Embed Size (px)
Citation preview

GLUCONEOGENESIS
Formation of glucose from noncarbohydrate sources
1

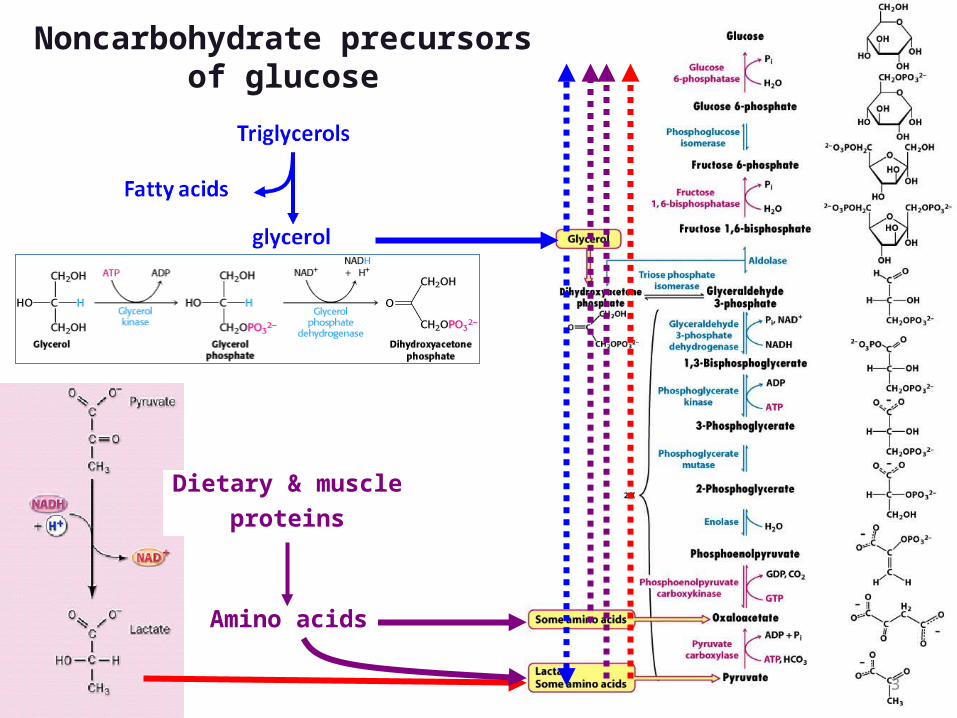
The source of pyruvate and oxaloacetate for gluconeogenesis during fasting or carbohydrate starvation is mainly amino acid catabolism.
Some amino acids are catabolized to pyruvate, oxaloacetate, or precursors of these.
Muscle proteins may break down to supply amino acids. These are transported to liver where they are deaminated and converted to gluconeogenesis inputs.
Glycerol, derived from hydrolysis of triacylglycerols in fat cells, is also a significant input to gluconeogenesis.
2
Dietary & muscleproteins
Amino acids
Noncarbohydrate precursors of glucose
3

Main sites of gluconeogenesis:
• Major site: Liver.• Minor site: Kidney.• Very little:
– Brain.– Muscle (skeletal and heart).
In liver and kidney it helps to maintain the glucose level in the blood so that brain and muscle can extract sufficient glucose from it to meet their metabolic demands.
4

Gluconeogenesis Versus Glycolysis:
• 7 steps are shared between glycolysis and gluconeogenesis.
• 3 essentially irreversible steps shift the equilibrium far on the side of glycolysis.
• Most of the decrease in free energy (consuming energy) in glycolysis takes place during these 3 steps.
5

6

In gluconeogenesis the three reactions are bypassed by a set of separate enzymes.
1. Phosphoenolpyruvate is formed from pyruvate:
2. Fructose 6-phosphate is formed from fructose 1,6-bisphosphate:
3. Glucose is formed by hydrolysis of glucose 6-phosphate:
7

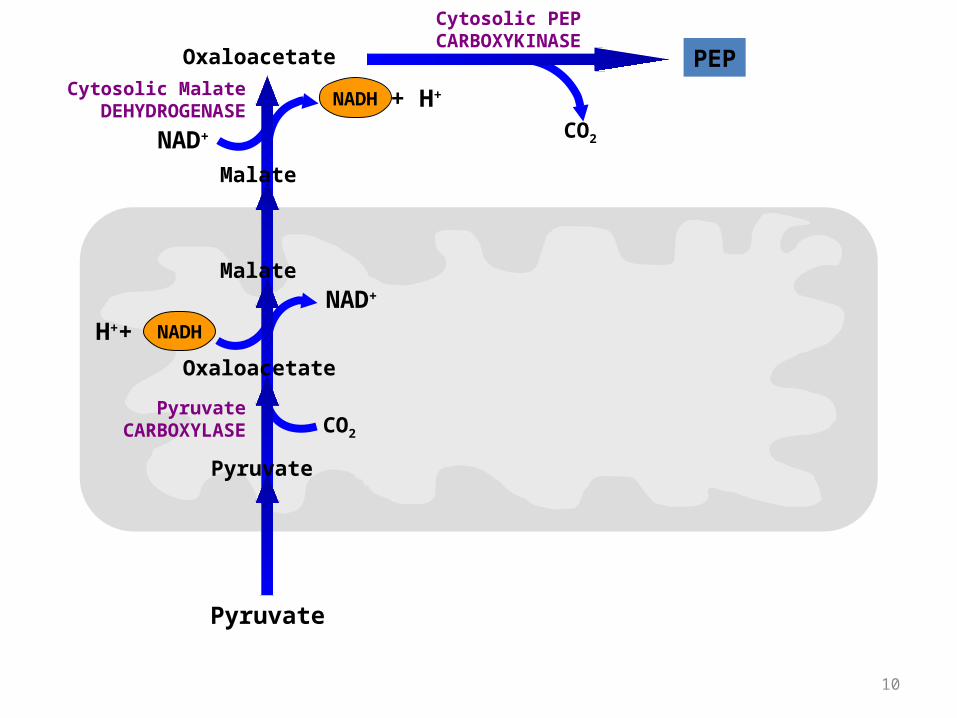
Bypass of Pyruvate Kinase:
Pyruvate Kinase (last step of Glycolysis) catalyzes: phosphoenolpyruvate + ADP pyruvate + ATP
For bypass of the Pyruvate Kinase reaction, cleavage of 2 ~P bonds is required.
G for cleavage of one ~P bond of ATP is insufficient to drive synthesis of phosphoenolpyruvate (PEP).
PEP has a higher negative G of phosphate hydrolysis than ATP.
8

