Embed Size (px)
Citation preview

H+/ATP Ratio during ATP Hydrolysis by Mitochondria: Modification of the ChemiosmoticTheoryAuthor(s): Martin D. Brand and Albert L. LehningerSource: Proceedings of the National Academy of Sciences of the United States of America,Vol. 74, No. 5 (May, 1977), pp. 1955-1959Published by: National Academy of SciencesStable URL: http://www.jstor.org/stable/67144 .
Accessed: 08/05/2014 09:26
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
National Academy of Sciences is collaborating with JSTOR to digitize, preserve and extend access toProceedings of the National Academy of Sciences of the United States of America.
http://www.jstor.org
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 09:26:06 AMAll use subject to JSTOR Terms and Conditions

Proc. Natl. Acad. Sci. USA Vol. 74, No. 5, pp. 1955-1959, May 1977 Biochemistry
H+/ATP ratio during ATP hydrolysis by mitochondria: Modification of the chemiosmotic theory
(oxidative phosphorylation/proton transport/phosphate transport/adenine nucleotide transport/cation transport)
MARTIN D. BRAND* AND ALBERT L. LEHNINGERt
Department of Physiological Chemistry, The Johns Hopkins University, School of Medicine, 725 N. Wolfe Street, Baltimore, Maryland 21205
Contributed by Albert L. Lehninger, March 7, 1977
ABSTRACT The stoichiometry of H+ ejection by mito- chondria during hydrolysis of a small pulse of ATP (the H+/ATP ratio) has been reexamined in the light of our recent observation that the stoichiometry of H+ ejection during mitochondrial electron transport (the H+/site ratio) was previously underes- timated. We show that earlier estimates of the H+/ATP ratio in intact mitochondria were based upon an invalid correction for scalar H+ production and describe a modified method for determination of this ratio which utilizes mersalyl or N-ethyl- maleimide to prevent complicating transmembrane movements of phosphate and HW. This method gives a value for the H+/ATP ratio of 2.0 without the need for questionable corrections, compared with a value of 3.0 for the H+/site ratio also obtained by pulse methods.
A modified version of the chemiosmotic theory is presented, in which 3 H+ are ejected per pair of electrons traversing each energy-conserving site of the respiratory chain. Of these, 2 H+ return to the matrix through the ATPase to form ATP from ADP and phosphate, and 1 H+ returns through the combined action of the phosphate and adenine nucleotide exchange carriers of the inner membrane to allow the energy-requiring influx of Pi and ADP3- and efflux of ATP4-. Thus, up to one-third of the energy input into synthesis of extramitochondrial ATP may be required for transport work. Since other methods suggest that the H+/site significantly exceeds 3.0, an alternative possibility is that 4 H+ are ejected per site, followed by return of 3 H+ through the ATPase and 1 H+ through the operation of the proton-coupled membrane transport systems.
The passage of electrons from substrates to oxygen via the res- piratory chain can develop an electrochemical gradient of H+ across the inner membrane of intact mitochondria (1-4). The number of H+ equivalents ejected during passage of a pair of electrons through each of the energy-conserving sites is desig- nated the H+/site ratio (5). According to the chemiosmotic theory of oxidative phosphorylation (1-3, 6), the electro- chemical H+ gradient so produced may drive H+ back into the mitochondrial matrix through the mitochondrial ATPase, causing coupled synthesis of ATP. The number of H+ equiva- lents translocated inward per ATP synthesized is the H+/ATP ratio. Since there is no net production or utilization of H+ during oxidative phosphorylation in vivo, the chemiosmotic theory postulates that the H+/site and the H+/ATP ratios are equal.
The original measurements of the H+/site ratio by Mitchell and Moyle (7, 8) yielded the value 2.0 (see also ref. 9). Since measurements of the H+/ATP ratio during hydrolysis of added ATP also gave the value 2.0 in both mitochondria (7, 10-14) and submitochondrial particles (14, 15), the postulated condition of equality (H+/site = H+/ATP) was apparently fulfilled. However, we have recently demonstrated that the original oxygen-pulse experiments of Mitchell and Moyle (7, 8) un- derestimated the H+/site ratio; when interfering secondary H+
* Present address: Department of Biochemistry, University of Cam- bridge, Tennis Court Road, Cambridge CB2 1QW, U.K.
t To whom reprint requests should be addressed.
movements were suppressed, H+/site ratios of at least 3.0 were observed (5, 16-20), which are in agreement with thermody- namic data (9, 21-23).
