Embed Size (px)
Citation preview

421
During the past year, significant advances have been made inour understanding of stomatal development and its response toclimate change, and in our knowledge of how guard cell Ca2+
oscillations encode environmental signals. Recent studies on(de)phosphorylation mechanisms have provided new informationon how guard cells respond to abscisic acid and blue light.
AddressesBiology Department, Pennsylvania State University, 208 Mueller Laboratory, University Park, Pennsylvania 16802-5301, USA*e-mail: [email protected]†e-mail: [email protected]
Current Opinion in Plant Biology 2001, 4:421–428
1369-5266/01/$ — see front matter© 2001 Elsevier Science Ltd. All rights reserved.
AbbreviationsAAPK ABA-activated protein kinaseABA abscisic acidabi1-1 ABA-insensitive1-1 cADPR cyclic ADP-riboseHIC HIGH CARBON DIOXIDEIP3 inositol 1,4,5-trisphosphateNAADP nicotinic acid adenine dinucleotide phosphatePLC phosphoinositide-specific phospholipase CPLD phospholipase DS1P sphingosine 1-phosphatesdd1 stomatal density and distribution1tmm too many mouths
IntroductionStomata, located in the plant epidermis, consist of a pair ofguard cells and the pore they enclose (Figure 1a,b).Stomata open and close through turgor changes driven bymassive ion fluxes, which occur mainly through the guardcell plasma membrane and tonoplast.
The guard cell has become a model system for the studyof signal transduction. Guard cell signaling is consideredto be intriguing, not only because stomates play animportant role in the plant’s response to its variableenvironment but also because the guard cell is mechanicallyseparate from the surrounding cells, and because guardcells have sensitive reversible reactions to internal andexternal stimuli. A detailed analysis of guard cell signaltransduction may be useful for studies of other plantsignaling systems such as, pulvinar movement, root gravitropism and pollen-tube growth.
Almost every kind of ion transporter known to function inplants has been described in the guard cell (Figure 1c,d)[1–3]. In the plasma membrane, H+-ATPases pump H+ outof the cytosol. The plasma membrane is thus hyperpolarized,which drives ion influx by other transporters. Inward K+
channels and a presumed Cl– transporter are responsiblefor ion influx during stomatal opening. Sucrose uptake,likely occurring via a H+–sucrose co-transport mechanism,also appears to play a significant role in osmotic build-upunder some stomatal opening conditions [4–6]. Although theguard cell inward K+ channel, KAT1, was identified severalyears ago, sensitive reverse transcription polymerase chainreaction techniques have also shown expression of threeother inwardly rectifying K+ channels — AKT1, AKT2/3,and KAT2 — in guard cells. KAT1 may not play asdominant a role in K+ uptake as previously believed [7•].
Outward K+ channels [8] and anion channels are the mainconduits for passive ion efflux from the cytosol to theapoplast during stomatal closure. Evidence is accumulatingthat the guard cell S-type anion channel may consist of, orbe regulated by, an ATP-binding cassette (ABC)-type trans-porter [9•]. In the tonoplast, anion channels, fast-vacuolarchannels (FV), vacuolar K+-selective channels (VK), slow-vacuolar channels (SV), and vacuolar voltage-dependentCa2+ channels (VV) have been characterized (Figure 1) [2,3].
How so many ion transporters are organized in response tovarious stimuli over both the short term and the long termis a key question in stomatal physiology. This review willprovide an overview of some of the advances in this areaover the past 18 months. Other valuable recent reviewscovering various aspects of stomatal physiology are alsoavailable [1,10••–12••,13].
Stomata are modified by environmental factorsThe stomatal signaling system has been moulded byenvironmental factors over both long (millions of years)and short (hours, minutes, or even seconds) timescales.Considerable progress has been made in the elucidation ofenvironmental perception and ion transport mechanismsoccurring over the short timescale. Our understandingof what has taken place over the developmental andevolutionary timescales remains limited, but severalpapers have broken new ground. In 1987, Woodward [14]pointed to evidence in the fossil record that plants haveresponded to post-industrial increases in atmospheric CO2levels with decreases in stomatal density. This responsemay have been selected for because it allowed reducedwater loss, while photosynthesis was maintained due to theincreased availability of carbon dioxide. Conversely, anincrease in stomatal density can lead to increased waterloss, accompanied by increases in leaf cooling.
In 2001, it has been hypothesized that the evolution of thebroad lamina typical of most present-day leaves was madepossible by an increase in stomatal density that occurred
From milliseconds to millions of years: guard cells andenvironmental responsesSarah M Assmann* and Xi-Qing Wang†

following a drastic decrease in atmospheric CO2 levelsthat occurred in the late Paleozoic, 410–363 million yearsago. It is speculated that this increased stomatal densitypromoted evaporative cooling, which in turn kept broaderundissected leaves with their attendant thicker boundarylayers from overheating [15•,16].
Over a much shorter developmental timescale, some plantspecies also show an acclimatory response to atmosphericCO2, with a decrease in stomatal density observed underelevated CO2 conditions [17]. Recently, Gray et al. [18••]demonstrated the expression in guard cells of the geneHIC (HIGH CARBON DIOXIDE). In plants in which theHIC gene was disrupted by insertion of the β-glucuronidasegene, the stomatal index increased upon doubling ofexperimental CO2 levels. The hic mutants thus exhibited areversal of the normal inverse correlation between stomatalindex and ambient CO2 concentration. On the basis ofnucleotide and derived peptide sequence homologies, HICencodes a putative 3-keto acyl coenzyme A synthase.Enzymes of this type are involved in the synthesis of very-long-chain fatty acids. In plants, long-chain fatty acids arefound in waxes and cutins, which are constituents of thecuticle, as well as in glycerolipids and sphingolipids. Somesubstance that normally diffuses from pre-existing guard
cells under high CO2 concentrations, and prevents neigh-boring cells from differentiating into guard cells, may beabsent in the hic mutants or may be immobilized as a result ofalterations in the composition of the cuticular layer [18••,19].
Other wax mutants also lend support to the idea that disruptionof the extracellular matrix leads to a change in stomatalindex: eceriferum-1 and eceriferum-6 mutants of Arabidopsis[20], both of which have alterations in wax composition onthe leaf surface, exhibit higher stomatal indices than dowild-type plants.
