Embed Size (px)
Citation preview

Plant Ph-siol. (1967) 42, 946-952
Growth and Synthesis of Nucleic Acid and Proteinby Excised Radish Cotyledons1
R. H. Nieman and L. L. PoulsenUnited States Salinity Laboratory, Riverside, California 92052
Received March 20, 1967.
Summtiary. Nutritional and light requireiments for growth and synthesis of RNA,DNA, and protein by cotyledons excised from 5-day-old seedlings of Rthapiwnssativits L. were investigated, and the course of synthesis was followed through thecell cycle. The minimum requirements for a net increase in nucleic acid and proteinwere sugar, nitrate, and ligh;t. The cotyledons used nitrite at low concentratioll,buit not ammonium ion. Light was required for preliminary steps in synthesis ofRNA, DNA, and protein, but the actual polymerization reactions occurredI in thedark. The cotyledons contained stufficient endogenouis growth factors for abouthalf of the cells to complete 1 cycle on a medium of 1 % sucrose, 80 mMI KNO3.The increase in DNA was limited to about 50 % and was accompanied by a com-parable increase in cell number. Fresh weight, RNA, and protein tende-d to increasein proportion to DNA. Growth of the isolated cotyledons commenced with cellenlarge,ment. RNA began to increase after about 4 hours, DNA after abouit 12.The major in,crease in protein also began at about 12 hoturs. The maximum rate ofincrease for all 3 occurred between 12 and 16 hours. Cell counts indicated that by28 hours most of the cells which had replicated DNA had also completed cell division.
Excised leaf tisstie offers certaiin advantagesfor the stutdy of ce-lluilar processes responsible forleaf growth and of the effect of environmentalstress suich as salinitv and drouight on theseprocesses. Ideally, the tissue shoul.d be capable ofgrowth by both cell enlargement and cell divisionon a simple, (lefined medium, and it would beadvantageous if division could be induiced or pre-vente,d at will. Earlier experiments (7) indicatedthat the cotyledonary leaves of yotng radish seed-lings came close to meeting these requiirements.This paper gives the results of stuidies on thenutri'tional and light requirements of this tissuieand on the couirse of RNA, DNA, and proteinsynthesis throuigh the cell cycle.
Materials and Methods
Seedling Culture. Selectedl seeds of Raphanutssatiztus L., Cherry Bell, were germinated in pansof washed, expanded vermicuilite at 25 + 10 with1000 ft-c constant white light (fluorescent-plus-ttungsten lamps). The cuilttires were irrigated withdeionized water. At the desired seedling age (5clays, for most experiments), a number of cotyle-
1 Contribuitioni from the United States Salinitv Lab-oratory, Soil and Water Conservation Research Division,ARS, USDA, Riverside, California, in cooperationiNviththe 17 Western States and Hawaii.
946
dons were meastured to get an estimate of the meanwidth. Those within 1 mm of the mean, as judgedby comparison wiith a template, were excisecl andplaced directly into a htumid cham,ber. They wereweighed in grotups of 20, and these grouips werekept intact through all subsequenit manipulations.
The cotyledons varied with different seedbatches. In general, RNA, protein, and, to alesser extent, DNA decline,d with seed cage. Hence,only seed from cturrent crops was uised. The freshweight of the cotyledons also varied wx ith the rela-tive humidity in the growth room, which was notcontrolled. An attempt was made to minimize thiseffect by covering the pans with perforatecl clearplastic. This increased the fresh weight but italso increased the incidence of microbial contami-nation and decreased seedling vigor, so the practicewas dliscontinued.
Incubation. The cotyledlons wxere surface-disin-fected by shaking them for 30 seconds in a 2 %aqtueouis solution of neutral detergenit. followed bya thorough rinsing with sterile distilled\water.Several groups were frozen at this stIge, to serveas pre-incuibation controls. The rest were trans-ferred aseptically in difflise light to -sterile 9-cmpetri dishes, 1 group per (lish. The cotyledonswere supported, adaxial side tip, on a sterile padconsisting of 4 layers of filter paper and( 1 ofwashed cheese-cloth. The pads were moistenedwith 10 ml of sterile medium. Each group) wvasplaced in the (lark immediately- after transfer- and
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

NIEMAN AND POULSEN-RADISH COTYLEDONS
kept there until all had been transferred. Transfertook about 2 minutes per group. The dishes withcotyledons were placed in a forced-air incubator,either in the dark or under glass exposed tofluorescent-plus-tungsten lamps. The air tempera-ture beside the dishes was 25 ± 10. Rarely wasthere any sign of microbial contamination. WVhereit was suspected for any reason, the culture wasdiscarded.