In view of the new values of at least 3.0 for the H+/site ratio, two fundamental questions arise. Are the previously reported (7, 10-14) values of 2.0 for the H+/ATP ratio correct or have they also been underestimated? Second, is the equality of the H+/site and H+/ATP ratios postulated in the chemiosmotic theory necessarily valid? This paper describes a reexamination of the H+/ATP ratio in intact rat liver mitochondria with the ATP pulse method (12, 14), as well as the postulated equality of the H+/site and H+/ATP ratios.
EXPERIMENTAL DETAILS Rat liver mitochondria were isolated in 250 mM sucrose, washed three times, and resuspended in 250 mM sucrose to yield a stock suspension containing 50 mg of protein per ml.
ATP pulse experiments were carried out anaerobically in the apparatus previously described for oxygen pulse experiments (5, 19), consisting of pH and oxygen electrodes inserted into a 2-ml chamber with a restricted opening over which a stream of nitrogen was blown. Mitochondria (10 mg of protein) were incubated in 2.0 ml of an anaerobic medium of 120 mM KCI, 3 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (Hepes), 1 mM EDTA, 5 AM rotenone, 1 M antimycin A, 100 ng of valinomycin per mg of protein, and 25 ng of aurovertin at pH 7.10 and 28?. Valinomycin was present to prevent for- mation of transmembrane potentials by allowing K+ move- ments across the membrane, EDTA to chelate Mg2+ and thus inhibit adenylate kinase, and aurovertin to ensure rapid and complete hydrolysis of added ATP (13, 14, 24). At zero time 5 Al of 10 mM disodium ATP (50 nmol; 5 nmol/mg of protein), which had been brought to the same pH as the medium with KOH, was injected and the H+ ejection recorded. Buffer ca- pacity of the system was determined by titration with small amounts of 100 mM HCI. ATP, ADP, and AMP were assayed enzymically (25-27). A fresh solution of N-ethylmaleimide (Sigma Chemical Co., St. Louis, MO) was prepared daily. Au- rovertin was a gift of Dr. E. N. Moudrianakis.
RESULTS
