Embed Size (px)
Citation preview

ORIGINAL ARTICLE Biology
Growth of juvenile chub mackerel Scomber japonicusin the western North Pacific Ocean: with applicationand validation of otolith daily increment formation
Masanori Takahashi • Michio Yoneda •
Hajime Kitano • Atsushi Kawabata •
Mami Saito
Received: 31 May 2013 / Accepted: 26 December 2013
� The Japanese Society of Fisheries Science 2014
Abstract The growth of juvenile chub mackerel Scomber
japonicus collected in the western North Pacific Ocean in
2007 and 2009 was examined based on the evidence of
otolith daily increment formation in captive specimens.
There was a significant difference in the relationship
between known age and number of increments in the frontal
and sagittal planes. Repeated markings on the otolith using
Alizarin complexone and the coefficient of variation in
number of increments suggest that the increments in the
frontal plane of the otolith are more suitable for age esti-
mation than those in the long and short axes of the sagittal
plane. The increments in the frontal plane formed daily, and
the first ring was usually deposited 3 days after hatch. Age of
wild juveniles ranged from 24 to 211 days after hatch based
on the frontal plane method. The estimated hatching periods
of specimens ranged from February to June, but the April-
hatched specimens were collected throughout the sampling
periods of 2007 and 2009. The Gompertz growth model
showed a difference in growth pattern in specimens between
2007 and 2009. The juveniles in 2009 appeared to grow more
quickly than those in 2007 until summer, but thereafter the
2009 specimens seemed to grow more slowly.
Keywords Alizarin complexone � Growth � Otolith �Scomber japonicus � Validation � Western North Pacific
Ocean
Introduction
Chub mackerel Scomber japonicus is one of the most
important commercial fishes around the coastal waters of
Japan, and it has been managed by the total allowable catch
system since 1997. Among the Pacific stock of chub
mackerel, spawning mainly occurs from April to June in
the waters around the Izu Islands, central Japan [1–4]. The
larvae and juveniles drift northeast into the Kuroshio-Oy-
ashio transition area, which functions as a nursery ground,
and then they migrate to the feeding ground around the
waters off the Kuril Islands from June to October [1, 5–8].
The size of the Pacific stock of chub mackerel has
decreased dramatically since the 1980s, and has remained
low over the last few decades [8]. The higher fishing
mortality rates found in younger chub mackerel and the
lower abundance of recruits were considered to prevent
recovery of the population, although relatively large year
classes occurred in several years in the 1990s and 2000s.
M. Takahashi � A. Kawabata
National Research Institute of Fisheries Science, Fisheries
Research Agency, 2-12-4 Fukuura, Kanazawa-Ku, Yokohama,
Kanagawa 236-8648, Japan
Present Address:
M. Takahashi (&)
National Research Institute of Fisheries and Environment of
Inland Sea, Fisheries Research Agency, 2-17-5 Maruishi,
Hatsukaichi, Hiroshima 739-0452, Japan
e-mail: [email protected]
M. Yoneda
Hakatajima Station, National Research Institute of Fisheries and
Environment of Inland Sea, Fisheries Research Agency,
2780 Kinourakou, Hakatachou, Imabari, Ehime 794-2305, Japan
H. Kitano
Fisheries Research Institute of Karatsu, Department of Joint
Research, Faculty of Agriculture, Kyushu University,
6-10-1 Hakozaki, Higashi-Ku, Fukuoka 812-8581, Japan
M. Saito
Environmental Science Laboratory, JAPAN NUS Co. Ltd,
1-1 Fukuura, Kanazawa-Ku, Yokohama,
Kanagawa 236-0004, Japan
123
Fish Sci
DOI 10.1007/s12562-013-0698-2

The long-term trend in the relationship between fish size at
age 0 (about 6 months old) and the abundance of recruit-
ment have indicated that the growth pattern during the
early life stage could be one of the key factors for under-
standing the mechanisms of recruitment fluctuation in the
Pacific stock of chub mackerel [9]. However, little infor-
mation on growth characteristics and survival process of
juvenile chub mackerel in these waters was available.
Microstructural increments (ring marks) in the otolith are
widely used for age determination and to estimate the
growth pattern of fish species during early life stages [10,
11]. In many cases, it has been assumed that microstructural
increments on the otolith are formed daily (e.g., Ma et al.
[12]). However, this assumption may be invalid, as the
otolith increment deposition may not be daily, or easily
discernible in some species [13, 14]. For chub mackerel,
estimates of spawning dates and growth pattern of juveniles
[15, 16] were based on an examination of microstructural
increments on the sagittal otolith, using evidence from
captive Atlantic mackerel Scomber scombrus [17] (Migoya,
unpublished, 1989) or spotted mackerel S. australasicus
[18]. However, no study of the validation of daily-based
formation of microstructural increments on the otolith has
been made in chub mackerel. Also, the microstructure of the
sagittal otolith of chub mackerel shows the occurrence of
accessory primordia (AP) at a specific time in the early life
stage [15]. Thereafter, the otolith grows rapidly from the
AP, giving the otolith a fan-like morphology in the sec-
ondary growth layers. These morphometric characteristics
would make it difficult to obtain a single plane of sectioning
through the core of the otolith to its edge. It is therefore
reasonable to assume that the interpretation and validation
of the microstructural growth pattern on the sagittal otolith
should be a prerequisite to estimating the age and growth of
chub mackerel during the early life stage.