The medium of Scott and Liverman (9) wasused in the first series of experiments. It con-tained 2 % sucrose plus the following salts: 80mM KNO3; 18 mnm sodium potassium tartrate; 7.4mm KH.,PO4; 0.3 mmt Co(N03,) 2. The pH was5.6. This medium was modified as described inthe text as the requirements of the cotyledons wereascertained.
After incubation, the cotyledons were blotted,weighed, and frozen.
Anallysis. RNXA and DNA were extracted anddetermined spectrophotometrically as described pre-viously (8). The coefficients used for a 1-cmlight path were: RNA pxg/ml = 31 X OD,,,, DNA/Ag/ml = 34(0D265-O0D30). The protein extractedalong with the nucleic acids in 0.3 N NaOH wasdetermined by the biuret reaction (1). Cell countswere made, following a pro-cedure suggested by R.Klein (2 and personal communication). Twentycotyledons were macerated for 24 houirs in 30 mlof 1 N HCl containing 5 % chromic acid. Themacerating solution was gently decanted and re-placed with several changes of distilled water.The cells were separated by shaking the cotvledonsin distilled water, followed by several passes of theresulting stuspension through a hypodermic syringewith a No. 20 needle. This procedture gave goodcell separation with very little breakage. Longerperiods of maceration increased cell breakage. Thecells were stained lightly with fast green, thesuspens.ion made to 100 ml with water, and the cellscouinte(l in a plankton counting chamber.
Results
Twenty cotyledons from ungerminated seed con-tained, on the average, 30 jug of DNA, 400 jg of
RNA, and from 6 to 10 mg of protein. Theamounts of DNA and RNA about doubled duringthe first 4 days of germination in the light ondeionized water (table I, zero incubation time).Under these conditions, there was no increase inDNA and RNA and little or no increase in proteinafter the fourth day. Crowth stopped after thesixth day. Cotyledons which were excised on thefourth, fifth, and sixth days and inctubated in thelight on Scott-Liverman medium increased theirfresh weight, DNA, and RNA. The 5-day-oldcotyledons appeared to be the most active and,hence, were used at this age in all subsequentexperiments.
Nutritional Requirements. The results of aseries of experiments undertaken to ascertain thenutritional requirements of the isol,ated cotyledonsare summarized in table II. The cotyledons re-quire sugar and a nitrogen source for net synthesisof RNA, DNA, and protein. Omission of nitratefrom the Scott-Liverman medium caused a loss ofall 3; omission of sucrose prevented lnet synthesisof DNA and protein but caused a loss only ofR'NA. Assuming that little or no cell divisionoccurred in the absence of net DNA synthesis,omission of either nitrate or sucrose from themedium limited growth to cell enlargement. Cobalt,which promotes the expansion of leaf disks undersome conditions (6,9), had little or nio effect onthe gain in fresh weight of the cotyledons andactually appeared to suppress synthesis of RNA,DNA, and protein. Reducing the sucrose concen-tration from 2 to 1 % had a slight beneficial effect.A suicrose solution alone, like distilled water, gaveonly a small increase in fresh weight with no in-crease in RNA, DNA, or protein. It appeared, infact, to promote the loss of DNA anid protein.Sucrose plus KNO3. on the other hand, gave astubstantial increase in fresh weight, RNA, DNA,and protein, but only in the light. Growth in thedark evidently was restricted to cell enlargementand was approximately equal to that obtained inthe light on Scott-Liverman medium without nitrateor on sucrose-KCl. NaN'0, was about as effectiveas KN03 in promoting growth and the synthesisof RNA and DNA. The potassium salt, however,
Table I. Effects of Seedling Age and Incubation after Excisioni on Fresh Weight, DNA, RNA, anid Proteini ofCotyledons
Seed'ings were grown on vermiculite Nx ith deionized ix-ater. Four groups of cotyledons were excised on the daysindicated; 2 -vere analyzed directly, 2 -%vere incubated for 28 hours (Scott-Liverman medium, 250, 800 Xl-c of light)and tlhen analx zed. Each value is the average of 2 groups.