Determination of H+/ATP ratio according to Moyle and Mitchell (14) The first set of measurements with the ATP pulse method was carried out exactly as described by Moyle and Mitchell (14) (see Fig. 1). The data were corrected for the back-decay of the H+ gradient and for the scalar H+ production exactly as they de- scribed. In a large number of experiments we observed a value of 2.00 + 0.09 (SD) for the corrected H+/ATP ratio, in close agreement with Mitchell and Moyle (7, 12-14). Addition of oligomycin in the presence or absence of proton-conducting
1955
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 09:26:06 AMAll use subject to JSTOR Terms and Conditions

1956 Biochemistry: Brand and Lehninger Proc. Natl. Acad. Sci. USA 74 (1977)
Moyle and Mitchell (1973) Present method
50 F
ng-ions
Plus FCCP | AH+
30 sec ATP ATP
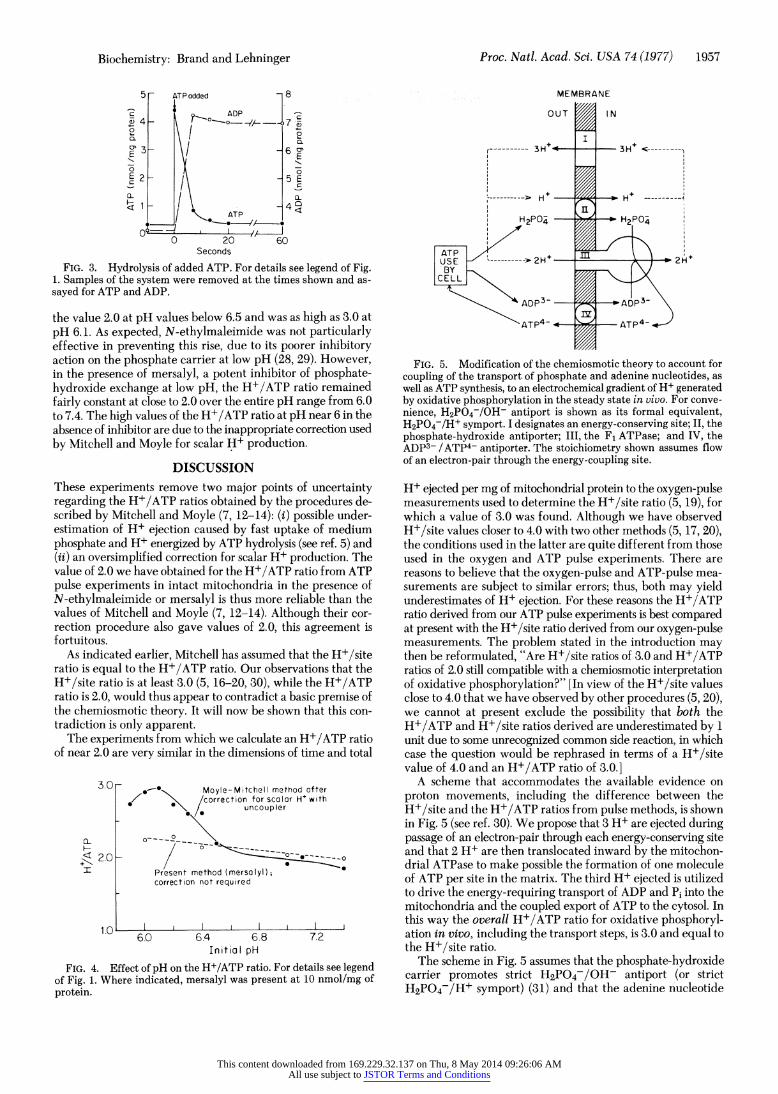
FIG. 1. Determination of vectorial H+ ejection (zH+v). For particulars see Experimental Details. H+ ejection (upwards) was initiated by addition of 5 nmol of ATP per mg of protein. Additions to the experimental medium, where shown, were N-ethylmaleimide, 40 nmol/mg of protein; carbonyl cyanide p-trifluoromethoxyphe- nylhydrazone (FCCP), 1 ,uM. (Left) In the procedure of Moyle and Mitchell (14) the apparent scalar H+ ejection (AH+,) is subtracted from the total H+ ejected to give the vectorial H+ ejection AH+v. This correction is invalid (see text). (Right) Both inward and outward movements of H2PO4- (and thus of OH- and scalar H+) were pre- vented by N-ethylmaleimide; no correction for scalar H+ is neces- sary.
uncouplers greatly inhibited ATP-induced H+ ejection; similar results were obtained with venturicidin and dicyclohexylcar- bodiimide.
However, ATP pulse experiments as carried out by Mitchell and Moyle (7, 12-14) are subject to two points of uncertainty. First, they can be expected to suffer froin the same type of er- rors and interference from secondary ion movements as the original oxygen pulse experiments of these authors (ref. 8; see refs. 5 and 19), since the general experimental conditions and maneuvers, as well as the magnitude of the observed H+ movements, are almost identical.
