
The prokaryote nature of these algae is indicated bythe small size of the cells (typically <10µm diame-ter) and by the presence of central regions of nucleoidDNA (not enclosed by a nuclear membrane). Thesenucleoid regions (Fig. 1.4) can be observed both bylight microscopy (as pale central areas within livingcells) and by transmission electron microscopy (asgranular regions in chemically fixed cells) – wherethe absence of a limiting membrane can be clearlyseen. Although these algae lack the cytological com-plexity of eukaryote organisms (no membrane-boundorganelles such as mitochondria, plastids, micro-somes, Golgi bodies) they do have a range of sim-ple granular inclusions such as carboxysomes, cyno-phycin granules, polyphosphate bodies and glycogenparticles (Fig. 1.4). Photosynthetic pigments are asso-ciated with thylakoid membranes, which are typicallydispersed throughout the peripheral protoplasm.
As with other prokaryotes, cell walls are made upof a peptidoglycan with a lipopolysaccharide layeroutside and are generally quite thin. Some speciesalso have a mucilage layer outside the cell wall whichmay be dense or watery, structured or unstructured.The outside layers of the cell wall can sometimesbecome stained straw coloured or brownish from ironand other compounds in the surrounding water as inScytonema and Gloeocapsa.
The fundamental bacterial nature of these organ-isms distinguishes them from all other algae, anddetermines a whole range of features – including

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.3 BLUE-GREEN ALGAE 13
molecular biology, physiology, cell size, cell struc-ture and general morphology.
Gas vacuoles
In some species gas vacuoles may be formed, appear-ing under the light microscope as highly refractive orquite dark structures. Gas vacuoles aid buoyancy inplanktonic species allowing the cells to control theirposition in the water column. Cells may then con-gregate at a depth of optimal illumination, nutrientconcentration or other factor for that species. This isnot necessarily at the surface as there the light inten-sity may be too great and cause photoinhibition orpermanent cell damage. Movement up and down inthe water column can enhance nutrient uptake as it al-lows the cells to migrate to depths where essential nu-trients, e.g. phosphates, are more abundant and thenup towards the surface for light energy absorption.When gas vacuole containing species are present in asample of water they often tend to float to the surface(this can cause problems when preparing the samplefor cell counts; see Chapter 2).
In addition to gas vacuole-mediated movementsof planktonic blue-greens within the water column,other types of motility also occur – including glidingmovements of filamentous algae (such as Oscillato-ria) on solid substrata.
1.3.2 Morphological and taxonomic diversity
Blue-green algae are remarkable within the prokary-ote kingdom for showing the range of size and formnoted above, with some organisms forming quitelarge, complex, three-dimensional colonies. Lim-ited differentiation (Figs. 4.24a–c) can occur withincolonies, with the formation of heterocysts (nitrogen-fixation) and akinetes (thick-walled resistant cells). Infilamentous forms the cells may be the same widthalong the filament or in some genera they may narrowor taper towards the end, even forming a distinct hair-like structure in the case of Gloeotrichia (Fig. 4.22).Clear branching occurs in the most complex forms,with a fundamental distinction between ‘true branch-ing’ as in Stigonema (Fig. 4.20), or ‘false branching’as in Tolypothrix (Fig. 4.21). True branching involvesdivision of a single cell within the filament, giving
rise to two or more daughter cells which themselvesform branches. In contrast, false branching involveslateral extension of the filament without the singlecell division and daughter cell development as above.
Freshwater blue-green algae can be divided intofour main groups (Table 1.5) in relation to generalmorphology, presence/absence of specialized cellsand the nature of branching in filamentous forms.These four groups form the basis for current taxon-omy of this phylum, as adopted by John et al. (2002),Komarek et al. (2003a,b).� Chroococcales. The simplest blue-green algae, oc-
curring essentially as solitary cells (no filamentousforms), typically enclosed by a thin layer of mu-cilage. The cells may remain as single cells, orbe aggregated into plate-like or globular colonies.Typically planktonic with some colonial forms (e.g.species of Microcystis) forming massive surfaceblooms containing individual colonies that are rec-ognizable with the naked eye. They typically lackspecialized cells, though one group (typified byChaemaesiphon) forms exospores.
� Oscillatoriales. Filamentous algae, lacking hetero-cysts and akinetes. These relatively simple algaeoccur as planktonic (some bloom forming) or ben-thic aggregations. In some cases they form densemats on mud or rocky substrata which secondar-ily detach as metaphyton into to the main body ofwater (Fig. 2.1).
� Nostocales. A diverse group of filamentous al-gae, planktonic or benthic, with heterocysts andakinetes but not showing true branching. Filamentsmay be unbranched or show false branching. Thesealgae are able form large colonies by lateral asso-ciation of filaments (bundles), 3-D tangles or asradiating filaments from the centre of the sphere.
� Stigonematales. Filamentous algae, with hetero-cysts and akinetes and showing true branching.Structurally the most complex blue-green algae,with some thalli (e.g. Fischerella) differentiatedinto multiseriate/uniseriate basal filaments anduniseriate erect branches. Largely benthic algae,with genera such as Stigonema commonly attachedto substrata in standing and flowing waters, detach-ing to form planktonic masses.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
14 1 INTRODUCTION TO FRESHWATER ALGAE
Table 1.5 Taxonomic and Morphological Diversity in Freshwater Blue-Green Algae
Taxonomic OrderMajor Morphotype Colony Size and Form Attached or Planktonic Example*
Chroococcales(a) Unicellular to spheroid
colonies, lackingspecialized vegetative,resistant or reproductivecells
Single cells Planktonic or attached to plantand substrate surfaces
Synechococcus (4.31)
Small colonies(4–32 cells)
Free-floating, tangled-up withfilamentous algae orattached to substrates
Chroococcus (4.33)Gleocapsa (4.32)
Flat plate of cells Free-floating or sedentary MerismopediaLarge solid spherical colony Planktonic Aphanocapsa (4.35)
Microcystis (4.34)Large hollow spherical colony Planktonic Gomphosphaeria (4.30)
Coelosphaerium(b) Unicellular, forming
exosporesCells remain single or form
multi-layered coloniesAttached to surfaces of
aquatic plants, algae andinorganic substrate
Chamaesiphon
OscillatorialesFilamentous algae, lacking
heterocysts and akinetesElongate straight filaments Planktonic or benthic
Benthic matOscillatoria (4.27)†
Phormidium (4.29)Elongate spiral filaments Planktonic or on mud surfaces Spirulina (4.26)
NostocalesFilamentous algae forming
heterocysts and akinetes,but no true branching
Bundles of Elongate filaments Planktonic or benthic Aphanizomenon (4.23)Nostoc (4.25)
3-D tangle of filaments Planktonic Anabaena (4.24)Spherical colony of radiating
filamentsPlanktonic Gloeotrichia (4.22)
StigonematalesFilamentous algae forming
heterocysts and akinetes,with true branching
Branched mass of filaments Benthic or planktonic Stigonema (4.20)Differentiation into basal
filaments and erectbranches
Benthic StauromatonemaFischerella
*Figure numbers in brackets; † Planktonic species of Oscillatoria are placed in the genera Planktothrix, Pseudanabaena or Limnothrixby some authors.
The range of size and form noted in Table 1.5indicates a wide morphological diversity, which isuseful in taxonomic identification. The relative im-portance of molecular versus morphological charac-teristics in relation to taxonomy reflects the debateas to whether these organisms should be treated asbacteria (cyanobacteria – Stanier et al., 1978) or al-gae (blue-green algae: Lewin, 1976). Although theyfundamentally resemble bacteria in their prokaryotefeatures, they also differ from bacteria in carrying outphotosynthesis coupled to O2 evolution, and in thecomplexity of their morphology. Although currenttaxonomy is based primarily on phenotypic charac-
ters (see above), ultrastructural and molecular dataprovide useful supplementary information (Komareket al., 2003a). Current molecular analyses supportthe separation of non-heterocystous and heterocys-tous genera (Rudi et al., 2000), and are particularlyrelevant in the case of unicellular blue-greens, wheremorphological features are not adequate. The classi-cal (eukaryote) use of morphology to define species isalso problematic in this group, since the species con-cept and definition of species is limited by a completeabsence of sexual reproduction.
In the absence of sexual processes, reproductionin this group is by vegetative or specialized asexual