Seedl ing Incubationiage time
da-s hr4
5
'5
0280
280
28
Frwt
DNA RNAper 20 cotyledons
mg3097564398900,4949
516751734768
0961.340.761.430 701.26
Protein
6.1
6.4
6.8
947
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
gave a larg-er increase in protein, possibly indi-cating a requirement for potassium in protein syn-thesis (4). The addition of KCI or NaCl to a
suicrose solution increased cell enlargement butfailed to promote net synthesis of nucleic acid andprotein. The small gain in fresh- weight with(NH4)2SI04, no greater than that obtained withdistilled water alone, suggests that this salt was
not taken up by the cotyledon cells. The cotyle-dons were able to uise nitrite at a reduced concen-
tration buit it became inhibitory at 80 mm.In other experiments (not shown), growth was
measured simply as a gain in fresh weight on 1 %suicrose containing variouis nitrogen sources, all atpH 6. At a concentration of 40 mm, nitrite suip-
ported growth as well as nitrate; glycine gave a
small amouint of growth, urea essentially none, andhydroxylamine was lethal.
The results in table II suiggest that growth andthe synthesis of nucleic acid and protein were in-
fluienced by light intensity. [This effect was ex-
amined in a later experiment (see table V).]Nutritiontal Status and Grozwth Potenttial. The
preceding experiments used cotyledons from seed-lings germinated with water. If the seedlings were
irrigated with nutrient solution, the cotyledons at5 days were from 20 to 30 % larger, and thequiestion arose whether the larger cotyledons mightnot also have a greater potential for growth afterisolation. They might, for example, contain a moreactive nitrate reductase or a greater reserve ofnitrogen precuirsors for the synthesis of nucleicacid and protein. To answer this question, severalhuindred seedlings were germinated in vermiculite
irrigated with nutrient solution containilng: 0.5 mmC,a(-N03)2, 0.6 mAi KNO3, 0.3 mM :MgSO4, and0.1 mms KH,PO,. Control seedlings were irrigatedwith water. Cotyledons were excised at 5 days;some were analyzed directly; others were incubatedfor 28 hours on 1 %,sucrose or sucrose-plus-KNO3.
As a second part of the experiment, growth ofcotyledons on nutrient-irrigated seedlings was com-
pared with the growth of isolated cotylefdons on
sucrose-KNO3. At the same time that cotyledonswere excised for the inicubation studies, others ofthe same size were tagged with string an,d left on
the seedlings. The intact plants were kept for 28hours under the same light and temperatuire condi-tions as the isolated cotyledons. At the encd ofthis time, the cotyledons were excised, weighed,and analyzed along with those which ha(l beeninctibated in petri dishes.
Irrigation with nutrient soluition increasedl thefresh weight and RNA, DNA, and protein contentof the cotyledons at 5 days (table III), butt itdid not increase their potential for growth andsynthesis on the suicrose-KNO3 mecliuim. Further,it did not appear to affect the requirement for an
exogenous supply of nitrate or for light. Cotyle-dons of seedlings given nuitrient soltution evidentlyfailed to store any nitrogen comipouindls which couildbe uised for net synthesis of nuicleic acid and pro-
tein after they were excised.Cotyledons of intact seedlings were similar to
excised cotyledons with respect to the increase infresh weight and DNA btut were considerably more
active in synthesizing RNA and proteini.Timiie Couirse I. A time-couirse experiment was
Table II. Influenice of M1edium Comiipositioni and Light on the Percent Chanigc in Freslh Weight, D.,VI, RNA, andProtein of Cotyledons durina Inculatiion
Duiplicate groups of cotyledons were incubated for 28 hours at 250 on each of the differenit media. The bufferwas 5 mAi potassium phosphate (pH 6.5). The values indicate the average percent increase or decrease (-) duringincubationl.