The effect of the hic mutation differs from that of otherstomatal mutants such as too many mouths (tmm) [21] andstomatal density and distribution1 (sdd1) [22•], in that thestomata of the hic mutants show increased density but donot show abnormal clustering. TMM has not yet beencloned, but SDD1 encodes a subtilisin-like serine protease,suggesting that the processing of a proteinaceous signalmay be involved in stomatal patterning. It is interesting tospeculate that TMM, SDD1, or other gene productsimplicated in stomatal patterning may, like HIC, provesensitive to environmental factors that regulate stomatalmovements, such as light, drought, air pollutants, andhumidity. Some studies have already shown that long-termdrought stress and abscisic acid (ABA) treatment result inincreased stomatal density and reduced guard cell size([23] and references therein). The ABA effect on cell sizeindicates that not only stomatal patterning but also guardcell development is influenced by the environment. Normaldifferentiation of the stomatal complex involves cytoskeletalreorganization, and some studies have implicated a sensitivityof guard cell actin filament and microtubules arrays tosignals such as ABA and light [24,25].
The role of Ca2+ in guard cell responses toABA, cold, and oxidative stressGuard cells can sense and rapidly respond to many signals,including not only external stimuli but also internal stimulisuch as ABA and auxin [26,27]. This area of study isadvancing rapidly, and this past year has seen particularadvances in our understanding of the inter-relationshipsbetween ABA and Ca2+ signaling.
Changes in cytosolic calcium are elicited by many of thesignals to which plants respond, and cytosolic calcium hasbeen proposed to be a universal second messenger. How isthe specificity and reliability of Ca2+ signaling achieved?Perhaps a given external stimulus could produce a specifictype of Ca2+ response that amplifies the signal upstream ofCa2+, such as signals from receptors and G proteins, andthen signals to Ca2+-regulated downstream molecules, forexample, calcium-dependent protein kinases [28], inwardK+ channels and anion channels.
Along these lines, recent studies have utilized proteina-ceous Ca2+ reporters to measure Ca2+ oscillations in guardcells, with particular reference to ABA signaling. Allen et al.
422 Cell signalling and gene regulation
Figure 1
Ion transport in guard cells. (a) Stomatal complex of Vicia faba. Scalebar = 10 µm. (b) Stomatal complex of Arabidopsis thaliana. Scale bar= 10 µm. (c) Ion channels and transporters discussed in this articlethat reside at the guard cell plasma membrane. The Cl– transporterand the sucrose transporter are inferred from physiologicalexperiments but have yet to be characterized by molecular orelectrophysiological techniques. (d) Ion channels and transportersdiscussed in this article that reside at the guard cell tonoplast. FV,fast-vacuolar channel; SV, slow-vacuolar channel; VK, vacuolar K+-selective channel; VV, vacuolar voltage-dependent Ca2+ channel.
Current Opinion in Plant Biology
(c)
(d)
(a) (b)
H+
H+
H+-ATPase
Cl–
K+out
Anion (R-type)
(Stretch-activated)
Anion (S-type)
K+in
Ca2+in
Ca2+
Ca2+
Ca2+
AnionK+
K+
K+
K+
H+
Sucrose
V-ATPase
Anion
Anionchannel VK
SV
FVVV
[29••] transformed a calcium reporting green fluorescentprotein construct or ‘cameleon’ into Arabidopsis, andassessed guard cell Ca2+ responses to several distinct stim-uli. They found that ABA caused both repetitive cytosolictransients and more prolonged oscillations. Cold stimulat-ed small, repetitive cytosolic Ca2+ transients, whereasH2O2 induced one or two separate Ca2+ transients. Thus,each stimulus led to a Ca2+ response with distinct features.Even for a single signal, Ca2+ responses may vary in a man-ner that reflects signal strength. Staxen et al. [30] foundthat different ABA concentrations administered toCommelina communis epidermis led to guard cell Ca2+
oscillations of different periods, with higher concentrationsof the hormone eliciting lower frequency oscillations. Thisphenomenon may arise from differential contributions ofCa2+ uptake and internal Ca2+-release pathways at differentABA concentrations [12••].
How are Ca2+ oscillations produced?Ca2+ oscillations (periodic Ca2+ concentration changes overtime) and Ca2+ waves (spatial changes in calcium concentra-tion) are produced through the integration of Ca2+ influx,release of Ca2+ from the endomembrane system, Ca2+
efflux from the cytosol to the apoplast, and Ca2+ sequestrationinto intracellular stores. Different stimuli presumably usedifferent Ca2+ sources or releasing pathways to establishcharacteristic Ca2+ responses, and this hypothesis wasrecently lent credence by two studies. Wood et al. [31•]used aequorin to report Ca2+ responses to several stimuli inthe presence of Ca2+ channel blockers and the Ca2+ chelatorEGTA. They concluded that mechanical and ABA signalingin guard cells rely heavily on Ca2+ release from intracellularstores, whereas low-temperature signaling relies heavilyon Ca2+ uptake from the apoplast. Allen, Schroeder, andcolleagues [29••] studied Ca2+ oscillations in the Arabidopsisdet3 mutant. This mutant exhibits reduced expression ofthe C-subunit of the V-type ATPase, accompanied by
reduced endomembrane energization. They found thatthe det3 mutation changed the Ca2+ oscillations observedwhen stomata were challenged with external Ca2+ and H2O2,but they found no detectable changes in response to ABAand cold stimuli [29••].
The result with cold treatment is consistent with theresults of Wood et al. [31•]. However, it is perhaps unex-pected that the det3 mutation affected the H2O2 responsebut not the ABA response, as Schroeder’s group has alsoimplicated H2O2 as a secondary messenger for ABA action(see below) [32•]. The guard cell may utilize multipleredundant pathways, thus ensuring adequacy of the vitalABA response. In fact, this idea of redundancy is consistentwith the multiplicity of Ca2+ pathways implicated in guardcells, as described below.
