Embed Size (px)
Citation preview

ECOLOGICALMODELLING
ELSEVIER Ecological Modelling 149 (2002) 41-51
www.elsevier.comflocatefecolmodel
Functional branch analysis as tool for fractal scaling above-and belowground trees for their additive and non-additive
properties
Meine
van Noordwijk *, Rachmat MuliaICRAF South East Asia Regional Research Program, PO Box 161, Bogor 16001, Indonesia
Abstract
Empirical allometric scaling equations (Y = aDb) for tree biomass on the basis of stem diameter D are often usedin forest inventories and for assessment of carbon and nutrient stocks in vegetation. When shifting from plantationforestry to mixed forestry or multi-species agroforestry systems, however, short-cuts to the empirical approach forestablishing such equations are desirable. Fractal branchin& models provide a transparent scheme for derivingtree-specific scaling rules (especially, the b parameter) on the basis of easily observable, non-destructive methods. Therelation between link (section of stem or branch between two branching points) length and link diameter has a directinfluence on the b parameter of the allometric equation in the range 2-3.5, providing substantial variation around theclaims of a universal value of 8/3. Apart from the total tree biomass, the models can provide rules for total leaf area;relative allocation of current growth to leaves, branches or stem. The power of the allometric scaling relation (b)necessarily has the same value, for a given tree, for all properties that are dominated by the endpoints of thebranching process, and that are thus 'additive'. Below ground, similar descriptions hold for individual root axes,where the proximal root diameter can be used for predicting total length or biomass of all its branches. Samplingerror was analyzed to derive rules for the number of branching points that should be observed for reliable estimatesof the fractal branching parameters. For the inherent parameter variability that was chosen as default setting aminimum number of 50 branching points should be observed. A spreadsheet model (functional branch analysis, FBA)is made available through the WWW that allows users to derive results for new parameter combinations and/or seeknew applications. @ 2002 Elsevier Science B. V. All rights reserved.
Keywords: Allometrics; Fractal dimension; Root biomass; Tree architecture; Tree biomass
1.
Introduction
Trees cover a broader range of scale in theirsize than any other organism (Thomas, 2000) andthe concept 'tree' needs a scalar before any mean-ingful statements about biomass, nutrient stocksor uptake, water use, carbon sequestration, pro-
* Corresponding author. Tel.: + 62-251-315-234; fax: + 62-251-315-567.
E-mail address: [email protected] (M. van No-ordwijk).
0304-3800/02/$ -see front matter @ 2002 Elsevier Science B.Y. All rights reserved.PII: S0304-3800(01)00513-0

42 M. van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51
ductivity or any other functional ecological prop-erty can be made. Foresters have long term expe-rience in scaling trees on the basis of stemdiameter at a specified height above the ground(Brown, 1997), but their empirical rules need re-calibration for each tree species and stand density,and can not be easily applied in mixed speciesforests or in situations where trees grow in moreopen conditions rather than in a closed stand(Brown et al., 1995; Ketterings et al., 2001). Whencomparisons include several orders of magnitude,simple scaling rules may give the impression thatall trees (West et al., 1999) or even all forests(Enquist and Niklas, 2001) of the world are be-having in the same way and can be captured by asingle allometric scaling rule. If the target is ananswer of the right order of magnitude, we mayindeed use a generic scaling rule with a power of8/3 or 2.67. If we care about errors of a factor 3,however, a method that predicts the power forscaling relations for each tree species (or evenindividual tree...) is relevant. Thus, the need re-mains for achieving a better understanding of theempirical scaling rules for trees and, ideally, ascheme that allows simple observations of treeshape to be translated into reasonable estimates ofthe scaling rules.
Trees can not have any form-there are clearconstraints in 'designing' trees, as all parts mustbe connected to ensure transport, it must meetminimum mechanical stability and it should beable to grow, i.e. all steps between its currentform and a start (usually from a seed) must havebeen possible as well. Beyond these minimumrequirements, there are considerations of resourcecapture and resource use efficiency, which proba-bly lead to differences in 'fitness' of differentshapes (Niklas, 1994).
We can explore the apparent logic of real worldtrees and use non-destructively observable proper-ties to reconstruct allometric equations-this iswhat we do in 'functional branch analysis' orFBA.
Aboveground trees, root systems, rivers androad networks share common properties, whichhave gradually been recognized on the basis of'fractal' analysis (Mandelbrot, 1983), althoughsome of the concepts date back to Leonardo da
Vinci. Fractal properties can emerge if a relativelysimple set of rules is applied consistently across arange of scales, e.g. stating that the diametersafter a branching point are a certain proportionof the diameter before branching. If such rules areapplied repeatedly, a form can be constructedwith remarkable similarity to a tree or river sys-tem. Different reasons may be given for such arule, for example a constant total transport re-quirement before and after the branching point,or the need for mechanical stability which re-quires each branch to be designed to carry itsshare of the total weight load (Niklas, 1992).
Fractal or self-repeating branching models areessentially based on assumptions of 'self-similar-ity' across scales: this means that a picture of abranching point looks the same, whether it istaken from the first or last branching point of atree, relative to the diameter of the branches. Thissuggests a simple test: measure the diameter ofbranches before and after a branching point andtest in regression analysis whether or not thederived parameters depend on current diameter.