PYRUVATE PHOSPHOENOLPYRUVATE
A
B
9
2 Enzymes needed to bypass pyruvate kinase
Pyruvate
Pyruvate
Oxaloacetate
Malate
Malate
Oxaloacetate PEP
CO2
NADHH++NAD+
NADH + H+
NAD+ CO2
Pyruvate CARBOXYLASE
Cytosolic PEP CARBOXYKINASE
Cytosolic Malate DEHYDROGENASE
10
NADH + H+
NAD+
Lactate
Pyruvate
Oxaloacetate
PEP
Pyruvate
Pyruvate
Oxaloacetate
Malate
Malate
Oxaloacetate PEP
CO2
CO2
CO2
NADHH++NAD+
NADH + H+
NAD+ CO2
Pyruvate CARBOXYLASE
Pyruvate CARBOXYLASE
Mitochondrial PEP CARBOXYKINASE
Cytosolic PEP CARBOXYKINASE
Lactate DEHYDROGENASE
Cytosolic Malate DEHYDROGENASE
Pyruvate
11

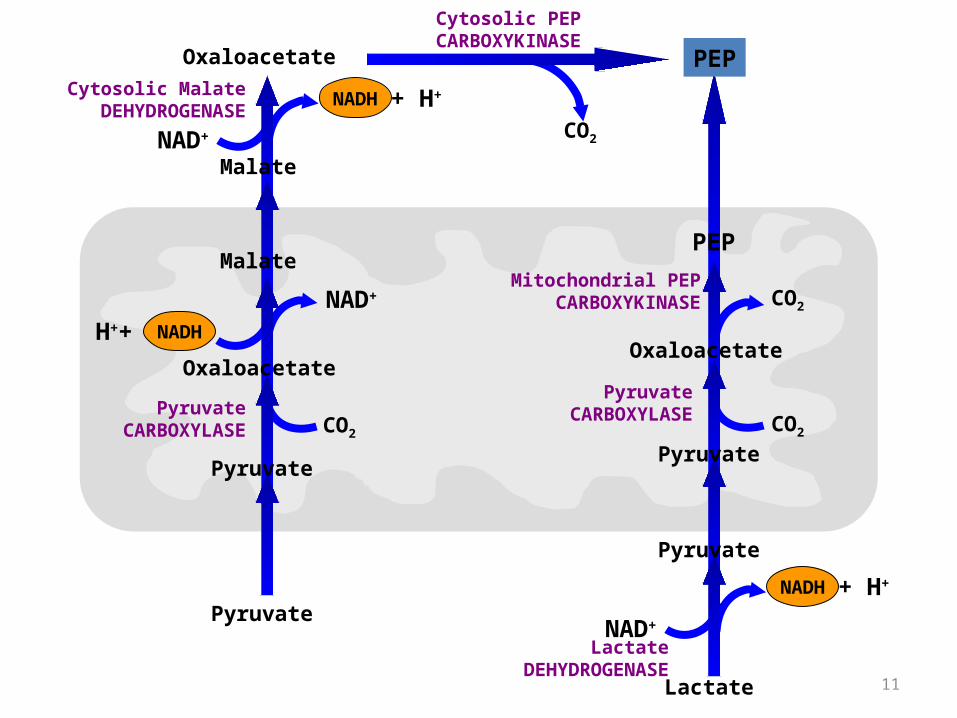
• This pathway predominates when lactate is the precursor.
• The conversion of lactate to pyruvate in the hepatocyte cytosol yields NADH.
• Thus no Malate transport is needed any more for this purpose.
• The mitochondrial and cytosolic PEP CARBOXYKINASE enzymes are encoded by separate nuclear genes. (two different enzymes catalyzing the same reaction in different localizations)
12

• To keep glucose inside the cell, the generation of free glucose is controlled in two ways:1.The enzyme responsible for the conversion of
glucose 6-phosphate into glucose, glucose 6-phosphatase, is regulated.
2.The enzyme is present only in tissues whose metabolic duty is to maintain blood-glucose balanced by releasing glucose into the blood (the liver and to a lesser extent the kidney).
13
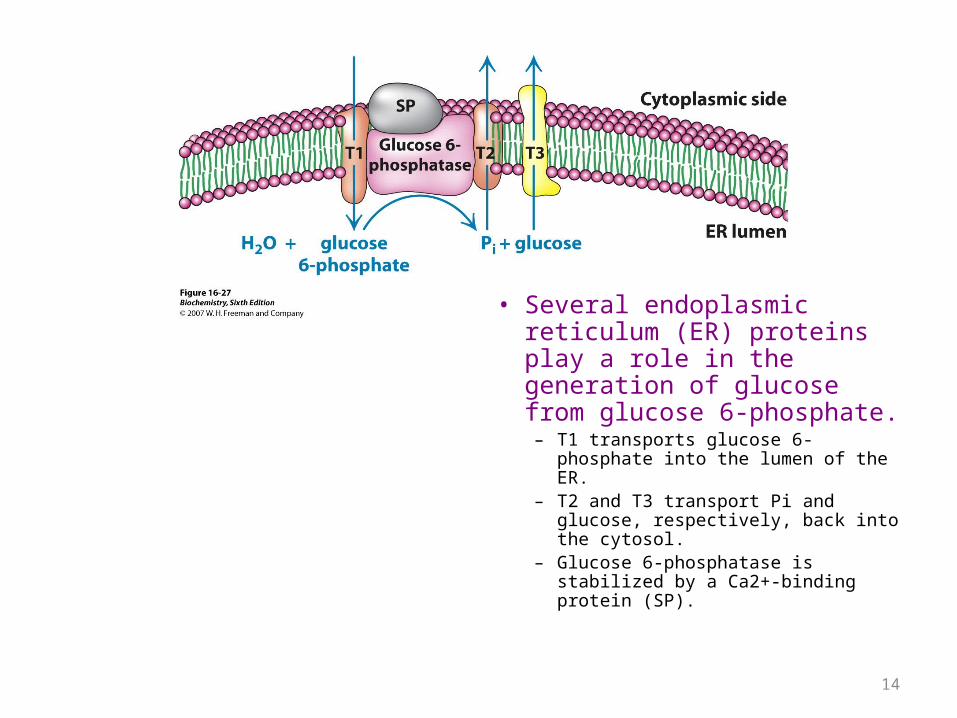
• Several endoplasmic reticulum (ER) proteins play a role in the generation of glucose from glucose 6-phosphate.– T1 transports glucose 6-phosphate
into the lumen of the ER.– T2 and T3 transport Pi and glucose,
respectively, back into the cytosol.– Glucose 6-phosphatase is stabilized
by a Ca2+-binding protein (SP).
14