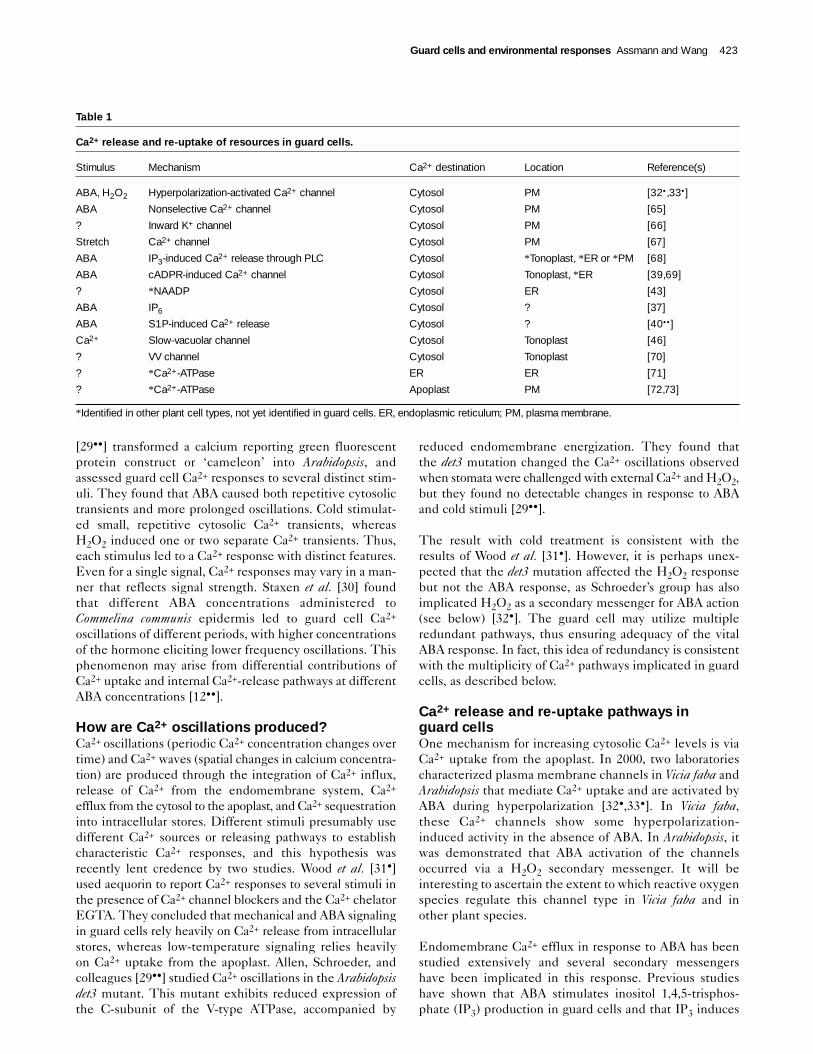
Ca2+ release and re-uptake pathways inguard cellsOne mechanism for increasing cytosolic Ca2+ levels is viaCa2+ uptake from the apoplast. In 2000, two laboratoriescharacterized plasma membrane channels in Vicia faba andArabidopsis that mediate Ca2+ uptake and are activated byABA during hyperpolarization [32•,33•]. In Vicia faba,these Ca2+ channels show some hyperpolarization-induced activity in the absence of ABA. In Arabidopsis, itwas demonstrated that ABA activation of the channelsoccurred via a H2O2 secondary messenger. It will beinteresting to ascertain the extent to which reactive oxygenspecies regulate this channel type in Vicia faba and inother plant species.
Endomembrane Ca2+ efflux in response to ABA has beenstudied extensively and several secondary messengershave been implicated in this response. Previous studieshave shown that ABA stimulates inositol 1,4,5-trisphos-phate (IP3) production in guard cells and that IP3 induces
Guard cells and environmental responses Assmann and Wang 423
Table 1
Ca2+ release and re-uptake of resources in guard cells.
Stimulus Mechanism Ca2+ destination Location Reference(s)
ABA, H2O2 Hyperpolarization-activated Ca2+ channel Cytosol PM [32• ,33•]
ABA Nonselective Ca2+ channel Cytosol PM [65]
? Inward K+ channel Cytosol PM [66]
Stretch Ca2+ channel Cytosol PM [67]
ABA IP3-induced Ca2+ release through PLC Cytosol *Tonoplast, *ER or *PM [68]
ABA cADPR-induced Ca2+ channel Cytosol Tonoplast, *ER [39,69]
? *NAADP Cytosol ER [43]
ABA IP6 Cytosol ? [37]
ABA S1P-induced Ca2+ release Cytosol ? [40•• ]
Ca2+ Slow-vacuolar channel Cytosol Tonoplast [46]
? VV channel Cytosol Tonoplast [70]
? *Ca2+-ATPase ER ER [71]
? *Ca2+-ATPase Apoplast PM [72,73]
*Identified in other plant cell types, not yet identified in guard cells. ER, endoplasmic reticulum; PM, plasma membrane.

cytosolic Ca2+ elevation and partial stomatal closure [34–36].IP3 may also serve as a precursor to IP6 (inositol hexakis-phosphate), which has also been linked to Ca2+-dependentABA responses [37]. Staxen et al. [30] characterized ABA-induced Ca2+ oscillations in detail and found that theseoscillations and stomatal closure were both partially inhibitedby U-73122 (1-[6-{(17β-3-methoxxyestra-1,3,5(10)-trien-17-yl)amino}hexyl]-1H-pyrrole-2,5-dione), an inhibitor ofphosphoinositide-specific phospholipase C (PLC).
PLC and another phospholipase, PLD, which cleaves thephospholipid headgroup to yield phosphatidic acid, bothshow transient increases in activity following ABA application.PLD seems to function in the same pathway as PLC:inhibitors of PLC and PLD each partially oppose ABA-induced stomatal closure, but simultaneous application ofboth inhibitors does not result in enhanced attenuation ofABA-induced closure [38•]. The relative positions of the twophospholipases in their signaling chain remains unknown,but it does seem that PLD activation is downstream of —or independent of — Ca2+ release because the directproduct of PLD action, phosphatidic acid, does not elicitcytosolic Ca2+ increases in guard cells.
The lack of complete reversal of ABA-induced stomatalclosure by combined PLC and PLD inhibitors implicatesother Ca2+ release pathways in the ABA response, and aparallel pathway appears to be that mediated by cyclicADP-ribose (cADPR). In contrast to the situation withPLC and PLD, simultaneous application of PLD andcADPR antagonists does increase the extent to whichABA-induced stomatal closure is reduced. For example,simultaneous application of nicotinamide, an inhibitor ofcADPR action, and 1-butanol, an inhibitor of phosphatidicacid production, results in an almost complete block ofABA-induced stomatal closure in Vicia faba epidermalpeels [38•]. Patch clamping of isolated guard cell vacuolesconfirms that cADPR activates Ca2+ release from thisorganelle via Ca2+-selective channels, and microjection ofcADPR into guard cells elicits cytosolic Ca2+ increases andstomatal closure [39].