The main purpose of this study is to standardize the
method of juvenile chub mackerel otoliths for the purposes
of age estimation. The microstructural increments on the
different planes of the otolith of captive individuals were
compared through rearing experiment, accompanied by
repeated marking of the otolith using Alizarin complexone
(ALC) at set intervals. The hatch dates and growth patterns
of juveniles were also examined, using specimens collected
from the Kuroshio-Oyashio transition area to the Oyashio
area in 2007 and 2009.
Materials and methods
Experimental set up
Juvenile chub mackerel were caught from the waters off
western Shikoku Island, Ehime Prefecture, and maintained
in the sea cages of a fish-farming company under natural
photoperiod and sea temperature from autumn 2007. In
April 2009, the 2-year-old fish in sea cages were trans-
ferred to the Fishery Research Laboratory of Kyushu
University, Fukuoka Prefecture, and kept in circular tanks
(3 t) under ambient temperature and a natural photoperiod.
After anesthetizing the specimens with 2-phenoxyethanol
(0.02 %), females with oocytes greater than 600 lm in
diameter and spermiating males were injected intramus-
cularly with GnRHa [des Gly10-(D-Ala6) LHRH ethyla-
mide; Sigma-Aldrich, St. Louis, MO, USA] [400 lg/kg
body weight (BW)], combined with coconut oil [19]. After
GnRHa injection, specimens to be returned into the circular
tanks were fed about 1 % BW per day of commercial dry
pellets (Higashimaru Co. Ltd., Tokyo, Japan). Fertilized
eggs were able to be collected every day from 2 days after
GnRHa injection, for about a month [19]. About 1,000 eggs
were transferred into 0.5 t circular tanks under ambient
temperature and a natural photoperiod, and larvae were
hatched out 3 days after spawning. They were fed with
rotifers, type S, enriched with docosahexaenoic acid
(DHA)-enriched fresh-water chlorella (Super Fresh Chlo-
rella-V12; Pacific Trading Co. Ltd., Fukuoka, Japan) from
2 days after they hatched. Artemia nauplii, enriched with a
commercial emulsion of n-3 polyunsaturated fatty acids
(Marine Alfa; Nisshin Marine Tech Co. Ltd., Tokyo,
Japan), were also fed to juveniles from 13 days after
hatching. Fish were fed formula feed (New Alteck;
Marubeni Nisshin Feed Co. Ltd., Tokyo, Japan) 15 days
after hatching.
ALC marking
Three repeated luminescent markings were made to vali-
date the periodicity of microstructural increments on the
sagittal otolith [17, 20]. Specimens were exposed to
25 ppm ALC solution, buffered to rearing pH with NaOH,
for 5.5 h 28 days after hatching. Second and third ALC
treatments were conducted 48 and 68 days after hatching.
Eighty-nine days after they hatched, all specimens were
collected and measured to the nearest 0.1 mm fork length
(FL) and BW. Identification of the developmental stage of
the individuals followed Watanabe [1]. Otoliths of speci-
mens were removed and cleaned by removing tissue frag-
ments, and stored dry until mounting.
Otolith preparation
To determine the optimal plane of sagitta (henceforth ot-
oliths) to observe the microstructural increments, the left
side of the otolith was used to analyze increments in the
frontal plane, while the right side was used for the sagittal
plane (Fig. 1). In both otoliths of a specimen, the rostrum
Fish Sci
123

side was folded to avoid loss by polishing of the curved
otolith edge. The left otolith was embedded in epoxy resin
(Petropoxy 154, Maruto Instrument Co. Ltd., Tokyo,
Japan) and mounted on a glass slide with hot water-soluble
wax (Aqua wax 80, Maruto Instrument Co. Ltd., Tokyo,
Japan), with the ventral side up. The otoliths were polished
with waterproof sandpaper (#600-2000) and alumina pol-
ishing suspension (Baikalox 3.0CR, Baikowski Japan Co.
Ltd., Chiba, Japan) until the nucleus became visible, and
thenthe wax was removed from the glass slide using hot
water. The otolith was turned over and mounted on a glass
slide with epoxy resin, with the dorsal side up. The dorsal
side was polished again with waterproof sandpaper and
alumina polishing suspension above the nucleus. The right
otolith was mounted in clear fingernail enamel, distal side
up, on a glass slide, and polished with waterproof sand-
paper and alumina polishing suspension until the nucleus
became visible.
The number of microstructural increments and ALC
luminescent markings on each plane of the otolith were
counted using an otolith measurement system (Ratoc Sys-
tem Engineering, Tokyo, Japan), comprising a transmitting
light or reflected UV light microscope at 200–400 magni-
fication. To validate daily-based increment deposition on
the otolith, the cumulative number of increments marked
ALC on the otolith and days after hatching ALC treatments
(ages 28, 48 and 68 days) were compared. Also, to deter-
mine the microstructural increments on the optimal planes
of the otolith for age estimation, the number of increments
in three different planes—frontal plane (postrostrum
direction), long axis (postrostrum direction) of sagittal
plane, and short (posterior or anterior direction) axis of
sagittal plane—were compared.