The objectives of our research were to developa procedure for deriving allometric equations forbiomass and other properties of trees on the basisof parameters that can be measured non-destruc-tively, and in future may be derived by visualestimation on individual trees in mixed stands. Indeveloping this procedure we need to know,which parameters have major impact on the finalresult, and we need guidelines for the number ofreplicate observations needed to derive the inputparameters for the model, for the type of variabil-ity to be expected in real trees.
The questions considered in this paper are:.can we design a consistent and transparent
system for describing trees in terms of branch-ing properties and relate this to total sizeproperties,
.can allometric equations for relating total treesize to initial stem diameter be predicted fromproperties of the branching system;
.which properties of the branching system havea major impact on the allometric equations andwhich a minor one (sensitivity analysis);
.how many observations will be required inpractical applications to reduce uncertainty toan acceptable level?

M. van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51
.can we relate the fractal dimension of various L(D) = Lm( l + r(~ )'
properties of a tree (e.g. leaf area, biomass) to DUlineach other, for a given branching pattern? Canwe identify scaling rules for different propertiesof the same set of trees from characteristics ofthese properties?
.can we relate a property such as 'specific rootlength' to the diameter of proximal roots?
43
(2)
For fractal (scale-independent) models to apply,the parameters p and q should be independent ofcurrent diameter D. Subsequent work on tree rootsystems, as well as, aboveground trees (Van No-ordwijk and Purnomosidhi, 1995) has shown thatthis assumption of independence may be used asfirst approximation for a substantial range ofdiameters, but at small diameters the average ~may increase, although the scatter in calculatedvalues from observations is often large. If a(weak) dependence of p or q on diameter is found,the branching pattern can still be reconstructed byrepeated applications of the same set of rules, butthe program needs some modification.
2. Methods
2.1. Basic fractal branching rules
2.2.
Rules for end-structures
The basic model can be extended by providingrules for 'end structures' such as leaves or fineroots (of a diameter less than DmiJ, as a functionof current diameter. In the simplest form we needtwo parameters:
lend describes the number of endstructures perunit link length, andDendstr, max indicates the maximum diameter thatstill carries endstructures.
2.3.
Fractal branching model
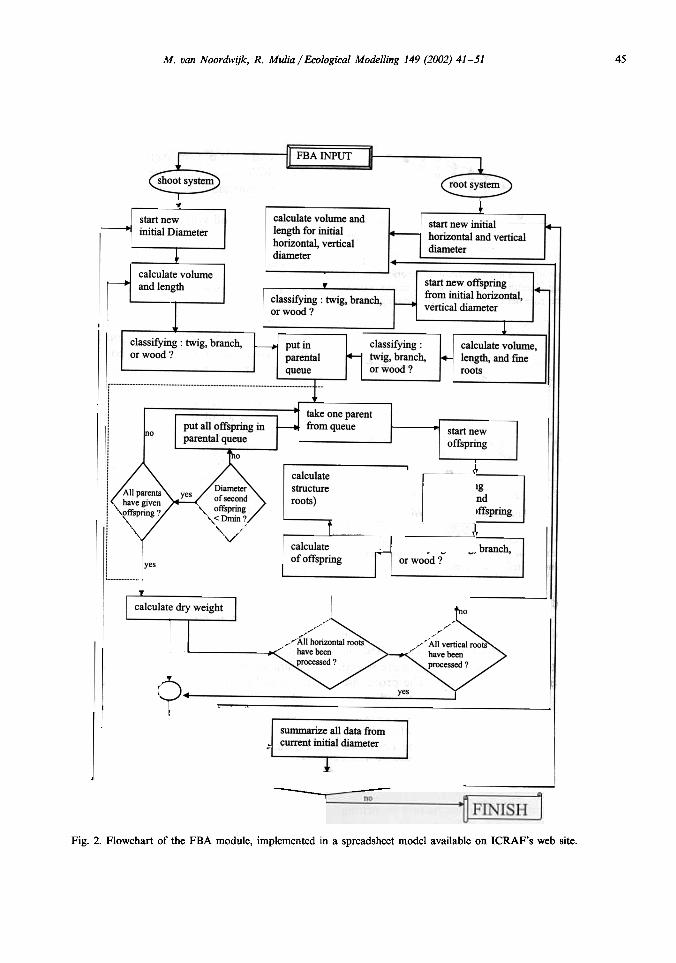
With these basic rules, a founding link of di-ameter Do can be split into links that can be putin a 'parental queue' and be subsequently split, aslong as more than 1 of their offspring has adiameter exceeding the minimum diameter Dmin'In the flowchart (Fig. 2), a central concept is thatof a 'parental queue' if links that have yet to be'split' to derive their offspring. Without a mini-mum diameter for links, this process would beinfinite, as each branching event adds more linksto the parental queue than it removed. But with aminimum diameter (DmiJ rule, splits in rootswith:
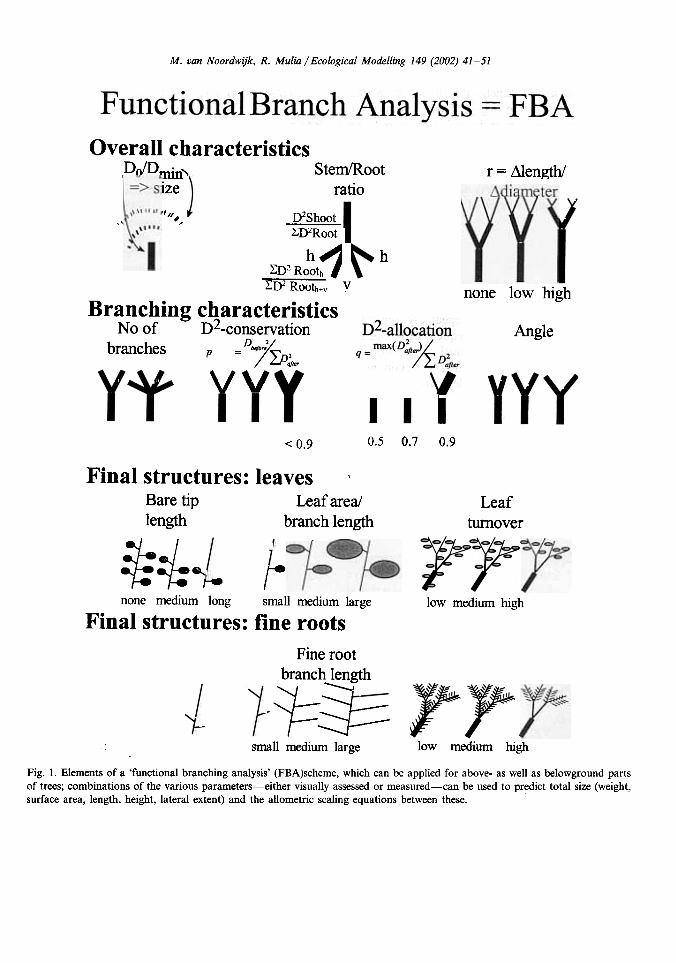
Fractal branching models repeatedly apply thesame equations to derive subsequent orders of thebranching process. For practical applications, arule is added for stopping when a certain mini-mum size is reached. The rules can refer to thediameter, length and/or orientation of the nextorder of branches. Van Noordwijk et al. (1994)and Spek and Van Noordwijk (1994) applied rulesthat are primarily based on the diameter to simu-late (tree) root systems. Five parameters (n, p, q,Lm and r) are needed in this approach (Fig. 1):
n = number of branches into which the currentlink (section of stem or branch between twobranching points) splits at the followingbranching point; n ;;::: 2p = D~/(~JDii I,), describes the change indiameter from link order i to its n derivativelinks of order i + 1 and hence cross-sectionalarea (cssa) of the stem ( this parameter wasearlier referred to as cx);,q=Di~l, 1/(Di~l, I+Di~l, 2), for n=2 andDl+ I, I> Di+ I, 2 (0,5 < q ~ 1) to describe therelative equity among the branches. With thesedefinitions we obtain for n = 2):
Di+ 1 1 = Di ~, ~p
Hl-q)Di+I,2=Di (1)
p
Lm is the length of a link of minimum diameter,and r is the relative increment in link length perunit relative increment in diameter, hence:
M. van Noordwijk, R. Mulia/Ecological Modelling 149 (2002) 41.-51
Overall characteristicsPoIDrnin"l=> size
Stem/Rootratio
r = 111ength/
1"'" f'" I'"" " '"
, ,, ~, ,
02Shoot","2.;D2R~LD2 Ro~t~ I ~
2.;IY ROOtltf'V Y
Branching characteristicsNo of D2-conservation
D ;J5branches p = bef... ~~2LJ.I after
VV'
h
low high
noneAngle
y0.5 0.7
I
0.9< 0.9
Leafturnover
<D,.I
Final structures: leavesBare tip Leaf areallength branch length:f::i:~ I. -~
none medium long small medium large
Final structures: fine rootslow medium high
Fine
rootturnover
Bare tip
length-ft,
none medium long
Fine rootbranch length
~§::::small medium large low medium high
Fig. 1. Elements of a 'functional branching analysis' (FBA)scheme, which can be applied for above- as well as belowground partsof trees; combinations of the various parameters-either visually assessed or measured-can be used to predict total size (weight,surface area, length, height, lateral extent) and the allometric scaling equations between these.
M. van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51 45
calculate volume andlength for initialhorizontal, verticaldiameter
start newinitial Diameter
start new initialhorizontal and verticaldiameter
calculate volumeand length start new offspring
from initial horizontal,vertical diameterclassifying: twig, branch,
or wood?
classifying: twig, branch,or wood?
classifying:twig, branch,or wood?
calculate volume,length, and [meroots
put inparentalQueue
take one parentfrom queueput all offspring in
parental queuestart newoffspring
shoot I
root ~~have been have been
yes
100 (root)
.:
~::~~--",---,-o (shoot) ,iterntioncomplete? yes
. yes (Shoot).c::=:::::~§~~~~~~:=::::::: ~ yes (root) yes (shoot) Start new initial dia~ter? {
no .