One of the most recent calcium-mobilizing molecules tobe identified is sphingosine 1-phosphate (S1P). InCommelina communis, ABA-induced stomatal closure isopposed by a competitive inhibitor of sphingosine kinase,the enzyme responsible for producing S1P from sphingosine.Application of S1P to epidermal peels induces Ca2+
oscillations in guard cells and stomatal closure. An increasein S1P levels occurs in whole leaves following droughtstress, and it will be important to ascertain whether or notsuch responses also occur in guard cells [40••]. S1P isunusual in that it can serve as an intracellular signal or, iflocated extracellularly, interact with outwardly facingreceptors [41]. The idea of a mesophyll messenger thatcoordinates photosynthetic and stomatal functions hasintrigued stomatal physiologists for years [42]. Could S1Pplay such a role?
Yet another Ca2+ mobilization pathway has yet to beassayed in guard cells, although it has been detected inother types of plant cells. Navazio et al. [43] demonstratedthat endoplasmic reticulum vesicles from cauliflowerrespond to nicotinic acid adenine dinucleotide phosphate(NAADP), a Ca2+-releasing molecule recently described inboth vertebrates and invertebrates. In contrast to the IP3and cADPR pathways, NAADP did not elicit Ca2+ releasefrom tonoplast vesicles. The results with NAADP thushighlight the importance of the endoplasmic reticulum asa Ca2+ store in plants. Given the plethora of Ca2+ releasepathways described in guard cells to date, one wouldpredict that NAADP will also be found to mobilize Ca2+ inthis cell type.
Ca2+-independent pathwaysThe existence of multiple Ca2+ mobilization pathwaysand Ca2+ release sites undoubtedly contributes to thegeneration of stimulus-specific Ca2+ signals. It should bekept in mind, however, that Ca2+-independent pathwaysalso exist in the guard cell. ABA can inhibit K+ channelsinvolved in stomatal opening and in some circumstanceselicit stomatal closure without detectable calcium increasesin guard cells [31•,39,44]. Protons have been implicated asone of the other secondary messengers in the guard cellABA response. First, ABA also induces pH increases priorto stomatal closure; and second, elevated pH activates theplasma membrane outward K+ channel through which K+
efflux occurs during stomatal closure. This channel isCa2+-insensitive [45]. The activities of FV and SV channels,which release K+ and Ca2+ to the cytosol (Figure 1) [2] arealso stimulated by alkaline cytosolic pH, although the VKcurrent is inhibited under these conditions [3,46]. Thesedata imply that changes in pH function alongside changesin pCa to modulate stomatal movement.
Nonionic signaling in guard cell reponsesto stressThe guard cell not only uses Ca2+ oscillations and pHchanges to establish specific responses but also employsnonionic signaling molecules. Evidence that guard cellpathways have novel features not found universally in allplant cells has been provided recently by two studies.Sutton et al. [47] found that co-injection of oocytes withguard cell protoplast mRNA along with cRNA for KAT1resulted in the expression of an inward K+ channel that wasinhibited upon ABA application to the oocyte. However,oocytes co-injected with mesophyll mRNA and KAT1cRNA produced inward K+ currents that were not inhibitedby ABA [47]. These results show that the mesophyll ABAsignaling pathway is distinct from, and cannot substitutefor, the guard cell ABA signaling pathway.
One guard-cell-specific component may be AAPK (ABA-activated protein kinase), a serine-threonine proteinkinase whose biochemical activity has only been detectedin guard cells to date [48]. On the basis of sequencing oftwo peptides from AAPK using tandem mass spectrometry,
424 Cell signalling and gene regulation

Li et al. [49••] cloned the AAPK cDNA from Vicia fabaguard cells. Mutating Lys43 in AAPK to an alanineresidue resulted in a kinase with apparent dominant-negative effects, and guard cells transformed with AAPKK43A no longer closed in response to ABA application toepidermal peels. Guard cells expressing the mutantversion of the kinase also failed to activate plasma membraneanion channels, which play a central role in ABA-inducedstomatal closure.
Protein phosphorylation is reversed by protein phosphatases.abi1-1 and abi2-1 are two ABA-insensitive dominantmutant alleles of the protein phosphatase 2C (PP2C) genesABI1 and ABI2. Both of these mutations confer a wiltyphenotype and ABA insensitivity of stomatal closure.Recently, Gosti et al. [50••] isolated seven recessive allelesof ABI1 as intragenic revertants of the abi1-1 mutation.These revertants show hypersensitive responses to ABA interms of the inhibition of seed germination and seedlingroot growth. When water is withheld, the revertants alsoshow enhanced drought tolerance. The recessive nature ofthe revertants suggests that a loss of function in ABI1 isresponsible for the ABA-hypersensitive phenotype.
Consistent with the above hypothesis, recombinant proteinsderived from the intragenic revertant alleles do not exhibitdetectable PP2C activity. In the same assay, the PP2Cencoded by the dominant, possibly gain-of-function,abi1-1 allele exhibits residual phosphatase activity [50••].Taken together, the evidence suggests that wild-type ABI1phosphatase negatively regulates ABA responses, and it istempting to speculate that it reverses the action of theABA-activated kinase, AAPK.
Guard cells respond not only to drought stress but also tostresses such as salinity and air pollution [51]. In additionto their previously implicated roles in cell division andorgan development, polyamines have been found to accu-mulate upon plant exposure to such stresses [52]. Liu et al.thus addressed the hypothesis that polyamines areinvolved in stomatal action and, indeed, found that appli-cation of all of the natural polyamines inhibited stomatalopening and induced stomatal closure. Polyamines alsoinhibited whole-cell inward K+ currents, plausibly actingon the KAT1 inward K+ channel. This inhibition of inwardK+ channels was not observed in isolated membrane patches,suggesting that the response also requires cytoplasmicfactors [53]. These data suggest that polyamines may becomponents of stress-induced signal transduction in guardcells, and the next step will be to document the extent towhich guard cell polyamine concentrations change inresponse to stress signals.