The second point of uncertainty is the correction applied by Mitchell and Moyle for the scalar H+ produced during the hydrolysis of ATP by the reaction
ATP4- + H20 > ADP3- + (1 - n)H2PO4
+ nHPO42- + nH+ [1]
where the value of n increases with pH above about 6.1. They estimated the scalar H+ production by carrying out a parallel ATP pulse in the presence of a proton-conducting uncoupling agent, which eliminates the H+ gradient generated by the ejection of the vectorial protons. The observed H+ formation in the presence of uncoupler was then subtracted from the total H+ ejected in the absence of uncoupler to give the vectorial component of the observed H+ ejection, from which their H+/ATP ratio was calculated. However, this correction pro- cedure is invalid, because the H+ produced by reaction 1, which occurs within the mitochondrial matrix, will not necessarily appear in the extramitochondrial medium in which the pH changes produced during the ATP pulse are actually monitored, unless, of course, some means of quantitative outward transport of the scalar H+ is available. In the absence of uncoupler the most probable route for outward transport of scalar H+ is by efflux with the H2PO4- generated in the matrix by ATP hy- drolysis, down the concentration gradient of phosphate, via the phosphate-hydroxide exchange carrier. H2PO4- reaching the extramitochondrial space in this manner will ionize and cause further H+ production in the medium. However, opposed to the efflux of scalar H+ by this path is the out > in electro- chemical gradient of HW, which increases as the added ATP is utilized. As a result, simple subtraction of the uncoupler-in- sensitive H+ ejection from the total H? ejection, as carried out
2.0 -2.0
E C: 1.9- C 1.9
- 5 E xtrapolIated 15~-F- ~ ~H~/ATP=1 93
<1. T_-
o,*
L 1.7-- 10
l l I
12 24 36 Seconds
FIG. 2. Determination of the H+/ATP ratio in the presence of N-ethylmaleimide (data of Fig. 1). The arrow represents the point at which the ATP hydrolysis was calculated to be half complete.
by Mitchell and Moyle (7, 12-14), oversimplifies a complex set of circumstances and cannot correct quantitatively for scalar H+ production. [This criticism does not apply to experiments on inverted inner membrane vesicles (14, 15), since in this case reaction 1 occurs in the medium phase; moreover, those ex- periments were carried out at a low pH, where the value of n is negligibly small. ]
Both of these uncertainties associated with ATP pulse ex- periments with intact mitochondria may be eliminated by in- hibiting the phosphate-hydroxide antiporter. This maneuver will (i) abolish the secondary inward movement of extramito- chondrial phosphate, and thus prevent coupled inward move- ment of H+ (5), (ii) abolish outward transport of phosphate, arising from ATP hydrolysis, on the phosphate-hydroxide an- tiporter, (iii) prevent scalar H+ from leaving the mitochondrial matrix coupled to efflux of H2PO4-, and (iv) also prevent, pari passu, the subsequent ionization of extramitochondrial H2PO4-. Under these conditions no correction will then be necessary for the appearance of scalar H+ in the medium. The total amount of H+ ejected during the ATP pulse can then be used directly to calculate the H+/ATP ratio.
Modified ATP pulse method
We found that the addition of N-ethylmaleimide or mersalyl to inhibit the phosphate-hydroxide exchange carrier (see refs. 19, 20, 28, and 29) diminished the overall observed H+ ejection in ATP pulse experiments, as expected from the reasoning outlined above. The rapid ejection of H+ was followed by monophasic exponential decay of the H+ gradient, which proceeded to a point somewhat above the expected baseline, probably because of slow efflux of scalar H+ (Fig. 1). The decay of the H+ gradient was extrapolated back to a point determined to be halfway through the period of ATP hydrolysis (Fig. 2) from the time-course of H+ ejection in the presence of uncou- pler. The peak H+ ejection so extrapolated was divided by the amount of ATP added to yield the H+/ATP ratio (7, 12, 14); no correction was applied for scalar H+ production. The value of H+/ATP obtained from a large number of such experiments was 1.96 ? 0.09 (SD).
Fig. 3 shows that the added ATP was hydrolyzed completely to ADP within 8-12 sec; AMP formation was measured and found to be insignificantly small. Pulsing the mitochondria with ADP yielded no H+ ejection. Other inhibitors of mitochondrial anion transport (0.5 mM butylmalonate, 2 mM benzene- 1,2,3-tricarboxylate, 2 mg/ml of bovine serum albumin) or variation of the amount of EDTA present or ATP added (from 3 to 8 nmol/mg of protein) did not affect the observed H+/ATP ratios.