Field collection
Specimens were collected during eight research cruises
conducted to cover the distribution area of juveniles of the
Pacific stock of chub mackerel at the Kuroshio-Oyashio
transition zone and the Oyashio area, between May and
October in 2007 and 2009 (Table 1). A mid-water trawl net
was used (27–30 m mouth opening, 8–17 mm mesh cod-
end, 30–60 min duration at 3.5–5.0 knots). All samples
were frozen on board. If the catch was too numerous (more
than about 100 specimens), about 60 specimens were ran-
domly sampled. A total of 450 field samples of chub
mackerel were used for otolith microstructure analysis.
After measuring FL and wet BW, otoliths were removed
from these samples at a laboratory, and the increment count
was examined using the sagittal and frontal plane method.
Mann–Whitney U test was conducted to compare FL in
each sampling period of both years.
Hatch date and growth analyses
The number of growth increments in individual specimens
in the frontal plane of the otolith were counted and mea-
sured. Daily age of individuals was determined by calcu-
lating the total number of growth increments, plus 2, since
the microstructural increments usually appeared to start
forming 3 days from hatch (see ‘‘Results’’). Hatch date was
calculated by subtracting daily age from sampling date.
The estimated hatch dates were classified into three periods
of a given month: early (days 1–10), middle (days 11–20)
and late (days 21–31).
Prior to the estimate of growth trends of wild fishes,
Gompertz, Logistic and von Bertalanffy equation models
dorsal
ventral
rostrum postrostrumN
short axis
long axis
N
ventral
dorsal
rostrum postrostrum
Fig. 1 Overview of different
plane otolith (sagitta) of chub
mackerel. Upper sagittal plane
of right otolith from the distal
side. Lower frontal plane of left
otolith polished from dorsal
side. Dotted arrows indicate the
direction of increment counting.
N nucleus of otolith
Fish Sci
123
fitted to the observed FL at the age were compared using
Akaike’s Information Criterion (AIC) to select the best
model in this study. Of the three models, the Gompertz
growth model has the lowest AIC, indicating an appropri-
ate growth trend of specimens collected. This agrees with
the studies on growth of Atlantic mackerel S. scombrus
[21–24]. The Gompertz growth model [25] was expressed
as:
Lt ¼ L1e�e�kðt�t0Þ
where Lt is FL at age t (days from hatch), L? is asymptotic
FL at the end of the first growth year (0-year-class), k is a
growth coefficient, t is age (days from hatch) and t0 is point
of inflection (age of maximum growth). The biological
intercept followed Watanabe (L0 = 3.0) [1]. The parame-
ters were estimated by non-linear least-squares fitting using
MS Excel Solver (Microsoft, Redmond, WA, USA). A
measure of goodness of fit was provided by calculating a
coefficient of determination (r2) [26].
Results
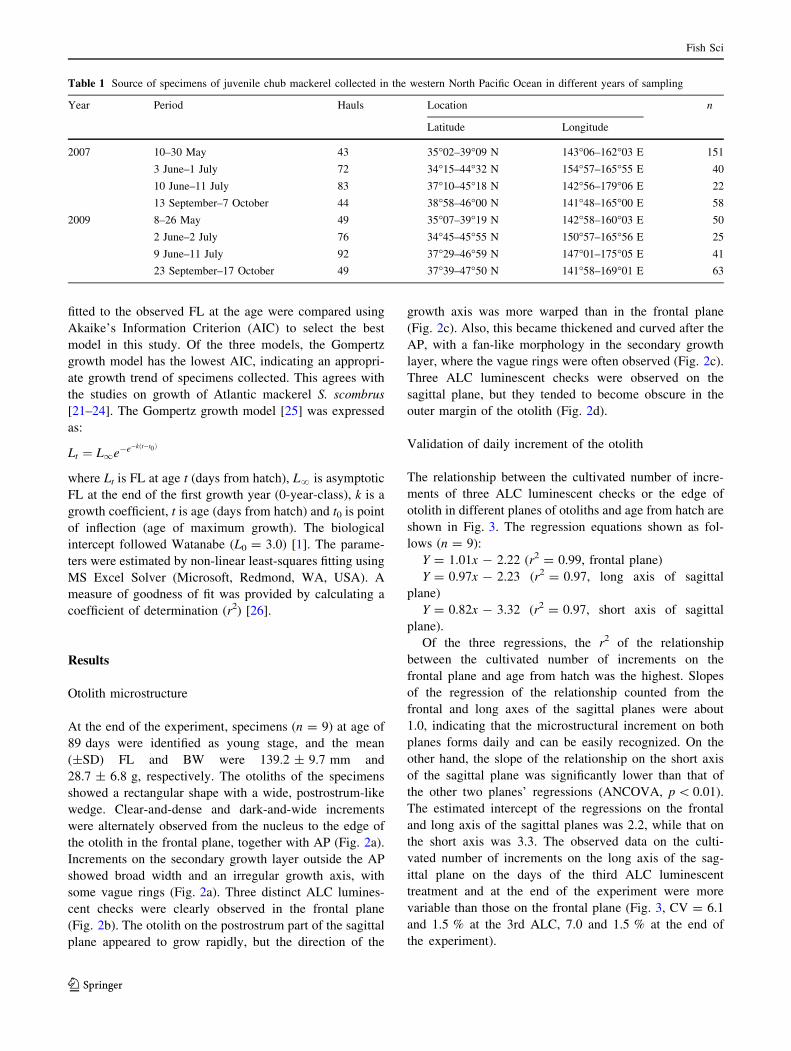
Otolith microstructure
At the end of the experiment, specimens (n = 9) at age of
89 days were identified as young stage, and the mean
(±SD) FL and BW were 139.2 ± 9.7 mm and
28.7 ± 6.8 g, respectively. The otoliths of the specimens
showed a rectangular shape with a wide, postrostrum-like
wedge. Clear-and-dense and dark-and-wide increments
were alternately observed from the nucleus to the edge of
the otolith in the frontal plane, together with AP (Fig. 2a).