Blue light and stomatal openingStomatal closure in response to ABA and stomatal openingin response to light comprise two major areas of investiga-tion by stomatal physiologists. Stomatal opening istriggered by both red light — acting through chlorophyll
— and blue light. Low-intensity blue light is much moreeffective in stimulating stomatal opening than comparablefluence rates of red light, pointing to a sensitive, non-chlorophyllous photoreceptor as the starting point for thisphotoresponse. Identification of this photoreceptor is stillunder investigation. Zeiger and colleagues have providedphotobiological evidence that zeaxanthin, a carotenoidintermediate in the xanthophyll cycle, is the specific blue-light photoreceptor [54]. Guard cells from the npq1 mutantof Arabidopsis, which lacks zeaxanthin, show reduced stomatalopening in response to blue light when assayed in isolatedepidermal peels [54]. Gas exchange experiments measuringblue-light-induced transpiration from whole leaves of thenpq1 mutant, however, show a wild-type stomatal responseto blue light. The same is true for all other blue-lightphotoreceptor mutants assayed to date (i.e. nph1, cry1, andcry2) [55,56].
Moreover, in Vicia faba at least, the ultraviolet action spectrumfor stomatal opening does not match the absorption spectrumof bulk (trans) zeaxanthin, and is instead consistent withthat of either a cis-carotenoid (including cis-zeaxanthin) ora flavin [57]. If zeaxanthin is indeed the guard cell blue-light photoreceptor, then the whole-leaf experiments withnpq1 imply that there must be a mesophyll blue-lightphotoreceptor that senses and reports the presence of bluelight to the zeaxanthin-less guard cells of npq1.
Although the early stages of the blue-light photoresponseremain mysterious, the ultimate targets of that signaltransduction chain are becoming clearer. Recently, twostudies [58••,59] have lent support to the idea that it is anH+-ATPase, rather than a redox pathway, that mediates theproton pumping that hyperpolarizes the plasma-membraneto drive K+ uptake and stomatal opening in response toblue light [60]. Shimazaki’s group used biochemicaltechniques to demonstrate that blue-light activation of theH+-ATPase involves blue-light-dependent phosphorylationof the carboxyl terminus of the H+-ATPase [58••]. Thisphosphorylation increases the affinity of the H+-ATPasefor a 14-3-3-protein (14-3-3 proteins are positive regulatorsof H+-ATPases). Three 14-3-3 cDNAs were identified in aVicia faba guard cell cDNA library, and analysis of thenative 14-3-3 protein co-immunoprecipitated with theH+-ATPase indicates that just one of the 14-3-3 isomers,14-3-3a, specifically associates with the H+-ATPase followingblue-light irradiation [61]. These elegant experiments havethus identified 14-3-3a as a key player in the stomatalresponse to blue light.
ConclusionsMassive progress has been made since the first successfulpatch clamp recordings from guard cells in 1984. A numberof important signaling elements have been identified thatfunction in guard cells on the timescale of milliseconds tominutes, although the ordering and interactions of thesesignaling molecules remain unknown in many cases. A fewkey genes have also been identified that play a role in
Guard cells and environmental responses Assmann and Wang 425

guard cell development. However, on the intermediatetimescale of the life-time of a single mature guard cell or asingle leaf, we still know very little about environmentaleffects on guard cell physiological function and geneexpression [62,63].
Many of the most telling experiments carried out overthe past few years have been conducted on isolated epi-dermal peels or guard cell protoplasts, or on whole leavesmonitored with gas-exchange techniques. It has beendemonstrated unequivocally, however, that in response tosignals such as ABA and atmospheric humidity, not all ofthe guard cells in a leaf act in unison. There exist domainsor ‘patches’ within the leaf that show differential responsesto an identical stimulus; furthermore, the boundaries andbehavior of such patches fluctuate over time. Such resultsindicate that stomata in the intact leaf do not functionindependently but rather are coupled, perhaps hydraulically,within a given patch. This ‘emergent collective behavior’[64••] adds one more layer of complexity to stomatal functionunder real-world conditions.
UpdateHeterotrimeric G proteins are one of the most importantcomponents of animal signaling pathways. Recent studieshave shown that the Arabidopsis G protein α subunit,GPA1, is also a vital regulator of plant growth and develop-ment [74••,75••], including guard cell signaling. Wang et al.[75••] used plants harbouring T-DNA insertions in GPA1 todemonstrate G protein involvement in ABA-inhibition ofstomatal opening and inward K+ channels, and in pH-inde-pendent ABA-activation of slow anion channels. gpa1mutant plants exhibit greater rates of water loss than theirwild-type counterparts [75••]. These results place G proteinsin the guard cell ABA signaling network.
Calcium is a universal signaling element. Allen et al. [76••]present elegant data showing that calcium oscillationnumbers frequency, and duration all regulate stomatal aperture[76••]. A detailed analysis is presented in Hetherington’sarticle on pp 415–420 of this issue.
AcknowledgementsResearch on guard cell signal transduction in the authors’ laboratory issupported by grants from the National Science Foundation (MCB 98-74438and MCB 00-86315) and the US Department of Agriculture (00-35100-9420and 01-35304-09916).
References and recommended readingPapers of particular interest, published within the annual period of review,have been highlighted as:
• of special interest••of outstanding interest
1. Assmann SM: Signal transduction in guard cells. Annu Rev CellBiol 1993, 9:345-375.
2. Ward JM, Pei ZM, Schroeder JI: Roles of ion channels in initiationof signal transduction in higher plants. Plant Cell 1995,7:833-844.
3. Allen GJ, Amtmann A, Sanders D: Calcium-dependent and calcium-independent K+ mobilization channels in Vicia faba guard cellvacuoles. J Exp Botany 1998, 49:305-318.
4. Zhao R, Dielen V, Kinet JM, Boutry M: Cosuppression of a plasmamembrane H+-ATPase isoform impairs sucrose translocation,stomatal opening, plant growth, and male fertility. Plant Cell 2000,12:535-546.
5. Ritte G, Rosenfeld J, Rohrig K, Raschke K: Rates of sugar uptake byguard cell protoplasts of Pisum sativum L. related to the soluterequirement for stomatal opening. Plant Physiol 1999, 121:647-656.
6. Talbott LD, Zeiger E: The role of sucrose in guard cellosmoregulation. J Exp Botany 1998, 49:329-337.
7. Szyroki A, Ivashikina N, Dietrich P, Roelfsema MRG, Ache P, • Reintanz B, Deeken R, Godde M, Felle H, Steinmeyer R et al.: KAT1 is
not essential for stomatal opening. Proc Natl Acad Sci USA 2001,98:2917-2921.