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.3 BLUE-GREEN ALGAE 15
means. Asexual spores (akinetes) consist of vegeta-tive cells that are larger than normal. They gener-ally have thickened walls and, in filamentous forms,are often produced next to heterocysts (Fig. 4.24c).Baeocysts (small spherical cells formed by divisionof a mother cell may) be produced in some coc-coid species and are released into the environment.In some filamentous forms deliberate fragmentationof the filament can occur – releasing the fragments(hormogonia) to produce new filaments.
1.3.3 Ecology
Blue green algae are thought to have arisen approx-imately 3.5 billion years ago (Schopf, 1993) duringwhich time they have been the dominant form of lifefor about 1.5 billion years. As a result of this long evo-lutionary history, they have adapted to (and frequentlydominate) all types of freshwater environment –including extreme conditions (thermal springs, des-iccating conditions), brackish (semisaline) condi-tions, high and low nutrient environments and plank-tonic/benthic habitats. At high latitudes, blue-greenalgae are particularly adapted to low temperatures(Tang & Vincent, 2002) – often dominating the ben-thic environment by forming dense mats. In milderclimates, these organisms frequently dominate sur-face waters, where they are able to out-competeother phytoplankton under eutrophic conditions(Section 3.2.3).
Algal blooms
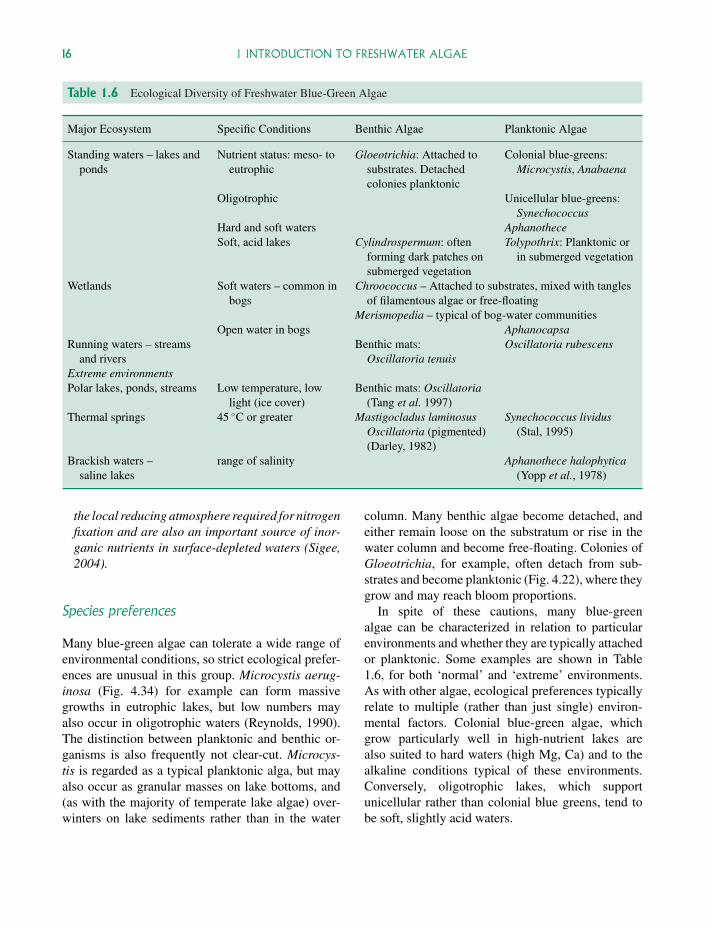
In mid to late summer, eutrophic temperate lakesfrequently develop massive populations of colonialblue-green algae These may rise to the surface of thelake, forming a thick layer of algal biomass at thetop of the water column, out-competing other algaeand having major impacts on zooplankton and fishpopulations (Sigee, 2004). The ability of blue-greensto out-compete other freshwater algae has been at-tributed (Shapiro 1990) to a range of characteristics,including:
� optimum growth at high temperatures – summertemperatures
50µm
Figure 1.5 Midsummer blue-green algal bloom in aeutrophic lake. SEM view of lake epilimnion phyto-plankton sample (August), showing the dense popu-lation of filamentous Anabaena flos-aquae that totallydominated the algal bloom (chlorophyll-a concentration140 µg ml−1). Copious mucilage associated with this algais seen as numerous fine strands, formed during the dehy-dration preparation process. See also Figs 2.8 (seasonalcycle) and 4.24 (live Anabaena). Photo: Sigee and Dean.
� low light tolerance – important within the densealgal bloom, and enabling algae to survive lowerin the water column
� tolerance of low N/P ratios – allowing continuedgrowth when N becomes limiting
� depth regulation by buoyancy – avoiding photoin-hibition during the early phase of population in-crease, and allowing algae to obtain inorganic nu-trients from the hypolimnion when the epilimnionbecomes depleted in mid to late summer
� resistance to zooplankton grazing – limiting theimpact of herbivory on algal growth
� tolerance of high pH/low CO2 concentrations – al-lowing continued growth of blue-greens (but notother algae) at the lake surface during intensebloom formation
� symbiotic association with aerobic bacteria – bac-terial symbionts at the heterocyst surface maintain
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
16 1 INTRODUCTION TO FRESHWATER ALGAE
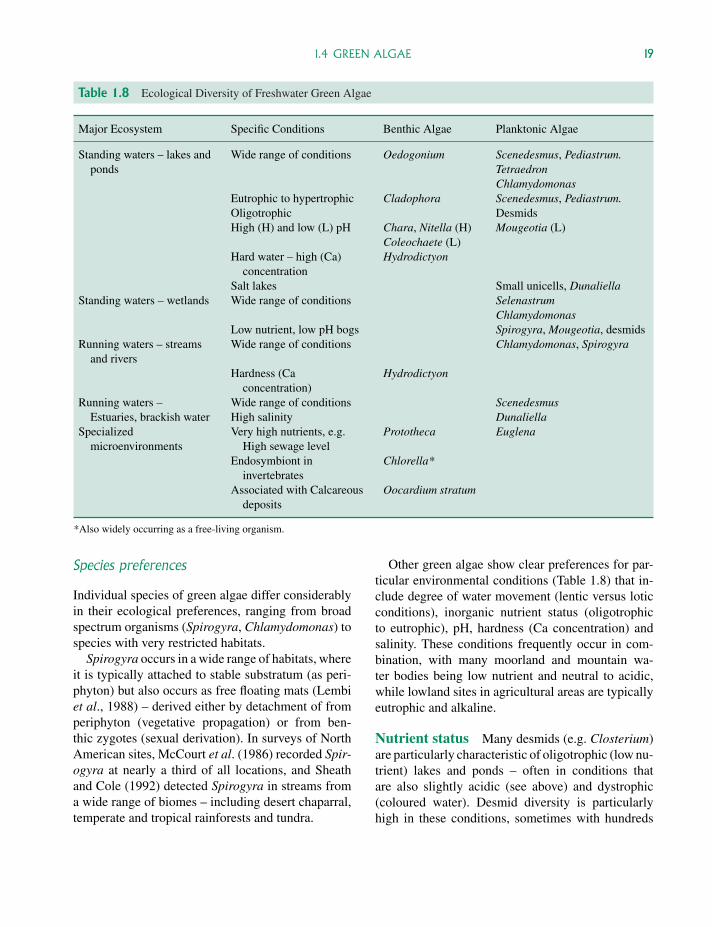
Table 1.6 Ecological Diversity of Freshwater Blue-Green Algae
Major Ecosystem Specific Conditions Benthic Algae Planktonic Algae
Standing waters – lakes andponds
Nutrient status: meso- toeutrophic
Gloeotrichia: Attached tosubstrates. Detachedcolonies planktonic
Colonial blue-greens:Microcystis, Anabaena
Oligotrophic Unicellular blue-greens:Synechococcus
Hard and soft waters AphanotheceSoft, acid lakes Cylindrospermum: often
forming dark patches onsubmerged vegetation
Tolypothrix: Planktonic orin submerged vegetation
Wetlands Soft waters – common inbogs
Chroococcus – Attached to substrates, mixed with tanglesof filamentous algae or free-floating
Merismopedia – typical of bog-water communitiesOpen water in bogs Aphanocapsa
Running waters – streamsand rivers
Benthic mats:Oscillatoria tenuis
Oscillatoria rubescens
Extreme environmentsPolar lakes, ponds, streams Low temperature, low
light (ice cover)Benthic mats: Oscillatoria
(Tang et al. 1997)Thermal springs 45 ◦C or greater Mastigocladus laminosus
Oscillatoria (pigmented)(Darley, 1982)
Synechococcus lividus(Stal, 1995)
Brackish waters –saline lakes
range of salinity Aphanothece halophytica(Yopp et al., 1978)
the local reducing atmosphere required for nitrogenfixation and are also an important source of inor-ganic nutrients in surface-depleted waters (Sigee,2004).
Species preferences
Many blue-green algae can tolerate a wide range ofenvironmental conditions, so strict ecological prefer-ences are unusual in this group. Microcystis aerug-inosa (Fig. 4.34) for example can form massivegrowths in eutrophic lakes, but low numbers mayalso occur in oligotrophic waters (Reynolds, 1990).The distinction between planktonic and benthic or-ganisms is also frequently not clear-cut. Microcys-tis is regarded as a typical planktonic alga, but mayalso occur as granular masses on lake bottoms, and(as with the majority of temperate lake algae) over-winters on lake sediments rather than in the water
column. Many benthic algae become detached, andeither remain loose on the substratum or rise in thewater column and become free-floating. Colonies ofGloeotrichia, for example, often detach from sub-strates and become planktonic (Fig. 4.22), where theygrow and may reach bloom proportions.
In spite of these cautions, many blue-greenalgae can be characterized in relation to particularenvironments and whether they are typically attachedor planktonic. Some examples are shown in Table1.6, for both ‘normal’ and ‘extreme’ environments.As with other algae, ecological preferences typicallyrelate to multiple (rather than just single) environ-mental factors. Colonial blue-green algae, whichgrow particularly well in high-nutrient lakes arealso suited to hard waters (high Mg, Ca) and to thealkaline conditions typical of these environments.Conversely, oligotrophic lakes, which supportunicellular rather than colonial blue greens, tend tobe soft, slightly acid waters.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.4 GREEN ALGAE 17
1.3.4 Blue-green algae as bio-indicators
As with other algal groups, the presence or absenceof particular species can be a useful indicator ofecological status (Table 1.6). The dominant presenceof colonial blue-greens, forming dense summerblooms (Fig. 1.5) has been particularly useful asan indicator of high nutrient status, and these algaeare a key component of various trophic indices(Chapter 3). Conversely, populations of unicellularblue-green algae are indicative of oligotrophic tomesotrophic conditions.
1.4 Green algae
In the freshwater environment, green algae (Chloro-phyta) range in size from microscopic unicellular or-ganisms to large globular colonies and extensive fil-amentous growths. They are characterized by a freshgreen coloration due to the presence of chlorophylls-aand -b, which are not obscured by accessory pigmentssuch as β-carotene and other carotenoids. In excep-tional cases the carotenoid pigments may occur inlarge amounts, obscuring the chlorophylls and givingthe alga a bright red colour. This is seen with the flag-ellate Haematococcus (Fig. 4.54), which frequentlycolours bird baths and other small pools bright red,and the snow alga Chlamydomonas nivalis – whereone of the red carotenoids has a major photoprotectivefunction.
1.4.1 Cytology
In addition to their characteristic pigmentation andother biochemical features (Table 1.3), green algaealso have a number of distinctive cytological aspects.
� Flagella, when present, occur in pairs. These areequal in length without tripartite tubular hairs, andhave a similar structure (isokont). Some generahave four or even eight flagella, but this is unusual.
� Chloroplasts vary in shape, size and number. Inunicellular species they tend to be cup-shaped(Fig. 4.55), but in filamentous forms may be an-
nular (Fig. 4.19), reticulate (Fig. 4.17), discoid orribbon-like (Fig. 4.13). They have a double outermembrane, with no enclosing periplastidal endo-plasmic reticulum.
� Production and storage of the photosynthetic re-serve (starch) occurs inside the plastid, with gran-ules frequently clustered around the pyrenoid. In allother eukaryote algae the storage material occursmainly in the cytoplasm.
In motile species an eyespot is frequently present,appearing red or orange (Fig. 4.38) in fresh spec-imens. The cell wall in green algae is made ofcellulose.
1.4.2 Morphological diversity
Green algae are the most diverse group of algae, withabout 17 000 known species (Graham and Wilcox,2000). This diversity is reflected by the varied of mor-phology, with organisms being grouped into a rangeof growth forms – depending on whether they areunicellular, colonial or filamentous (Table 1.7).The level of greatest morphological and reproductivecomplexity is represented by Charalean algae (e.g.Chara, Nitella), which can reach lengths of over a me-tre, have whorls of branches at nodes along the lengthof the thallus and have been frequently confused withaquatic higher plants such as Ceratophyllum.
In the past, this morphological diversity providedthe taxonomic basis for green algal classification(Bold and Wyne, 1985) – with orders primarily beingdefined largely on a structural basis. These includedthe orders Volvocales (flagellate unicells and simplecolonial forms), Chlorococcales (unicells and non-motile coenobial colonies), Ulotrichales (unbranchedfilaments) and Chaetophorales (branched filaments).More recently, the application of cytological, com-parative biochemical and molecular sequencing tech-niques has demonstrated the occurrence of extensiveparallel evolution. On the basis that classificationshould reflect phylogeny, these original groupingsare thus no longer valid, and a new classification isemerging where individual orders contain a mixture

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
18 1 INTRODUCTION TO FRESHWATER ALGAE
Table 1.7 Range of Morphologies in Freshwater Green Algae
Major Morphotype Attached or Planktonic Example*
Flagellate unicells Planktonic Chlamydomonas (4.55)Nonflagellate Unicells Planktonic or benthic (often
associated with periphyton)Crescent shaped – Selenastrum (4.78)Equatorial division – Micrasterias (4.81);
Closterium (4.80)Colonial Planktonic Net – Hydrodictyon (4.1)
Hollow sphere – Volvox (4.40), Coelastrum,Eudorina (4.39)
Solid sphere – Pandorina (4.36)Plate of cells – Gonium (4.38), Pediastrum (4.47)Small linear colonies – Scenedesmus (4.50)Branching colonies – Dictyosphaerium (4.45)
Disk-shaped plate, onecell thick
Attached to higher plants(epiphyte) or to stones
Coleochaete, Chaetosphaeridium
Unbranched filaments Typically attached, but maydetach to become planktonic
Uronema, Microspora, Oedogonium (4.17),Zygnema (4.15), Spirogyra (4.13)
Branched filaments Chaetophora, Draparnaldia, Cladophora (4.6)Large complex algae with
whorls of branchesAttached Chara, Nitella
*Figure numbers in brackets.
unicells, globular colonial and filamentous forms(Graham and Wilcox, 2000).
1.4.3 Ecology
Green algae are ecologically important as major pro-ducers of biomass in freshwater systems, either asplanktonic (standing waters) or attached (running wa-ters) organisms – where they respectively may formdense blooms and periphyton growths.
Algal blooms
In mesotrophic and eutrophic lakes, green algae donot normally produce the dense blooms seen withdiatoms and blue-green algae – but do become dom-inant or co-dominant in early summer, between theclearwater phase and midsummer mixed algal bloom.In smaller ponds, filamentous Spirogyra and colo-nial Hydrodictyon frequently form surface blooms orscums, while in garden pools and birdbaths Haemato-coccus may form dense populations. When nutrientlevels become very high, a switch may occur from
colonial blue-green to green algae as major bloomformers. This is can be seen in some managed fish-ponds, where organic and inorganic nutrients are ap-plied to enhance fish production by increasing car-bon flow through the whole food chain. In the Trebonbasin biosphere reserve (Czech Republic), applica-tion of lime (supply of carbonate and bicarbonateions), organic fertilizers and manure leads to high pHand hypertrophic nutrient levels. In these conditions,a short diatom bloom (Stephanodiscus) is replaced inearly summer by populations of rapidly growing uni-cellular and small colonial green algae (Scenedesmusand Pediastrum) which continue to dominate forthe rest of the annual cycle (Pokorny et al., 2002a;Pechar et al., 2002). Extensive growth of attachedalgae (periphyton) may also occur under high nutri-ent conditions. Branched thalli of Cladophora canform extended communities of shallow water vegeta-tion along the shores of eutrophic lakes and streams,breaking off in storms to form mass of loose biomasswhich can degrade to generate noxious odours.Another filamentous alga, Mougeotia, can form largemucilaginous subsurface growths in lakes that havebeen affected by acid rain.
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.4 GREEN ALGAE 19
Table 1.8 Ecological Diversity of Freshwater Green Algae
Major Ecosystem Specific Conditions Benthic Algae Planktonic Algae
Standing waters – lakes andponds
Wide range of conditions Oedogonium Scenedesmus, Pediastrum.TetraedronChlamydomonas
Eutrophic to hypertrophic Cladophora Scenedesmus, Pediastrum.Oligotrophic DesmidsHigh (H) and low (L) pH Chara, Nitella (H)
Coleochaete (L)Mougeotia (L)
Hard water – high (Ca)concentration
Hydrodictyon
Salt lakes Small unicells, DunaliellaStanding waters – wetlands Wide range of conditions Selenastrum
ChlamydomonasLow nutrient, low pH bogs Spirogyra, Mougeotia, desmids
Running waters – streamsand rivers
Wide range of conditions Chlamydomonas, Spirogyra
Hardness (Caconcentration)
Hydrodictyon
Running waters –Estuaries, brackish water
Wide range of conditions ScenedesmusHigh salinity Dunaliella
Specializedmicroenvironments
Very high nutrients, e.g.High sewage level
Prototheca Euglena
Endosymbiont ininvertebrates
Chlorella*
Associated with Calcareousdeposits
Oocardium stratum
*Also widely occurring as a free-living organism.
Species preferences
Individual species of green algae differ considerablyin their ecological preferences, ranging from broadspectrum organisms (Spirogyra, Chlamydomonas) tospecies with very restricted habitats.
Spirogyra occurs in a wide range of habitats, whereit is typically attached to stable substratum (as peri-phyton) but also occurs as free floating mats (Lembiet al., 1988) – derived either by detachment of fromperiphyton (vegetative propagation) or from ben-thic zygotes (sexual derivation). In surveys of NorthAmerican sites, McCourt et al. (1986) recorded Spir-ogyra at nearly a third of all locations, and Sheathand Cole (1992) detected Spirogyra in streams froma wide range of biomes – including desert chaparral,temperate and tropical rainforests and tundra.
Other green algae show clear preferences for par-ticular environmental conditions (Table 1.8) that in-clude degree of water movement (lentic versus loticconditions), inorganic nutrient status (oligotrophicto eutrophic), pH, hardness (Ca concentration) andsalinity. These conditions frequently occur in com-bination, with many moorland and mountain wa-ter bodies being low nutrient and neutral to acidic,while lowland sites in agricultural areas are typicallyeutrophic and alkaline.
Nutrient status Many desmids (e.g. Closterium)are particularly characteristic of oligotrophic (low nu-trient) lakes and ponds – often in conditions thatare also slightly acidic (see above) and dystrophic(coloured water). Desmid diversity is particularlyhigh in these conditions, sometimes with hundreds