FINISH.
Fig. 2. Flowchart of the FBA module, implemented in a spreadsheet model available on ICRAF's web site.
46 M. van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51
100000000
10000000
1000000
100000
10000
1000
100
of properties of the tree (or root system) as awhole can be related to Do, for example by fittingan allometric equation of the type Y = aDb to thedata. In a spreadsheet model (available viawww.icraf.cgiar.orgjseajAgroModelsjWaNuICASjindex.htm), the relations between theinput parameters n, p, q, Lm and r and the outputparameters a and b can be explored, for length,surface area and volume (or weight) of the tree.
3.
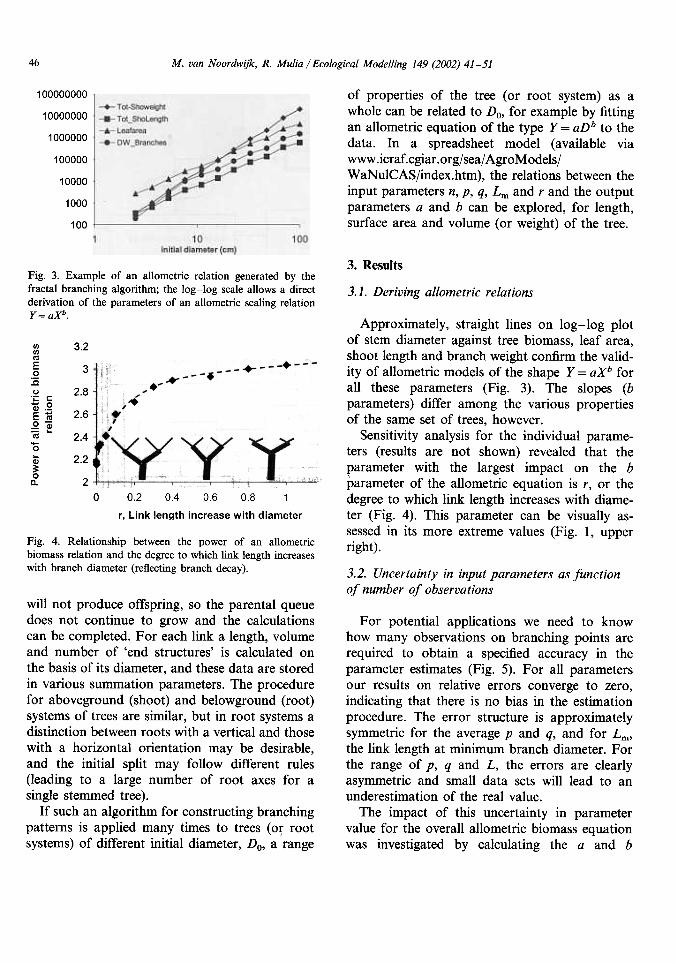
ResultsFig. 3. Example of an allometric relation generated by thefractal branching algorithm; the log-log scale allows a directderivation of the parameters of an allometric scaling relationY = aXb.
3.1.
Deriving allometric relations
3.2
c:,2m~
3
2.8
2.6
2.4
2.2
2
Approximately, straight lines on log-log plotof stem diameter against tree biomass, leaf area,shoot length and branch weight confirm the valid-ity of allometric models of the shape Y = aXb forall these parameters (Fig. 3). The slopes (bparameters) differ among the various propertiesof the same set of trees, however.
Sensitivity analysis for the individual parame-ters (results are not shown) revealed that theparameter with the largest impact on the bparameter of the allometric equation is r, or thedegree
to which link length increases with diame-ter (Fig. 4). This parameter can bc visually as-sessed in its more extreme values (Fig. I, upperright).
InInco
E0:cu"':WE..Qiij-0
a;~0D.
0
0.2 0.4 0.6 0.8 1
r, Link len~th increase with diameter
Fig.
4. Relationship between the power of an allometricbiomass relation and the degree to which link length increases
with branch diameter (reflecting branch decay).
3.2.
Uncertainty in input parameters as functionof number of observations
will not produce offspring, so the parental queuedoes not continue to grow and the calculationscan be completed. For each link a length, volumeand number of 'end structures' is calculated onthe basis of its diameter, and these data are storedin various summation parameters. The procedurefor aboveground (shoot) and belowground (root)systems of trees are similar, but in root systems adistinction between roots with a vertical and thosewith a horizontal orientation may be desirable,and the initial split may follow different rules(leading to a large number of root axes for asingle stemmed tree).