Genetic disruption of the inward K+ channel KAT1 has no effect on stomatalbehavior in the assays reported in this paper. Other inward K+ channels arealso expressed in the guard cell and may compensate for the loss offunctional KAT1.
8. Ache P, Becker D, Ivashikina N, Dietrich P, Roelfsema MRG,Hedrich R: GORK, a delayed outward rectifier expressed in guardcells of Arabidopsis thaliana, is a K+-selective, K+-sensing ionchannel. FEBS Lett 2000, 486:93-98.
9. Gaedeke N, Klein M, Kolukisaoglu U, Forestier C, Müller A, • Ansorge M, Becker D, Mamnun Y, Kuchler K, Schulz B et al.: The
Arabidopsis thaliana ABC transporter AtMRP5 controls rootdevelopment and stomata movement. EMBO J 2001,20:1875-1887.
ABC (ATP-binding cassette) transporters constitute a large protein family inArabidopsis. This paper shows that the ABC transporter, AtMRP5, isexpressed in guard cells and is involved in stomatal opening.
10. Schroeder JI, Kwak JM, Allen GJ: Guard cell abscisic acid signalling •• and engineering drought hardiness in plants. Nature 2001,
410:327-330.This excellent review updates current knowledge of guard cell abscisic acid(ABA) responses, summarizing most of the components that have beenshown to be involved in this signaling pathway.
11. Blatt MR: Cellular signaling and volume control in stomatal •• movements in plants. Annu Rev Cell Dev Biol 2000, 16:221-241.This review describes some of the recent developments in stomatal physiology,with particular emphasis on membrane trafficking.
12. MacRobbie EAC: ABA activates multiple Ca2+ fluxes in stomatal •• guard cells, triggering vacuolar K+ (Rb+) release. Proc Natl Acad
Sci USA 2000, 97:12361-12368.This article provides a comprehensive overview of Ca2+ influx pathways acrossthe plasma membrane and Ca2+-release pathways from endomembranes.
13. McAinsh MR, Brownlee C, Hetherington AM: Calcium ions assecond messengers in guard cell signal transduction. PhysiolPlant 1997, 100:16-29.
14. Woodward FI: Stomatal numbers are sensitive to CO2 increasesfrom pre-industrial levels. Nature 1987, 327:617-618.
15. Beerling DJ, Osborne CP, Chaloner WG: Evolution of leaf-form in • land plants linked to atmospheric CO2 decline in the late
Palaeozoic era. Nature 2001, 410:352-354.Atmospheric CO2 concentration influences on stomatal density and evaporativecooling may have been key elements in the evolution of the broad leaf laminafound in many present day land plants.
16. McElwain JC, Beerling DJ, Woodward FI: Fossil plants and globalwarming at the Triassic–Jurassic boundary. Science 1999,285:1386-1390.
17. Woodward FI, Kelly CK: The influence of CO2 concentration onstomatal density. New Phytol 1995, 131:311-327.
18. Gray JE, Holroyd GH, van der Lee FM, Bahrami AR, Sijmons PC, •• Woodward FI, Schuch W, Hetherington AM: The HIC signalling
pathway links CO2 perception to stomatal development. Nature2000, 408:713-716.
The HIC gene identified in this report may be crucial for stomatal responseto global climate change. hic mutants show an unusual increase in stomatalindex under elevated CO2 concentrations. HIC is hypothesized to encodean enzyme involved in the synthesis of the leaf cuticle.
19. Serna L, Fenoll C: Plant biology. Coping with human CO2emissions. Nature 2000, 408:656-657.
20. Jenks MA, Tuttle HA, Eigenbrode SD, Feldmann KA: Leaf epicuticularwaxes of the eceriferum mutants in Arabidopsis. Plant Physiol1995, 108:369-377.
426 Cell signalling and gene regulation

21. Geisler M, Nadeau J, Sack FD: Oriented asymmetric divisions thatgenerate the stomatal spacing pattern in Arabidopsis aredisrupted by the too many mouths mutation. Plant Cell 2000,12:2075-2086.
22. Berger D, Altmann T: A subtilisin-like serine protease involved in • the regulation of stomatal density and distribution in Arabidopsis
thaliana. Genes Dev 2000, 14:1119-1131.Understanding of stomatal development at the molecular level remainslimited. The authors of this paper used genetic techniques to identify a role for asubtilisin-like serine protease in the regulation of stomatal density and distribution.
23. Franks PJ, Farquhar GD: The effect of exogenous abscisic acid onstomatal development, stomatal mechanics, and leaf gasexchange in Tradescantia virginiana. Plant Physiol 2001,125:935-942.
24. Marcus AI, Moore RC, Cyr RJ: The role of microtubules in guardcell function. Plant Physiol 2001, 125:387-395.
25. Hwang JU, Lee Y: Abscisic acid-induced actin reorganization inguard cells of dayflower is mediated by cytosolic calcium levelsand by protein kinase and protein phosphatase activities. PlantPhysiol 2001, 125:2120-2128.
26. Merritt F, Kemper A, Tallman G: Inhibitors of ethylene synthesisinhibit auxin-induced stomatal opening in epidermis detachedfrom leaves of Vicia faba L. Plant Cell Physiol 2001, 42:223-230.
27. Bauly JM, Sealy IM, Macdonald H, Brearley J, Dröge S, Hillmer S,Robinson DG, Venis MA, Blatt MR, Lazarus CM, Napier RM:Overexpression of auxin-binding protein enhances the sensitivityof guard cells to auxin. Plant Physiol 2000, 124:1229-1238.
28. Berkowitz G, Zhang X, Mercie R, Leng Q, Lawton M: Co-expressionof calcium-dependent protein kinase with the inward rectifiedguard cell K+ channel KAT1 alters current parameters in Xenopuslaevis oocytes. Plant Cell Physiol 2000, 41:785-790.
29. Allen GJ, Chu SP, Schumacher K, Shimazaki CT, Vafeados D, •• Kemper A, Hawke SD, Tallman G, Tsien RY, Harper JF et al.:
Alteration of stimulus-specific guard cell calcium oscillations andstomatal closing in Arabidopsis det3 mutant. Science 2000,289:2338-2342.
