Embed Size (px)
Citation preview

Journal of Ecology
2001
89
, 608–615
© 2001 British Ecological Society
608
Blackwell Science, Ltd
Growth strategies of a shade-tolerant tropical tree: the interactive effects of canopy gaps and simulated herbivory
ARTHUR G. BLUNDELL* and DAVID R. PEART
Department of Biological Sciences, Dartmouth College, Hanover, NH, USA
Summary
1
We hypothesized that the survival of shade-tolerant juvenile trees in the understoreydepends on allocation strategies that slow their growth but enhance recovery fromherbivore damage. In contrast, allocation patterns should maximize height growth in gapswhere competitors grow rapidly. We tested the interactive effects of canopy gaps andsimulated herbivory (by removing the apical meristem, or 10%, 50% or 90% of tissuefrom all leaves) on juvenile (< 1 cm diameter at breast height)
Shorea quadrinervis
Sloot(Dipterocarpaceae), a dominant canopy tree in Borneo.
2
Damage did not diminish survival over 8 months, except for 90% foliage removalfrom understorey plants.
Height growth in the understorey was negligible in both control and damaged plants.In gaps, height growth was stimulated by removal of either the apical meristem or 10%of leaf tissue; growth was reduced only after 90% foliage removal.
3
Higher damage levels increased subsequent net leaf loss. Leaf production was muchgreater, but leaf retention much lower, in gaps than in the understorey. Ninety per centfoliage removal drastically reduced production in gaps and retention in the understorey.After 10% defoliation, height growth was enhanced in gaps despite approximately 30%net leaf loss.
4
In a separate experiment, juveniles were moved from a shade house to gaps. After8 months, gap plants had more leaves and more total biomass, but only half theroot : shoot ratio of plants left in the shade.
5
Allocation patterns, together with the survival of understorey plants after all but thehighest levels of damage, indicate that root : shoot allocation there tends to reduce therisk of mortality following herbivore or mechanical damage at the expense of growth. Incontrast, in gaps, where interference competition for light is high, resource allocationapparently maximizes height growth.
The interaction between resource availability and herbivore damage provides the basisfor a more synthetic theory of resource allocation and growth strategies than one basedon resource availability alone.
Key-words
: artificial defoliation, Gunung Palung National Park, Kalimantan Barat,Indonesia, meristem damage, overcompensation, plant–herbivore interactions, seedlings,south-east Asia, tropics
Journal of Ecology
(2001)
89
, 608–615
Introduction
Although ‘feeding by herbivorous animals almostinvariably reduces plant performance and presumably
plant fitness’ (Crawley 1989), compensatory responsesmay mitigate the effects of animals on the dynamics ofindividual plants, populations and communities. Indeed,for tropical forest seedlings, evidence for herbivoreeffects is mixed. As little as 8% foliage loss to herbivoresdecreased the 18-month survival of newly germinated
Dipteryx panamensis
(Fabaceae) seedlings at La Selva,Costa Rica (Clark & Clark 1985). However, removal ofmammalian herbivores had no effect on the 2-yearsurvival of juveniles (< 1 m tall) in the understorey in
*Present address and correspondence: Arthur G. Blundell,National Center for Environmental Assessment, AmericanAssociation for the Advancement of Science, US EnvironmentalProtection Agency, 1200 Pennsylvania Street NW, MailStop 8601 D, Washington, DC 20460, USA (fax 202.565.0059;e-mail [email protected]).
JEC581.fm Page 608 Saturday, July 14, 2001 1:35 PM

609
Effects of herbivory and gaps on juvenile performance
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 608–615
Panama and Peru (Terborgh & Wright 1994), and onlysevere defoliation reduced growth and survival ofneotropical shrubs (Marquis 1984; Dominguez &Dirzo 1994) and recently germinated seedlings of twoCentral American pioneer species (Núñez-Farfán &Dirzo 1991). Similarly, artificial defoliation of 25% ofeach leaf (double the naturally occurring partial leaf loss)had no effect on 2-year survival of newly germinated
Shorea leprosula
and
S. maxwelliana
(Dipterocarpaceae)seedlings (Becker 1983).
In contrast to the variable and often slight effectsof herbivory, canopy gaps have clear, consistent andstrongly positive effects on growth, as well as providingjuveniles with physical access to the canopy (Denslow1980; Platt & Strong 1989). The comparison betweenherbivore effects in gaps and understorey is particularlyinteresting for juveniles of shade-tolerant trees thatapparently require gaps to grow to maturity (Hartshorn1978; Martínez-Ramos
et al
. 1988, 1989). Juveniles ofsuch trees (including our study species,
Shorea quadrin-ervis
Sloot (Dipterocarpaceae) ) must be able to persistfor long periods in the shaded understorey, but alsocompete effectively with neighbours when exposed tohigh light in gaps (Swaine & Whitmore 1988; Uhl
et al
.1988).
Accordingly, we predicted an interaction betweenherbivore effects and canopy cover, specifically thatunderstorey plants should be less able to cope withherbivory than fast-growing juveniles in gaps. Theextremely slow growth frequently reported for seedlingsin the understorey of tropical forests (Swaine 1996)suggests that juveniles have little residual energy, eitherfor height growth or replacement of lost tissue. Thus,effects of herbivory on understorey plant performanceshould be expressed as higher mortality, whereas gapplants may respond to detrimental foliage loss witheither increased mortality or lower growth. Further,because of the high potential carbon gain in high light,we predicted that greater foliage loss would be requiredto cause mortality in gaps than in the understorey.