Change during incubationLight Fr
Medium intensity Wt DNA RNA Protein-~~~~~~~~~~~~~~f-ft-c %0 % % %Scott-Livermani 800 112 23 38 46
KNO, 800 61 -18 -36 --18- Sucrose 800 99 0 -22 0
Co 800 106 51 67 53With 1 % suierose 800 138 31 39 51
H.,O 800 30 0 -28 71 % Sucrose 400 36 -12 -21 251 % Suicrose plus
80 nir KNO3 800 143 51 62 8280 nmi KNOX Dark 65 1 -2 --680 mui KNO3 400 119 27 38 4080 in Ai NaNO., 400 116 21 39 2780 n AiKCI 400 70 -2 -22 2780 inm NaCI 400 79 7 - 9 -2140 mM (NH4)9SO4 400 31 7 9 - 3Btuffer, 80 nmia NaNO. 1000 145 31 86 41Buffer, 80 niAm NaNO., 1000 54 12 17 9Buf fer, 60 noirA NaNO. 1000 101 17 54 25
GAR
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
949NIEMAN AND POULSEN-RADISH COTYLEDONS
Table III. Influence of Seedling Nuttrition and Excision on Growth ond Synthesis of DNA, RNA, and Proteinby Cotyledons
Seedlings were grown on vermiculite irrigated with either deionized water or nutrient solution. Cotyledons wereexcised at 5 days and incubated for 28- hours at 250 on 1-% sucrose, with or without 8& mm KNO3. The lightintensitv was 800 ft-c. Selected seedlings were left intact on the nutrient culture and kept for 28 hours underthese same light and temperature conditions. Cotyledons were then excised. weighed, and analyzed. All values areaverages of 2 groups.
Seedling Incubationculture condition Fr wt per 20 cotyledons Protein
mg ,ug mg mgBefore incubation
540 57 1.03 9.0Water After incubation
Sucrose, KNO3, light 1089 73 1.42 13.7Sucrose, KNO3, dark 921 59 1.06 8.5
Before incubationNutrient 693 62 1.19 10.7
solution After incubationSucrose, KNO3, light 1210 81 1.58 15.2Sucrose, KNO3, dark 1051 68 1.23 10.8Sucrose, --, light 859 62 1.07 10.3Sucrose, --, dark 808 61 0.96 8.6
Nutrient Intactsolution seedlings, light 1197 87 1.96 18.5
Table IV. The Chantge in Fr-esh Weight, DNA, RNA, and Protein of Cotyledons with TimeTwo groups of cotyledons were analyzed before and after incubation for the indicated time. Incubation conditions:
1 % sucrose in 80 mMi KNO3, 250, 800 ft-c white light. Each value is the mean and range for 2 grouips.
Fr wt DNA RNATime (X 102) Proteinhr per 20 cotyledons (X 10)
mg mg mg0 411 + 0 50 ± 2 68 ±+ 2 62 214 631 ± 9 53 ±+ 2 100 ± 1 67 026 819 2 66 ± 1 135 + 2 80 638 916 36 76 ± 3 132 ± 7 89 350 838 7 74 4 106 ± 6 85 262 872 ± 21 79 9 94 ± 3 86 4
performed to find out how long the isolated coty-ledons would grow and synthesize RNA, DNA, andprotein when supplied with nothing more thansucrose, KNO3, and light. The results (table IV)indicate that growth and synthesis terminatedwithin 38 hours. RNA, along with fresh weight,began to increase early in the incubation period.The RNA content reached a maximum within 26hours and appeared to remain constant for thenext 12 hours, then to decline when growth stopped.DNA and protein increased very little during thefirst 14 hours. Both increased during the next 24hours and then remained constant. The delay in
the synthesis of DNA and protein indicated by thisexperiment was reexamined with shorter timeintervals in a later experiiment (see figs 1-4).
Light Intensitv. The experimental results sum-
marized in table II stuggested that growth andsynthesis of nucleic acid and protein were influ-enced by light intensity. This effect was studied
by incubating cotyledons on sucrose-KN03 mediumfor 28 hours at 5 different light intensities (tableV). Intensities below 1000 ft-c were obtained by
Table V. Influence of Liqht Intensity on Fresh Wieight,DNA, RNA, and Protein of Cotyledons
Two groups of cotyledons were incubated at eachlight intensity for 28 hours at 250. The medium was1 % suicrose in 80 mM KNO3. Each value is the meanand range for 2 groups.