Increments on the secondary growth layer outside the AP
showed broad width and an irregular growth axis, with
some vague rings (Fig. 2a). Three distinct ALC lumines-
cent checks were clearly observed in the frontal plane
(Fig. 2b). The otolith on the postrostrum part of the sagittal
plane appeared to grow rapidly, but the direction of the
growth axis was more warped than in the frontal plane
(Fig. 2c). Also, this became thickened and curved after the
AP, with a fan-like morphology in the secondary growth
layer, where the vague rings were often observed (Fig. 2c).
Three ALC luminescent checks were observed on the
sagittal plane, but they tended to become obscure in the
outer margin of the otolith (Fig. 2d).
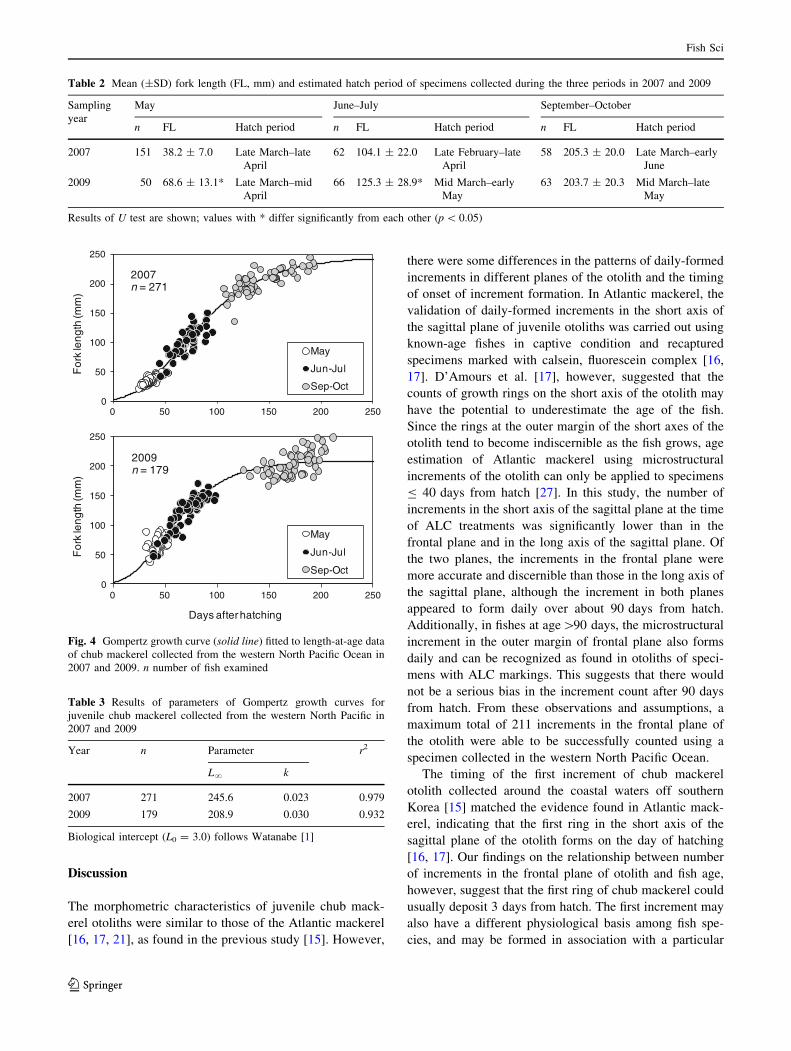
Validation of daily increment of the otolith
The relationship between the cultivated number of incre-
ments of three ALC luminescent checks or the edge of
otolith in different planes of otoliths and age from hatch are
shown in Fig. 3. The regression equations shown as fol-
lows (n = 9):
Y = 1.01x - 2.22 (r2 = 0.99, frontal plane)
Y = 0.97x - 2.23 (r2 = 0.97, long axis of sagittal
plane)
Y = 0.82x - 3.32 (r2 = 0.97, short axis of sagittal
plane).
Of the three regressions, the r2 of the relationship
between the cultivated number of increments on the
frontal plane and age from hatch was the highest. Slopes
of the regression of the relationship counted from the
frontal and long axes of the sagittal planes were about
1.0, indicating that the microstructural increment on both
planes forms daily and can be easily recognized. On the
other hand, the slope of the relationship on the short axis
of the sagittal plane was significantly lower than that of
the other two planes’ regressions (ANCOVA, p \ 0.01).
The estimated intercept of the regressions on the frontal
and long axis of the sagittal planes was 2.2, while that on
the short axis was 3.3. The observed data on the culti-
vated number of increments on the long axis of the sag-
ittal plane on the days of the third ALC luminescent
treatment and at the end of the experiment were more
variable than those on the frontal plane (Fig. 3, CV = 6.1
and 1.5 % at the 3rd ALC, 7.0 and 1.5 % at the end of
the experiment).