This article provides evidence that different stimuli elicit distinct Ca2+ oscil-lations, and that different stimuli use different Ca2+ resources in the productionof those oscillations. det3 is an Arabidopsis mutant with a 60% reduction inthe V-type H+-ATPase. det3 exhibits altered Ca2+ oscillations in response toCa2+ and H2O2, but ABA and cold-treatment-induced Ca2+ oscillationsretain the wild-type phenotype.
30. Staxen I, Pical C, Montgomery LT, Gray JE, Hetherington AM,McAinsh MR: Abscisic acid induces oscillations in guard-cellcytosolic free calcium that involve phosphoinositide-specificphospholipase C. Proc Natl Acad Sci USA 1999, 96:1779-1784.
31. Wood NT, Allan AC, Haley A, Viry-Moussaïd M, Trewavas AJ: The • characterization of differential calcium signalling in tobacco guard
cells. Plant J 2000, 24:335-344.The authors of this paper identify differential use of Ca2+ resources in guardcell responses to various environmental stimuli.
32. Pei ZM, Murata Y, Benning G, Thomine S, Klüsener B, Allen GJ, • Grill E, Schroeder JI: Calcium channels activated by hydrogen
peroxide mediate abscisic acid signalling in guard cells. Nature2000, 406:731-734.
This report characterizes a guard cell Ca2+ channel from the model plantspecies, Arabidopsis, and presents evidence that reactive oxygen speciesmediate ABA signaling in guard cells.
33. Hamilton DWA, Hills A, Köhler B, Blatt MR: Ca2+ channels at the • plasma membrane of stomatal guard cells are activated by
hyperpolarization and abscisic acid. Proc Natl Acad Sci USA 2000,97:4967-4972.
A plasma membrane Ca2+ channel in Vicia faba guard cells that is activatedby hyperpolarization and ABA is described.
34. Lee YS, Choi YB, Suh S, Lee J, Assmann SM, Joe CO, Kelleher JF,Crain RC: Abscisic acid-induced phosphoinositide turnover inguard cell protoplasts of Vicia faba. Plant Physiol 1996,110:987-996.
35. Gilroy S, Read ND, Trewavas AJ: Elevation of cytoplasmic calciumby caged calcium or caged inositol trisphosphate initiatesstomatal closure. Nature 1990, 346:769-771.
36. Blatt MR, Thiel G, Trentham DR: Reversible inactivation of K+
channels of Vicia stomatal guard cells following the photolysis ofcaged inositol 1,4,5-trisphosphate. Nature 1990, 346:766-769.
37. Lemtiri-Chlieh F, MacRobbie EAC, Brearley CA: Inositolhexakisphosphate is a physiological signal regulating the K+-inward rectifying conductance in guard cells. Proc Natl Acad SciUSA 2000, 97:8687-8692.
38. Jacob T, Ritchie S, Assmann SM, Gilroy S: Abscisic acid signal • transduction in guard cells is mediated by phospholipase D
activity. Proc Natl Acad Sci USA 1999, 96:12192-12197.Phospholipase D (PLD) activity following ABA application to guard cells wasassessed by direct biochemical assays and use of pharmacologicalinhibitors. PLD appears to function in a pathway that is distinct from thecADPR pathway to mediate stomatal responses to ABA.
39. Leckie CP, McAinsh MR, Allen GJ, Sanders D, Hetherington AM:Abscisic acid-induced stomatal closure mediated by cyclic ADP-ribose. Proc Natl Acad Sci USA 1998, 95:15837-15842.
40. Ng CKY, Carr K, MacAinsh MR, Powell B, Hetherington AM: Drought-•• induced guard cell signal transduction involves sphingosine-1-
phosphate. Nature 2001, 410:596-599.Sphingosine-1-phosphate appears to be a novel calcium-mobilizing agent inguard cells. Sphingosine-1-phosphate could induce Ca2+ oscillations andstomatal closure. Disruption of sphingosine-1-phosphate production led tothe partial inhibition of ABA-induced stomatal closure.
41. Young KW, Nahorski SR: Intracellular sphingosine 1-phosphateproduction: a novel pathway for Ca2+ release. Semin Cell Dev Biol2001, 12:19-25.
42. Wong SC, Cowan IR, Farquhar GD: Stomatal conductancecorrelates with photosynthetic capacity. Nature 1979,282:424-426.
43. Navazio L, Bewell MA, Siddiqua A, Dickinson GD, Galione A,Sanders D: Calcium release from the endoplasmic reticulum ofhigher plants elicited by the NADP metabolite nicotinic acidadenine dinucleotide phosphate. Proc Natl Acad Sci USA 2000,97:8693-8698.
44. Romano LA, Jacob T, Gilroy S, Assmann SM: Increases in cytosolicCa2+ are not required for abscisic acid-inhibition of inward K+
currents in guard cells of Vicia faba L. Planta 2000, 211:209-217.
45. Blatt MR, Armstrong F: K+ channels of stomatal guard cells:abscisic-acid-evoked control of the outward rectifer mediated bycytoplasmic pH. Planta 1993, 191:330-341.
46. Ward JM, Schroeder JI: Calcium-activated K+ channels andcalcium-induced calcium release by slow vacuolar ion channels inguard cell vacuoles implicated in the control of stomatal closure.Plant Cell 1994, 6:669-683.
47. Sutton F, Paul SS, Wang XQ, Assmann SM: Distinct abscisic acidsignaling pathways for modulation of guard cell versusmesophyll cell potassium channels revealed by expressionstudies in Xenopus laevis oocytes. Plant Physiol 2000,124:223-230.
48. Li J, Assmann SM: An abscisic acid-activated and calcium-independent protein kinase from guard cells of fava bean. PlantCell 1996, 8:2359-2368.
49. Li J, Wang XQ, Watson MB, Assmann SM: Regulation of abscisic •• acid-induced stomatal closure and anion channels by guard cell
AAPK kinase. Science 2000, 287:300-303.The authors of this paper report cloning of a guard-cell-specific proteinkinase, AAPK. In guard cells expressing a dominant loss-of-function AAPKallele, ABA failed to activate anion channels or induce stomatal closure.
50. Gosti F, Beaudoin N, Serizet C, Webb AAR, Vartanian N, Giraudat J: •• ABI1 protein phosphatase 2C is a negative regulator of abscisic
acid signaling. Plant Cell 1999, 11:1897-1909.The abi1-1 mutant is a wilty, ABA-resistant Arabidopsis mutant that hasbeen heavily used in stomatal physiology. The authors of this paper describethe whole-plant phenotype and phosphatase activity of intragenic revertantsof abi1-1. The results indicate that the wild-type ABI1 phosphatase is anegative regulator of ABA responses.