If such an algorithm for constructing branchingpatterns is applied many times to trees (o~ root
systems) of different initial diameter, Do, a range
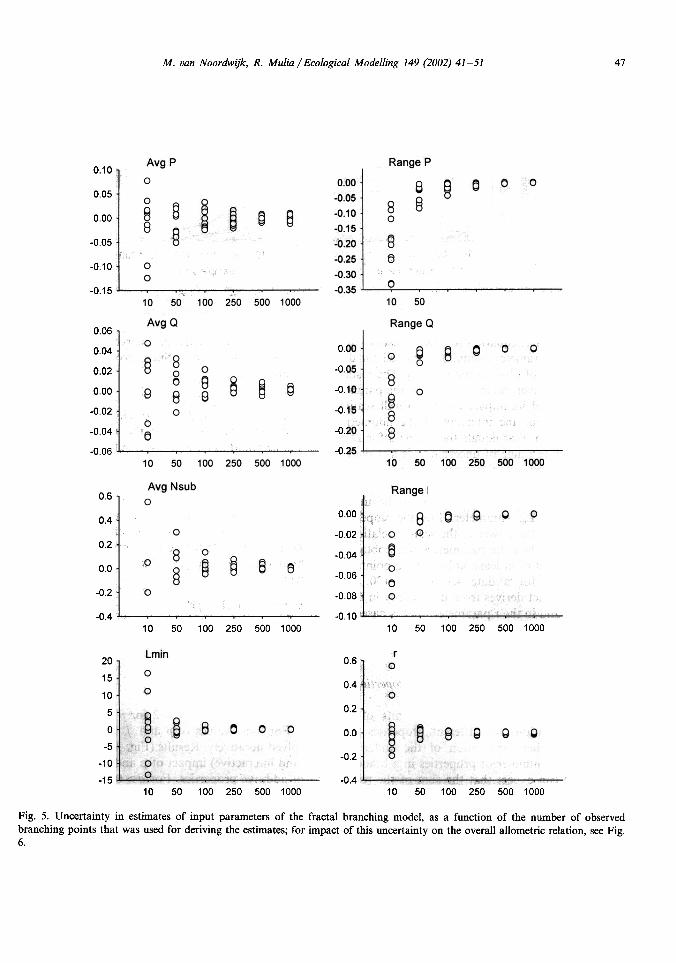
For potential applications we need to knowhow many observations on branching points arerequired
to obtain a specified accuracy in theparameter estimates (Fig. 5). For all parametersour results on relative errors converge to zero,indicating that there is no bias in the estimationprocedure.
The error structure is approximatelysymmetric for the average p and q, and for Lm,the
link length at minimum branch diameter. Forthe range of p, q and L, the errors are clearlyasymmetric
and small data sets will lead to anunderestimation of the real value.
The impact of this uncertainty in parametervalue for the overall allometric biomass equationwas
investigated by calculating the a and b
M. van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51 47
0.10
0,05
0,00
-0.05
-0.10
-0.15
100 250 500 1000
o.
o.
o.
o.
-0.
-0.
-0.
0.1
-0.1
-0.
-0.
-0.:
-0.:
0.6
0.00
-0.02
-0.04
-006
-008
-0.10
0.4
0.2
0.0
-0.2
-0.4
10 50 100 250 500 1000 10 50 100 250 500 1000
20
15
10
5
0
-5
-10
-15
0.6
0.4
0.2
0.0
-0.2
-0.4
10 50 100 250 500 1000 10 50 100 250 500 1000
Fig. 5. Uncertainty in estimates of input parameters of the fractal branching model, as a function of the number of observedbranching points that was used for deriving the estimates; for impact of this uncertainty on the overall allometric relation, see Fig.6.
06
04
02
00
02
04
06
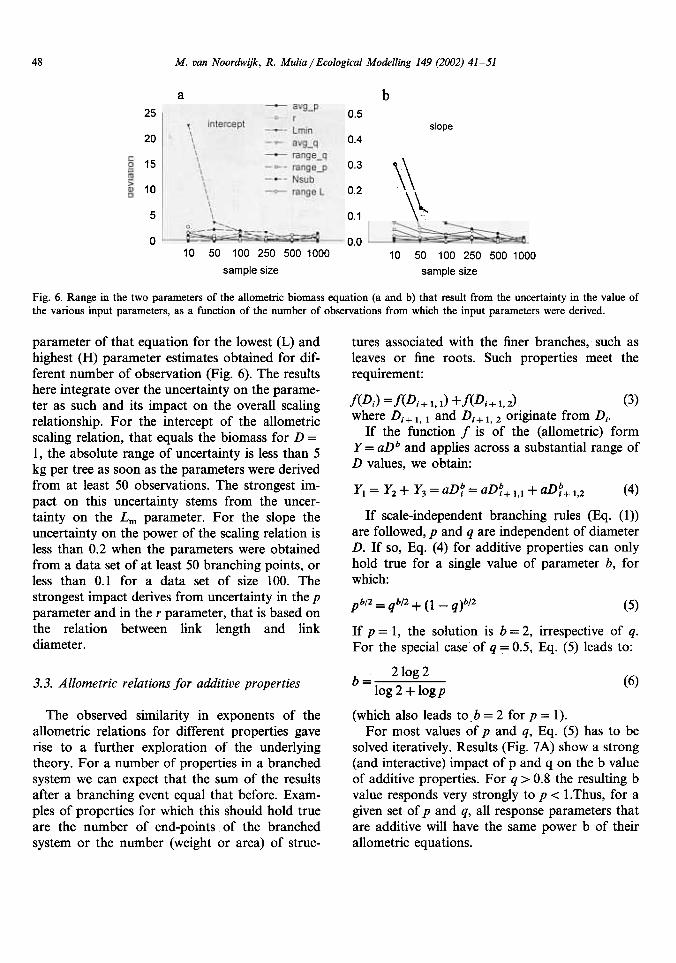
48 M. van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51
ba25 0.5
slope20 0.4
15 0.3
\\10 0.2
\\
5
0.1
0 0.010 50 100 250 5001000
sample size10 50 100 250 5001000
sample size
Fig.