REGULATION OF CARBOHYDRATE METABOLISM
15

• The rate of conversion of glucose into pyruvate is regulated to meet two major cellular needs:
1. The production of ATP, generated by the degradation of glucose.
2. The provision of building blocks for synthetic reactions, such as the formation of fatty acids.
16

Regulation and control of enzyme activity
• Substrate level control.• Allosteric effectors• Covalent modification• Enzyme concentration:
1. increased synthesis2. generation of active enzyme by processing
• Substrate cycles
17
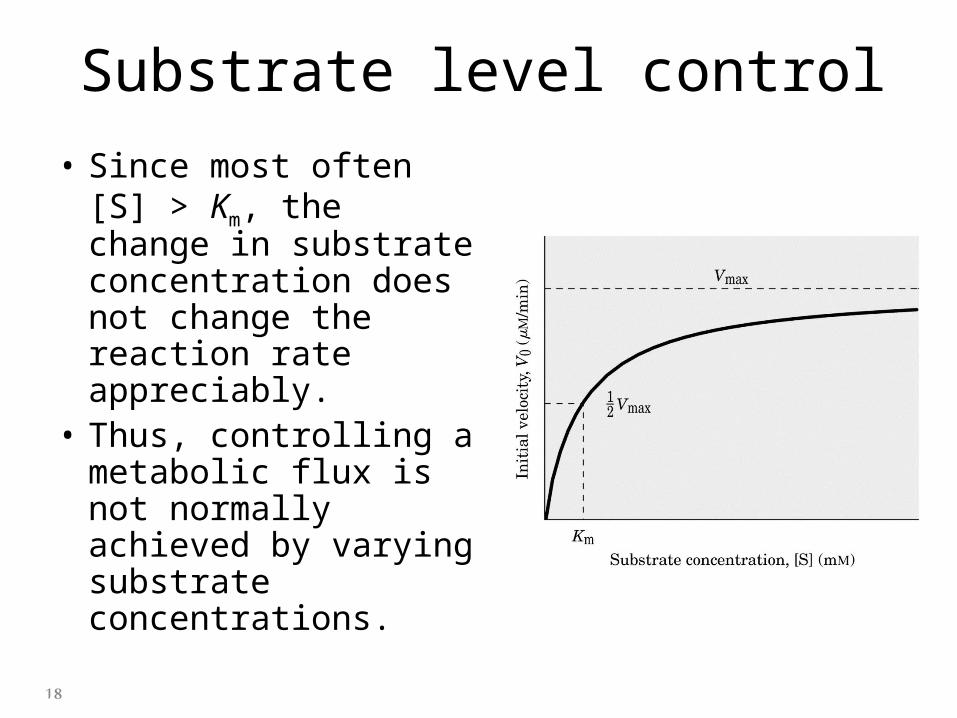
Substrate level control
• Since most often [S] > Km, the change in substrate concentration does not change the reaction rate appreciably.
• Thus, controlling a metabolic flux is not normally achieved by varying substrate concentrations.
18
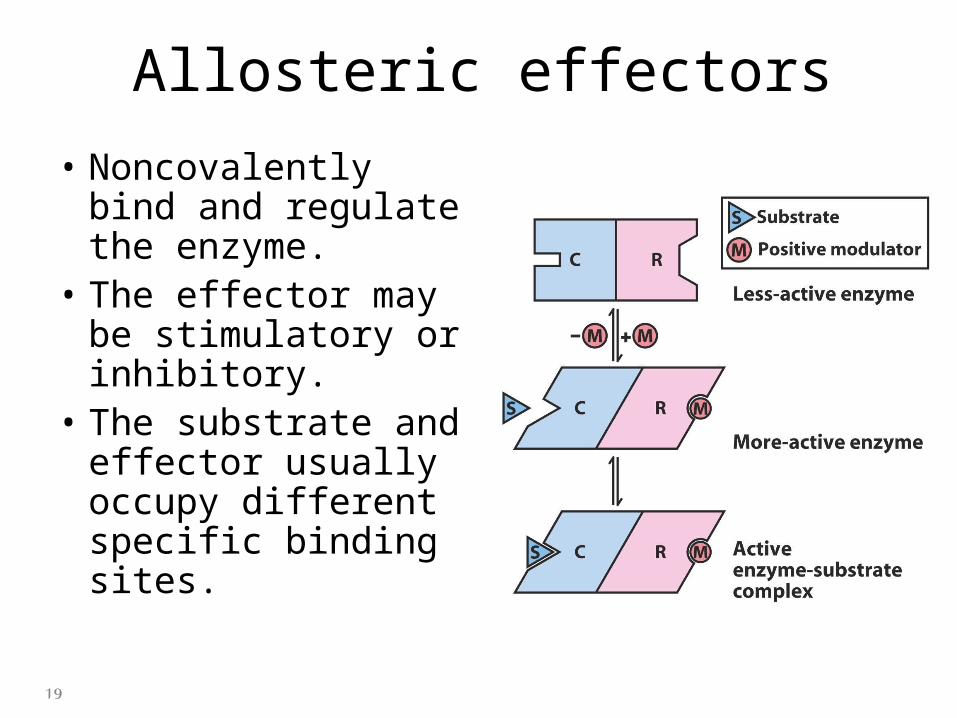
Allosteric effectors
• Noncovalently bind and regulate the enzyme.
• The effector may be stimulatory or inhibitory.
• The substrate and effector usually occupy different specific binding sites.
19
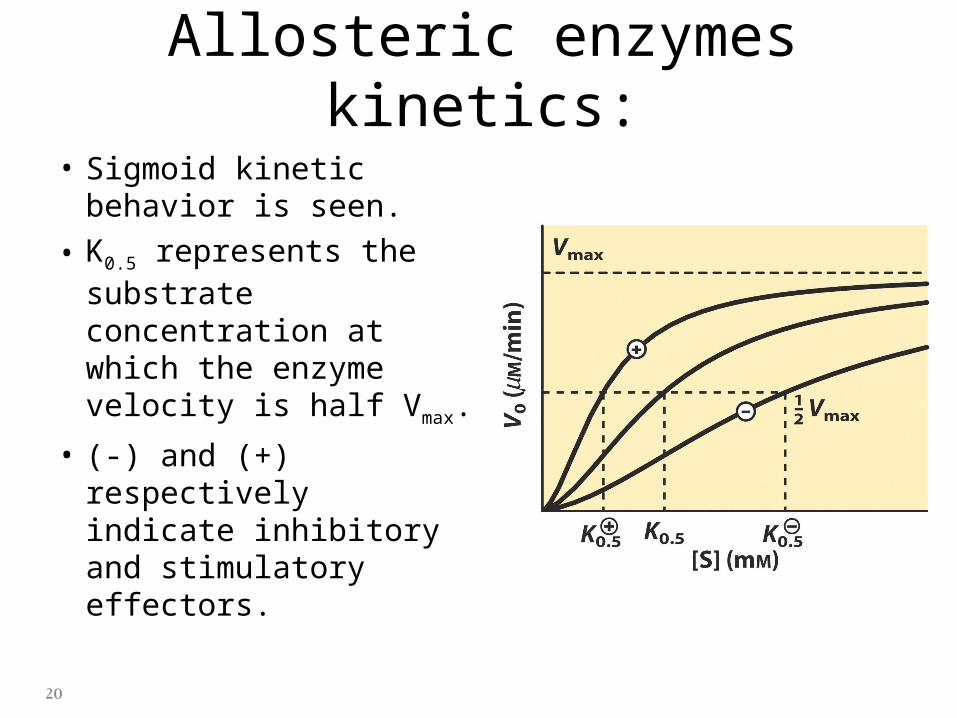
Allosteric enzymes kinetics:
• Sigmoid kinetic behavior is seen.
• K0.5 represents the substrate concentration at which the enzyme velocity is half Vmax.
• (-) and (+) respectively indicate inhibitory and stimulatory effectors.
20
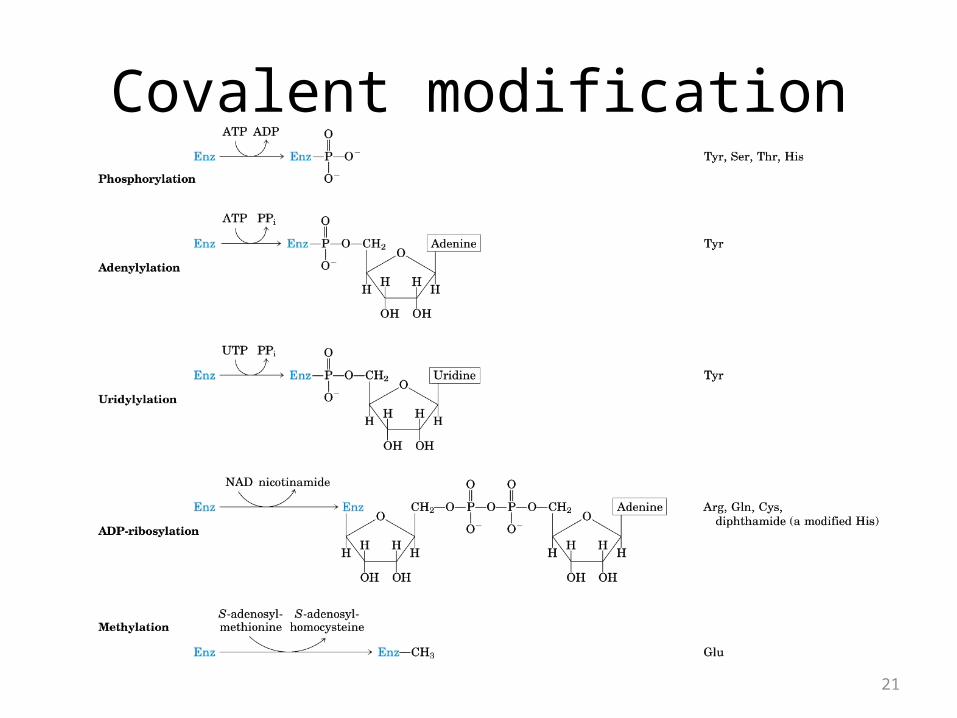
Covalent modification
21