Table 1 Source of specimens of juvenile chub mackerel collected in the western North Pacific Ocean in different years of sampling
Year Period Hauls Location n
Latitude Longitude
2007 10–30 May 43 35�02–39�09 N 143�06–162�03 E 151
3 June–1 July 72 34�15–44�32 N 154�57–165�55 E 40
10 June–11 July 83 37�10–45�18 N 142�56–179�06 E 22
13 September–7 October 44 38�58–46�00 N 141�48–165�00 E 58
2009 8–26 May 49 35�07–39�19 N 142�58–160�03 E 50
2 June–2 July 76 34�45–45�55 N 150�57–165�56 E 25
9 June–11 July 92 37�29–46�59 N 147�01–175�05 E 41
23 September–17 October 49 37�39–47�50 N 141�58–169�01 E 63
Fish Sci
123
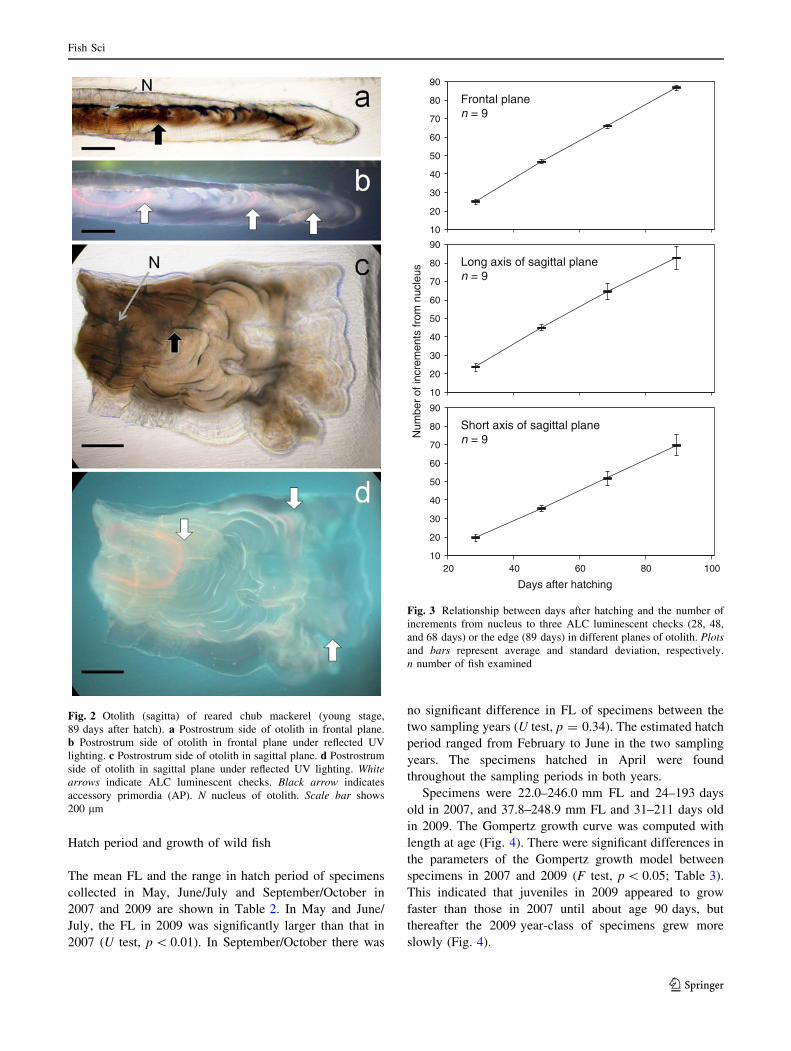
Hatch period and growth of wild fish
The mean FL and the range in hatch period of specimens
collected in May, June/July and September/October in
2007 and 2009 are shown in Table 2. In May and June/
July, the FL in 2009 was significantly larger than that in
2007 (U test, p \ 0.01). In September/October there was
no significant difference in FL of specimens between the
two sampling years (U test, p = 0.34). The estimated hatch
period ranged from February to June in the two sampling
years. The specimens hatched in April were found
throughout the sampling periods in both years.
Specimens were 22.0–246.0 mm FL and 24–193 days
old in 2007, and 37.8–248.9 mm FL and 31–211 days old
in 2009. The Gompertz growth curve was computed with
length at age (Fig. 4). There were significant differences in
the parameters of the Gompertz growth model between
specimens in 2007 and 2009 (F test, p \ 0.05; Table 3).
This indicated that juveniles in 2009 appeared to grow
faster than those in 2007 until about age 90 days, but
thereafter the 2009 year-class of specimens grew more
slowly (Fig. 4).
