Embed Size (px)
Citation preview

doi: 10.1006/cyto.1999.0533, available online at http://www.idealibrary.com on
HIGH ALTITUDE INCREASES CIRCULATINGINTERLEUKIN-6, INTERLEUKIN-1 RECEPTOR
ANTAGONIST AND C-REACTIVE PROTEIN
G. Hartmann,2 M. Tschop,1 R. Fischer, C. Bidlingmaier,2 R. Riepl, K. Tschop,2
H. Hautmann,1 S. Endres,2 M. Toepfer
Hypoxic pulmonary vasoconstriction is associated with but may not be sufficient for thedevelopment of high-altitude pulmonary oedema (HAPO). Hypoxia is known to induce aninflammatory response in immune cells and endothelial cells. It has been speculated thathypoxia-induced inflammatory cytokines at high altitude may contribute to the development ofHAPO by causing capillary leakage in the lung. We were interested if such an inflammatoryresponse, possibly involved in a later development of HAPO, is detectable at high altitude inindividuals without HAPO. We examined the plasma levels of interleukin 6 (IL-6), interleukin1 receptor antagonist (IL-1ra) and C-reactive protein (CRP) in two independent studies: studyA, Jungfraujoch, Switzerland, three overnight stays at 3458 m, n=12; study B: Capanna ReginaMargherita, Italy, 3 overnight stays at 3647 m and one overnight stay at 4559 m, n=10. In bothstudies, probands showed symptoms of acute mountain sickness but no signs of HAPO. At theJungfraujoch, IL-6 increased from 0.1�0.03 pg/ml to 2.0�0.5 pg/ml (day 2, P=0.03), IL-1rafrom 101�21 to 284�73 pg/ml (day 2, P=0.01), and CRP from 1.0�0.4 to 5.8�1.5 �g/ml(day 4, P=0.01). At the Capanna Margherita, IL-6 increased from 0.5�0.2 pg/ml to2.0�0.8 pg/ml (P=0.02), IL-1ra from 118�25 to 213�28 pg/ml (P=0.02), and CRP from0.4�0.03 to 3.5�1.1 �g/ml (P=0.03). IL-8 was below the detection limit of the ELISA(<25 pg/ml) in both studies. The increase of IL-6 and IL-1ra in response to high altitude wasdelayed and preceded the increase of CRP. We conclude that: (1) circulating IL-6, IL-1ra andCRP are upregulated in response to hypobaric hypoxic conditions at high altitude, and (2) themoderate systemic increase of these inflammatory markers may reflect considerable localinflammation. The existence and the kinetics of high altitude-induced cytokines found in thisstudy support the hypothesis that inflammation is involved in the development of HAPO.
� 2000 Academic Press
From the 1Mountain Medicine Study Group and 2Divisionof Clinical Pharmacology, Medizinische Klinik, Ludwig-Maximilians-University, Munich, Germany; Current address:G. Hartmann, Department of Internal Medicine, University ofIowa, USA
Correspondence to: Hartmann Gunther, Departmant of InternalMedicine, University of Iowa, 540 EMRB, Iowa City, IA 52242,USA; E-mail: [email protected]
Received 17 October 1998; received in revised form 24 February1999; accepted for publication 6 May 1999
� 2000 Academic Press1043–4666/00/030246+07 $35.00/0
KEY WORDS: C-reactive protein/cytokines/high altitude/hypoxia/interleukin-6/interleukin-1 receptor antagonist
Several protective physiological responses to localhypoxia have been evolved during evolution. In thelung, vasoconstriction of the pulmonary arteries inregions with low alveolar oxygen contributes to opti-mal oxygen saturation of the blood in regions withhigh oxygen (known as Euler-Lilijenstrand mechan-ism). The brain increases oxygen delivery by local
246
vasodilatation leading to enhanced cerebral bloodflow. Low oxygen co-activates immune cells1–4 and theendothelium,5–9 which is involved in tissue injury andtissue repair. All of these mechanisms are locallyrestricted and are beneficial in a normal oxygenenvironment. However, in a low oxygen environmentat high altitude, these normally protective mechanismscause high altitude-associated diseases.