6. Range in the two parameters of the allometric biomass equation (a and b) that result from the uncertainty in the value ofthe various input parameters, as a function of the number of observations from which the input parameters were derived.
parameter of that equation for the lowest (L) andhighest (H) parameter estimates obtained for dif-ferent number of observation (Fig. 6). The resultshere
integrate over the uncertainty on the parame-ter as such and its impact on the overall scalingrelationship.
For the intercept of the allometricscaling relation, that equals the biomass for D =1, the absolute range of uncertainty is less than 5kg
per tree as soon as the parameters were derivedfrom at least 50 observations. The strongest im-pact on this uncertainty stems from the uncer-tainty on the Lm parameter. For the slope theuncertainty on the power of the scaling relation isless
than 0.2 when the parameters were obtainedfrom a data set of at least 50 branching points, orless
than 0.1 for a data set of size 100. Thestrongest impact derives from uncertainty in the pparameter and in the r parameter, that is based onthe relation between link length and linkdiameter.
tures associated with the finer branches, such asleaves or fine roots. Such properties meet therequirement:
f(DJ = f(Di+I, J + f(Di+I,2) (3)where Di+ 1,1 and Di+ 1,2 originate from Di'
If the function f is of the (allometric) formY = aDb and applies across a substantial range ofD values, we obtain:
Yi= Y2+ Y3=aDf=aDf+i,i+aDf+i,2 (4)
If scale-independent branching rules (Eq. (I))are followed, p and q are independent of diameterD. If so, Eq. (4) for additive properties can onlyhold true for a single value of parameter b, forwhich:
pbl2 = qbl2 + (1 -q)bl2 (5)
If p = 1, the solution is b = 2, irrespective of q.For the special case of q = 0.5, Eq. (5) leads to:
2 log 2log 2 + logp
b= (6)
3.3.
Allometric relations for additive properties
The observed similarity in exponents of theallometric relations for different properties gaverise to a further exploration of the underlyingtheory. For a number of properties in a branchedsystem we can expect that the sum of the resultsafter a branching event equal that before. Exam-ples
of properties for which this should hold trueare the number of end-points of the branchedsystem or the number (weight or area) of struc-
(which also leads to.b = 2 for p = 1).For most values of p and q, Eq. (5) has to be
solved iteratively. Results (Fig. 7A) show a strong(and interactive) impact of p and q on the b valueof additive properties. For q > 0.8 the resulting bvalue responds very strongly to p < 1. Thus, for agiven set of p and q, all response parameters thatare additive will have the same power b of theirallometric equations.
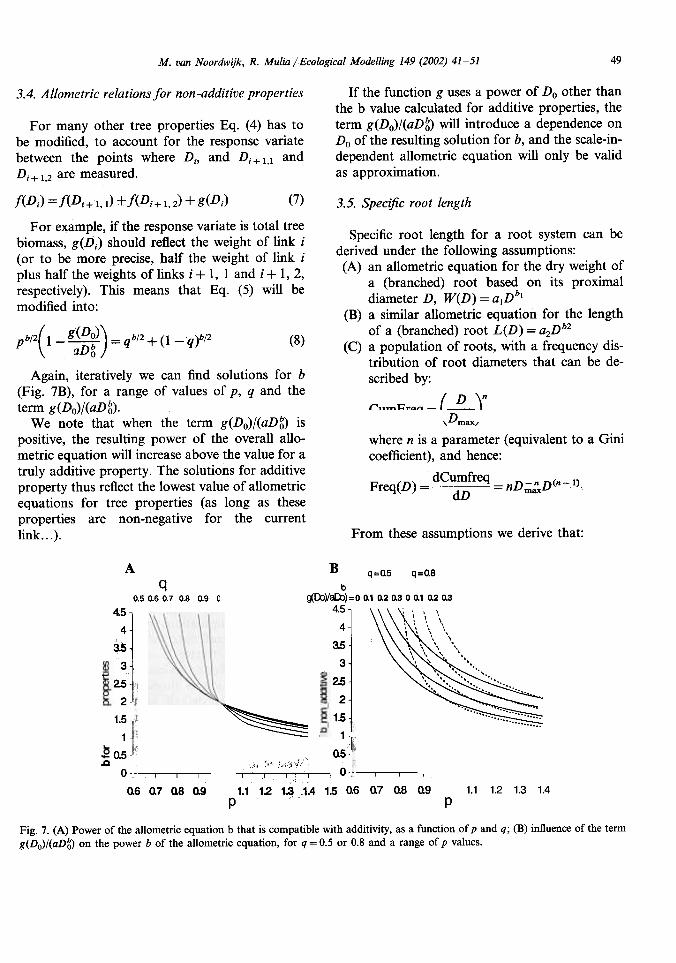
49M. van Noordwijk, R. Mulia jEcological Modelling 149 (2002) 41-51
If the function g uses a power of Do other thanthe b value calculated for additive properties, theterm g(Do)/(aDg) will introduce a dependence onDo of the resulting solution for b, and the scale-in-dependent allometric equation will only be validas approximation.