Fig. 2 Otolith (sagitta) of reared chub mackerel (young stage,
89 days after hatch). a Postrostrum side of otolith in frontal plane.
b Postrostrum side of otolith in frontal plane under reflected UV
lighting. c Postrostrum side of otolith in sagittal plane. d Postrostrum
side of otolith in sagittal plane under reflected UV lighting. White
arrows indicate ALC luminescent checks. Black arrow indicates
accessory primordia (AP). N nucleus of otolith. Scale bar shows
200 lm
10
20
30
40
50
60
70
80
90
20 40 60 80
10
20
30
40
50
60
70
80
90
20 40 60 80
10
20
30
40
50
60
70
80
90
20 40 60 80 100
Num
ber
of in
crem
ents
from
nuc
leus
Days after hatching
Frontal planen = 9
Long axis of sagittal planen = 9
Short axis of sagittal planen = 9
Fig. 3 Relationship between days after hatching and the number of
increments from nucleus to three ALC luminescent checks (28, 48,
and 68 days) or the edge (89 days) in different planes of otolith. Plots
and bars represent average and standard deviation, respectively.
n number of fish examined
Fish Sci
123
Discussion
The morphometric characteristics of juvenile chub mack-
erel otoliths were similar to those of the Atlantic mackerel
[16, 17, 21], as found in the previous study [15]. However,
there were some differences in the patterns of daily-formed
increments in different planes of the otolith and the timing
of onset of increment formation. In Atlantic mackerel, the
validation of daily-formed increments in the short axis of
the sagittal plane of juvenile otoliths was carried out using
known-age fishes in captive condition and recaptured
specimens marked with calsein, fluorescein complex [16,
17]. D’Amours et al. [17], however, suggested that the
counts of growth rings on the short axis of the otolith may
have the potential to underestimate the age of the fish.
Since the rings at the outer margin of the short axes of the
otolith tend to become indiscernible as the fish grows, age
estimation of Atlantic mackerel using microstructural
increments of the otolith can only be applied to specimens
B 40 days from hatch [27]. In this study, the number of
increments in the short axis of the sagittal plane at the time
of ALC treatments was significantly lower than in the
frontal plane and in the long axis of the sagittal plane. Of
the two planes, the increments in the frontal plane were
more accurate and discernible than those in the long axis of
the sagittal plane, although the increment in both planes
appeared to form daily over about 90 days from hatch.
Additionally, in fishes at age[90 days, the microstructural
increment in the outer margin of frontal plane also forms
daily and can be recognized as found in otoliths of speci-
mens with ALC markings. This suggests that there would
not be a serious bias in the increment count after 90 days
from hatch. From these observations and assumptions, a
maximum total of 211 increments in the frontal plane of
the otolith were able to be successfully counted using a
specimen collected in the western North Pacific Ocean.
The timing of the first increment of chub mackerel
otolith collected around the coastal waters off southern
Korea [15] matched the evidence found in Atlantic mack-
erel, indicating that the first ring in the short axis of the
sagittal plane of the otolith forms on the day of hatching
[16, 17]. Our findings on the relationship between number
of increments in the frontal plane of otolith and fish age,
however, suggest that the first ring of chub mackerel could
usually deposit 3 days from hatch. The first increment may
also have a different physiological basis among fish spe-
cies, and may be formed in association with a particular
Table 2 Mean (±SD) fork length (FL, mm) and estimated hatch period of specimens collected during the three periods in 2007 and 2009
Sampling
year
May June–July September–October
n FL Hatch period n FL Hatch period n FL Hatch period
2007 151 38.2 ± 7.0 Late March–late
April
62 104.1 ± 22.0 Late February–late
April
58 205.3 ± 20.0 Late March–early
June
2009 50 68.6 ± 13.1* Late March–mid
April
66 125.3 ± 28.9* Mid March–early
May
63 203.7 ± 20.3 Mid March–late
May
Results of U test are shown; values with * differ significantly from each other (p \ 0.05)
0
50
100
150
200
250
0 50 100 150 200 250
May
Jun-Jul
Sep-Oct
0
50
100
150
200
250
0 50 100 150 200 250
May
Jun-Jul
Sep-Oct
Days after hatching
Fo
rk le
ng
th (m
m)
For
k le
ngth
(mm
)
2007n = 271
2009n = 179
Fig. 4 Gompertz growth curve (solid line) fitted to length-at-age data
of chub mackerel collected from the western North Pacific Ocean in
2007 and 2009. n number of fish examined
Table 3 Results of parameters of Gompertz growth curves for
juvenile chub mackerel collected from the western North Pacific in
2007 and 2009
Year n Parameter r2
L? k
2007 271 245.6 0.023 0.979
2009 179 208.9 0.030 0.932
Biological intercept (L0 = 3.0) follows Watanabe [1]
Fish Sci
123

developmental stage [28]. For example, in Clupea haren-
gus, the first prominent increment of the otolith was formed
well after hatching and towards the end of the yolk-sac
stage [13]. In our experiment, the yolk-sac absorption and
mouth opening of chub mackerel were found about 2 days
from hatch, and first feeding was observed at age 3 and
4 days, as found in a previous study of captive fish [1].
Although the cause of the timing of onset of increment
formation on chub mackerel otolith needs to be clarified,
the first increment can be used as the reference point for
age estimation of this species during the early life stage.
The estimated hatch period of wild juveniles ranged
from February to June in 2007 and 2009. This is consistent
with the spawning period of the Pacific stock of chub
mackerel [1, 29]. In 2007 and 2009, the April-hatched
specimens were found throughout the sampling periods,
although the range of hatch period differed between sam-
pling periods. Watanabe [29] showed that a relatively
higher abundance of chub mackerel eggs collected in the
spawning grounds has been found between May and June
in recent years. Further research of the relationship
between the abundances of eggs and surviving cohort in
these waters needs to clarify the mechanism of recruitment
of this species.