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
20 1 INTRODUCTION TO FRESHWATER ALGAE
of species occurring together at the same site(Woelkerling, 1976). Desmids are also typical ofnutrient-poor streams, where they are permanent res-idents of periphyton – making up to 10% of totalcommunity biomass. These desmids are associatedwith plants such as the bryophyte Fontinalis, achiev-ing concentrations as high as 106 cells g−1 of sub-strate (Burkholder and Sheath, 1984). Desmids oftenconstitute a significant proportion of algal biomass inwetlands (bogs and fens) where they are also a majoraspect of species diversity.
Some species of desmids – such as Closteriumaciculare, are more typical of high nutrient, slightlyalkaline lakes – and are indicators of eutrophic con-ditions. Periphytic algae typical of eutrophic watersinclude Cladophora.
Acid lakes Although desmids are typical of neu-tral to slightly acid lakes, other green algae areadapted to more extreme conditions. These includethe filamentous green alga, Mougeotia, which canform substantial sub-surface growths in acidified wa-ters – and is widely regarded as an indicator of earlyenvironmental change (Turner et al., 1991). Whetheracidification is the result of acid precipitation, exper-imentation (Webster et al., 1992) or industrial pol-lution, the acid conditions also tend to be associatedwith:
� relatively clear waters, due to low levels of phyto-plankton
� increases in the concentration of metals such as Aland Zn
� reduced levels of dissolved organic carbon,with derivation largely from external lake (al-lochthonous) rather than internal (autochthonous)sources
� food web changes, including a reduction in thenumbers of herbivores.
Laboratory studies (Graham et al., 1996a,b)showed that Mougeotia was physiologically adaptedto such conditions, with the ability to photosynthe-sis over a wide range of irradiances (300–2300 µmolquanta m−2 s−1) and tolerance to a broad range of
both pH (pH 3–9) conditions and metal concentra-tions. The physiological adaptations of this alga, cou-pled with reduced grazing from herbivores, probablyaccounts for the extensive growth and domination ofthe benthic environment in many acidic waters.
Alkaline and calcium-rich waters Someplanktonic (Closterium aciculare) and benthic algae(e.g. Nitella and Chara) are adapted to grow in alka-line habitats, many of which are also rich in calcium(hard waters). Nitella produces lush meadows on thebottoms of neutral to high pH lakes, and Chara canform dense, lime encrusted lawns in shallow alkalinewaters. Hydrodictyon is also typical of hard-waterconditions, occasionally blanketing the surface ofponds and small lakes, and occurring widely in largeralkaline standing waters and hardwater streams. Ex-tensive growths of this alga can also be observed inagricultural situations such as irrigation ditches, ricepaddy fields and fish farms – where there is somedegree of eutrophication.
Some calcium-loving algae occur in very restrictedenvironments. The unusual, slow-growing desmidOocardium stratum, for example, occurs in calcare-ous streams and waterfalls at the top of branched cal-careous tubes, in association with deposits of ‘traver-tine’ and ‘tufa’.
Saline waters Various green algae are adapted tosaline conditions in salt lakes and estuaries, wheresalt levels range from low (brackish waters) to highlevels. With some widely occurring genera such asScenedesmus, brackish conditions are at one end ofthe continuum of environmental tolerance. Other al-gae, such as Dunaliella, are specifically adapted toa more extreme situation. This organism occurs inthe highly saline waters of commercial salt pans(salterns) and natural salt lakes such as the Great SaltLake of Utah (USA). Dunaliella is able to tolerate ex-ternal salt concentrations of >5 M, by balancing thehigh external osmotic pressure (OP) with a high in-ternal OP generated by photosynthetically producedglycerol.
Specialized environments In addition to thefree-living occurrence of particular algae in lakes andrivers that have a range of characteristics (see above),other green algae are adapted to more specialized

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.5 EUGLENOIDS 21
conditions. Some of these involve a change in themode of nutrition from autotrophic to heterotrophic,and include various species of the non-flagellateunicell Chlorella – a widespread endosymbiont offreshwater invertebrates. Other green algae retaintheir free-living existence, but are able to supplementphotosynthesis (mixotrophy) by absorption ofexogenous dissolved carbon (Tuchman, 1996) suchas amino acids and sugars through their cell surface(osmotrophy). The ability to use external organiccarbon may be important in two situations – whenphotosynthesis is limiting, and when external solubleorganic material is in excess.
� Limiting photosynthesis. This may occur, for ex-ample, under conditions of chronic low light inten-sity (e.g. arctic lakes). It is also typical of manyacid lakes, where low pH reduces the level of dis-solved inorganic carbon to levels that are unableto saturate photosynthesis. Laboratory experimentson Coleochaete, an acidophilic alga have demon-strated an ability to use dissolved organic carbonin the form of hexose sugars and sucrose (Grahamet al., 1994)
� High levels of external dissolved organic carbon.Some osmotrophic green algae are photo-heterotrophic, only able to use organic carbon whenlight is present, and when their photosynthesisis inhibited by the high availability of dissolvedorganic carbon (Lewitus and Kana, 1994). Thecolourless unicellular alga Prototheca has com-pletely lost the ability to photosynthesize and ob-tains all its carbon from external sources presentin soils and freshwater environments that are con-taminated by sewage. This evolutionary relation-ship of this obligate heterotroph to green algaeis indicated by the presence of starch-containingplastids.
1.4.4 Green algae as bioindicators
Contemporary algae
Habitat preferences of contemporary green algae(Table 1.8) are frequently useful in providing in-formation on physicochemical characteristics of the
aquatic environment. Filamentous green algae, forexample, often dominate environments stressed bycultural eutrophication, acidification and metal con-tamination (Cattaneo et al., 1995).
Fossil algae
Green algae do not typically produce resistantwalls that persist in aquatic sediments, and aremuch less useful than diatoms and chrysophytes asbioindicators in terms of the fossil record. There aresome exceptions to this, where the cell wall containsfatty acid polymers known as algaenans, that canwithstand millions of years of burial (Gelin et al.,1997). Scenedesmus and Pediastrum are among themost common members of green algae that havefossil records, containing algaenans and also somesilicon. The desmid Staurastrum is also frequentlyseen as fossil remains, due to the impregnation ofcell wall material with polyphenolic compoundswhich confer resistance to bacterial decay (Gunnisonand Alexander, 1975).
1.5 Euglenoids
Euglenoid algae (Euglenophyta) are almost entirelyunicellular organisms, with a total of 40 genera world-wide (about 900 species) – most of which are fresh-water. Cells are typically motile, either via flagella or(in non-flagellate cells) by the ability of the body tochange shape (referred to as ‘metaboly’).
About one-third of the Euglenoids are photosyn-thetic and classed amongst the algae. The rest arecolourless, being either heterotrophic or phago-trophic, and are usually placed in the Protozoa. Inphotosynthetic organisms, pigmentation is closelysimilar to the green algae (Table 1.3), but the variablepresence of carotenoid pigments means that theseorganisms can routinely vary in colour from freshgreen (Fig. 4.51) to yellow-brown. In some situa-tions, the accumulation of the carotenoid astaxan-thin gives cells a bright red coloration. This is seenparticularly well in organisms such as Euglena san-guinea, which forms localized blooms in ponds andditches.
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
22 1 INTRODUCTION TO FRESHWATER ALGAE
1.5.1 Cytology
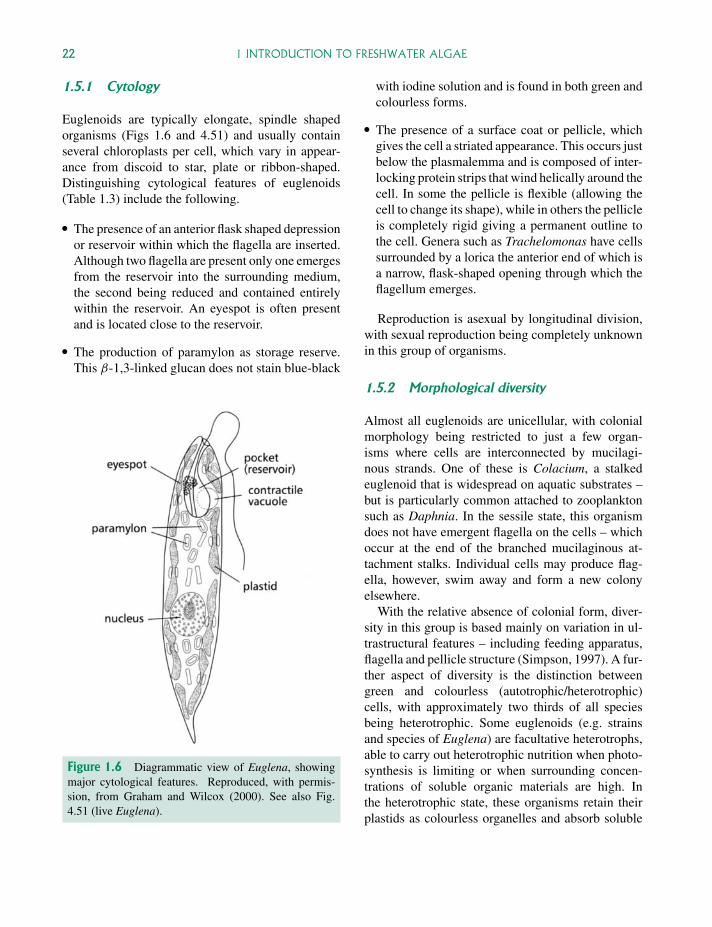
Euglenoids are typically elongate, spindle shapedorganisms (Figs 1.6 and 4.51) and usually containseveral chloroplasts per cell, which vary in appear-ance from discoid to star, plate or ribbon-shaped.Distinguishing cytological features of euglenoids(Table 1.3) include the following.
� The presence of an anterior flask shaped depressionor reservoir within which the flagella are inserted.Although two flagella are present only one emergesfrom the reservoir into the surrounding medium,the second being reduced and contained entirelywithin the reservoir. An eyespot is often presentand is located close to the reservoir.
� The production of paramylon as storage reserve.This β-1,3-linked glucan does not stain blue-black
Figure 1.6 Diagrammatic view of Euglena, showingmajor cytological features. Reproduced, with permis-sion, from Graham and Wilcox (2000). See also Fig.4.51 (live Euglena).
with iodine solution and is found in both green andcolourless forms.
� The presence of a surface coat or pellicle, whichgives the cell a striated appearance. This occurs justbelow the plasmalemma and is composed of inter-locking protein strips that wind helically around thecell. In some the pellicle is flexible (allowing thecell to change its shape), while in others the pellicleis completely rigid giving a permanent outline tothe cell. Genera such as Trachelomonas have cellssurrounded by a lorica the anterior end of which isa narrow, flask-shaped opening through which theflagellum emerges.
Reproduction is asexual by longitudinal division,with sexual reproduction being completely unknownin this group of organisms.
1.5.2 Morphological diversity
Almost all euglenoids are unicellular, with colonialmorphology being restricted to just a few organ-isms where cells are interconnected by mucilagi-nous strands. One of these is Colacium, a stalkedeuglenoid that is widespread on aquatic substrates –but is particularly common attached to zooplanktonsuch as Daphnia. In the sessile state, this organismdoes not have emergent flagella on the cells – whichoccur at the end of the branched mucilaginous at-tachment stalks. Individual cells may produce flag-ella, however, swim away and form a new colonyelsewhere.
With the relative absence of colonial form, diver-sity in this group is based mainly on variation in ul-trastructural features – including feeding apparatus,flagella and pellicle structure (Simpson, 1997). A fur-ther aspect of diversity is the distinction betweengreen and colourless (autotrophic/heterotrophic)cells, with approximately two thirds of all speciesbeing heterotrophic. Some euglenoids (e.g. strainsand species of Euglena) are facultative heterotrophs,able to carry out heterotrophic nutrition when photo-synthesis is limiting or when surrounding concen-trations of soluble organic materials are high. Inthe heterotrophic state, these organisms retain theirplastids as colourless organelles and absorb soluble