Howe (1990) noted that the relative influences ofcanopy condition and herbivory on juvenile dynamicswere virtually unexplored for any species. However, thelimited evidence is consistent with our predictions. Foryoung seedlings of the neotropical canopy tree,
Virolasurinamensis
(Myristicaceae), survivorship was low inthe understorey and herbivory retarded growth ingaps (Howe 1990). Similarly, for six tree species in twotropical rain forests in Queensland, Australia, herbivoryincreased mortality of seedlings in the understoreymore than in gaps (Osunkjoya
et al
. 1993). In threetemperate tree species, browsing reduced seedlinggrowth rates more in high than in low light (Canham
et al
. 1994; Saunders & Puettmann 1999). Whitmore &Brown (1996) demonstrated that herbivory by cricketsin gaps (but not the understorey) altered the compet-itive hierarchy in juveniles of five dipterocarp species.
If plant allocation strategies distribute resourcesoptimally to maximize growth, understorey plants should
invest more in leaves, relative to gap plants, thus increas-ing access to the limited resource of sunlight. In con-trast, plants with access to high light should allocaterelatively more to roots, thus increasing access to below-ground resources, to support high rates of photosynthesisper unit leaf area (Bloom
et al
. 1985).However, the opposite trend is predicted if two
additional factors are included: the need to controlthe risk of mortality due to herbivore damage in thelight-poor understorey, and the need for an aggressivegrowth response in gaps. Resource allocation to above-and below-ground tissue is likely to influence theresponse of juveniles to herbivore damage, for tworeasons. First, because above-ground tissue is moreavailable to most herbivores, greater allocation belowground can decrease the risk of tissue loss. Second, thereplacement of above-ground tissue lost to herbivoresmay be facilitated by greater allocation below ground,as stored energy can be mobilized for reconstructionabove ground (Tilman 1988; Chapin
et al
. 1990; Saunders& Puettmann 1999), especially refoliation (Kobe 1997;Canham
et al
. 1999). However, selective pressures differbetween gaps and the understorey. In gaps, which arerare in space and time, yet provide the only access tothe canopy, competitive and, eventually, reproductivesuccess depends on rapid increase in height wheneverit is energetically possible (Humphrey & Swaine 1997).In summary, allocation strategy in the low light of theunderstorey should place a premium on survival over themany years that elapse before a gap opens, by exposingless biomass to herbivores and allocating substantialresources below ground (Kitajima 1994); in gaps, heightgrowth should take priority, to avoid suppression byneighbours in competition for light.
In the understorey,
S. quadrinervis
juveniles havea mean mortality rate of approximately 10% year
–1
and a mean height growth of approximately 1 cmyear
–1
(Blundell 1999). Thus, less than 40% of a cohortof juveniles would be expected to survive a periodof 10 years during which they would (on average)grow only 10 cm in height. While ‘waiting’ for a canopyopening, priority allocation to the less vulnerablebelow-ground tissue (reflected in a high root : shootratio) would provide a source of energy that can betapped for replacement of leaf tissue (Kobe 1997;Canham
et al
. 1999) lost to either herbivores orphysical damage (e.g. from falling litter; Clark & Clark1989).
Evaluating the trade-offs between opposing predic-tions will depend on quantitative estimates of effectsand risks. In this paper, we focus on comparing theeffects of foliage loss between juveniles (< 1 cm diameterat breast height, d.b.h.) of
S. quadrinervis
growing ingaps and the understorey, using simulated herbivoredamage. We also include a simple experiment to test forchanges in root : shoot allocation in plants that experi-ence a transition from shade conditions to exposure incanopy gaps.
We address the following questions:
JEC581.fm Page 609 Saturday, July 14, 2001 1:35 PM

610
A. G. Blundell, & D. R. Peart
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 608–615
1.
What are the effects of removal of the apicalmeristem, and various proportions of leaf tissue, onthe growth and survival of juveniles under naturalconditions, and how do the effects of damage dependon canopy cover (understorey vs. gaps)?
2.
What are the leaf dynamics of juveniles subjected tosimulated herbivory in the understorey and in gaps?
3.
Does exposure of a juvenile to a canopy gap alterthe allocation of biomass to above- and below-groundtissues?
Methods
Experiments were conducted in a 75-ha area ongranite-derived soils (100–350 m above sea level) at theCabang Panti Research Station (Curran & Leighton2000), Gunung Palung National Park, KalimantanBarat, Indonesian West Borneo (1.13
°
S, 110.7
°
E).There is no hunting or logging in the study area. Theclimate is aseasonal, except for a slightly drier period inJuly and August, with 4.5 m of rain annually (Webb &Peart 1999). ‘El Niño’ years often include especiallydry periods; one occurred over the final 3 months ofour 8-month study and was associated with haze fromforest fires burning in disturbed areas of Borneo. Theseconditions may explain why all plants in our study(including non-manipulated controls) experienced anet loss of leaves (see Results).
The family Dipterocarpaceae dominates the rainforests of south-east Asia.
S. quadrinervis
is an abundantcanopy species throughout the lowlands of Borneo(Ashton 1988) and is the second most common
Shorea
species at the study site, comprising 5% of basal area(C.O. Webb, unpublished data). Like most dipterocarps(Ashton 1988),
S. quadrinervis
is mast fruiting, exclusivelywind-dispersed and facultatively ectomycorrhizal.Section Mutica of the genus
Shorea
, which includes
S. quadrinervis
, contains some of the most light-responsivedipterocarp species (Ashton 1982). Like all dipterocarps,juveniles of
S. quadrinervis
have a monopodial stemwith opposite leaves presented horizontally on multiplebranches that facilitate light interception in the shadedunderstorey (Webb & Curran 1996).
Natural levels of herbivore damage on 3499 juvenilesof
S. quadrinervis
(in the understorey only) weremeasured by Blundell (1999). The mean percentage ofpartial tissue loss on extant leaves was approximately10%, but ranged up to 90%. Only for the 12% of juvenilesthat had lost greater than 60% of tissue was mortalitysignificantly increased over the following 17 months.