The pH dependence of the H+/ATP ratio is shown in Fig. 4. In the absence of inhibitors of phosphate transport, in which case the data were corrected for scalar H+ production according to Moyle and Mitchell (14), the H+/ATP ratio increased above
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 09:26:06 AMAll use subject to JSTOR Terms and Conditions
Biochemistry: Brand and Lehninger Proc. Natl. Acad. Sci. USA 74 (1977) 1957
5 ATPodded 78
a l 0 ADP 7, 4 - ll - 7/ o
3 20 60 o Ea E82 - 58E
F-~~~~~~~~~~~~a ATP <___
0 20 60 Seconds
FIG. 3. Hydrolysis of added ATP. For details see legend of Fig. 1. Samples of the system were removed at the times shown and as- sayed for ATP and ADP.
the value 2.0 at pH values below 6.5 and was as high as 3.0 at pH 6.1. As expected, N-ethylmaleimide was not particularly effective in preventing this rise, due to its poorer inhibitory action on the phosphate carrier at low pH (28, 29). However, in the presence of mersalyl, a potent inhibitor of phosphate- hydroxide exchange at low pH, the H+/ATP ratio remained fairly constant at close to 2.0 over the entire pH range from 6.0 to 7.4. The high values of the H+/ATP ratio at pH near 6 in the absence of inhibitor are due to the inappropriate correction used by Mitchell and Moyle for scalar H+ production.
DISCUSSION These experiments remove two major points of uncertainty regarding the H+/ATP ratios obtained by the procedures de- scribed by Mitchell and Moyle (7, 12-14): (i) possible under- estimation of H+ ejection caused by fast uptake of medium phosphate and H+ energized by ATP hydrolysis (see ref. 5) and (ii) an oversimplified correction for scalar H+ production. The value of 2.0 we have obtained for the H+/ATP ratio from ATP pulse experiments in intact mitochondria in the presence of N-ethylmaleimide or mersalyl is thus more reliable than the values of Mitchell and Moyle (7, 12-14). Although their cor- rection procedure also gave values of 2.0, this agreement is fortuitous.
As indicated earlier, Mitchell has assumed that the H+/site ratio is equal to the H+/ATP ratio. Our observations that the H+/site ratio is at least 3.0 (5, 16-20, 30), while the H+/ATP ratio is 2.0, would thus appear to contradict a basic premise of the chemiosmotic theory. It will now be shown that this con- tradiction is only apparent.
The experiments from which we calculate an H+/ATP ratio of near 2.0 are very similar in the dimensions of time and total
3.0 - Moyle-Mitchell method after correction for scalar H+ with
* X /. uncoupler
a- I- o- -_-_s
2.0 - 0
Present method (mersolyi) 0
correction not required
10 , I , , I I I ,I I _
6.0 6.4 6.8 7.2 Initial pH
FIG. 4. Effect of pH on the H+/ATP ratio. For details see legend of Fig. 1. Where indicated, mersalyl was present at 10 nmol/mg of protein.
MEMBRANE
OUT IN
+ +
:, 3F -H 3H +
H2PO~ H2P0_4
ATP + i USE ------ - *2H211 BY
CELL
ADp3-s ADP-
ATP4- ATP4-
FIG. 5. Modification of the chemiosmotic theory to account for coupling of the transport of phosphate and adenine nucleotides, as well as ATP synthesis, to an electrochemical gradient of H+ generated by oxidative phosphorylation in the steady state in vivo. For conve- nience, H2P04-/OH- antiport is shown as its formal equivalent, H2PO4-/H+ symport. I designates an energy-conserving site; II, the phosphate-hydroxide antiporter; III, the F1 ATPase; and IV, the ADP3- /ATP4- antiporter. The stoichiometry shown assumes flow of an electron-pair through the energy-coupling site.
H-t ejected per mg of mitochondrial protein to the oxygen-pulse measurements used to determine the H+/site ratio (5, 19), for which a value of 3.0 was found. Although we have observed H+/site values closer to 4.0 with two other methods (5, 17, 20), the conditions used in the latter are quite different from those used in the oxygen and ATP pulse experiments. There are reasons to believe that the oxygen-pulse and ATP-pulse mea- surements are subject to similar errors; thus, both may yield underestimates of H+ ejection. For these reasons the H+/ATP ratio derived from our ATP pulse experiments is best compared at present with the H+/site ratio derived from our oxygen-pulse measurements. The problem stated in the introduction may then be reformulated, "Are H+/site ratios of 3.0 and H+/ATP ratios of 2.0 still compatible with a chemiosmotic interpretation of oxidative phosphorylation?" [In view of the H+/site values close to 4.0 that we have observed by other procedures (5, 20), we cannot at present exclude the possibility that both the H+/ATP and H+/site ratios derived are underestimated by 1 unit due to some unrecognized common side reaction, in which case the question would be rephrased in terms of a H+/site value of 4.0 and an H+/ATP ratio of 3.0.]