Enzyme concentration: increased synthesis
22
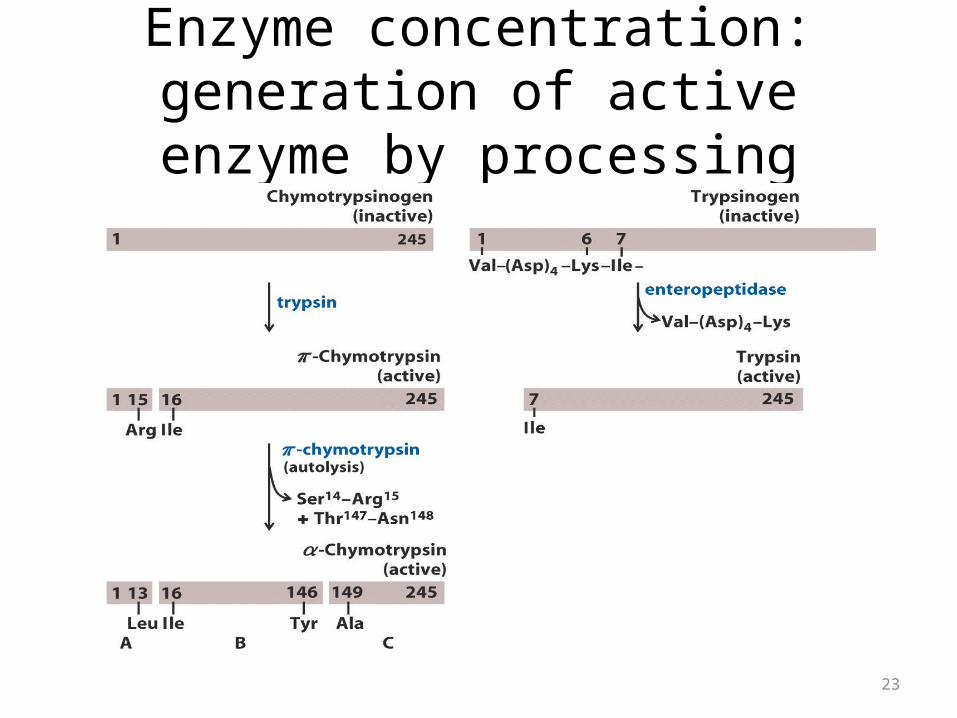
Enzyme concentration: generation of active enzyme by processing
23