Amongst invertebrates, caterpillars and grasshopperscause most damage on
S. quadrinervis
juveniles, butleaf rollers, gall-forming insects and leaf miners alsooccur. Vertebrate browsers are present (Blundell 1996),including sambar deer (
Cervus unicolor
), barking deer(
Muntiacus atherodes
) and mousedeer (
Tragulus javanicus
,
T. napu
), but we do not have evidence of their impact on
dipterocarp juveniles at the study site (Webb 1997;Blundell 1999).
In January 1997, we simulated herbivory by removingleaf or meristem tissue from juveniles and measured theeffects over the following 8 months. The experimentprovides a controlled comparison of the effects of her-bivory in gaps and the understorey, and an evaluationof the interaction between the effects of tissue loss andcanopy cover. It may not provide accurate estimatesof natural herbivore effects for given percentages offoliage removal because we removed a fixed percentageof each leaf, whereas under natural conditions damagevaries among leaves. The coefficient of variation ofpartial tissue loss among
S. quadrinervis
leaves is 120%(A.G. Blundell, unpublished data). Because plants canapparently compensate better for a given amount ofherbivore damage if it is evenly distributed over theplant (Marquis 1992), the effects we measured prob-ably underestimate the impact of similar rates of tissueremoval by real herbivores.
The 75-ha study site contained six gaps largerthan 100 m
2
; we marked and recorded the height of all
S. quadrinervis
juveniles (< 1 cm d.b.h.;
n
= 208) inthese gaps. We found no juveniles < 10 cm tall. Each gapwas paired with an understorey sample area centredat a point 50 m away from the gap edge on a randomazimuth. In each understorey location we markedthe juveniles nearest this central point, increasing theradius until the sample size equalled that in the pairedgap sample. We will refer to each such pair of samplesas a sampling location. Light levels were recorded onall juveniles between 10.00 and 14.00 on overcastdays during 1–7 September 1997. Although no gap waslarger than 500 m
2
, juveniles in gaps experienced morethan six times the light levels (278 lux
±
21.7; mean
±
1SE) of those in the understorey (43 lux
±
3.1; Wilcoxontest
χ
2
= 155.8,
P
< 0.0001; hand-held meter, ExtechInstruments, Waltham, Massachusetts, USA). Relativeto completely open sky, the largest gap received 16%of sunlight (Blundell 1999; as measured at gap centre,50 cm above ground, using a fish-eye lens and GeneralLight Index software (Canham 1988) ).
We simulated herbivory using scissors, cutting acrossthe midrib to remove either 10%, 50% or 90% of tissuefrom every leaf on each juvenile (
n
= 40 for each damagetreatment
×
habitat combination). The removal treat-ments covered the range of foliage loss found on plantsunder natural conditions (see above). We removed theapical meristem from an additional 38 juveniles in gapsand 40 in the understorey. After treatment, herbivoreshad normal access to all juveniles. Controls consistedof a further 50 non-manipulated juveniles in gaps anda large sample (
n
= 3499) of marked, non-manipulatedunderstorey juveniles from a separate study (see above).All controls were in the same study area and within200 m of one of the six gaps.
JEC581.fm Page 610 Saturday, July 14, 2001 1:35 PM

611
Effects of herbivory and gaps on juvenile performance
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 608–615
At the end of the experiment we recorded changein height to apical meristem (cm) and plant mortality.As an index of natural herbivory experienced by eachplant, we measured partial tissue loss on the newestleaf produced over the 8-month period. Percentageloss, rather than absolute area lost from the newestleaf, was used to facilitate comparison among leavesof different size (Lowman 1984). We used the newestleaf because differences in longevity among leaves(Kikuzawa 1995) suggest that older leaves are lesscomparable in chronological age than are the youngest.Further, most herbivory on tropical forest plants occursduring leaf expansion (Coley & Barone 1996).
For all juveniles, we recorded the net change inleaf number over the 8 months. For juveniles in thesimulated herbivory treatments, we also monitored leafdynamics, by counting the number of damaged leavesretained at the end of the experiment, calculating thenumber of leaves that had been shed, and counting thenumber of new leaves produced since the initiation ofthe experiment.
Sampling location did not influence the results of theexperimental treatments (i.e. there was no significantinteraction between treatment and location for any ofthe dependent variables;
P
> 0.19), so data were pooledover locations. Although chosen by the same criterion(i.e.
S. quadrinervis
juveniles < 1 cm d.b.h.), gap plantsdiffered in both height and leaf number from under-storey plants. At the beginning of the experiment,juveniles in the understorey were only 57% as tall as gapplants (60.1
±
2.0 cm (mean
±
1 SE) vs. 34.0
±
0.3 cm;Wilcoxon test
χ
2
= 201.3,
P
< 0.0001) and had 50%as many leaves (26.3
±
1.1 leaves vs. 13.0
±
1.1 leaves;Wilcoxon test
χ
2
= 181.2,
P
< 0.0001). We controlledfor height by regressing change in height against initialheight and then using the residuals of change in heightas the dependent variable in statistical tests. This methodwas preferred to the more traditional approach ofcalculating relative growth rate (RGR). Use of RGRis problematic in woody plants because the ratio ofphotosynthetic to non-photosynthetic tissue increaseswith plant size (Kozlowski & Pallardy 1997). The analysesusing residuals produced the same significant trendsas those using RGR (the latter are not reported here).Similarly, we controlled for total leaf number per plantin analyses in which a measure of leaf dynamics (i.e. changein leaf number, leaf production and leaf loss) was thedependent variable. This was done by using the residuals,separately obtained for each measure of leaf dynamics,in regressions against the initial number of leaves.