Fr wt DNA RNA ProteinLight (X 102) (X 10)
intensity per 20 cotyledons
ft-c mg ltg mg mg0 823+19 56±1 107+5 120+5
200 975±14 72±6 132±6 127±6400 1007±39 73+7 143±9 141±+5600 1035±28 74±4 160±4 146+41000 1103±24 74±4 174±6 150+5
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
interposing cheese-cloth screens betweeii the lampsand petri dishes. Fresh weight and RNA increasedwith light intensity, though not proportionally, over
the entire range. DNA and protein, on the otherhand, indicated little or no increase with lightintensitr above 200 and 40 ft-c, respectively.
pH Of the Medium. The infltuence of the pHof the meditum on growth and synthesis of nucleicacid and protein was examined by inctubating coty-leclons on sticrose-KNO3 meditum buffered at pH5.0, 5.5, 6.0, 6.5, and 7.0 with 5 mm potassiumphosphate. There was no clear pH effect in thisrange, btut phosphate appeared to be beneficial andso was made a regular constittuent of the meditum.
Tim e Course II. In a second time-course ex-
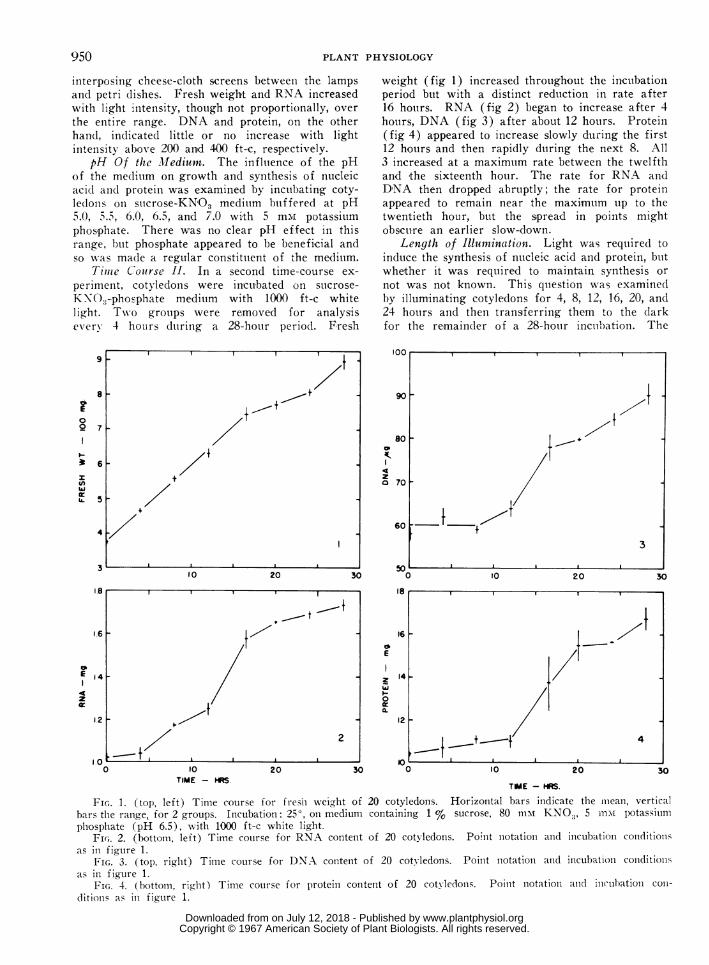
periment, coty)ledons were incubated on suicrose-
KN\O8-phosphate medium with 1000 ft-c whitelight. Tw o groups were removed for analysiseverv 4 hou: rs duiring a 28-hotur period. Fresh
9
r
0
0
I-
U
8
7
6
5
4
alE
4
10TIME - HtS.
20
weight (fig 1) increased throughout the incubationperiod but with a distinct reduction in rate after16 hoturs. RNA (fig 2) began to increase after 4hours, DNA (fig 3) after about 12 hours. Protein(fig 4) appeared to increase slowly during the first12 hours and then rapidly during the next 8. All3 increased at a maximum rate between the twelfthand the sixteenth hour. The rate for RNA andD,NA then dropped abruptly; the rate for protein
appeared to remain near the maximutm up to thetwentieth hour, but the spread in points mightobscure an earlier slow-down.