Glycolysis & Gluconeogenesis are both spontaneous. If both pathways were simultaneously active in a cell, it would constitute a "futile cycle" that would waste energy. Glycolysis: glucose + 2 NAD+ + 2 ADP + 2 Pi 2 pyruvate + 2 NADH + 2 ATPGluconeogenesis: 2 pyruvate + 2 NADH + 4 ATP + 2 GTP glucose + 2 NAD+ + 4 ADP + 2 GDP + 6 Pi
Questions:1. Glycolysis yields how many ~P ? 2. Gluconeogenesis expends how many ~P ? 3. A futile cycle of both pathways would waste how many ~P per cycle ?
26
4 24

Gluconeogenesis and Glycolysis Are Reciprocally Regulated
• The amounts and activities of the distinctive enzymes of each pathway are controlled so that both pathways are not highly active at the same time.
• The interconversion of fructose 6-phosphate and fructose 1,6-bisphosphate is stringently controlled.– Phosphofructokinase and fructose 1,6-
bisphosphatase are reciprocally controlled by fructose 2,6-bisphosphate in the liver
25
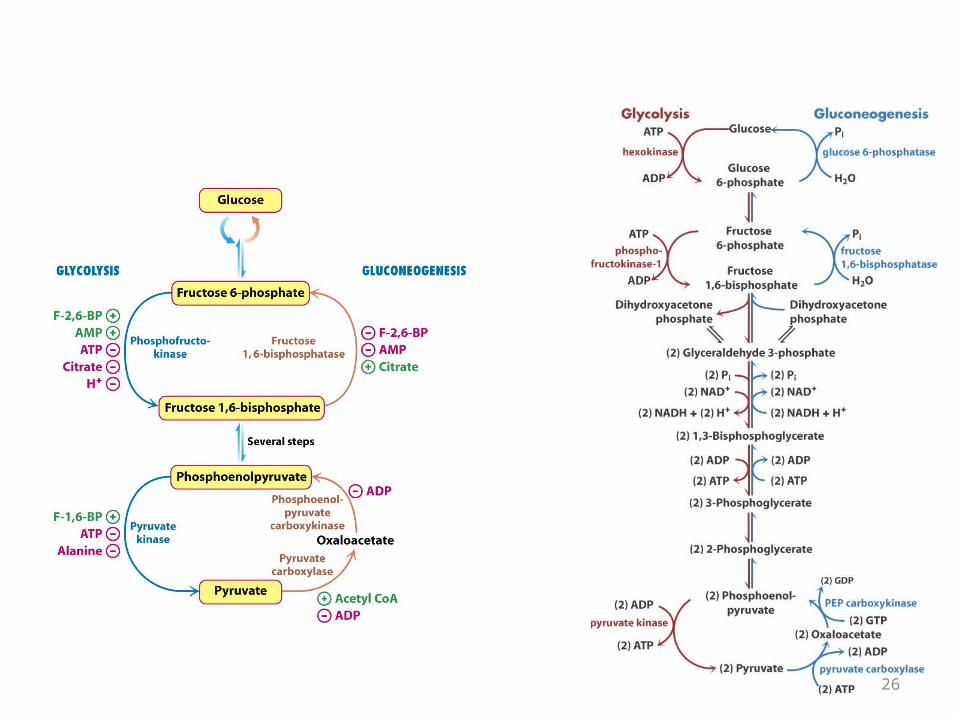
26

ALLOSTERIC REGULATORS OF PFK-1 and FBPase-1
27

PHOSPHOFRUCTOKINASE: The most important control element in the mammalian
glycolytic pathway.• PFK in the liver is a
tetramer of 4 identical subunits.
• The allosteric effectors binding site is distinct from the catalytic site.
28
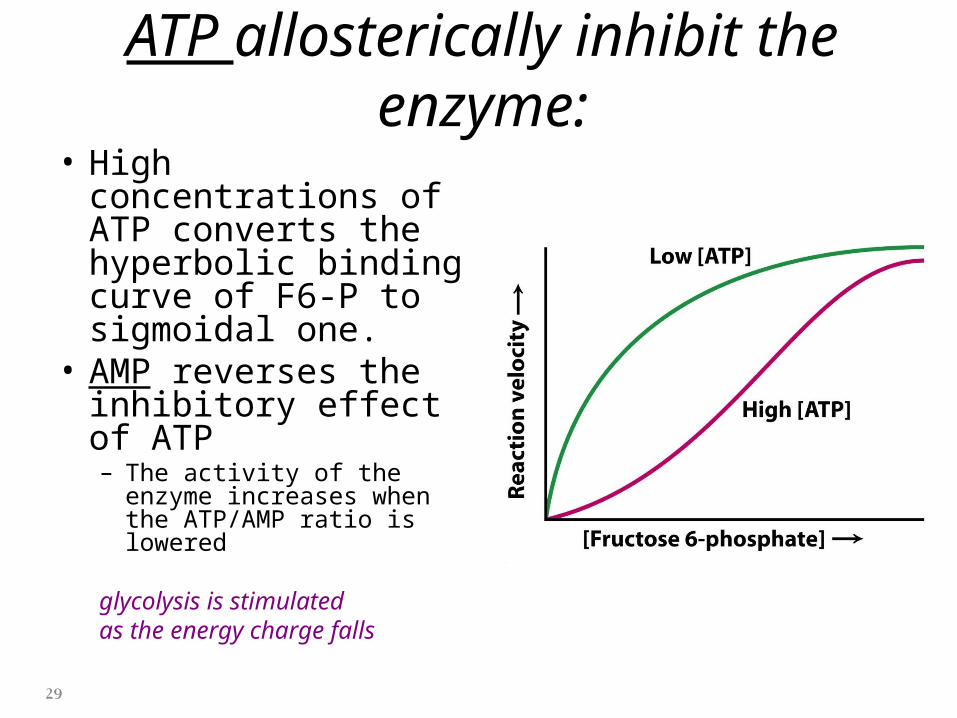
ATP allosterically inhibit the enzyme:• High concentrations of
ATP converts the hyperbolic binding curve of F6-P to sigmoidal one.
• AMP reverses the inhibitory effect of ATP– The activity of the enzyme
increases when the ATP/AMP ratio is lowered
glycolysis is stimulatedas the energy charge falls
29

AMP but not ADP positive regulator of PFK-1
• When ATP is utilized rapidly, the enzyme Adenylate Kinase forms ATP and AMP from ADP:
• AMP becomes the signal for low energy charge.
ADP + ADP ATP + AMP
30