Twenty-four
S. quadrinervis
juveniles (
c.
15 cm tall)were collected from the understorey around an adult700 m from the study site, with main roots intact andsoil attached. Roots and soil were placed in polyethylenebags (25 cm high and 15 cm diameter) and the plants
were moved to a shade house under tree cover near theresearch station. Plants were watered liberally to reducetransplant shock. Light level in the shade house wasadjusted to be similar to that in the understorey of thestudy site. After 4 months, 12 juveniles were randomlyselected and placed into a large artificial gap 20 maway; the remaining 12 were kept in the shade houseto protect them from falling debris. Water availabilityin the soils at the study site is high (Webb 1997) exceptduring EL Niño drought periods, so all juveniles werewatered liberally. After a further 8 months (January–September 1997) we removed the plants from thebags and soaked the roots for 36 hours. This allowedsoil to be shaken free, preserving most of the fine rootsintact. There was no indication that any plants wererootbound (A.G. Blundell, personal observation).We measured the wet-weight of above- and below-ground tissue (hereafter referred to as root and shoot)and dried the tissue at 50
°
C in an oven until dry-weights stabilized.
Results
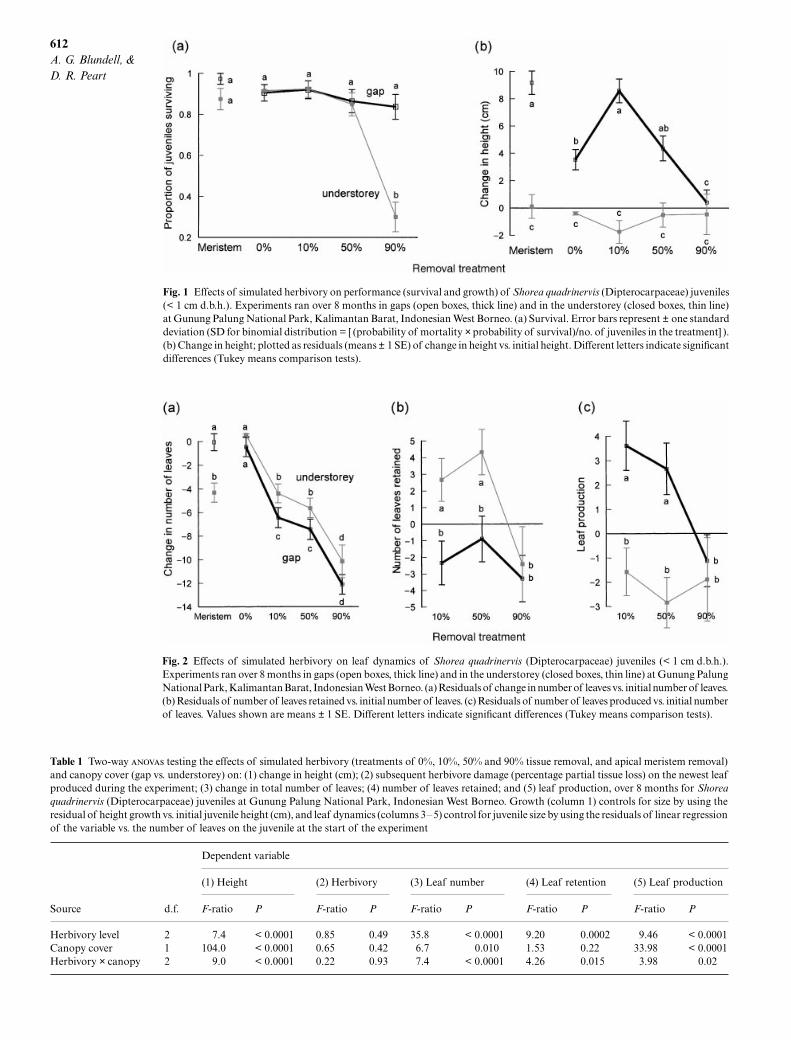
In gaps, juvenile survival over 8 months was high(85–90%) and not significantly affected by the removalof the apical meristem or 10%, 50% or even 90% of extantfoliage. In the understorey, survival was similarly highin all but the 90% removal treatment. Heavy defoliationkilled
c.
70% of understorey plants (Fig. 1a; chi-squaretest,
χ
2
= 72.56,
P
< 0.0001).Understorey and gap plants differed greatly in their
growth after damage, both in the magnitude and trendsof their responses (Table 1). In the understorey, growthwas negligible even in controls, and there was no effectof the removal treatments (Fig. 1b). In contrast, gapplants grew significantly in height and also respondeddramatically to damage. There was a clear stimulationof height growth after 10% defoliation, compared withcontrols. Even after 50% foliage loss, height growth wassimilar to that of undamaged controls. Only after 90%defoliation in gaps was height growth reduced to thelow level characteristic of understorey plants (Fig. 1b).Removal of the apical meristem had similar effects tomild (10%) defoliation, i.e. stimulation of growth in gaps(Fig. 1b).