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.6 YELLOW-GREEN ALGAE 23
nutrients over their whole surface (osmotrophy).Other euglenoids (Petalomonas, Astasia, Peranema)are obligate heterotrophs and have lost their plastidscompletely. Many of these organisms consume partic-ulate organic material (phagocytic) and have evolveda complex feeding apparatus.
1.5.3 Ecology
Euglenoids are generally found in environmentswhere there is an abundance of decaying organicmaterial. This is in line with the heterotrophic na-ture of many of these organisms and the ability totake up complex organic material either in the sol-uble or particulate state. Typical habitats includeshallow lakes, farm ponds, wetlands, brackish sandand mudflats. Within these environments, euglenoidsare particularly associated with interfaces such assediment-water and air-water boundaries (Walneand Kivic, 1990) and should probably not be re-garded as open water truly planktonic algae (Lackey,1968).
Certain euglenoid algae are able to tolerateextreme environmental conditions. One of these,Euglena mutabilis, is able to grow in very low pHwaters. This alga has a pH optimum of pH 3.0, cantolerate values below pH 1.0, and is typical of acidicmetal-contaminated ponds and streams drainingmines. Other euglenoids, found in brackish habitats,are able to tolerate wide ranges in salinity (Walneand Kivic, 1990).
1.5.4 Euglenoids as bioindicators
Euglenoid algae are not particularly useful as en-vironmental bioindicators, either in terms of con-temporary populations or fossil records. Althoughpresent-day algae show some adaptations to specificenvironments (see above), there is no established en-vironmental library and species may be difficult toidentify. Within lake sediments, the lack of calci-fied or silicified structures that are resistant to decaymeans that hardly any remains have survived in thefossil record.
1.6 Yellow-green algae
Yellow-green algae (Xanthophyta) are non-motile,single celled or colonial algae, with a distinctive pig-mentation that gives the cells a yellow or fresh greenappearance (e.g. Fig. 4.18 – Tribonema). Althoughthere is a wide range in morphology (see below), thisphylum contains relatively few species (compared tomajor groups such as green algae) and the algae tendto be ecologically restricted to small water bodies anddamp soils.
1.6.1 Cytology
Yellow-green algae are most likely to be confusedwith green algae when observed in the fresh condi-tion, but differ from them (Table 1.3) in a number ofkey features.
� Distinctive pigmentation, with chlorophylls (a, c1
and c2 – but not b), carotenoids (especially β-carotene) and three xanthophylls (diatoxanthin,vaucheriaxanthin, heteroxanthin).
� Carbohydrate storage as oil droplets or chrysolam-inarin (usually referred to as leucosin) granules.
� Walls composed mainly of pectin or pectic acid(sometimes with associated cellulose or siliceousmaterial). The walls often occur as two splicedand overlapping sections, breaking into ‘H’-shapedpieces on dissociation of the filaments.
� When flagella are present they are of unequal length(heterokont) the longer one pointing forward andthe shorter (which can be more difficult to see usinga light microscope) sometimes pointing forwards orbackwards
� Chloroplast fine structure – four outer membranes(incorporating an endoplasmic reticulum) and thy-lakoids occurring in threes.
Cells contain two or more discoid and green toyellow-green chloroplasts, with associated pyrenoidsrarely seen. An eyespot may be present.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
24 1 INTRODUCTION TO FRESHWATER ALGAE
1.6.2 Morphological diversity
Yellow-green algae show a range of morphology,from unicellular to colonial and filamentous (Ta-ble 1.9). Unicellular forms are non-flagellate in themature (vegetative) state, with motility being re-stricted to biflagellate zoospores (asexual) or motilegametes (sexual) reproductive stages. The zoosporeshave unequal and morphologically dissimilar flag-ella (heterokont algae – Fig. 1.7). Unicellular al-gae include simple free-floating forms (Botrydiop-sis, Tetraedriella), or may have associated rhizoidalsystems or an attachment stalk. Some yellow-greenalgae form simple globular colonies, which may bemucilaginous and free floating (e.g. Gloeobotrys)or attached via a stalk (Ophiocytium). Filamentousforms may be unbranched (Tribonema – Fig. 4.18)or branched (Heterococcus), and reach their mostmassive state in the coenocytic siphonaceous generaBotrydium and Vaucheria (Fig. 4.3).
1.6.3 Ecology
Yellow-green algae are rather limited in their ex-ploitation of aquatic habitats, tending to occur ondamp mud (at the edge of ponds) and soil, but notoccurring extensively in either lentic or lotic sys-tems. Where planktonic forms do occur, they tend
to be present in ditches or small ponds. Tribonemaspecies are often found as free-floating filaments intemporary waters, and Botrydiopsis arrhiza is foundat the edge of ponds and in patches of water insphagnum bogs, where it may form yellowish wa-ter blooms. Even Gloeobotrys limneticus, which haswidespread occurrence as mucilaginous colonies inlake plankton, never forms extensive or dominantpopulations (as seen, for example, by blue-green al-gae). Various yellow-green algae are epiphytic, in-cluding Mischococcus and Ophiocytium (attached tofilamentous algae) and Chlorosaccus (attached tomacrophytes).
As with other algal groups, some yellow-green al-gae have developed alternative modes of nutrition.Chlamydomyxa is an amoeboid, naked form thathas retained its photosynthetic capability (still haschloroplasts) but has also become holozoic, ingest-ing desmids, diatoms and other algae and digestingthem within internal food vacuoles. Chlamydomyxais typical of low-nutrient acid bogs, and the holozoicmode of nutrition may be important for supplement-ing nitrogen uptake in such adverse conditions.
1.6.4 Yellow-green algae as bioindicators
Yellow-green algae have not been widely usedas bioindicator organisms, partly because they are
Table 1.9 Range of Morphologies in Yellow-Green Algae
Major Morphotype Attached or Planktonic Example*
UnicellsCoccoid Planktonic or benthic BotrydiopsisPear-shaped, with rhizoidal system Present on mud surface BotrydiumOvoid cells on stalk Epiphytic on attached or planktonic algae CharaciopsisAmoeboid Benthic, often in hollow cells of Sphagnum Chlamydomyxa
ColonialSmall to large globular colonies Planktonic GloeobotrysBranched colony on stalk Planktonic or attached Ophiocytium
FilamentsUnbranched Typically planktonic, occasionally attached
when youngTribonema (4.18)
Branched Attached, damp soils HeterococcusCoenocytic, (siphonaceous) Attached Botrydium, Vaucheria (4.3)
*Figure numbers in brackets
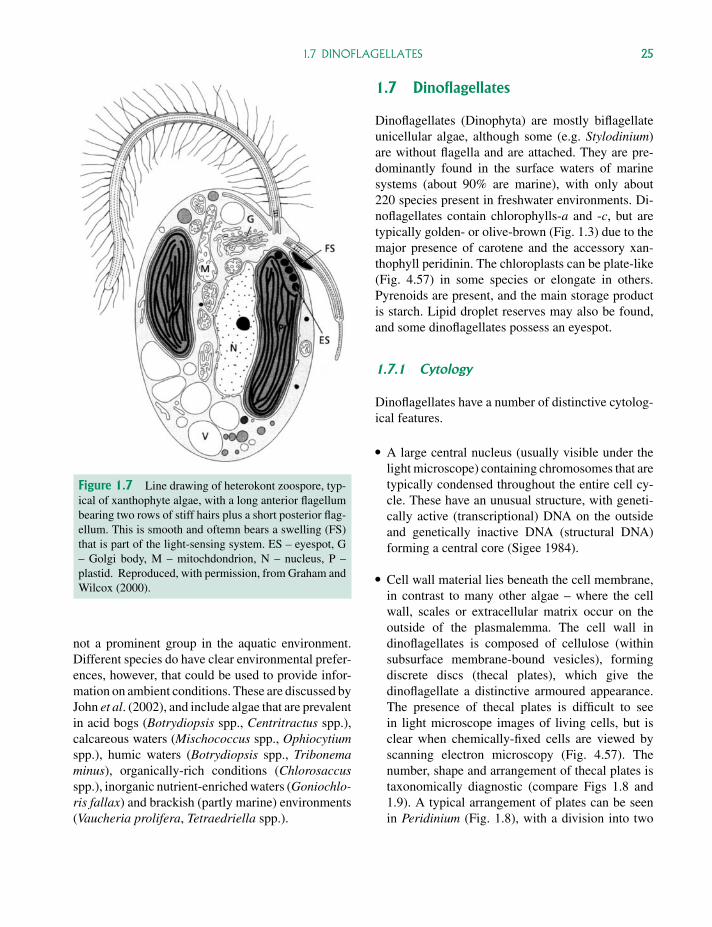
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.7 DINOFLAGELLATES 25
Figure 1.7 Line drawing of heterokont zoospore, typ-ical of xanthophyte algae, with a long anterior flagellumbearing two rows of stiff hairs plus a short posterior flag-ellum. This is smooth and oftemn bears a swelling (FS)that is part of the light-sensing system. ES – eyespot, G– Golgi body, M – mitochdondrion, N – nucleus, P –plastid. Reproduced, with permission, from Graham andWilcox (2000).
not a prominent group in the aquatic environment.Different species do have clear environmental prefer-ences, however, that could be used to provide infor-mation on ambient conditions. These are discussed byJohn et al. (2002), and include algae that are prevalentin acid bogs (Botrydiopsis spp., Centritractus spp.),calcareous waters (Mischococcus spp., Ophiocytiumspp.), humic waters (Botrydiopsis spp., Tribonemaminus), organically-rich conditions (Chlorosaccusspp.), inorganic nutrient-enriched waters (Goniochlo-ris fallax) and brackish (partly marine) environments(Vaucheria prolifera, Tetraedriella spp.).
1.7 Dinoflagellates
Dinoflagellates (Dinophyta) are mostly biflagellateunicellular algae, although some (e.g. Stylodinium)are without flagella and are attached. They are pre-dominantly found in the surface waters of marinesystems (about 90% are marine), with only about220 species present in freshwater environments. Di-noflagellates contain chlorophylls-a and -c, but aretypically golden- or olive-brown (Fig. 1.3) due to themajor presence of carotene and the accessory xan-thophyll peridinin. The chloroplasts can be plate-like(Fig. 4.57) in some species or elongate in others.Pyrenoids are present, and the main storage productis starch. Lipid droplet reserves may also be found,and some dinoflagellates possess an eyespot.
1.7.1 Cytology
Dinoflagellates have a number of distinctive cytolog-ical features.
� A large central nucleus (usually visible under thelight microscope) containing chromosomes that aretypically condensed throughout the entire cell cy-cle. These have an unusual structure, with geneti-cally active (transcriptional) DNA on the outsideand genetically inactive DNA (structural DNA)forming a central core (Sigee 1984).
� Cell wall material lies beneath the cell membrane,in contrast to many other algae – where the cellwall, scales or extracellular matrix occur on theoutside of the plasmalemma. The cell wall indinoflagellates is composed of cellulose (withinsubsurface membrane-bound vesicles), formingdiscrete discs (thecal plates), which give thedinoflagellate a distinctive armoured appearance.The presence of thecal plates is difficult to seein light microscope images of living cells, but isclear when chemically-fixed cells are viewed byscanning electron microscopy (Fig. 4.57). Thenumber, shape and arrangement of thecal plates istaxonomically diagnostic (compare Figs 1.8 and1.9). A typical arrangement of plates can be seenin Peridinium (Fig. 1.8), with a division into two
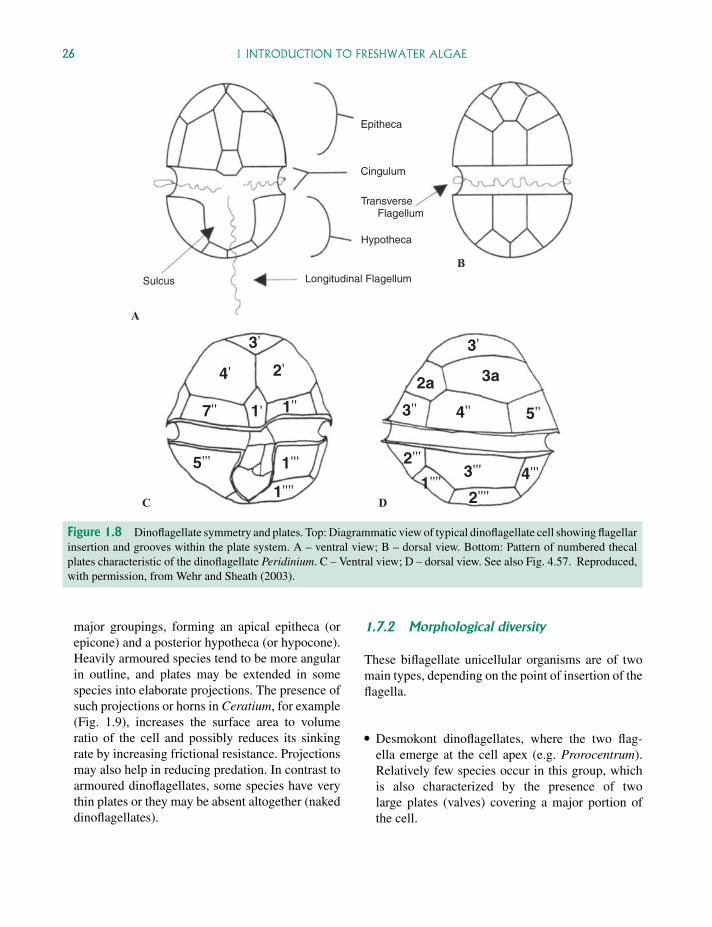
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
26 1 INTRODUCTION TO FRESHWATER ALGAE
A
B
C D
Epitheca
Cingulum
Hypotheca
Longitudinal FlagellumSulcus
4'
4''
4'''
7''
5'''
5''
1''''
2'
2''''
2'''
3'
3'''
3''
3a2a
3'
1''1'
1'''1''''
TransverseFlagellum
Figure 1.8 Dinoflagellate symmetry and plates. Top: Diagrammatic view of typical dinoflagellate cell showing flagellarinsertion and grooves within the plate system. A – ventral view; B – dorsal view. Bottom: Pattern of numbered thecalplates characteristic of the dinoflagellate Peridinium. C – Ventral view; D – dorsal view. See also Fig. 4.57. Reproduced,with permission, from Wehr and Sheath (2003).
major groupings, forming an apical epitheca (orepicone) and a posterior hypotheca (or hypocone).Heavily armoured species tend to be more angularin outline, and plates may be extended in somespecies into elaborate projections. The presence ofsuch projections or horns in Ceratium, for example(Fig. 1.9), increases the surface area to volumeratio of the cell and possibly reduces its sinkingrate by increasing frictional resistance. Projectionsmay also help in reducing predation. In contrast toarmoured dinoflagellates, some species have verythin plates or they may be absent altogether (nakeddinoflagellates).
1.7.2 Morphological diversity
These biflagellate unicellular organisms are of twomain types, depending on the point of insertion of theflagella.
� Desmokont dinoflagellates, where the two flag-ella emerge at the cell apex (e.g. Prorocentrum).Relatively few species occur in this group, whichis also characterized by the presence of twolarge plates (valves) covering a major portion ofthe cell.
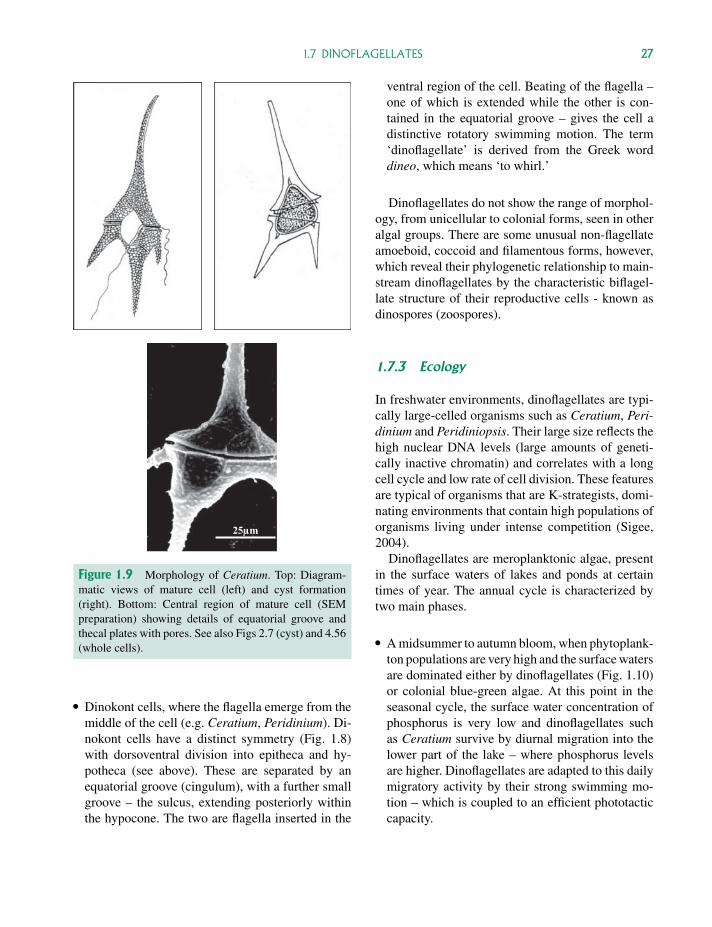
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.7 DINOFLAGELLATES 27
25µm
Figure 1.9 Morphology of Ceratium. Top: Diagram-matic views of mature cell (left) and cyst formation(right). Bottom: Central region of mature cell (SEMpreparation) showing details of equatorial groove andthecal plates with pores. See also Figs 2.7 (cyst) and 4.56(whole cells).
� Dinokont cells, where the flagella emerge from themiddle of the cell (e.g. Ceratium, Peridinium). Di-nokont cells have a distinct symmetry (Fig. 1.8)with dorsoventral division into epitheca and hy-potheca (see above). These are separated by anequatorial groove (cingulum), with a further smallgroove – the sulcus, extending posteriorly withinthe hypocone. The two are flagella inserted in the
ventral region of the cell. Beating of the flagella –one of which is extended while the other is con-tained in the equatorial groove – gives the cell adistinctive rotatory swimming motion. The term‘dinoflagellate’ is derived from the Greek worddineo, which means ‘to whirl.’
Dinoflagellates do not show the range of morphol-ogy, from unicellular to colonial forms, seen in otheralgal groups. There are some unusual non-flagellateamoeboid, coccoid and filamentous forms, however,which reveal their phylogenetic relationship to main-stream dinoflagellates by the characteristic biflagel-late structure of their reproductive cells - known asdinospores (zoospores).
1.7.3 Ecology
In freshwater environments, dinoflagellates are typi-cally large-celled organisms such as Ceratium, Peri-dinium and Peridiniopsis. Their large size reflects thehigh nuclear DNA levels (large amounts of geneti-cally inactive chromatin) and correlates with a longcell cycle and low rate of cell division. These featuresare typical of organisms that are K-strategists, domi-nating environments that contain high populations oforganisms living under intense competition (Sigee,2004).
Dinoflagellates are meroplanktonic algae, presentin the surface waters of lakes and ponds at certaintimes of year. The annual cycle is characterized bytwo main phases.
� A midsummer to autumn bloom, when phytoplank-ton populations are very high and the surface watersare dominated either by dinoflagellates (Fig. 1.10)or colonial blue-green algae. At this point in theseasonal cycle, the surface water concentration ofphosphorus is very low and dinoflagellates suchas Ceratium survive by diurnal migration into thelower part of the lake – where phosphorus levelsare higher. Dinoflagellates are adapted to this dailymigratory activity by their strong swimming mo-tion – which is coupled to an efficient phototacticcapacity.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
28 1 INTRODUCTION TO FRESHWATER ALGAE
200µm
Figure 1.10 Autumn dinoflagellate bloom in a eu-trophic lake. SEM view of epilimnion phytoplanktonsample (September), showing almost complete domi-nance by Ceratium hirundinella. See also Fig. 2.8 (sea-sonal cycle) and Fig. 4.56 (live Ceratium). Photo: Sigeeand Dean.
� An overwintering phase, where dinoflagellates sur-vive on the sediments as resistant cysts. Thesenon-flagellate cells (Fig. 2.7) lack an equatorialgroove and initially form in surface waters at theend of the summer/autumn bloom before sink-ing to the bottom of the lake. Germination ofcysts occurs in early summer, either on sedi-ments or recently mixed waters, giving rise tovegetative cells which take up newly availablephosphorus.
Ceratium and Peridinium are geographicallywidely dispersed, occurring particularly in waters thathave high calcium-ion concentrations (hard waters)and low levels of inorganic nutrients (see above). Al-though these two organisms are phototrophic, obtain-ing their nutrients in inorganic form, other dinoflag-ellates exhibit some degree of heterotrophy. Someare able to ingest food particles by engulfment ofwhole cells, formation of a feeding veil (pallium)or use of a feeding tube (phagopod). In some cases,these dinoflagellates are pigmented and mixotrophic,combining heterotrophic nutrition with photosynthe-sis. Other dinoflagellates such as Peridiniopsis areobligate heterotrophs and colourless. The common
freshwater heterotrophic dinoflagellate Peridiniopsisberolinensis uses a fine cytoplasmic filament to makecontact with suitable prey such as insect larvae, theningests the contents.
Heterotrophic dinoflagellates are particularlyadapted to conditions where photosynthesis is lim-iting. A number of colourless species, for example,are found under the ice of frozen lakes during thewinter season, where photosynthesis is severely lim-ited by low irradiance levels – which are about 1% ofsurface insolation.
1.8 Cryptomonads
Cryptomonads (Cryptophyta) are a group of rela-tively inconspicuous algae that are found in both ma-rine and freshwater environments. They are generallysmall to medium-sized unicells, and in many standingwaters are a relatively minor part of the phytoplank-ton assemblage – both in terms of cell numbers andbiomass.
1.8.1 Cytology
The fine structure of cryptomomads is illustrated bythe light microscope images of Rhodomonas (Fig.4.52) and Cryptomonas (Fig. 4.53), and by the linedrawing shown in Fig. 1.11. Each cell bears two flag-ella, which are about the same length as the celland slightly unequal. One of these flagella propelsthe cell, while the other is stiff and non-motile. Theflagella are inserted near to an anterior ventral de-pression, the vestibulum. The cells tend to be moreconvex on one side than the other and the front end,where the flagella arise, tends to be obliquely trun-cated. On this side arises a gullet, which is not al-ways clearly visible using a light microscope. Twoelongate chloroplasts are present in Cryptomonas,lying along the length of the cell (Fig. 4.53) andon either side of the midline. The colour of cryp-tomonad chloroplasts may be blue, blue-green, red-dish, olive green, brown or yellow-brown depend-ing upon the accessory pigments present. Thesemay include phycocyanin, phycoerythrin, caroteneand xanthophylls. A pyrenoid is present and starch