Length of Illumnination. Light was reqtuired toinduce the syn'thesis of nucleic acid and protein, btitwhether it was required to maintain synthesis or
not was not known. This qtuestion was examinedby illuminating cotyledons for 4, 8, 12, 16, 20, and24 hoturs and then transferring them to the clarkfor the remainder of a 28-hour inctubation. The
100
90 _
80 _ob
*I4
a 70 _
60
E
I0
30
50 L
O 10 20 30
4~ ~ ~
10 I I *
V ^^_3010TIE - HFS.
Fic.. 1. (top, left) Time course for fres-i weight of 20 cotyledons. Horizontal bars indicate the mlean, verticalbars the range, for 2 groups. Incubation: 250, on medium containing 1 % sucrose, 80 illm KNO.,, 5 mni potassiumphosphate (pH 6.5), wxith 10(X ft-c white light.
Fic,. 2. (bottom, left) Time course for RNA content of 20 cotyledons. Point niotation and incubation coniditioinsas in figure 1.
FIG. 3. (top, right) Time course for DNA content of 20 cotyledons. Point notationi and incubation conditionsas in figure 1.
FIG. 4. (bottom, righlt) Time course for protein content of 20 cotyledons. Point notation aind inctuhation coli-ditionis as in figure 1.
950
/+
/
/~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.[j+_t'I~~~~~3
10 20 30
.8 I I I I8~~.t
6 - r
4~~~~~~~~~~~4
2Z
nr - - f ,i I~~~~~~
1.~
1.1
Il
1.
0 20
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
951NIEMAN AND POULSEN-RADISH COTYLEDONS
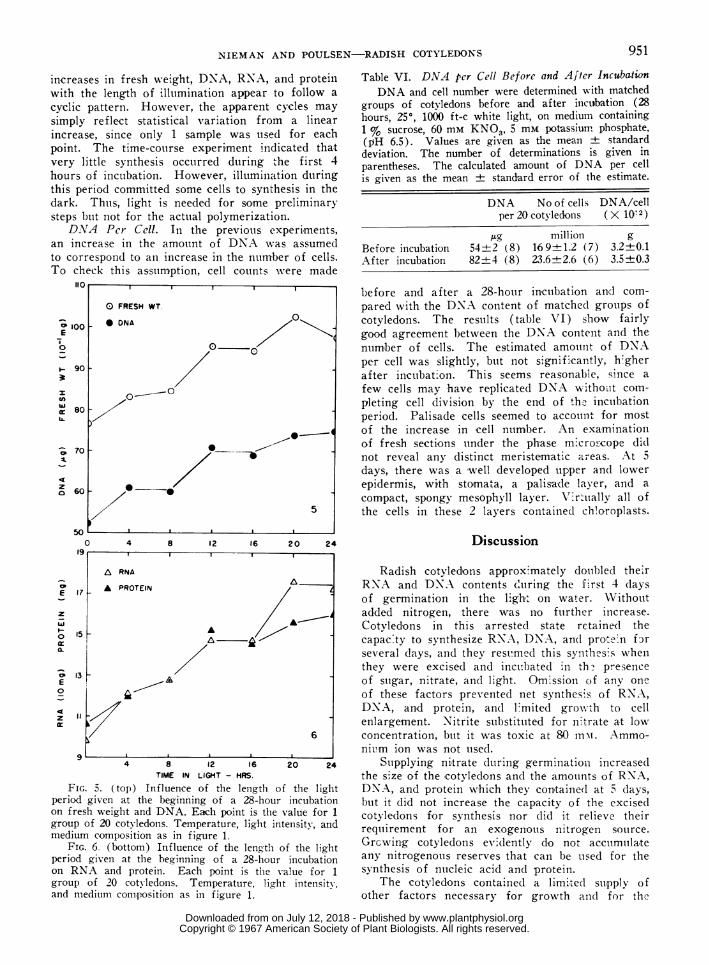
increases in fresh weight, DNA, RNA, and proteinwith the length of illumination appear to follow acyclic pattern. However, the apparent cycles maysimply reflect statistical variation from a linearincrease, since only 1 sample was used for eachpoint. The time-course experiment indicated thatvery little synthesis occurred during the first 4hours of inctubation. However, illumination duringthis period committed some cells to synthesis in thedark. Thus, light is needed for solme preliminarysteps buit not for the acttual polymerization.DNA Per Cell. In the previouis experiments,
an increase in the amount of DNA was assumedto correspond to ani increase in the ntumber of cells.To check this assuimption, cell cotunts were made
1101 I
E
0
I.-
a:cnIL
100
90
80
70
Cib4.