3.4.
Allometric relations for non-additive properties
3.5.
Specific root length
For many other tree properties Eq. (4) has tobe modified, to account for the response variatebetween the points where D;, and D;+ 1,1 andD;+ 1,2 are measured.f(DJ
= f(D;+I, J+ f(D;+ 1,2) +g(DJ (7)
For e~ample, if the response variate is total treebiomass, g(DJ should reflect the weight of link i(or to be more precise, half the weight of link iplus half the weights of links i + I, 1 and i + 1, 2,respectively). This means that Eq. (5) will bemodified into:
pb/2(
1-~)= qb/2 + (1 _q)b/2 (8)
Specific root length for a root system can bederived under the following assumptions:
(A) an allometric equation for the dry weight ofa (branched) root based on its proximaldiameter D, W(D) = a[Dbl
(B) a similar allometric equation for the lengthof a (branched) root L(D) = a2Db2
(C) a population of roots, with a frequency dis-tribution of root diameters that can be de-scribed by:
aD8
CumFreq = ( -£- )n
Again, iteratively we can find solutions for b(Fig. 7B), for a range of values of p, q and theterm g(Do)/(aDg).
We note that when the term g(Do)/(aDg) ispositive, the resulting power of the overall allo-metric equation will increase above the value for atruly additive property. The solutions for additiveproperty thus reflect the lowest value of allometricequations for tree properties (as long as theseproperties are non-negative for the currentlink...).
,Dmax,where n is a parameter (equivalent to a Ginicoefficient), and hence:
dCumfreqFreq(D) = = nD;;;:XD(n-l)-..dD
From these assumptions we derive that:
A Bq=Q5 q=o.s
q0.5 0.6 0.7 0.8 0.9 0.95
b\t()))/a:b)=O 0.1 0.20.300.1 0.20.3
4.5 '.\
4 ...'\
3.5 ",3 "'.
.-2.5 2
~~~- 1.5 ".'-..;:::::::::::~ 1.J
0.5~C" ;...' fA.'.!'!";
I I ",. I 0 I "~c
1.1 12 ~ 1A 1.5 0.6 0.7 0.8 0.9
4.:\~5
3
2.5-
2
1.5 ]
1
0.5.a 0] I I I
0.6 0.7 Qa Q.9
1.1
1.2 1.3 1.4pp
Fig.
7. (A) Power of the allometric equation b that is compatible with additivity, as a function of p and q; (B) influence of the termg(Do)/(aDS) on the power b of the allometric equation, for q = 0.5 or 0.8 and a range of p values.

50
M.
van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51
i Dmax w=
t 0
FreqWdD
(11
and
SumD2 = n {w~~} 2/bl (13)
r (2 + n) t (aln)
Similarly, from assumptions Band C weobtain:
(14)
and
the relation between link length and link diame-ter. Fractal dimensions of various properties of atree (e.g. leaf area, biomass) appear to followspecific patterns, depending on the degree towhich the property in question follows rules for'additivity'. The fractal branching procedure canalso be used to estimate specific root length as afunction of tree stem diameter. Other applicationsare in the estimation of 'green' versus 'brown'light interception in savannah trees and the impli-cations this has for modifying temperature with orwithout competition for water with other parts ofthe vegetation (Van Noordwijk and Ong, 1999).Applicability of these equations depend onwhether or not the assumptions underlying thefractal (scale-independent) process are met in thereal world. A procedure for testing this on indi-vidual trees is available (Van Noordwijk andPurnomosidhi, 1995; Van Noordwijk et al., 1996).The basic assumptions underlying fractal branch-ing have been tested and found to be applicable asacceptable first approximation for a wide range oftropical trees, above, as well as belowground,although a number of extensions of the theoryhave been suggested for various situations (Pageset al., 2000; Ozier-Lafontaine et al., 1999; Rowe,1999; Smith, 2001).
If the basic assumptions do inde~d apply, un-certainty on parameter values becomes the nextissue. Results presented here suggest that a mini-mum of 50 but preferably 100 branching pointsshould be used for deriving the main parametersof the fractal branching process; from the graphsand table presented here one can derive morespecific recommendations once the required preci-sion in the overall outcome is specified.
Overall, the fractal branching analysis offerspromise for a better understanding of why, whenand how to modify allometric scaling relationsfrom the generic 'default' values recommended inthe forestry literature. Although the 8/3 rule forallometric scaling of trees has a biomechanicalinterpretation that may appear to be generic(West et al., 1999), our results suggest that scalingrules in the range 2-3.5 are consistent with shapesas can be observed in real trees (Fig. 4).
We are currently testing the degree to which theparameters of the fractal branching process arecharacteristic of tree species, and thus can be
4.