Citrate inhibits PFK-1 enzyme
• A high level of citrate means that biosynthetic precursors are abundant and additional glucose should not be degraded for this purpose.
• Citrate inhibits PFK-1 by enhancing the inhibitory effect of ATP.
31

F2,6-BP allosterically activates PFK-1 and inhibits FBPase:
• Phosphofructokinase and fructose 1,6-bisphosphatase are reciprocally controlled by fructose 2,6-bisphosphate in the liver.
32

How is the concentration of F 2,6-BP appropriately controlled?
• F2,6-BP is formed in a reaction catalyzed by Phosphofructokinase-2 (PFK-2)
• It is hydrolyzed to F6-P by Fructose Bisphosphatase-2 (FBPase-2)
• Both PFK-2 and FBPase-2 are part of the same 55Kd polypeptide chain.
• The bifunctional enzyme Exists in 5 isozymic forms.– L-isoform in liver.– M-isoform in muscle.
33

Why is phosphofructokinase rather than hexokinase the pacemaker of
glycolysis? • Glucose 6-phosphate is not solely a glycolytic
intermediate.– It can also be converted into glycogen or it can be oxidized by
the pentose phosphate pathway to form NADPH.
• The first irreversible reaction unique to the glycolytic pathway, the committed step, is the phosphorylation of fructose 6-phosphate to fructose 1,6-bisphosphate.
34
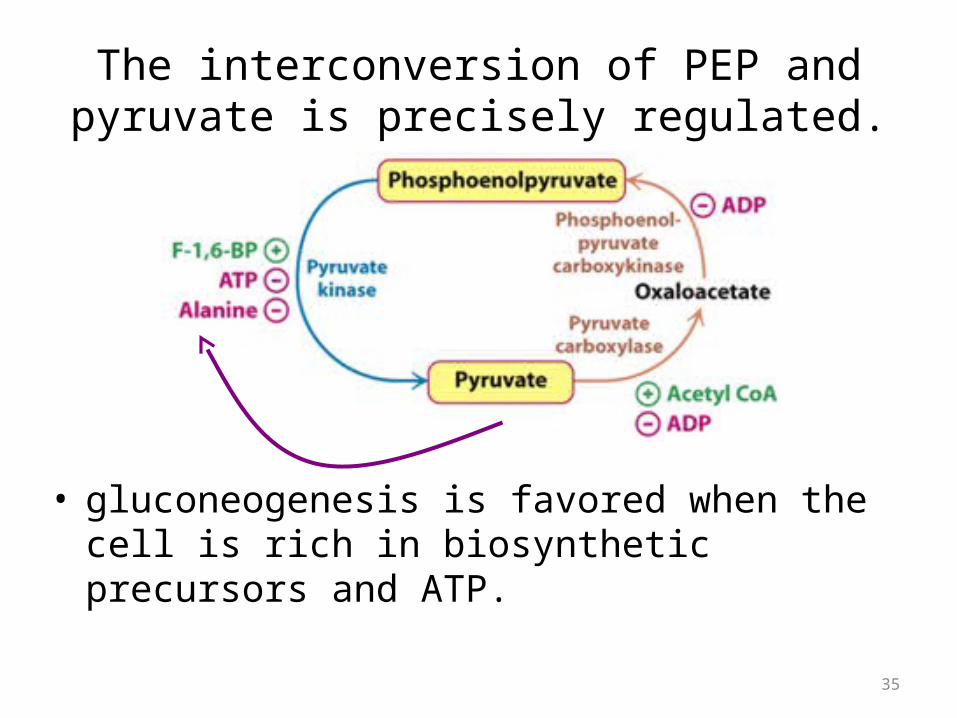
The interconversion of PEP and pyruvate is precisely regulated.
• gluconeogenesis is favored when the cell is rich in biosynthetic precursors and ATP.
35

GLUCOSE TRANSPORTERS
36

HORMONES control the amount and activities of essential enzymes
• Hormones affect gene expression primarily by:1. changing the rate of transcription2. regulating the degradation of mRNA.
• Transcriptional control in eukaryotes is much slower than allosteric control;– it takes hours or days in contrast with seconds to
minutes.
37

Insulin
• Rises subsequent to eating and stimulates the expression of:–Phosphofructokinase–pyruvate kinase–PFK-2/FBPase-2
38