z
a 60 [
50
E
z
w
1.-
0
a.
E
0
4
z
4 8 12 16 20 24
4 8 12 16 20 24
TIME IN LIGHT HRS.
FIG. 5. (top) Influence of the length of the lightperiod given at the beginning of a 28-hour incubationon fresh weight and DNA. Earch point is the value for 1group of 20 cotyledons. Temperature, light intensity, andmedium composition as in figure 1.
FIG. 6. (bottom) Influence of the length of the lightperiod given at the beginning of a 28-hour incubationon RNA and protein. Each point is the alue for 1group of 20 cotyledons. Temperature, light intensity,and medium comiposition as in figure 1.
Table VI. DNA per Cell Before anid Al fer IncubationDNA and cell number were determined with matched
groups of cotyledons before and after inicuibation (28hours, 250, 1000 ft-c white light, on medium containing1 % sucrose, 60 mm KNO, 5 mm potassium. phosphate,(IpH 6.5). Values are given as the mean + standarddeviation. The number of determinations is given inparentheses. The calculated amount of DNA per cellis given as the mean ± standard error of the estimate.
DNA No of cells DNA/cellper 20 cotyledons (X 10' 2)
millioni gBefore incubation 54±2 (8) 169+1.2 (7) 3.2+0.1After incubation 82+4 (8) 23.6±2.6 (6) 3.5±+0.3
before and after a 28-hour inccubation and com-pared with the DN'A content of matched grouips ofcotyledons. The results (table VI) show fairlygood agreement between the DNA content and thenumber of cells. The estimated amount of DNAper cell was slightly, btut not significantly, h gherafter incubation. This seems reasonable, since afew cells may have replicated DNA withoiLt com-pleting cell division by the end of thc incuibationperiod. Palisade cells seemed to accoulnt for mostof the increase in cell number. An examinationof fresh sections uinder the phase microscope didnot reveal any distinct meristematic areas. At 5days, there was a -well developed upper an(d lowerepidermis, wiith stomata, a palisade layer, and acompact, spongy mesophyll layer. Vir tually all ofthe cells in these 2 layers contained chloroplasts.
Discussion
Radish cotyledons approximately douibled theirRNA and D,NA contents dluring the first 4 (laysof germination in the light on water. Withoutadded nitrogen, there was no fuirther increase.Cotyledons in this arrested state retained thecapac:ty to synthesize R.\A, DNA, andl protei,:n f:rseveral days, and they restmed this synthbs.s whenthey were excised and incubated in th- presenceof suigar, nitrate, and light. Omission of any oneof these factors prevented net synthesis of RNA,DNA, and protein, and l mited growth to cellenlargement. Nitrite suibstituited for nitrate at lowconcentration, buit it was toxic at 80 mari. Ammo-niuLm ion was not uised.
Supplying nitrate duiring germination increasedthe size of the cotyledons and the amouints of RNA,DNA, and protein which they containe,d at 5 days,but it did not increase the capacity of the excisedcotyledons for synthesis nor didl it reliev\e theirrequiirement for an exogenotus nitrogen soturce.Grcwing cotyledonis evidently do not accumulateany nitrogenous reserves that can be tused for thesynthesis of nucleic acid and protein.