The growth pattern of specimens differed significantly
between the two year classes. The specimens in 2009 grew
faster than those in 2007 until about 90 days from hatch.
This is likely to reflect a difference in FL of specimens in
May and June/July between the two sampling years,
although the ranges of hatch periods were almost identical.
In chub mackerel, the effect of temperature on growth was
evident, with higher temperature resulting in acceleration
of growth rate of larvae and early juveniles under captive
condition [30, 31]. On the other hand, Watanabe and Yatsu
[9] hypothesized that improved feeding conditions may
result in acceleration of juvenile mackerel growth. Thus the
temperature and prey availability that larvae and juveniles
have experienced could affect their growth rate during the
early life stage, although the cause of variation in growth
pattern between specimens in 2007 and 2009 remains
unknown.
In conclusion, our findings showed for the first time that
the daily-formed increments in the frontal plane of the
otolith of chub mackerel were more suitable for estimating
the age of juveniles than those in the short and long axes of
the sagittal planes. The first increment in the frontal plane
of the otolith was usually deposited 3 days from hatch.
These findings led us to estimate the hatch period and
growth pattern of wild juveniles collected in the western
North Pacific Ocean. The range of the estimated hatch
period of specimens was consistent with the previously
reported spawning period of chub mackerel in these waters
[1, 29], but specimens hatched in April were found
throughout the sampling periods in 2007 and 2009. Also,
there was a significant difference in growth pattern of
specimens between the two year classes. Recent field sur-
veys indicate a positive relationship between the mean FL
of juveniles collected in the Kuroshio-Oyashio transition
area and the abundance of recruits in a given year class [8].
Kawabata et al. [8] have suggested that this may be
attributable to variations in the hatch date and growth rates
during larval and early juvenile stages among the year-
classes. Our findings could assist in the proper assessment
of their life history traits to clarify the mechanism of
recruitment dynamics of the Pacific stock of chub
mackerel.
Acknowledgments We thank the staff and students of the Fishery
Research Laboratory and the Laboratory of Marine Biology at Kyu-
shu University for their support in rearing and measuring the speci-
mens. We are grateful to Drs. K. Mori and N. Yamashita of the
Hokkaido National Fisheries Research Institute, Fisheries Research
Agency of Japan (FRA); Drs. Y. Ueno and S. Honda of the National
Research Institute of Fisheries Science, FRA; and Drs. S. Suyama and
M. Nakagami of the Hachinohe Station, Tohoku National Fisheries
Research Institute, FRA, for sample collection; and to the captain,
officers and crews of the training vessels Hokuho-Maru and Omi-
Maru for helping with collection and measurement. We also thank Dr.
T. Akamine for critical comments on the statistical analysis. This
research was funded by a sub-project on studies on the prediction and
application of fish species alternation (SUPRFISH), financed by the
Agriculture, Forestry and Fisheries Research Council of Japan as part
of the Population Outbreak of Marine Life (POMAL) Project, and
financially supported by the Fisheries Agency of Japan. The Fisheries
Agency is not responsible for the content of this paper.
References
1. Watanabe T (1970) Morphology and ecology of early stages of
life in Japanese common mackerel, Scomber japonicus HOU-
TTUYN, with special reference to fluctuation of population. Bull
Tokai Reg Fish Res Lab 62:1–283 (in Japanese with English
abstract)
2. Usami S (1973) Ecological studies of life pattern of the Japanese
mackerel, Scomber japonicus HOUTTUYN, on the adult of the
Pacific sub-population. Bull Tokai Reg Fish Res Lab 76:71–178
(in Japanese with English abstract)
3. Murayama T, Mitani I, Aoki I (1995) Estimation of the spawning
period of the Pacific mackerel Scomber japonicus based on the
changes in gonad index and the ovarian histology. Bull Jpn Soc
Fish Oceanogr 59:11–17 (in Japanese with English abstract)
4. Watanabe C, Hanai T, Meguro K, Ogino R, Kubota Y, Kimura R
(1999) Spawning biomass estimated of chub mackerel Scomber
japonicus of Pacific subpopulation off central Japan by a daily
egg production method. Nippon Suisan Gakkaishi 65:695–702 (in
Japanese with English abstract)
5. Odate K (1994) Zooplankton biomass and its long-term variation
in the western north Pacific ocean, Tohoku sea area, Japan. Bull
Tohoku Natl Fish Res Inst 56:115–173 (in Japanese with English
abstract)
6. Kawasaki T (1966) Structure of the Pacific population of the
mackerel. Bull Tokai Reg Fish Res Lab 47:1–34 (in Japanese
with English abstract)
Fish Sci
123

7. Watanabe C, Nishida H (2002) Development of assessment
technique for pelagic fish stocks: applications of daily egg pro-
duction method and pelagic trawl in the northwestern Pacific
ocean. Fish Sci 68:97–100
8. Kawabata A, Watanabe C, Nishida H, Honda S (2012) Stock
assessment and evaluation for the Pacific stock of chub mackerel
(fiscal year 2011). In: Marine fisheries stock assessment and
evaluation for Japanese waters (fiscal year 2011/2012). Fisheries
Agency and Fisheries Research Agency of Japan, Tokyo,
pp 151–186 (in Japanese)