Glucagon
• Rises during starvation:• inhibits the expression of:– Phosphofructokinase– pyruvate kinase– PFK-2/FBPase-2.
• stimulates instead the production of two key gluconeogenic enzymes:– phosphoenolpyruvate carboxykinase– fructose 1,6-bisphosphatase
39
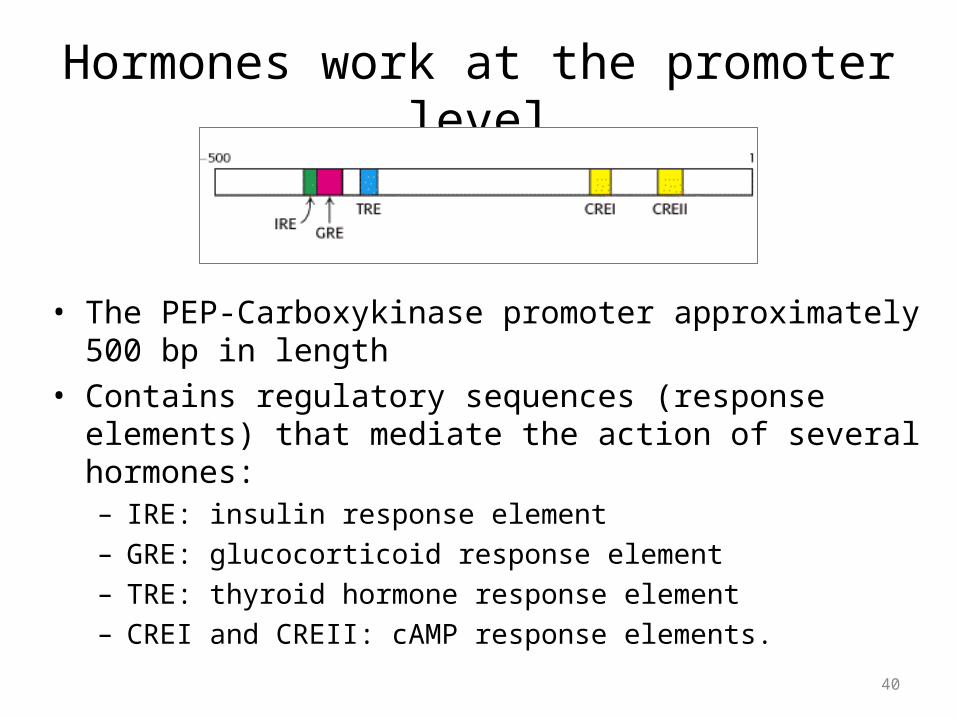
Hormones work at the promoter level
• The PEP-Carboxykinase promoter approximately 500 bp in length• Contains regulatory sequences (response elements) that
mediate the action of several hormones:– IRE: insulin response element– GRE: glucocorticoid response element– TRE: thyroid hormone response element– CREI and CREII: cAMP response elements.
40
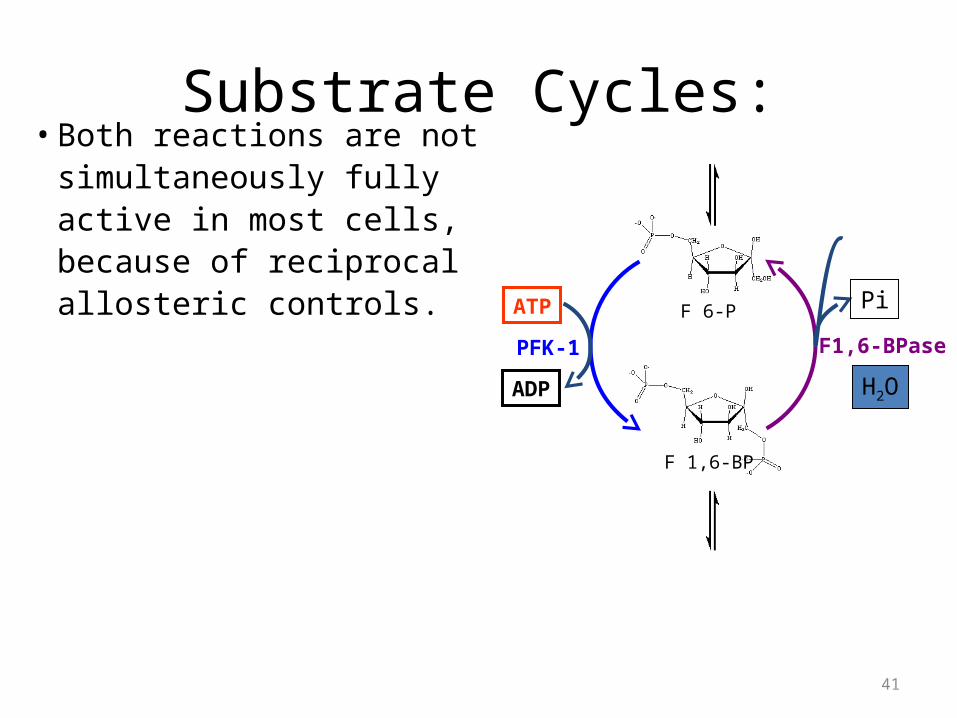
Substrate Cycles:• Both reactions are not
simultaneously fully active in most cells, because of reciprocal allosteric controls.
F 6-P
F 1,6-BP
H2O
Pi
F1,6-BPase
ATP
ADP
PFK-1
41

Substrate cycles are biologically important:
1. Substrate cycles amplify metabolic signals:– This amplification is made possible by
the rapid hydrolysis of ATP.
If an allosteric effector reciprocally increases A to B and decreases B to A by 20% eachThen a 20% change in the rates of the opposing reactions has led to a 480% (=100x48/10) increase in the net flux.
2. generation of heat produced by the hydrolysis of ATP.
42

Lactate produced by active skeletal muscle and erythrocytes is a source of energy for other
organs.• The only purpose of the reduction of pyruvate to
lactate is to regenerate NAD+ so that glycolysis can proceed in active skeletal muscle and erythrocytes.
• lactate is a dead end in metabolism.– It must be converted back into pyruvate before it can be
metabolized.
43