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.8 CRYPTOMONADS 29
Figure 1.11 Cryptomonad morphology. Diagram ofa non-photosynthetic cryptomonad with two flagellaemerging from a depression. Reproduced, with permis-sion from Graham and Wilcox (2000).
is stored. Further distinctive cytological features,include:
� a prominent contractile vacuole
� large ejectile organelles (ejectisomes), which dis-charge their content into the vestibular cavity andprobably act as a defence response to herbivorouszooplankton
� the presence of an internal ‘cell wall’ (the periplast)below the plasmalemma, which is composed ofround, oval, rectangular or hexagonal organicplates
� a complex flagellar root apparatus
� distinctive plastids, with the associated pres-ence of a much reduced nucleus – referred toas a nucleomorph; this phylogenetic survival isthought to be derived from the original (red algal)
symbiont that lead to the establishment of plastidsin cryptomonads.
1.8.2 Comparison with euglenoid algae
Cyptomonads share a number of similarities witheuglenoid algae. In both cases:
� algae are fundamentally biflagellate unicells, withflagella emerging from an apical depression
� cells are essentially naked, with a rigid covering ofthe cell occurring inside the plasmalemma
� species able to produce thick walled cysts that cansurvive adverse conditions
� different species occur in a variety of aquatic envi-ronments, both freshwater and marine
� heterotrophic (colourless) algae are well repre-sented alongside autotrophic (green) species.
In spite of these similarities, there are a numberof major differences between the algal groups, indi-cating a distinct phylogenetic origin: Key biochemi-cal differences include the presence of starch (stainsblue-black with iodine) in crypomonads, with pho-tosynthetic pigments which include phycobilins andshow some similarity to red algae. Ultrastructural dif-ferences also occur in terms of flagellar ornamenta-tion, and flagellar roots are different from euglenoids.The environmental biology of cryptomonads is alsoquite distinct from euglenoids, indicating a differentecological role for these organisms. Cryptomonadsare genuinely planktonic algae, widespread amongstthe plankton of a range of water bodies – in contrastto euglenoids, which tend to occupy mud, sand andwater interfaces.
1.8.3 Biodiversity
Cryptomonads are a relatively small group of algae,with a total of about 12 genera (van den Hoek et al.,1995) containing approximately 100 known freshwa-ter species and 100 known marine species.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
30 1 INTRODUCTION TO FRESHWATER ALGAE
Table 1.10 Variation in Plastid Number and Col-oration in Cryptomonads
Trophic State Plastid Characteristics*
Autotrophic Rhodomonas – a single boat-shaped,red coloured plastid (4.52)
Chroomonas – a single H-shapedblue-green plastid with a pyrenoidon the bridge.
Cryptomonas – two plastids, each witha pyrenoid (4.53)
Heterotropohic Chilomonas – a colourless cell thatcontains a plastid (leucoplast) thatlacks pigmentation. No pyrenoid
*Figure numbers in brackets.
The low diversity in terms of number of speciesis reflected in the uniformity of morphology. Thereare no clear colonial forms, though some species areable to form irregular masses embedded in mucilage(palmelloid stage) under adverse conditions. Differ-ences do occur in relation to plastids – includingpresence or absence, number and pigmentation (Ta-ble 1.10). Variations also occur in other key cytologi-cal features such as the presence or absence of nucle-omorphs, occurrence of ejectisomes and structure ofthe periplast – providing a basis for the classificationof these organisms (Novarino and Lucas, 1995).
1.8.4 Ecology
Cryptomonads are typical of temperate, high latitudestanding waters that are meso- to oligotrophic. Theyappear to be particularly prominent in colder waters,becoming abundant in many lakes in early spring,when they may commence growth under ice. Algaesuch as Rhodomonas may form red blooms whenthe ice melts in early spring. In other (Antarctic)lakes, where ice persists, cryptomonad species suchas Cryptomonas sp. and Chroomonas lacustris maycontribute up to 70% of the phytoplankton biomass,dominating the algal flora for the whole of the limitedsummer period (Spaulding et al., 1994).
Cryptomonads are also typical of the surface wa-ters of temperate lakes during the clearwater phaseof the seasonal cycle, between the diatom spring
bloom and the beginning of the mixed summerbloom. During this period, when algal populationsare severely predated by zooplankton populations,the limited competition between algal cells favoursrapidly-growing r-selected organisms such as cryp-tomonads (Sigee, 2004).
In oligotrophic freshwater lakes, cryptomonadsoften form large populations about 15–20 m deepwithin the lake, where oxygenated surface watersinterface with the anoxic lower part of the watercolumn. In this location, these algae form deepwater accumulations of photosynthetic organisms,referred to as ‘deep-chlorophyll maxima’. Studies oncultures of Cryptomonas phaseolus and C. undulataisolated from deep chlorophyll maxima (Gervais,1997) showed that these organisms had optimalgrowth under light-limiting conditions, suggestingphotosynthetic adaptation to low light intensities.Other factors which may be important in the abilityof these organisms to form large depth populationsinclude – the ability for heterotrophic nutrition,relative absence of predation by zooplankton,access to inorganic nutrients regenerated by benthicdecomposition and tolerance of sulfide generated inthe reducing conditions of the hypolimnion.
The ability of cryptomonads to carry out het-erotrophic nutrition, using ammonium and organicsources as a supply of carbon and nitrogen is eco-logically important. Evidence for uptake of partic-ulate organic material comes from direct observa-tion of phagocytosis and has been documented inboth pigmented (e.g. Cryptomonas ovata) and vari-ous colourless species (Gillott, 1990). Ultrastructuralstudies also provide evidence that many pigmentedcryptomonads are mixotrophic, capable of both au-totrophic and heterotrophic nutrition. The blue-greencryptomonad, Chroomonas pochmanni, has a special-ized vacuole for capturing and containing bacterialcells. Bacteria are drawn into this through a smallpore at the cell surface, and subsequently pass intodigestive vesicles within the cytoplasm.
1.8.5 Cryptomonads as bioindicators
Although they are common in oligo- and mesotrophicconditions, cryptomonads also occur in eutrophiclakes (Fig. 2.16) and have limited use as