The number of leaves on almost all juveniles (evencontrol plants) decreased over the 8-month experiment.The reduction in leaf number increased progressivelywith the severity of foliar damage in both gaps andunderstorey (Fig. 2a, Table 1). Total leaf number wasreduced by approximately one-third following 10%and 50% tissue removal, and by over one-half after90% removal (calculated from original data, not theresiduals shown in Fig. 2a). In the foliage removal
JEC581.fm Page 611 Saturday, July 14, 2001 1:35 PM
612
A. G. Blundell, & D. R. Peart
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 608–615
Table 1
Two-way
s testing the effects of simulated herbivory (treatments of 0%, 10%, 50% and 90% tissue removal, and apical meristem removal)and canopy cover (gap vs. understorey) on: (1) change in height (cm); (2) subsequent herbivore damage (percentage partial tissue loss) on the newest leafproduced during the experiment; (3) change in total number of leaves; (4) number of leaves retained; and (5) leaf production, over 8 months for
Shoreaquadrinervis
(Dipterocarpaceae) juveniles at Gunung Palung National Park, Indonesian West Borneo. Growth (column 1) controls for size by using theresidual of height growth vs. initial juvenile height (cm), and leaf dynamics (columns 3–5) control for juvenile size by using the residuals of linear regressionof the variable vs. the number of leaves on the juvenile at the start of the experiment
Dependent variable
(1) Height (2) Herbivory (3) Leaf number (4) Leaf retention (5) Leaf production
Source d.f.
F
-ratio
P F
-ratio
P F
-ratio
P F
-ratio
P F
-ratio
P
Herbivory level 2 7.4 < 0.0001 0.85 0.49 35.8 < 0.0001 9.20 0.0002 9.46 < 0.0001Canopy cover 1 104.0 < 0.0001 0.65 0.42 6.7 0.010 1.53 0.22 33.98 < 0.0001Herbivory
×
canopy 2 9.0 < 0.0001 0.22 0.93 7.4 < 0.0001 4.26 0.015 3.98 0.02
Fig. 1
Effects of simulated herbivory on performance (survival and growth) of
Shorea quadrinervis
(Dipterocarpaceae) juveniles(< 1 cm d.b.h.). Experiments ran over 8 months in gaps (open boxes, thick line) and in the understorey (closed boxes, thin line)at Gunung Palung National Park, Kalimantan Barat, Indonesian West Borneo. (a) Survival. Error bars represent
±
one standarddeviation (SD for binomial distribution = [ (probability of mortality
×
probability of survival)/no. of juveniles in the treatment] ).(b) Change in height; plotted as residuals (means
±
1 SE) of change in height vs. initial height. Different letters indicate significantdifferences (Tukey means comparison tests).
Fig. 2
Effects of simulated herbivory on leaf dynamics of
Shorea quadrinervis
(Dipterocarpaceae) juveniles (< 1 cm d.b.h.).Experiments ran over 8 months in gaps (open boxes, thick line) and in the understorey (closed boxes, thin line) at Gunung PalungNational Park, Kalimantan Barat, Indonesian West Borneo. (a) Residuals of change in number of leaves vs. initial number of leaves.(b) Residuals of number of leaves retained vs. initial number of leaves. (c) Residuals of number of leaves produced vs. initial numberof leaves. Values shown are means
±
1 SE. Different letters indicate significant differences (Tukey means comparison tests).
JEC581.fm Page 612 Saturday, July 14, 2001 1:35 PM
613
Effects of herbivory and gaps on juvenile performance
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 608–615
treatments, net leaf loss tended to be greater in gapsthan in understorey plants at the same level of damage;this difference was significant for 10% and 50% foliageremoval (Fig. 2a, Table 1). In the meristem removaltreatment, net leaf loss was significantly greater in theunderstorey.
The clear and consistent patterns in net leaf loss area consequence of more complex underlying patternsin leaf production and leaf retention (Fig. 2b,c). Pro-duction and retention showed opposite trends whentheir responses to gap and understorey environmentswere compared. With low to moderate levels of foliageremoval, leaf retention was much greater in the under-storey than in gaps, while the reverse was true for leafproduction. In the understorey, the effects of severe(90%) defoliation were manifested in sharply reducedleaf retention, in the minority of plants that sur-vived (Fig. 2b, Table 1). In gaps, the effects of severedefoliation were seen as an equally sharp decline in leafproduction (Fig. 2c, Table 1).
During the experiment, the youngest leaf on juvenilesincurred natural tissue loss to herbivores of 11
±
1.5%(mean
±
1 SE). This value was not significantly differ-ent between gap and understorey plants, and was notrelated to the amount of artificial herbivory (Table 1).
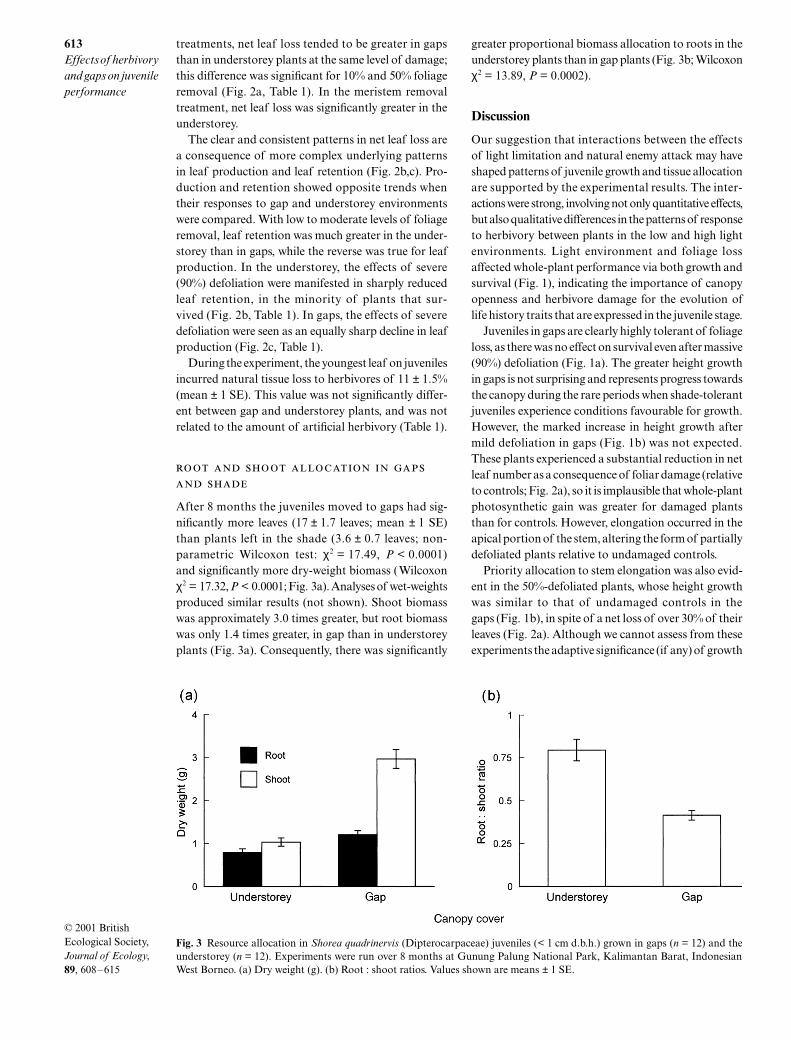
After 8 months the juveniles moved to gaps had sig-nificantly more leaves (17
±
1.7 leaves; mean
±
1 SE)than plants left in the shade (3.6
±
0.7 leaves; non-parametric Wilcoxon test:
χ
2
= 17.49,
P
< 0.0001)and significantly more dry-weight biomass (Wilcoxon
χ
2
= 17.32,
P