The cotyledtons containedl a limited supply ofother factors necessary for growth andcl for the
0 FRESH WT.
- *DNA 0
0
00
0--b
0~~~ 0
5
-°-
%I
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
synithesis of nucleic acid and protein. This supplywas depleted within 38 hours when they wereexcised and incubated in the light on sucrose-KNO,medium. Growth under these conditions was lim-ited to a 50 % increase in cell numiber an,d a com-parable increase in DNA, protein, and average cellsize (fr wt/D,NA). RNA and total fresh weightabout doubled. Growth began with cell enlarge-ment. RNA began to increase after about 4 hours,DNA and protein after about 12 hours. All 3increasedl maximally between 12 and 16 hours. Therate o,f increase in fresh weight, RNA, and DNAdecreased sharply after the sixteenth hour, possiblymarking the beginning of mitosis. Protein syn-thesis may have begun to decline at about the sametime, definitely after the twentieth houir. After28 hours, most of the cells that had replicate(d DNAhad also completed cell division.
Light was required to initiate the synthesis ofRNA, DNA, andI proteini, bLut the actual polymer-ization reactions occurredl in the dark, in the caseof DNA and protein, some 8 hours after illumina-tion. The increase in DNA and in protein appearedto be independent of light intensity above about200 and 400 ft-c, respectively. RNA and freshweight, on the other hand, increased with lightintensity to at least 1000 ft-c.
The status of DNA replication in nitrogen-de-prived cotyledons is probably similar to that inbacterial (3,5) and protozoan (10) ce'lls deprivedof an essential amino acid. DNA replication begunbefore the deficiency (leveloped probably goes tocompletion, but no new cycles are started. Whenthe cotyledons were excised and supplied with sugarand KNO3, some cells were induced to replicateDNA by illuiminating them for 4 hours; othersrequired longer illumintation, or longer incubationbefore they were capable of responding to light.The amount of preparation required for inductionmay have been dleci,ded by the stage in the previouiscycle at which nitrogen became limiting.
Since, at most, only half of the cotyledon cellsreplicated DNA, it is entirely possible that non-replicating cells contributed to the increase in fresh
weight, RNA, and protein. In the long run, how-ever, fresh weight, RNA, and protein tended toincrease proportionally with DNA, which stuggeststhat they were increased by cells which also repli-cated DNA. In the second time-course experiment,for example, after DNA began to inicrease, thefresh weight, RNA, and protein per ,ug of DNAtended to remain constant at 10 mg, 20 jug, and185 ,ug, respectively.
Literature Cited
1. GORNALL, A. G., C. S. BARDAWILL, AND M. M.DAVID. 1949. Determination of serum proteinsby means of biuret reaction. J. Biol. Chem. 177:751-66.
2. KLEIN, R. M. AND J. WANSOR. 1963. Effects ofnon-ionizing radiation on expansion of disks fromleaves of dark-grow-n plants. Planit Physiol. 38:5-10.
3. LARK, K. G. 1966. Regulationi of clhromosomereplication and segregation in bacteria. Bacteriol.Rev. 30: 3-22.
4. LUBIN, M. AND H. L. ENNIS. 1964. In1tracellularpotassium in protein synthesis. Biollimil. Biophys.Acta 80: 614-31.
5. MVAALOE, 0. AND P. C. HANAWALT. 1961. Thy-mine deficiency and the normal DNA replicationcxcle. J. Mol. Biol. 3: 144-55.
6. MITLLER, C. 0. 1951. Proimotinig eflfect of cobal-tous and nickelous ions on expansion of etiolatedbean leaf disks. Arch. Biocihem. Biophys. 32:216-18.
7. NIEMAN, R. H. AND L. L. POULSEN. 1962. Thelight dependence of nucleic acid anid protein s5yn-thesis by isolated radish cotvledonarv leaves. PlantPh' 1siol. 37: xxi.
8. NIEMAN, R. H. AND L. L. POULSEN. 1963. Spec-trophotometric estimationi of nucleic aci l of Iplantleaves. Plant Phyvsiol. 38: 31-35.
9. SCOTT, R. A., JR. AND J. L. LIVERMAN. 19,56. Pro-motion of leaf expansion by kinetin anid benzinl-aminopurine. Plant Phxysiol. 31: 321-22.
10. STONE, G. E. AND D. -M. PRESCOTT. 1964. Celldivision and DNA synthesis in Tct:,-7avmcna pr-formis deprived of essential amino acids. J. CellBiol. 21: 275-81.
952
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.





