51. Torsethaugen G, Pell EJ, Assmann SM: Ozone inhibits guard cell K+
channels implicated in stomatal opening. Proc Natl Acad Sci USA1999, 96:13577-13582.
52. Evans PT, Malmberg RL: Do polyamines have roles in plantdevelopment? Annu Rev Plant Physiol Plant Mol Biol 1989,40:235-269.
53. Liu K, Fu H, Bei Q, Luan S: Inward potassium channel in guardcells as a target for polyamine regulation of stomatal movements.Plant Physiol 2000, 124:1315-1326.
Guard cells and environmental responses Assmann and Wang 427

54. Zeiger E, Zhu J: Role of zeaxanthin in blue light photoreceptionand the modulation of light-CO2 interactions in guard cells. J ExpBotany 1998, 49:433-442.
55. Eckert M, Kaldenhoff R: Light-induced stomatal movement of selectedArabidopsis thaliana mutants. J Exp Botany 2000, 51:1435-1442.
56. Lasceve G, Leymarie J, Olney MA, Liscum E, Christie JM,Vavasseur A, Briggs WR: Arabidopsis contains at least fourindependent blue-light-activated signal transduction pathways.Plant Physiol 1999, 120:605-614.
57. Eisinger W, Swartz TE, Bogomolni RA, Taiz L: The ultraviolet actionspectrum for stomatal opening in broad bean. Plant Physiol 2000,122:99-106.
58. Kinoshita T, Shimazaki KI: Blue light activates the plasma •• membrane H+-ATPase by phosphorylation of the C-terminus in
stomatal guard cells. EMBO J 1999, 18:5548-5558. The H+-ATPase is a key component in guard cell signaling. This paperelegantly shows that blue light can stimulate phosphorylation of the carboxy-terminus of the H+-ATPase, and induce H+-ATPase activity. 14-3-3 proteinbinding to the H+-ATPase is promoted by blue-light illumination.
59. Taylor AR, Assmann SM: Apparent absence of a redox requirementfor blue light activation of pump current in broad bean guard cells.Plant Physiol 2001, 125:329-338.
60. Assmann SM, Simoncini L, Schroeder JI: Blue light activateselectrogenic ion pumping in guard cell protoplasts of Vicia faba.Nature 1985, 318:285-287.
61. Emi T, Kinoshita T, Shimazaki KI: Specific binding of vf14-3-3aisoform to the plasma membrane H+-ATPase in response to bluelight and fusicoccin in guard cells of broad bean. Plant Physiol2001, 125:1115-1125.
62. Plesch G, Kamann E, Mueller-Roeber B: Cloning of regulatorysequences mediating guard-cell-specific gene expression. Gene2000, 249:83-89.
63. Smart LB, Cameron KD, Bennett AB: Isolation of genespredominantly expressed in guard cells and epidermal cells ofNicotiana glauca. Plant Mol Biol 2000, 42:857-869.
64. Mott KA, Buckley TN: Patchy stomatal conductance: emergent •• collective behaviour of stomata. Trends Plant Sci 2000, 5:258-262.This insightful review summarizes knowledge and hypotheses concerningpatchy stomatal behavior in the intact leaf.
65. Schroeder JI, Hagiwara S: Repetitive increases in cytosolic Ca2+ ofguard cells by abscisic acid activation of nonselective Ca2+
permeable channels. Proc Natl Acad Sci USA 1990, 87:9305-9309.
66. Fairley-Grenot KA, Assmann SM: Permeation of Ca2+ through K+
channels in the plasma membrane of Vicia faba guard cells.J Membr Biol 1992, 128:103-113.
67. Cosgrove DJ, Hedrich R: Stretch-activated chloride, potassium, andcalcium channels coexisting in plasma membranes of guard cellsof Vicia faba L. Planta 1991, 186:143-153.
68. Muir SR, Sanders D: Inositol 1,4,5-trisphosphate-sensitive Ca2+
release across nonvacuolar membranes in cauliflower. PlantPhysiol 1997, 114:1511-1521.
69. Navazio L, Mariani P, Sanders D: Mobilization of Ca2+ by cyclicADP-ribose from the endoplasmic reticulum of cauliflower florets.Plant Physiol 2001, 125:2129-2138.
70. Allen GJ, Sanders D: Two voltage-gated, calcium release channelscoreside in the vacuolar membrane of broad bean guard cells.Plant Cell 1994, 6:685-694.
71. Hwang I, Sze H, Harper JF: A calcium-dependent protein kinasecan inhibit a calmodulin-stimulated Ca2+ pump (ACA2) located inthe endoplasmic reticulum of Arabidopsis. Proc Natl Acad SciUSA 2000, 97:6224-6229.
72. Bonza MC, Morandini P, Luoni L, Geisler M, Palmgren MG,De Michelis MI: At-ACA8 encodes a plasma membrane-localized calcium-ATPase of Arabidopsis with acalmodulin-binding domain at the N-terminus. Plant Physiol 2000,123:1495-1506.
73. Chung WS, Lee SH, Kim JC, Heo WD, Kim MC, Park CY, Park HC,Lim CO, Kim WB, Harper JF, Cho MJ: Identification of acalmodulin-regulated soybean Ca2+-ATPase (SCA1) that islocated in the plasma membrane. Plant Cell 2000, 12:1393-1407.
74. Ullah H, Chen JG, Young JC, Im KH, Sussman MR, Jones AM:•• Modulation of cell proliferation by heterotrimeric G protein in
Arabidopsis. Science 2001, 292:2066-2069.See ‘Update’.
75. Wang XQ, Ullah H, Jones AM, Assmann SM: G protein regulation of•• ion channels and abscisic acid signaling in Arabidopsis guard
cells. Science 2001, 292:2070-2072.See ‘Update’.
76. Allen GJ, Chu SP, Harrington CL, Schumacher K, Hoffmann T, •• Tang YY, Grill E, Schroeder JI: A defined range of guard cell
calcium oscillation parameters encodes stomatal movements.Nature 2001, 411:1052-1057.
See ‘Update’.
428 Cell signalling and gene regulation





