< 0.0001; Fig. 3a). Analyses of wet-weightsproduced similar results (not shown). Shoot biomasswas approximately 3.0 times greater, but root biomasswas only 1.4 times greater, in gap than in understoreyplants (Fig. 3a). Consequently, there was significantly
greater proportional biomass allocation to roots in theunderstorey plants than in gap plants (Fig. 3b; Wilcoxon
χ
2
= 13.89,
P
= 0.0002).
Discussion
Our suggestion that interactions between the effectsof light limitation and natural enemy attack may haveshaped patterns of juvenile growth and tissue allocationare supported by the experimental results. The inter-actions were strong, involving not only quantitative effects,but also qualitative differences in the patterns of responseto herbivory between plants in the low and high lightenvironments. Light environment and foliage lossaffected whole-plant performance via both growth andsurvival (Fig. 1), indicating the importance of canopyopenness and herbivore damage for the evolution oflife history traits that are expressed in the juvenile stage.
Juveniles in gaps are clearly highly tolerant of foliageloss, as there was no effect on survival even after massive(90%) defoliation (Fig. 1a). The greater height growthin gaps is not surprising and represents progress towardsthe canopy during the rare periods when shade-tolerantjuveniles experience conditions favourable for growth.However, the marked increase in height growth aftermild defoliation in gaps (Fig. 1b) was not expected.These plants experienced a substantial reduction in netleaf number as a consequence of foliar damage (relativeto controls; Fig. 2a), so it is implausible that whole-plantphotosynthetic gain was greater for damaged plantsthan for controls. However, elongation occurred in theapical portion of the stem, altering the form of partiallydefoliated plants relative to undamaged controls.
Priority allocation to stem elongation was also evid-ent in the 50%-defoliated plants, whose height growthwas similar to that of undamaged controls in thegaps (Fig. 1b), in spite of a net loss of over 30% of theirleaves (Fig. 2a). Although we cannot assess from theseexperiments the adaptive significance (if any) of growth
Fig. 3 Resource allocation in Shorea quadrinervis (Dipterocarpaceae) juveniles (< 1 cm d.b.h.) grown in gaps (n = 12) and theunderstorey (n = 12). Experiments were run over 8 months at Gunung Palung National Park, Kalimantan Barat, IndonesianWest Borneo. (a) Dry weight (g). (b) Root : shoot ratios. Values shown are means ± 1 SE.
JEC581.fm Page 613 Saturday, July 14, 2001 1:35 PM

614
A. G. Blundell, & D. R. Peart
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 608–615
of gap plants in response to damage, it is reasonable toassume that replacement of at least some of the losttissue is necessary to translate the high light-availabilityin gaps into growth. As the gap vegetation increasesin height and fills the canopy opening, the new foliageis most effectively placed if it is as high as possiblerelative to that of neighbours, thus requiring allocationof resources to stem elongation. This argument appliesequally to undamaged plants, but there may be additionalbenefits of enhanced height growth after damage if itplaces new foliage out of reach of herbivores, such asground-based vertebrates. While we have not documentedsignificant damage by vertebrates (see Introduction),vertebrate herbivores are present at the study site(Blundell 1996) and may have occurred at higherdensities in the past.
Gap plants did not grow significantly after extreme(90%) defoliation, but even this severe treatment did notincrease mortality (Fig. 1a). The ability to maintainessential physiological processes even after heavydefoliation and major net leaf loss must be due to highrates of carbon gain per unit leaf area in gap plants. Thecombination of high light-availability and the highpriority for resource allocation to above-ground tissuesin gaps help to explain the observed patterns of leafdynamics. When a juvenile encounters a gap created bythe death of a canopy tree, old ‘shade’ leaves should bereplaced rapidly with new ones that are physiologicallyadapted to high light (Bazzaz & Carlson 1982). Youngerfoliage generally has higher photosynthetic rates (Aide1993), and the placement of new leaves at the greatestpossible height increases the differential betweenphotosynthetic gain in old vs. new leaves. Consequently,where resources are available for leaf replacement,rapid leaf turnover should increase net carbon gain.In the experiments, gap plants discarded many more ofthe damaged leaves than did shaded plants and (exceptin the 90% defoliation treatments) gap plants maintainedmuch higher rates of leaf production (Fig. 2c).
The combination of field survey data (Blundell 1999)and the experiments reported here indicate that the riskof mortality following herbivore damage is significantfor this species. While 90% foliage removal is uncommon,such high levels of loss are occasionally encountered inunderstorey individuals in the field. Further, the experi-mental results (
c.