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.9 CHRYSOPHYTES 31
bioindicators. Individual species do not provide thecontemporary diagnostic range for different habitatsseen in other algal groups, and cryptomonads do notsurvive as clearly identifiable remains within lakesediments.
1.9 Chrysophytes
Chrysophytes (Chrysophyta), commonly referred toas the ‘golden algae’, are a group of microscopic al-gae that are most readily recognized by their golden-brown colour (Figs 1.3 and 4.37). This is due tothe presence of the accessory pigment fucoxan-thin within the chloroplasts, masking chlorophylls-aand -c (Table 1.3). There are around 200 generaand 1000 species of chrysophytes, present mainly infreshwater although some may be found in brackishand salt waters.
1.9.1 Cytology
Distinguishing cytological features (Table 1.3) in-clude the presence of two dissimilar (long andshort) flagella (heterokont condition) in motile organ-isms, plastids with four outer membranes plus triplet
thylakoids, chrysolaminarin as the main storage prod-uct (present in special vacuoles) and cell walls com-posed of pectin – covered in some species with smallsilica spines or scales. Lipid droplets also frequentlyoccur, and an eyespot may be visible in some species,associated with the chloroplast. In order to surviveadverse conditions round walled cysts (stomatocysts)may be produced, with silica scales present in somespecies. Although most chrysophyte species are pho-tosynthetic, some may be partially heterotrophic oreven fully phagotrophic (Pfandl et al., 2009). Manyspecies are small and are important members of thenanoplankton. They are widely distributed through-out freshwater systems, and are of particular interestin relation to their morphological diversity, ecologyand as indicator organisms. Details of taxonomy andultrastructure are given by Kristiansen (2005).
1.9.2 Morphological diversity
Chrysophytes exhibit considerable diversity in theirgeneral organization, ranging from unicellular tospherical (e.g. Synura – Fig. 4.37) and branching(e.g. Dinobryon – Fig. 4.2) colonial types. The greatmorphological diversity shown by these organisms(Table 1.11) follows a similar diversity seen in other
Table 1.11 Range of Body Structure and Form in Chrysophytes
General Structure Colony Status Examples*
Flagellate cells (monads)Simple cell wall Single cells: Chromulina,
OchromonasColonies Uroglena
Surrounded by envelope (lorica) Colony Dinobryon (4.2)Cells covered in silica scales Single cells Mallomonas
Colony Synura (4.37)Non-flagellate unicellsMotile by pseudopod Golden amoebae ChrysamoebaNon-motile Cells surrounded by a jelly Chrysocapsa
Cells surrounded by solid cell wall StichogloeaColonies with distinctive
morphologySimple branched filaments Sphaeridiothrix
PhaeothamnionTwo-dimensional cell plates Phaeoplaca
*Figure numbers in brackets.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
32 1 INTRODUCTION TO FRESHWATER ALGAE
algal phyla and includes flagellates, amoeboid forms,cells in jelly (capsal), filamentous algae and plate-like(thalloid) organisms. Each of these types, however,can transform into another phase (e.g. flagellates canbecome amoeboid) – so that grouping in relation tomorphology is dependent on the stage of life cycle.The occurrence of a resistant (stomatocyst) phase (seeabove) is also a key part of the life cycle.
1.9.3 Ecology
Chrysophytes are ecologically important in a numberof ways (Kristiansen, 2005).
� In many ecosystems they play an important roleas primary producers. This is particularly the casefor adverse conditions – such low nutrient and acidlakes (e.g. Nedbalova et al., 2006).
� They have a versatile nutrition, with many mem-bers (e.g. Dinobryon) being mixotrophic. These or-ganisms are able, in addition to photosynthesis, toobtain their carbon from organic sources – eitherby surface absorption of organic compounds or byphagocytosis of particulate organic matter.
� They can be nuisance algae, giving a fishy smellto drinking water reservoirs when they reach highpopulation levels.
1.9.4 Chrysophytes as bioindicators
Their diverse ecological preferences (Table 1.12)make chrysophyte species potentially useful as envi-ronmental indicator organisms – both in terms of con-temporary water assessment and sediment analysis.
Contemporary water quality
Although potentially very useful, chrysophytes arerarely used in monitoring projects. The main reasonfor this is that the best ecologically-studied species(with silica scales) require electron scanning mi-
Table 1.12 Chrysophyte Species as Bioindicators
Strongly acid,often humicconditions
Dinobryon pediforme, Synurasphagnicola, Mallomonaspaludosa, M. hindonii,M. canina
Weakly buffered,slightly acidclear water lakes
Mallomonas hamata, Synuraechinulata, Dinobryonbavaricum var. vanhoeffenii
Alkalineconditions
Mallomonas punctifera, Synurauvella
Alkaline/salineconditions
Mallomonas tonsurata,M. tolerans
croscopy for identification – a technique not routinelyused as a tool in water quality assessment.
Although chrysophytes have traditionally beenheld to indicate oligotrophic conditions, the situa-tion is more complex. Studies by Kristiansen (2005)have shown that the presence of a few species such asUroglena or Dinobryon may indicate oligotrophy, butgreater chrysophyte species diversity at lower overallbiomass is more indicative of eutrophic conditions.The value of individual chrysophyte species as eco-logical indicators varies considerably (Kristiansen,2005). Synura petersenii, with a broad ecologicalrange, is of limited use – but other narrow-rangespecies (Table 1.12) can be used particularly in re-lation to pH and salinity.
Sediment analysis
The main application of chrysophytes as environ-mental bioindicators is in paleoecology. Their ma-jor advantage for this (as with diatoms) is the pres-ence of non-degradable silica cell wall material, sothat they remain undamaged in the lake sediments.The use of these algae in paleoecology depends onaccurate means of classification and identification,undisturbed sediment layers and absolute dating ofthe layers – e.g. by the lead isotope Pb210.
Two main types of chrysophyte remains are usefulin freshwater sediments – silica scales (from veg-etative planktonic cells) and stomatocysts (resistantspores). Stomatocysts have the advantage of betterpreservation compared to scales, but the disadvantage