A scheme that accommodates the available evidence on proton movements, including the difference between the H+/site and the H+/ATP ratios from pulse methods, is shown in Fig. 5 (see ref. 30). We propose that 3 H+ are ejected during passage of an electron-pair through each energy-conserving site and that 2 H1+ are then translocated inward by the mitochon- drial ATPase to make possible the formation of one molecule of ATP per site in the matrix. The third H+ ejected is utilized to drive the energy-requiring transport of ADP and Pi into the mitochondria and the coupled export of ATP to the cytosol. In this way the overall H+/ATP ratio for oxidative phosphoryl- ation in vivo, including the transport steps, is 3.0 and equal to the H+/site ratio.
The scheme in Fig. 5 assumes that the phosphate-hydroxide carrier promotes strict 112P04/OH- antiport (or strict 112PO4-/11 symport) (31) and that the adenine nucleotide
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 09:26:06 AMAll use subject to JSTOR Terms and Conditions

1958 Biochemistry: Brand and Lehninger Proc. Natl. Acad. Sci. USA 74 (1977)
carrier promotes strict ADP3-/ATP4- antiport (32, 41) between the site of ATP synthesis and the extramitochondrial com- partment, in accordance with the bulk of the available evidence but contrary to suggestions that the ADP-ATP exchange may be largely electroneutral during steady-state oxidative phos- phorylation (33). Although H2PO4-/H+ symport is electro- neutral, it is driven in the direction of phosphate entry at the expense of the H+ gradient generated by electron transport; thus, influx of one H?out is required to bring in each H2P04-. Since the net outward movement of one negative charge on ATP4- via the adenine nucleotide carrier is electrically equivalent to entry of one positive charge, the two carriers to- gether promote entry of one H+ per turnover. Thus the ex- change of ADP3-out and Pi-out for ATP4-in occurs at the ex- pense of a portion of the protonmotive force generated by electron transport, more precisely, at the expense of one of the 3 H+ ejected at each energy-conserving site (or, if the H+/site ratio is 4.0 and the H+/ATP ratio is 3.0, at the expense of one of the 4 H+ ejected at each site). In contrast, the hydrolysis of ATPout in ATP pulse experiments such as those described here and by Mitchell and Moyle (7, 12-14) requires no protonmotive force to bring about the electrogenic exchange of ATP4- with ADP3- since the test system contains valinomycin, which permits K+ to move through the membrane with ATP4-, thus rendering the net exchange of ADP3- for ATP4- + K+ elec- troneutral.
The formulation in Fig. 5 is in full agreement with thermo- dynamic data, indicating that 3 H+ must be ejected per 2e- per site in order that the observed protonmotive force generated by electron transport will be sufficient to balance the maximum phosphorylation potentials against which mitochondria can phosphorylate external ADP (9, 21-23). Our data also agree with and explain the observation that the phosphorylation po- tential achieved by intact rat liver mitochondria in the ex- tramitochondrial medium during steady-state respiration in state 4, about 16 kcal/mol, is substantially higher than the phosphorylation potential of the inner compartment, about 12 kcal/mol (ref. 32; see also refs. 33-38). The difference between these values closely agrees with our proposal that one-fourth or more of the total protonmotive force generated by electron flow is required for the transport steps alone (33-35, 37, 38). In concord with data of Davis et al. (39), our formulation also makes unnecessary the assumption (1, 37, 38, 40) that the P/O ratio of oxidative phosphorylation must be less than 1.0 per site whenever the phosphorylation potential achieved in the ex- ternal medium exceeds that of the matrix compartment.
Ionization of H2PO4- to give HP042- + H+ may occur on either side of the membrane during the oxidative phosphoryl- ation cycle in vivo (Fig. 5). However, this reaction is a dead end in the steady-state in vivo and cannot contribute to the H+ fluxes. This is obviously not true in isolated mitochondria carrying out net oxidative phosphorylation from added ADP and Pi, which results in net alkalinization of the medium, or carrying out hydrolysis of ATP to yield ADP and Pi, which results in net acidification.