70% of understorey plants suffering90% foliage died within 8 months) suggest that mostindividuals that experience a single severe defoliationevent would probably die before they could be observedin a static field survey. Patterns of leaf dynamics in thesurvivors of heavy defoliation (Fig. 2) suggest that theywere also on a trajectory towards death; high leafretention characteristic of lightly damaged understoreyplants could not be maintained when juveniles wereseverely defoliated. This, combined with their low leaf-production rate, makes maintenance of essential func-tions unlikely.
In summary, severe defoliation events occur, probablymore commonly than is generally suspected, and few, if
any, understorey juveniles can survive such an attack.Repeated attacks, even if they inflict little foliage loss perattack, are also likely to increase mortality risk. Overdecades, the cumulative risk imposed by herbivoredamage must be considerable, and the overall riskassociated with damage in the understorey of tropicalrain forest is further increased by chronic exposure tofalling debris (Clark & Clark 1989).
Understorey plants are presented with very differentchallenges from those in gaps. Traits conferring highfitness in juveniles should therefore be expressed differ-ently in the two environments (Kitajima 1994) and theexpression must change rapidly when gaps are created.Resource availability has contributed much to thetheory of plant anti-herbivore defences (Coley
et al
. 1985).However, the implications of herbivory for the evolu-tion of resource allocation strategies in shade-tolerantplants have been largely ignored, probably because theeffects of natural enemies are much more cryptic thanthe more obvious, and clearly demonstrated, effectsof light availability on photosynthesis and growth. Thefindings reported here show that the interactions betweenlight availability and herbivore damage can provide thebasis for a more synthetic and ecologically rich theoryof resource allocation and growth strategies.
Acknowledgements
We thank L. Curran, M. Leighton and Cam Webb forinvaluable assistance at Cabang Panti, the IndonesianInstitute of Science (LIPI, Jakarta and Bogor, especiallyDr Dedy Darnaedi), the Conservation Agency of theDepartment of Forestry (PHPA) and KSDA Kalbarfor their assistance and access to Taman NasionalGunung Palung, Republik Indonesia. Usdik, Kaslan,Emily Harwell, Tim Laman and Cam Webb assisted inthe field, and Matt Ayres, Carol Folt, Lindsay Haddon,Matt Landis, Pete Marra, Miguel Martínez-Ramos,Mark McPeek, Scott Sillett, Cam Webb, Joe Wright andScot Zens commented on the manuscript. We especiallythank James Dykes and Scot Zens for statistical help.For financial assistance, we thank the US Departmentof State (Fulbright Award), National GeographicSociety, the Conservation, Food and Health Foundation,Inc., Sigma Xi, and the Explorer’s Club. Note: The viewsexpressed are the authors’ own and do not necessarilyrepresent official EPA policy.
References
Aide, T.M. (1993) Patterns of leaf development and herbivoryin a tropical understorey community.
Ecology
,
74
, 455–466.Ashton, P.S. (1982) Dipterocarpaceae.
Flora Malesiana
,Series I,
9
, 237–552.Ashton, P.S. (1988) Dipterocarp reproductive biology.
TropicalRain Forest Ecosystems B
(eds H. Lieth. & M.J.A. Wager),pp. 220–240. Elsevier Science, Amsterdam.
Bazzaz, F.A. & Carlson, R.W. (1982) Photosynthetic acclima-tion to variability in the light environment of early and latesuccessional plants.
Oecologia
,
54
, 313–316.
JEC581.fm Page 614 Saturday, July 14, 2001 1:35 PM

615
Effects of herbivory and gaps on juvenile performance
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 608–615
Becker, P. (1983) Effects of insect herbivory and artificial defoli-ation on survival of
Shorea
seedlings.
Tropical Rainforest:Ecology and Management
(eds S.L. Sutton, T.C. Whitmore& A.C. Chadwick), pp. 241–252. Blackwell Science, Oxford.
Bloom, A.J., Chapin,F.S. III & Mooney, H.A. (1985) Resourcelimitation in plants – an economic analogy.
Annual Reviewof Ecology and Systematics
, 16, 363–392.Blundell, A.G. (1996) A preliminary checklist of mammals at
Cabang Panti Research Station, Gunung Palung NationalPark, West Kalimantan. Tropical Biodiversity, 3 (3), 251–259.
Blundell, A.G. (1999) Density-dependent dynamics in a tropicalrain forest canopy tree. PhD thesis, Dartmouth College,Hanover, New Hampshire.
Canham, C.D. (1988) An index for understorey light levels inand around canopy gaps. Ecology, 69, 1634–1638.
Canham, C.D., Kobe, R., Latty, E. & Chazdon, R. (1999)Interspecific and intraspecific variation in tree seedlingsurvival: effects of allocation to roots versus carbohydratereserves. Oecologia, 121, 1–11.
Canham, C.D., McAninch, J.B. & Wood, D.M. (1994) Effectsof the frequency, timing, and intensity of simulated brows-ing on growth and mortality of tree seedlings. CanadianJournal of Forestry Research, 24, 817–825.
Chapin, F.S. III, Schulze, E.-D. & Mooney, H.A. (1990) Theecology and economics of storage in plants. Annual Reviewof Ecology and Systematics, 21, 423–447.
Clark, D.B. & Clark, D.A. (1985) Seedling dynamics of atropical tree: impacts of herbivory and meristem damage.Ecology, 66 (6), 1884–1892.
Clark, D.B. & Clark, D.A. (1989) The role of physical damagein the seedling mortality regime of a neotropical rain forest.Oikos, 55, 225–230.