9. Watanabe C, Yatsu A (2004) Effect of density-dependence and
sea surface temperature on interannual variation in length-at-age
of chub mackerel (Scomber japonicus) in the Kuroshio-Oyashio
area during 1970–1997. Fish Bull 102:196–206
10. Campana SE, Neilson JD (1985) Microstructure of fish otoliths.
Can J Aquat Sci 42:1014–1032
11. Campana SE (2005) Otolith science entering the 21st century.
Mar Freshw Res 56:485–495
12. Ma T, Miller MJ, Shinoda A, Minagawa G, Aoyama J, Tsu-
kamoto K (2005) Age and growth of Saurenchelys (Nettastom-
atidae) and Dysomma (Synaphobranchidae) leptocephali in the
East China Sea. J Fish Biol 67:1619–1630
13. Geffen AJ (1982) Otolith ring deposition in relation to growth
rate in herring (Clupea harengus) and turbot (Scophthalmus
maximus) larvae. Mar Biol 71:317–326
14. Morales-Nin B (1992) Determination of growth in bony fishes
from otolith microstructure. FAO Fish Tech Paper 322:51
15. Hwang S, Lee TW (2005) Spawning dates and early growth of
chub mackerel Scomber japonicus as indicated by otolith
microstructure of juveniles in the inshore nursery ground. Fish
Sci 71:1185–1187
16. Takahashi M, Takagi K, Kawabata A, Watanabe C, Nishida H,
Yamashita N, Mori K, Suyama T, Nakagami M, Ueno Y, Saito M
(2010) Estimated hatching season of the Pacific stock of chub
mackerel Scomber japonicas and spotted mackerel S. austral-
asicus in 2007. Fish Biol Oceanogr Kuroshio 11:49–54 (in
Japanese)
17. D’Amours D, Landry JG, Lambert TC (1990) Growth of juvenile
(0-group) Atlantic mackerel (Scomber scombrus) in the gulf of St.
Lawrence. Can J Fish Aquat Sci 47:2212–2218
18. Watanabe C, Kobayashi K, Kawabata A, Nashida K (2002) Age
and growth of chub mackerel and spotted mackerel. Kaiyo Mon
34(4):261–265 (in Japanese)
19. Shiraishi T, Ohta K, Yamaguchi A, Yoda M, Chuda H, Ma-
tsuyama M (2005) Reproductive parameters of the chub mackerel
Scomber japonicus estimated from human chorionic gonadotro-
pin-induced final oocyte maturation and ovulation in captivity.
Fish Sci 71:531–542
20. Tsukamoto K (1988) Otolith tagging of Ayu embryo with fluo-
rescent substances. Nippon Suisan Gakkaishi 54:1289–1295
21. Simard P, Castonguay M, D’Amours D, Magnan P (1992)
Growth comparison between juvenile Atlantic mackerel
(Scomber scombrus) from the two spawning groups of the
Northwest Atlantic. Can J Fish Aquat Sci 49:2242–2248
22. Kendall AW Jr, Gordon D (1981) Growth rate of Atlantic
mackerel (Scomber scombrus) larvae in the middle Atlantic
Bight. Rapp P v Reun Cons Int Explor Mer 178:337–341
23. Ware DM, Lambert TC (1985) Early life history of Atlantic
mackerel (Scomber scombrus) in the southern Gulf of St. Law-
rence. Can J Fish Aquat Sci 42:577–592
24. Gluyas-Millan MG, Castonguay M, Quinonez-Velazquez CQ
(1998) Growth of juvenile Pacific mackerel, Scomber japonicus,
in the Gulf of California. Sci Mar 62:225–231
25. Ricker WE (1973) Growth rates and models. In: Hoar WS et al
(eds) Fish physiology, vol. 8. Acadmic Press, New York,
pp 677–743
26. Sokal RR, Rohlf FJ (1981) Biometry: the principles and practice
of statistics in biology research, 2nd edn. WH Freeman and Co.,
New York
27. Robert D, Castonruay M, Fortier L (2007) Early growth and
recruitment in Atlantic mackerel Scomber scombrus: discrimi-
nating the effects of fast growth and selection for fast growth.
Mar Ecol Prog Ser 337:209–219
28. Wright PJ, Panfili P, Morales-Nin B, Geffen AJ (2002) Types of
calcified structures. In: Panfili J et al (eds) Manual of fish scle-
rochronology. IRD Editions, Paris, pp 29–88
29. Watanabe C (2010) Change in the reproductive traits of the
Pacific stock of chub mackerel Scomber japonicus and their
effects on the population dynamics. Bull Jap Soc Oceanogr
74:46–50 (in Japanese with English abstract)
30. Hunter JR, Kimbrell CA (1980) Early life history of Pacific
mackerel, Scomber japonicus. Fish Bull 78:89–101
31. Murata O, Yamamoto S, Ishibashi R, Oka Y, Yoneshima H, Kato
K, Miyashita S, Kumai H (2005) Egg development and growth of
larval and juvenile cultured chub mackerel Scomber japonicus
(Perciformes: Scombridae) in a captive spawning experiment.
Aquacult Sci 53:319–324
Fish Sci
123





