In contrast to earlier chemiosmotic models (1, 38, 40), the scheme in Fig. 5 includes the energy cost of membrane trans- location steps in the measured P/O ratios of 3.0 for NAD-linked and 2.0 for FAD-linked substrates. It follows that inverted, fully coupled submitochondrial vesicles, in which transport of ade- nine nucleotides and phosphate is unnecessary and may be inhibited without impairing ATP production, will not require protonmotive force for transport steps. The P/O ratios of such inverted vesicles should consequently be 50% greater than in intact mitochondria, i.e., the overall P70 ratio should be 4.5 for
NADH and 3.0 for succinate oxidation if the chemiosmotic hypothesis is correct.
Because other experimental methods we have used (5, 17, 20) indicate that the H+/site ratio is actually 4.0, it is possible that the true H+/ATP ratio is correspondingly 3.0.
The data and considerations developed here, while com- patible with the chemiosmotic theory, do not prove that it is correct. On the other hand, alternative models of oxidative phosphorylation are valid only if they can ultimately account for the observed direction and stoichiometry of the H+ move- ments associated with electron transport and ATP synthesis or hydrolysis.
This work was supported by Grant BMS75-21923 from the National Science Foundation, Grant GM05919 from the National Institutes of Health, and Contract NO1-CP-45610 from the National Cancer In- stitute.
The costs of publication of this article were defrayed in part by the payment of page charges from funds made available to support the research which is the subject of the article. This article must therefore be hereby marked "advertisement" in accordance with 18 U. S. C. ?1734 solely to indicate this fact.
1. Mitchell, P. (1966) Chemiosmotic Coupling in Oxidative and Photosynthetic Phosphorylation (Glynn Research Ltd., Bod- min).
2. Mitchell, P. (1968) Chemiosmotic Coupling and Energy Transduction (Glynn Research Ltd., Bodmin).
3. Greville, G. D. (1969) in Current Topics in Bioenergetics, ed. Sanadi, D. R. (Academic Press, New York), Vol. 3, pp. 1-78.
4. Lehninger, A. L., Carafoli, E. & Rossi, C. S. (1967) Adv. Enzymol. 29, 259-320.
5. Brand, M. D., Reynafarje, B. & Lehninger, A. L. (1976) Proc. Natl. Acad. Sci. USA 73,437-441.
6. Mitchell, P. (1961) Nature 191, 144-148. 7. Mitchell, P. & Moyle, J. (1965) Nature 208, 147-151. 8. Mitchell, P. & Moyle, J. (1967) Biochem. J. 105, 1147-1162. 9. Papa, S. (1976) Biochim. Biophys. Acta 456, 39-84.
10. Bielawski, J. & Lehninger, A. L. (1966) J. Biol. Chem. 241, 4316-4322.
11. Rossi, C. S., Siliprandi, N., Carafoli, E., Bielawski, J. & Lehninger, A. L. (1967) Eur. J. Biochem. 2, 332-340.
12. Mitchell, P. & Moyle, J. (1968) Eur. J. Biochem. 4, 530-539. 13. Mitchell, P. & Moyle, J. (1970) FEBS Lett. 6, 309-311. 14. Moyle, J. & Mitchell, P. (1973) FEBS Lett. 30, 317-320. 15. Thayer, W. S. & Hinkle, P. C. (1973) J. Biol. Chem. 248,
5395-5402. 16. Lehninger, A. L. & Brand, M. D. (1976) in The Structural Basis
of Membrane Function, eds. Hatefi, Y. & Djavadi-Ohaniance, L. (Academic Press, New York), pp. 83-93.