Discussion
The results of our analysis show that a consis-tent and transparent system can indeed be de-signed for describing trees in terms of branchingproperties and relate this to total size propertiesby allometric equations. The parameter of thebranching system with the single largest effect onoverall allometrics for biomass, appeared to be
L = -~ { 2(n + 2) }b2/2
t (b2+n) SumDr-;;-

51M. van Noordwijk, R. Mulia / Ecological Modelling 149 (2002) 41-51
meaningfully entered in databases, or whetherthey essentially depend on site and managementconditions and thus require location specific tun-ing. The fact that the major parameters, at least intheir more extreme values, can be visually recog-nized should lead to more confidence in the use ofthe method. The allometric scaling relations asderived with the FBA module can be directly usedin the WaNuLCAS model of tree-soil-crop in-teractions (Van Noordwijk and Lusiana, 1999,2000).
Acknowledgements
Thanks are due to Dr Francis Putz and ananonymous reviewer for comments on an earlierversion of this manuscript.
References
Ozier-Lafontaine, H., Lecompte, F., Sinon, J.F., 1999. Fractalanalysis of the root architecture of Gliricidia sepium for thespatial prediction of root branching, size and mass: modeldevelopment and evaluation in agroforestry. Plant Soil209, 167-180.
Pages, L., Asseng, S., Pellerin, S., Diggle, A., 2000. Modellingroot system growth and architecture. In: Smit, A.L.,Bengough, A.G., Engels, C., van Noordwijk, M., Pellerin,S., van de Geijn, S.C. (Eds.), Root Methods, A Handbook.Springer, Berlin, pp. 113-146.
Rowe, E.C., 1999. The safety-net role of tree roots inhedgerow intercropping systems. PhD. Thesis, Wye Col-lege, University of London, UK, pp. 288
Spek, L.Y., Van Noordwijk, M., 1994. Proximal root diame-ters as predictors of total root system size for fractalbranching models. II. Numerical model. Plant and Soil
164,119-128.Smith, D.M., 2001. Estimation of tree root lengths using
fractal branching rules: a comparison with soil coring forGrevillea robusta. Plant Soil 229, 295-304.
Thomas, P., 2000. Trees: Their Natural History. CambridgeUniversity Press, Cambridge, UK, p. 286.
Van Noordwijk, M., Purnomosidhi, P., 1995. Root architec-ture in relation to tree-soil-crop interactions and shootpruning in agroforestry. Agroforestry Systems 30, 161-173.
Van Noordwijk, M., Lusiana, B., 1999. WaNuLCAS, a modelof water, nutrient and light capture in agroforestry sys-tems. Agroforestry Systems 43, 217-242.
Van Noordwijk, M., Ong, C.K., 1999. Can the ecosystemmimic hypotheses be applied to farms in African savan-nahs? Agroforestry Systems 45, 131-158.
Van Noordwijk, M. and Lusiana, B. 2000, WaNuLCAS 2.0,Background on a model of water nutrient and light capturein agroforestry systems. International Centre for Researchin Agroforestry (ICRAF), Bogor, Indonesia, pp. 186.
Van Noordwijk, M., Spek, L.Y., De Willigen, P., 1994. Proxi-mal root diameters as predictors of total root system sizefor fractal branching models. I. Theory. Plant and Soil 164,107-118.
Van Noordwijk, M., Lawson, G., Groot, J.J.R., Hairiah, K.,1996. Root distribution in relation to nutrients and compe-tition. In: Ong, C.K., Huxley, P.A. (Eds.), Tree-CropInteractions-a Physiological Approach. CAB Interna-tional, Wallingford, pp. 319-364.
West, G.B., Brown, J.H., Enquist, B.J., 1999. A generalmethod for the structure and'allometry of plant vascularsystems. Nature 400, 664-667.
Brown, S., 1997. Estimating biomass and biomass change oftropical forests, a primer. FAD Forestry paper 134, FAD,Rome.
Brown, I.F., Martinelli, L.A., Thomas, W.W., Moreira, M.Z.,Ferreira, C.A.C., Victoria, R.A., 1995. Uncertainty in thebiomass of Amazonian forests: an example from Rondo-nia, Brazil. Forest Ecology and Management 75, 175-189.
Enquist, B.J., Niklas, K.J., 2001. Invariant scaling relationsacross tree-doininated communities. Nature 410, 655-660.
Ketterings, Q.M., Coe, R., Van Noordwijk, M., Ambagau',Y., Palm, C.A., 2001. Reducing uncertainty in the use ofallometric biomass equations for predicting above-groundtree biomass in mixed secondary forest. Forest Ecologyand Management 146, 201-211.
Mandelbrot, B.B., 1983. The Fractal Geometry of Nature.Freeman, New York.
Niklas, K.J., 1992. Plant Biomechanics: an Engineering Ap-proach to Plant Form and Function. University ofChicago Press, Chicago, London, p. 607.
Niklas, K.J., 1994. Plant Allometry, the Scaling of Form andProcess. University of Chicago Press, Chicago, London, p.365.





