The Cori Cycle operates during exercise.
For a brief burst of ATP utilization, muscle cells utilize ~P stored as phosphocreatine.
Once phosphocreatine is exhausted, ATP is provided mainly by Glycolysis, with the input coming from glycogen breakdown and from glucose uptake from the blood.
(Aerobic fat metabolism, discussed elsewhere, is more significant during a lengthy period of exertion such as a marathon run.)
44
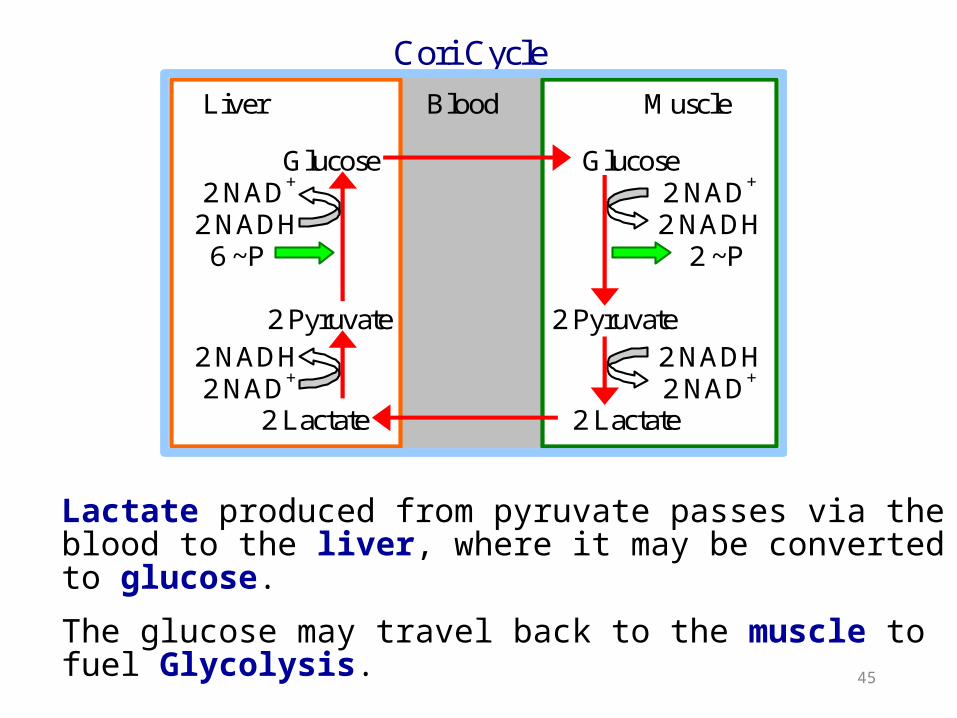
Lactate produced from pyruvate passes via the blood to the liver, where it may be converted to glucose.
The glucose may travel back to the muscle to fuel Glycolysis.
Cori Cycle
Liver Blood Muscle Glucose Glucose 2 NAD+ 2 NAD+
2 NADH 2 NADH 6 ~P 2 ~P 2 Pyruvate 2 Pyruvate 2 NADH 2 NADH 2 NAD+ 2 NAD+ 2 Lactate 2 Lactate
45
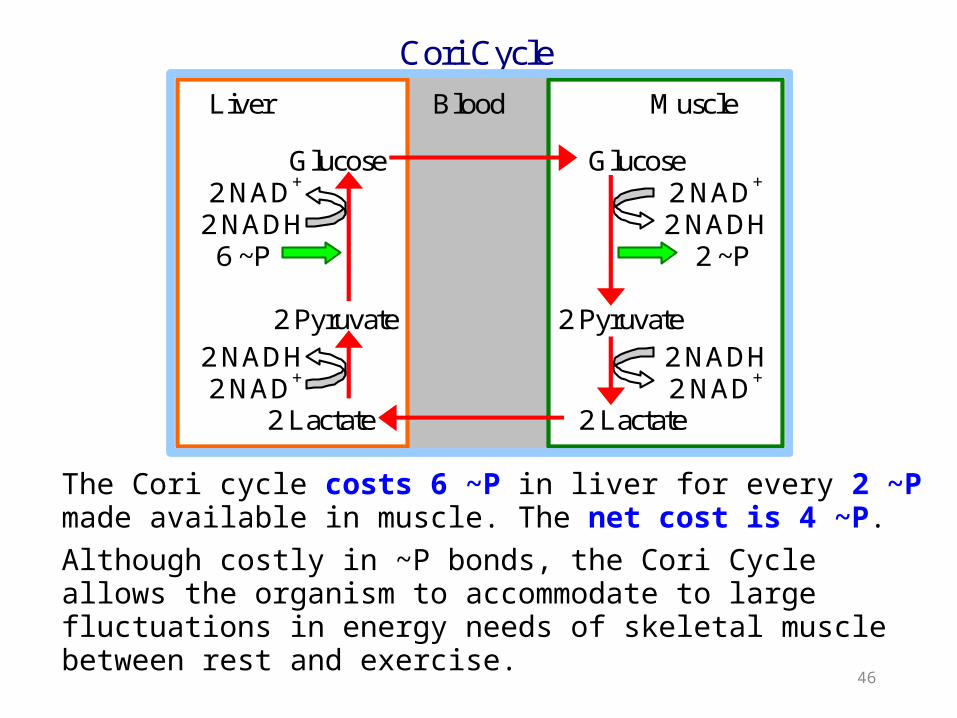
The Cori cycle costs 6 ~P in liver for every 2 ~P made available in muscle. The net cost is 4 ~P. Although costly in ~P bonds, the Cori Cycle allows the organism to accommodate to large fluctuations in energy needs of skeletal muscle between rest and exercise.
Cori Cycle
Liver Blood Muscle Glucose Glucose 2 NAD+ 2 NAD+
2 NADH 2 NADH 6 ~P 2 ~P 2 Pyruvate 2 Pyruvate 2 NADH 2 NADH 2 NAD+ 2 NAD+ 2 Lactate 2 Lactate
46

The equivalent of the Cori Cycle also operates during cancer.
If blood vessel development does not keep pace with growth of a solid tumor, decreased O2 concentration within the tumor leads to activation of signal processes that result in a shift to anaerobic metabolism.
47
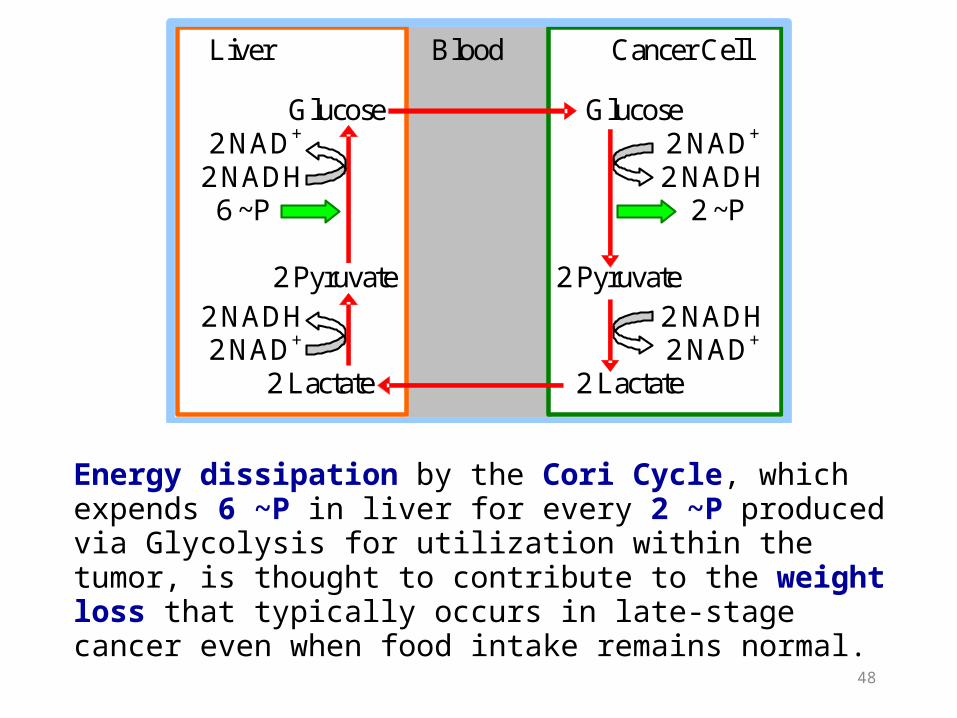
Energy dissipation by the Cori Cycle, which expends 6 ~P in liver for every 2 ~P produced via Glycolysis for utilization within the tumor, is thought to contribute to the weight loss that typically occurs in late-stage cancer even when food intake remains normal.
Liver Blood Cancer Cell Glucose Glucose 2 NAD+ 2 NAD+
2 NADH 2 NADH 6 ~P 2 ~P 2 Pyruvate 2 Pyruvate 2 NADH 2 NADH 2 NAD+ 2 NAD+ 2 Lactate 2 Lactate
48