Coley, P.D. & Barone, J.A. (1996) Herbivory and plantdefenses in tropical forests. Annual Review of Ecology andSystematics, 27, 305–335.
Coley, P.D., Bryant, J.P. & Chapin, F.S. III (1985) Resourceavailability and plant anti-herbivore defense. Science, 230,895–899.
Crawley, R.J. (1989) Insect herbivores and plant populationdynamics. Annual Review of Entomology, 34, 531–564.
Curran, L.M. & Leighton, M. (2000) Vertebrate responses tospatiotemporal variation in seed production of mast-fruitingDipterocarpaceae. Ecological Monographs, 70, 101–128.
Denslow, J.S. (1980) Gap partitioning among tropical rain-forest trees. Biotropica, (Suppl.) 12, 47–55.
Dominguez, C.A. & Dirzo, R. (1994) Effects of defoliation onErythroxylum havenensis: a tropical proleptic species.Ecology, 75, 1896–1902.
Hartshorn, G.S. (1978) Treefalls and tropical forest dynamics.Tropical Trees as Living Systems (eds P.B. Tomlinson &M.H. Zimmerman). Cambridge University Press, Cambridge.
Howe, H.F. (1990) Survival and growth of juvenile Virolasurinamensis in Panama: effects of herbivory and canopyclosure. Journal of Tropical Ecology, 6, 259–280.
Humphrey, J.W. & Swaine, M.D. (1997) Factors affecting thenatural regeneration of Quercus in Scottish oakwoods. 2.Insect defoliation of trees and seedlings. Journal of AppliedEcology, 34 (3), 585–593.
Kikuzawa, K. (1995) The basis for variation in leaf longevityof plants. Vegetatio, 121, 89–100.
Kitajima, K. (1994) Relative importance of photosynthetictraits and allocation patterns as correlates of seedling shadetolerance of 13 tropical trees. Oecologia, 98 (3–4), 419– 428.
Kobe, R.K. (1997) Carbohydrate allocation to storage as a
basis of interspecific variation in sapling survivorship andgrowth. Oikos, 80, 226–233.
Kozlowski, T.T. & Pallardy, S.G. (1997) Physiology of WoodyPlants, 2nd edn. Academic Press, San Diego.
Lowman, M.D. (1984) An assessment of techniques formeasuring herbivory: is rainforest defoliation more intensethan we thought? Biotropica, 16 (4), 264–268.
Marquis, R.J. (1984) Leaf herbivores decrease fitness of atropical plant. Science, 226, 537–539.
Marquis, R.J. (1992) A bite is a bite is a bite? Constraints onresponse to folivory in Piper arieianum (Piperaceae). Ecology,73 (1), 143–152.
Martínez-Ramos, M., Alvarez-Buylla, E. & Sarukhán, J.(1989) Tree demography and gap dynamics in a tropicalrain forest. Ecology, 70 (3), 555–558.
Martínez-Ramos, M., Sarukhán, J. & Piñero, D. (1988) Thedemography of tropical trees in the context of forest gapdynamics: the case of Asytrocaryum mexicanum at LosTuxtlas tropical rain forest. Plant Population Ecology (edsA.J. Davy, M.J. Hutchings, A.R. Watkinson), pp. 293–313.Blackwell Science, Oxford.
Núñez-Farfán, J. & Dirzo, R. (1991) Effects of defoliation onthe saplings of a gap-colonizing neotropical tree. Journal ofVegetation Science, 2, 459–464.
Osunkjoya, O.O., Ash, J.E., Graham, A.W. & Hopkins, M.S.(1993) Growth of tree seedlings in tropical rain forests ofNorth Queensland, Australia. Journal of Tropical Ecology,9, 1–18.
Platt, W.J. & Strong, D.R. (1989) Gaps in forest ecology. Eco-logy, 70 (3), 535.
Saunders, M.R. & Puettmann, K.J. (1999) Effects of overstoryand understorey competition and simulated herbivory ongrowth and survival of white pine seedlings. CanadianJournal of Forest Research, 295, 536–546.
Swaine, M.D. (1996) The Ecology of Tropical Forest Seedlings.UNESCO and The Parthenon Publishing Group, Paris.
Swaine, M.D. & Whitmore, T.C. (1988) On the definition ofecological species groups in tropical rain forests. Vegetatio,75, 81–86.
Terborgh, J. & Wright, S.J. (1994) Effects of mammalianherbivores on plant recruitment in two neotropical forests.Ecology, 75 (6), 1829–1833.
Tilman, D. (1988) Plant Strategies and the Dynamics andStructure of Plant Communities. Princeton UniversityPress, Princeton, New Jersey.
Uhl, C., Clark, K., Dezzeo, N. & Maquirino, P. (1988) Vege-tation dynamics in Amazonian treefall gaps. Ecology, 69(3), 751–763.
Webb, C.O. (1997) Seedling ecology and tree diversity in aBornean rain forest. PhD thesis, Dartmouth College,Hanover, New Hampshire.
Webb, C.O. & Curran, L.M. (1996) A field key to dipterocarpseedlings of the Gunung Palung National Park, WestKalimantan, Indonesia. Tropical Biodiversity, 3, 193–225.
Webb, C.O. & Peart, D.R. (1999) Seedling density dependencepromotes coexistence of Bornean rain forest trees. Ecology,80 (6), 2006–2017.
Whitmore, T.C. & Brown, N.D. (1996) Dipterocarp seedlinggrowth in rain forest canopy gaps during six and a half years.Philosophical Transactions of the Royal Society of LondonB, 351, 1195–1203.
Received 27 May 1999 revision accepted 18 December 2000
JEC581.fm Page 615 Saturday, July 14, 2001 1:35 PM





