17. Brand, M. D., Chen, C.-H. & Lehninger, A. L. (1976) J. Biol. Chem. 251, 968-974.
18. Brand, M. D., Reynafarje, B. & Lehninger, A. L. (1976) J. Cell. Physiol. 89, 595-602.
19. Brand, M. D., Reynafarje, B. & Lehninger, A. L. (1976) J. Biol. Chem. 251, 5670-5679.
20. Reynafarje, B., Brand, M. D. & Lehninger, A. L. (1976) J. Biol. Chem. 251, 7442-7451.
21. Nicholls, D. G. (1974) Eur. J. Biochem. 50, 305-315. 22. Rottenberg, H. (1975) J. Bioenerg. 7, 61-74. 23. Weichmann, A. H. C. A., Beem, E. P. & van Dam, K. (1975) in
Electron Transfer Chains and Oxidative Phosphorylation, eds. Quagliariello, E., Papa, S., Palmieri, F., Slater, E. C. & Siliprandi, N. (North-Holland, Amsterdam), pp. 335-342.
24. Ebel, R. E. & Lardy, H. A. (1975) J. Biol. Chem. 250, 4992- 4995.
25. Strehler, B. L. (1974) in Methods of Enzymatic Analysis, ed. Bergmeyer, H. U. (Verlag Chemie, Weinheim & Academic Press, New York), 2nd English edition, Vol. 4, pp. 2112-2126.
26. Lamprecht, W. & Trautschold, I. (1974) in Methods of Enzy- matic Analysis, ed. Bergmeyer, H. U. (Verlag Chemie, Wein-
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 09:26:06 AMAll use subject to JSTOR Terms and Conditions

Biochemistry: Brand and Lehninger Proc. Natl. Acad. Sci. USA 74 (1977) 1959
heim & Academic Press, New York), 2nd English edition, Vol. 4, pp. 2101-2110.
27. Jaworek, D., Gruber, W. & Bergmeyer, H. U. (1974) in Methods of Enzymatic Analysis, ed. Bergmeyer, H. U. (Verlag Chemie, Weinheim & Academic Press, New York), 2nd English edition, Vol. 4, pp. 2127-2131.
28. Klingenberg, M., Durand, R. & Guerin, B. (1974) Eur. J. Bio- chem. 42, 135-150.
29. Fonyo, A., Ligeti, E., Palmieri, F. & Quagliariello, E. (1975) in Biomembranes: Structure and Function, FEBS Symposium No. 35, eds. Gardos, G. & Szasz, I. (Akadamiai Kiad6, Budapest), pp. 287-306.
30. Brand, M. D., Reynafarje, B. & Lehninger, A. L. (1977) in FEBS Symposium on Biochemistry of Membrane Transport (Zurich, July 1976), in press.
31. McGivan, J. D. & Klingenberg, M. (1971) Eur. J. Biochem. 20, 392-399.
32. Klingenberg, M. (1970) Essays Biochem. 6, 119-159. 33. Klingenberg, M. (1975) in Energy Transformation in Biological
Systems, CIBA Foundation Symposium No. 31 (Elsevier, Am- sterdam), pp. 105-124.
34. Heldt, H. W., Klingenberg, M. & Milovancev, M. (1972) Eur. J. Biochem. 30, 434-440.
35. Slater, E. C., Rosing, J. & Mol, A. (1973) Biochim. Biophys. Acta 292,534-553.
36. Davis, E. J. & Lumeng, L. (1975) J. Biol. Chem. 250, 2275- 2282.
37. Klingenberg, M., Heldt, H. W. & Pfaff, E. (1969) in The Energy Level and Metabolic Control in Mitochondria, eds. Papa, S., Tager, J. M., Quagliariello, E. & Slater, E. C. (Adriatica Editrice, Bari), pp. 237-253.
38. Klingenberg, M., Wulf, R., Heldt, H. W. & Pfaff, E. (1969) in Mitochondria, Structure and Function, FEBS Symposium No. 17, eds. Ernster, L. & Drahota, Z. (Academic Press, London), pp. 59-77.
39. Davis, E. J., Lumeng, L. & Bottoms, D. (1974) FEBS Lett. 39, 9-12.
40. Mitchell, P. (1969) in The Molecular Basis of Membrane Functions, ed. Tosteson, D. C. (Prentice-Hall, Englewood Cliffs, NJ), pp. 483-518.
41. Klingenberg, M. & Rottenberg, H. (1977) Eur. J. Biochem. 73, 125-130.
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 09:26:06 AMAll use subject to JSTOR Terms and Conditions