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.10 DIATOMS 33
that (with a few exceptions) they cannot be referred toparticular species. Although stomatocysts cannot beidentified in terms of species, they can be recorded asdistinct morphological types (morphotypes). Thesecan be correlated with other microfossils (e.g. di-atoms) and with pollen to obtain environmental indi-cator values. Quantitative assessment of past aquaticenvironments in relation to fossil chrysophytes (stom-atocysts) parallels that of diatoms (Section 3.2.2) andcan be obtained in relation to species ratios (en-vironmental indices), transfer functions (Facher &Schmidt, 1996) and group analysis.
Species ratios The ratio of chrysophytes to otheralgal groups may provide a useful index to assess en-vironmental change. Studies by Smol (1985) on thesurface sediments of Sunfish Lake (Canada) demon-strated a marked increase in the ratio of diatoms tosomatocysts, coincident with the arrival of settlers tothe lake catchment area. The diatom/somatocyst ratiois a trophic index, signalling an increase in eutrophicstatus of the lake at the time of human settlement.The above ratio change resulted from a populationdecrease in the chrysophyte Mallomonas (indicatingoligotrophic conditions) and an increase in eutrophicdiatoms. The increase in trophic ratio parallels an in-crease in the pollen count of ragweed, an indicator offorest clearance and farming activity.
Transfer functions The environmental proper-ties of stomatocyst morphotypes can be assigned asnumerical descriptors or ‘transfer functions.’ Thesedefine each morphotype in relation to a range of en-vironmental data (pH, conductivity, phosphorus con-tents and maximal depth of the lake).
The transfer functions have been derived by mul-tivariate analysis of sediment calibration sets, ob-tained from a series of North American lakes inwhich both environmental data and stomatocyst as-semblages were available (Charles and Smol, 1994).
Group analysis The various environmental fac-tors which influence correlations between morpho-types can be identified by statistical analysis – partic-ularly principal component analysis (PCA). Studiesby Duff and Smol (1995) on 181 morphotypes ob-tained from 71 different lakes (in the United States)
showed that differences in pH and lake morphometryhad the greatest influence in determining statisticalassociations. PCA analysis separated morphotypesinto three main groups, respectively relating primarilyto acid environments, neutral/alkaline environmentsand lake morphometry (e.g. lake depth).
1.10 Diatoms
Diatoms (Bacillariophyta) are a very distinct groupof algae, identifiable under the light microscope bytheir yellow-brown coloration (Fig. 1.13) and by thepresence of a thick silica cell wall. This cell wall typ-ically appears highly refractive under the light micro-scope, giving the cell a well-defined shape. Removalof surface organic material by chemical oxidation(Section 2.5.4) reveals a complex cell wall ornamen-tation – as illustrated in the light microscope im-ages comparing living and digested cells of Stephan-odiscus (Fig. 4.58). Cell wall ornamentation can beseen even more clearly with the higher resolution ofthe scanning electron microscope and is important inspecies identification.
Diatoms occur as non-flagellate single cells, simplecolonies or chains of cells, and are widely-occurringin both marine and freshwater environments. Theirsuccess in colonizing and dominating a wide rangeof aquatic habitats is matched by their genetic diver-sity – with a worldwide total of 285 recorded genera,encompassing 10 000–12 000 species (Round et al.,1990; Norton et al., 1996). Diatoms are also veryabundant in both planktonic and benthic freshwaterenvironments, where they can form a large part of thealgal biomass, and are a major contributor to primaryproductivity.
1.10.1 Cytology
Distinctive cytological features (Table 1.3) include:
� plastids with periplasmic endoplasmic reticulumand the presence of girdle lamellae
� chrysolaminarin and lipid food reserves, presentoutside the plastid

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
34 1 INTRODUCTION TO FRESHWATER ALGAE
� a distinctive cell wall, the frustule, composed ofopaline silicon dioxide (silica) together with or-ganic coatings.
Silica cell wall
The presence of silica in the diatom cell wall can bedetected by cold digestion in a strongly oxidizing acidto remove organic matter (Section 2.54). If the cellwall resists this treatment it is probably silica. Thecell wall of diatoms differs from that of other algaein being almost entirely inorganic in composition. Ithas evolved as an energy-efficient structure, requiringsignificantly less energy to manufacture than the cel-lulose, protein and mucopeptide cell walls of otheralgae (Falkowski and Raven, 1997). This gives di-atoms a major ecological advantage at times whenphotosynthesis is limited (e.g. early in the seasonalcycle), but has a number of potential disadvantages.
� The cell wall of diatoms is very dense. These or-ganisms are only able to stay in suspension inturbulent conditions, limiting the development ofextensive planktonic diatom populations to unstrat-ifiied waters.
� The formation of the diatom cell wall is depen-dent on an adequate supply of soluble silica (silicicacid) in the surrounding water. The diatom springbloom of temperate lakes strips out large quantitiesof silica from the water, reducing concentrations toa level that becomes limiting.
� Unlike other types of cell wall material, silica isrigid and unable to expand. This means that daugh-ter cells are unable to enlarge and progressive celldivisions result in a gradual decrease in cell size.Ultimately this decrease reaches a critical level,at which point sexual reproduction is required tocompletely shed the original cell wall and formnew, large daughter cells.
Cell wall structure
The frustule is composed of two distinct halves – theepitheca and hypotheca – which fit together ratherlike the lid and base of a pill-box (Fig. 1.12). Atcell division, two new walls are formed internallyat the cell equator ‘back to back’, and become thehypothecae of the two daughter cells. The originalepitheca and hypotheca of the parent diatom become
A
B C
D E
1
2
3
4
5aa
aa
ta ta
e
h
e
h
eh
Figure 1.12 Diatom frustulestructure – comparison of centricand pennate diatoms. Centricdiatom: A. Separate views ofepitheca (e) 1. Epivalve 2. Mantleof epivalve. 3. Epicingulum andhypotheca (h) 4. Hypocingulum. 5.Hypovalve. B. Complete frustule –girdle or side view (showing overlapof cingula). C. Valve or face viewof epivalve. Pennate diatom: D.Complete frustule – girdle view.E. Valve view, showing apical orlongitudinal axis (aa) and transapicalor transverse axis (ta).

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.10 DIATOMS 35
the epithecae of the two daughter cells, so the epithecais always the oldest part of the frustule.
In both centric and pennate diatoms (see below)the epitheca consists of two main parts – a circu-lar disc (the valve) with a rim (the mantle) plusan attached band or girdle (the epicingulum). Thehypotheca correspondingly consists of a hypovalve,hypovalve mantle and hypocingulum. When the ep-itheca and hypotheca fit together to form the completefrustule the hypocingulum fits inside the epicingulumto give an overlap.
Because of frustule morphology (Fig. 1.12), di-atoms can be observed from either a girdle (side)view or from a valve (face) aspect – with girdle andvalve views of live diatoms being clearly distinguish-able under the light microscope (Fig. 1.13). The twopossible valve views, of the top (epivalve) or bottom(hypovalve) of the diatom, are not distinguishable inmany genera – but do differ in some cases (e.g. Coc-coneis, Achnanthes). Axes of symmetry in pennatediatoms are shown in Figs 1.12 and 1.13.
Frustule markings
A wide range of surface markings can be seen onthe face (epivalve and hypovalve) of diatoms. Thesehave been recorded in considerable detail (Barber andHaworth, 1981; Round et al., 1990; Wehr and Sheath,2003) and form the basis for the classification andidentification of these organisms (Chapter 4).
Clear visualization of the frustule markings re-quires the high resolution of either oil immersion(light microscopy) or scanning electron microscopy,and is normally carried out after the removal ofsurface organic matter by chemical (acid digestion)cleaning. Chemical fixation for scanning electron mi-croscopy may also strip away some of the overlyingorganic material to reveal frustule surface structure.
The terminology of diatom morphology (seeGlossary) includes various descriptors of frustulemarkings – including eye-shaped structures (ocelli),small pores (punctae) and fine lines (striae). Illus-trations of diatom surface markings are shown dia-grammatically in Fig. 1.14 and in various figures andplates in Chapter 4. Although the biological signifi-cance of much of this surface detail is obscure, the
presence of one major surface structure – the raphe– is clearly associated with locomotion. The secre-tion of mucus from this channel or canal promotesmovement on solid surfaces. In some diatoms suchas Nitzschia (Fig. 4.70a, b) the raphe is elevated fromthe main diatom surface as a keel, allowing more in-timate contact between the raphe and substrata. Suchkeeled diatoms are able to move particularly well onfine sediments, and reach their maximum abundancein the epipelon of pools and slowly-flowing streams(Lowe, 2003). A raphe is not always present in pen-nate diatoms, and is never seen in centric diatoms(see below).
1.10.2 Morphological diversity
The morphological diversity of diatoms can be con-sidered in relation to two main aspects – the distinc-tion between centric/pennate diatoms and the rangeof unicellular to colonial forms.
Centric and pennate diatoms
Diatoms can be separated into two major group-ings – centric and pennate diatoms, based primarilyon cell shape and frustule morphology (Fig. 1.12).Centric diatoms typically have a discoid or cyclindri-cal shape, with a radial symmetry when seen in face or‘valve’ view. Pennate diatoms have an elongate bilat-eral symmetry, with longitudinal and transverse axes,and often have the appearance of a ‘feathery’ (hence‘pennate’) ornamentation when seen in surface view(Fig. 1.13). Pennate diatoms have a range of shapes(Barber and Haworth, 1981), with broad division(Fig. 1.15) into isopolar (ends of valve similar in sizeand shape), heteropolar (ends of valve differing in sizeand shape), asymmetrical (outline dissimilar eitherside of the longitudinal axis) and symmetrical (outlineof valve similar either side of the longitudinal axis).
The fundamental differences between centric andpennate diatoms in relation to structure and symme-try reflect a range of other distinguishing features(Table 1.13) – including motility, number and sizeof plastids, sexual reproduction and ecology. Cen-tric diatoms are mainly planktonic algae, and theoccurrence of oogamy – with the production of a
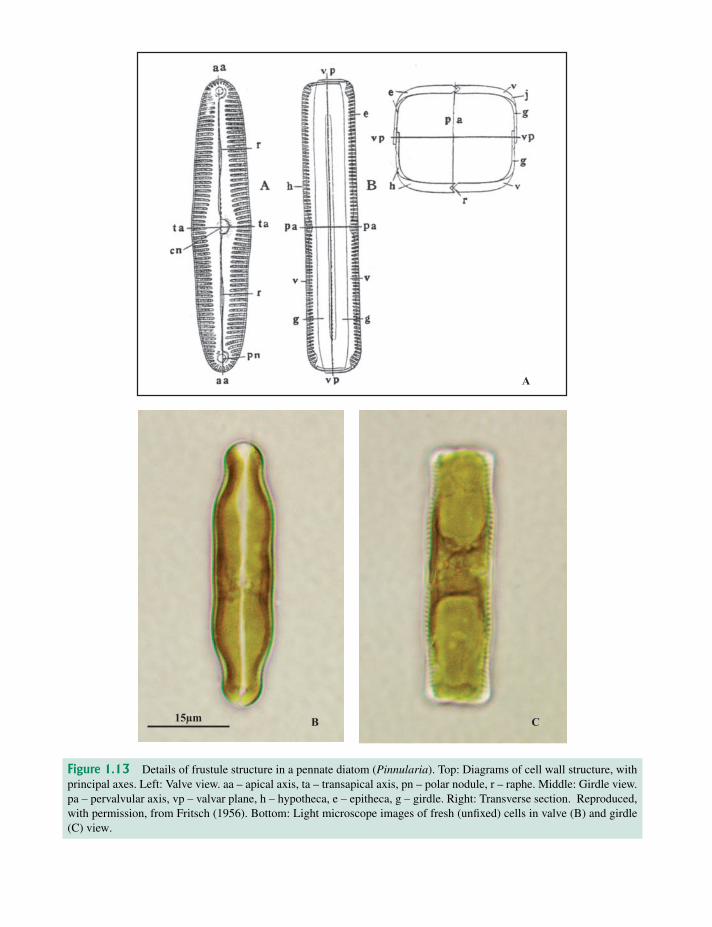
P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
A
B C 15µm
Figure 1.13 Details of frustule structure in a pennate diatom (Pinnularia). Top: Diagrams of cell wall structure, withprincipal axes. Left: Valve view. aa – apical axis, ta – transapical axis, pn – polar nodule, r – raphe. Middle: Girdle view.pa – pervalvular axis, vp – valvar plane, h – hypotheca, e – epitheca, g – girdle. Right: Transverse section. Reproduced,with permission, from Fritsch (1956). Bottom: Light microscope images of fresh (unfixed) cells in valve (B) and girdle(C) view.
36

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.10 DIATOMS 37
Figure 1.14 Diatom valve markings. 1 internal septa;2 transapical costae; 3 raphe in thickened ribs; 4 normalraphe; 5 shortened raphe in ribs; 6 parallel striae; 7 radiatestriae; 8 central area round; 9 central area transverse;10 central area small; 11 central area acute angled; 12punctae in radial rows interspersed with subradial rows;13 coarse areolae; 14 areolae grouped in segments; 15sections of valve alternately raised and depressed givingshaded appearance; 16 central region raised or depressed;17 pennate with raised or depressed areas; 18 ocelli onsurface; 19 rudimentary raphe as in Eunotia; 20 raphecurved to point at centre as in Epithemia. Reproduced,with permission, from Bellinger, 1992.
large number motile sperm, is regarded as a strategyfor increasing the efficiency of fertilization in openwater environments. Pennate diatoms are almost al-ways isogamous, with equal sized non-flagellate ga-metes (two per parent cell). In these algae, efficiencyof fertilization is promoted by the pairing of gameteparental cells prior to gamete formation, a possible
adaptation to more restricted benthic environments.Fossil and molecular evidence suggests that centricdiatoms arose prior to pennate ones, implying that inthese algae isogamy has been phylogenetically de-rived from oogamy (Edlund and Stoermer, 1997).This is in contrast to the evolutionary sequence nor-mally accepted for other algal groups - particularlygreen and brown algae.
Unicellular and colonial diatoms
The genetic diversity of diatoms is not matched bythe complexity of their morphological associations –which are limited to small chains and groups of cells.In planktonic diatoms, simple filamentous (e.g. Aula-coseira – Fig. 4.9) and radial (e.g. Asterionella –Figs 1.16, 4.42) colonies have evolved, but there isno formation of spherical globular colonies that are akey feature of some other algal groups.
Colonial forms of diatoms arise by means of simplelinkages, which are of three main types.
� Mucilage pads. These occur at the ends of cellsto join diatom cells in various ways. Asterionellahas elongate cells which are joined at their innerends to form stellate colonies. In Tabellaria, cellsare linked valve to valve as short stacks, with con-nection of the stacks at frustule edges by mucilagepads to form a zig-zag pattern. Mucilage pads areseen particularly clearly in Fig. 2.28, where chainsof Tabellaria are attached to Cladophora.
� Interlinking spines. In diatoms such as Aulacoseira,chains of cells are joined valve to valve by spines.The valves are of two main types (Davey and Craw-ford, 1986)
– Separation valves, with long tapering spines andstraight rows of pores,
– Linking valves, with short spines and curved rowsof pores
Cells are strongly linked into chains via the shortspines on the linking valves, but weakly linked viathe long spines of the separation valves – where

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
38 1 INTRODUCTION TO FRESHWATER ALGAE
Figure 1.15 Diatom morphology.1 pennate diatom with raphe; 2with pseudoraphe; 3 sulcus (s) onMelosira; 4 spines on margin ofStephanodiscus; 5 valve shapes: (a)isopolar, (b) heteropolar, (c) asym-metrical, (d) symmetrical. Re-produced, with permission, fromBellinger, 1992.
filament breakage easily occurs (Fig. 4.9). Thelength of filaments within a population of Aula-coseira depends on the proportion of separationvalves to linking valves.
� Gelatinous stalks. Biofilm diatoms such asRhoicosphenia (Fig. 4.67) and Gomphonema (Fig.2.29) have gelatinous stalks, which attach the di-atom to the substratum and join small groups ofcells together as a single colony. In these diatoms,the stalk material is secreted via apical pores. Some
species of Cymbella also produce a gelatinous tube-like filament.
1.10.3 Ecology
Diatoms are ubiquitous in both standing and runningfreshwaters – occurring as planktonic, benthic, epi-phytic (on higher plants and other algae) and epizoic(on animals, such as zooplankton) organisms. Theyare equally successful as free floating and attached
Table 1.13 Centric and Pennate Diatoms
Characteristic Centric Diatoms Pennate Diatoms
Symmetry Radial Bilateral
Examples* Stephanodiscus (4.58)Cyclotella (4.62)
Pinnularia (1.13, 4.71)Navicula (4.73)
Gliding motility Non-motile Some (raphid) diatoms are actively motile
Plastids Many discoid plastids Two large plate-like plastids
Sexual reproduction†
Initiation Independent formation of gametes from parentcells
Pairing of parental cells prior to gameteproduction
Egg cells Oogamous – production of one or two eggs perparent cell
Isogamous
Sperm cells 4–128 sperm per parent cell. Each with a singleflagellum bearing two rows of mastigonemes
Few amoeboid, non-flagellate sperm cells
Ecology Mainly planktonic, typical of open water Planktonic, epiphytic and benthic forms
*Figure numbers in brackets.†Hasle and Syvertsen, 1997.

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
1.10 DIATOMS 39
100µm A
Au
d
Figure 1.16 Spring diatom bloom in a eutrophiclake. SEM view of lake surface phytoplankton sample(March), showing numerous stellate colonies of Asteri-onella (A) with some filamentous Aulacoseira (Au). Thelarge amount of debris (d) occurs due to the completemixing of the water column at this time of year. See alsoFigs 2.8 (seasonal cycle), 4.8 (live Aulacoseira) and 4.42(live Asterionella). Photo: Sigee and Dean.
forms, where they may be respectively important inplankton bloom formation and biofilm development.
Planktonic diatoms: bloom formation
In many temperate lakes, diatoms such as Asteri-onella and Tabellaria dominate the phytoplanktonpopulation in Spring and early Summer (Fig. 1.16),at a time when inorganic nutrients (N, P, Si) are at highconcentration, light and temperature levels are risingand lake turbulence is maintained by moderate windaction. At this point in the seasonal cycle, diatoms areable to out-compete other micro-algae due to their tol-erance of low temperature and low light conditions,coupled with their ability to grow in turbulent wa-ter. Autumn blooms of Fragilaria and Asterionellaare also a common feature of many lakes, with thediatoms part of a mixed phytoplankton population.
Diatoms such as Aulacoseira italica are regardedas meroplanktonic, with major growth in very earlySpring, but spending the rest of the year as resis-tant cells on lake sediments. Other diatoms, such asAsterionella and Tabellaria also form major bloomsin Spring, but are additionally present as minor
constituents within the mixed phytoplankton popula-tion over much of the annual cycle – and are regardedas holoplanktonic.
Attached diatoms: biofilm formation
Diatoms are major components of biofilms, wherethey are present early in the colonization sequenceand also within the mature periphyton community.See later for examples of diatoms in river biofilm(Fig. 2.23), reed biofilm (Fig. 2.29) and as attachedepiphytes (Fig. 2.28).
1.10.4 Diatoms as bioindicators
The major use of diatoms as bioindicators of waterquality (both lotic and lentic systems) is detailed inChapter 3. These algae can be isolated from existinglive populations (contemporary analysis) or from sed-iments (fossil diatoms) and quantitative informationon water quality from species counts can be obtainedfrom taxonomic indices, multivariate analysis, trans-fer functions and species assemblage analysis.
Contemporary analysis
Many diatom species have distinct ecological prefer-ences and tolerances, making them useful indicatorsof contemporary ecological conditions. These prefer-ences relate to degree of water turbulence, inorganicnutrient concentrations, organic pollution, salinity,pH and tolerance of heavy metals. Round (1993) alsolists a number of other advantages in using diatomsas indicators – including easy field sampling, highsensitivity to water quality but relative insensitivityto physical parameters in the environment and easycell counts. Diatoms are also the most diverse groupof algae present in freshwaters, making them themost ideal assemblage for calculation of bioindices(Section 3.4.5).
Diatoms, once cleaned and mounted for identifi-cation, make excellent permanent slides which canform an important historical record for a location.Although there are hundreds of diatom species thatmay be present only dominant species need to beused in an assessment. Benthic diatoms have been

P1: OTA/XYZ P2: ABCc01 JWBK440/Bellinger March 15, 2010 11:55 Printer Name: Yet to Come
40 1 INTRODUCTION TO FRESHWATER ALGAE
particularly useful in assessing water quality of rivers,and the wide range of indices that have been devisedis discussed in Chapter 3.
Lake sediments
Diatoms, more than any other group of algae, are usedto monitor historical ecological conditions from sed-iment analysis – as discussed in Section 3.2.2. Themajor role of diatoms in this lies in the resistance ofthe diatom frustule to biodegradation, coupled withease of identification from frustule morphology andthe wide range of species with clear ecological pref-erences see in freshwater environments.
The importance of diatoms in environmental anal-yses is indicated by the European Diatom DatabaseInitiative (EDDI). This key web-based site includeselectronic images and data handling software, and isparticularly useful for lake sediment analysis (Sec-tion 3.2.2). Other databases, such as the stream ben-thic database of Gosselain et al. (2005), are moreapplicable to contemporary environmental analysis.
1.11 Red algae
Red algae (Rhodophyta) are predominantly ma-rine in distribution, with only 3% of over 5000species worldwide occurring in true freshwater habi-tats (Wehr and Sheath, 2003). Although termed red-algae, the level of accessory pigments (phycoery-thrin and phycocyanin) may not be sufficient to maskthe chlorophyll – resulting in an olive-green to blue(rather than red) coloration. The relatively commonriver alga Batrachospermum, for example, typicallyappears bluish-green in colour (Fig. 4.4) – giving noclue to its rhodophyte affinity.
Major cytological features of red algae (Table 1.3)include the absence of flagellae, presence of florideanstarch as the major food reserve, characteristic pho-tosynthetic pigments (chlorophyll-a only, presenceof phycobilins) and distinctive plastid structure(unstacked thylakoids, no external endoplasmicreticulum).
Many of the freshwater species (which occurmainly in streams and rivers) are quite large, being
visible to the naked eye when occurring in reasonablenumbers (Sheath and Hambrook, 1990). The range ofmorphologies include gelatinous filaments (e.g. Ba-trachospermum), pseudoparenchymatous forms (e.g.Lemanea) and a flat thallus of tiered cells (Hilden-bandia). Many of these shapes can be an advantage inresisting the forces exerted in swiftly flowing waters.
Although freshwater red algae (such as the large fil-amentous alga Batrachospermum) are largely foundin streams and rivers, Rhodophyta may also occuras marine invaders of lakes and brackish environ-ments. Certain freshwater red algae in the littoralzones of the Great Lakes Basin (United States), forexample, appear to be originally marine and to havelost the capacity for sexual reproduction. These in-clude the filamentous red alga Bangia atropurpurea(Lin and Blum, 1977), which reproduces only byasexual monospores – in contrast to marine specieswhich undergo alternation of generations and carryout sexual reproduction. Attached red algae (e.g.Chroodactylon ramosum) also contribute to the epi-phytic flora of lake periphyton.
1.12 Brown algae
As with red algae, brown algae (Phaeophyta) are al-most entirely marine – with less than 1% of speciespresent in freshwater habitats (Wehr and Sheath,2003). These species are entirely benthic, either inlakes or rivers, and have a very scattered distribution.
Cytological features of this group (Table 1.3) in-clude heterokont flagella (reproductive cells only),presence of laminarin as the major food reserve, char-acteristic photosynthetic pigments (chlorophyll-a and-c, β and ε carotenes) and distinctive plastid struc-ture (triple thylakoids, enclosing endoplasmic reticu-lum). Freshwater brown algae include genera suchPleurocladia and Heribaudiella, and are the leastdiverse of all freshwater algae. Their morphologiesare based on a relatively simple filamentous structure(forming mats or crusts), and they lack the complexmacro-morphology typical of the brown seaweeds.The freshwater brown-algae have not been fully stud-ied and their ecological characteristics are not wellknown. All are benthic and may be recognized bytheir possession of large sporangia (Wehr, 2003).





