The pathogenesis of high-altitude pulmonaryoedema (HAPO) and high-altitude cerebral oedema(HACO), two life-threatening diseases in mountain-eers, is not completely understood. HACO developsfrom acute mountain sickness (AMS) which is charac-terized by headache, dizziness, sleeplessness, nauseaand vomiting. Vasodilatation in the brain is in partresponsible for AMS and HACO.10 HAPO developsindependently from AMS but is often associated withit.11 Generalized pulmonary vasoconstriction resultingin an increased pulmonary artery pressure is associated
CYTOKINE, Vol. 12, No. 3 (March), 2000: pp 246–252
Cytokines at high altitude / 247
with but may not be sufficient for the development ofHAPO.11 Hypoxia reduces the formation of vasodila-tory nitric oxide in the endothelium of the pulmonaryarteries leading to pulmonary vasoconstriction.12 Thevasoconstrictor endothelin-1 is elevated in high alti-tude,13 but the impact of endothelin-1 in the patho-genesis of HAPO is still unclear.14 In addition topulmonary vasoconstriction, HAPO is associated withan increase in leakage of both fluid and proteins intothe alveolar space.15–17 Recently we found that twoother hormones, leptin18 and adrenomedullin,19 areelevated at high altitude. While leptin may play a keyrole in the disregulation of energy homeostasis athigh altitude, adrenomedullin seems to balance thevasoconstrictive activity of endothelin-1.
Recent observations of high pulmonary arterypressure in individuals who did not develop HAPOsuggest that additional factors like inflammation ordecreased fluid clearance from the lung may contributeto the development of HAPO.20 It has been speculatedthat hypoxia-induced immune activation is involved inthe pathogenesis of HAPO.16,20–22 This hypothesis isbased on the following observations: (1) patients withHAPO have high concentrations of inflammatorycytokines in the plasma and in the lungs;21 (2) ahigh incidence of preexisting respiratory infections incases of HAPO and an increased susceptibility ofrats with respiratory infection to HAPO has beendescribed;23 and (3) inflammatory cytokines enhancecapillary permeability and hypoxic pulmonaryvasoconstriction.24,25
It is known that hypoxia stimulates the expressionof inflammatory cytokines and adhesion moleculesin macrophages and endothelial cells in vitro andenhances NK-cell activity in humans exposed to highaltitude.1,5,9 If contributing to the development ofHAPO, an inflammatory response to high altitude hasto be present prior to the development of HAPO.Therefore, the goal of the present study was to examineif exposure to hypoxia at high altitude causes aninflammatory response in individuals without HAPO.Using sensitive assays we were indeed able to showelevated levels of circulating cytokines, which could beinvolved in a possible subsequent development ofHAPO in susceptible individuals.
RESULTS
Clinical examinationPassive transportation to the Jungfraujoch
(3458 m) caused symptoms of AMS (score >5) in twoof 12 probands after the first night (mean AMS scoreof all probands 3.7�0.7) and in three probands afterthe second night. The average of symptoms of AMSwas higher in the morning. On day 4, none of the
probands had AMS. Arterial oxygen saturationmeasured at 8 a.m. after the first night at high altitudewas 88.5% (83–92%), partial pressure of oxygen was51 mm Hg (43 to 62 mm Hg). None of the probandsdeveloped HAPO (clinical examination) or HACO(clinical symptoms). Symptoms of AMS were higherafter active ascent to Capanna Margherita (4559 m),although all probands were well adapted to an altitudeof 3647 m (three overnight stays). Eight of 10 probandssuffered from AMS 2 h after arrival at the CapannaMargherita (AMS score: mean of 5.6�0.7), but noneof them showed signs of HAPO. In the next morning,five of 10 probands had AMS (AMS score: mean of5.1�0.9). All probands had severe hypoxia (oxygensaturation: mean 81.9%, range 75–90%).
3
0
8 a.m.
IL-6
(pg
/ml)
Day 3Day 2
2
1
8 p.m. 8 p.m. 8 a.m.
P = 0.03
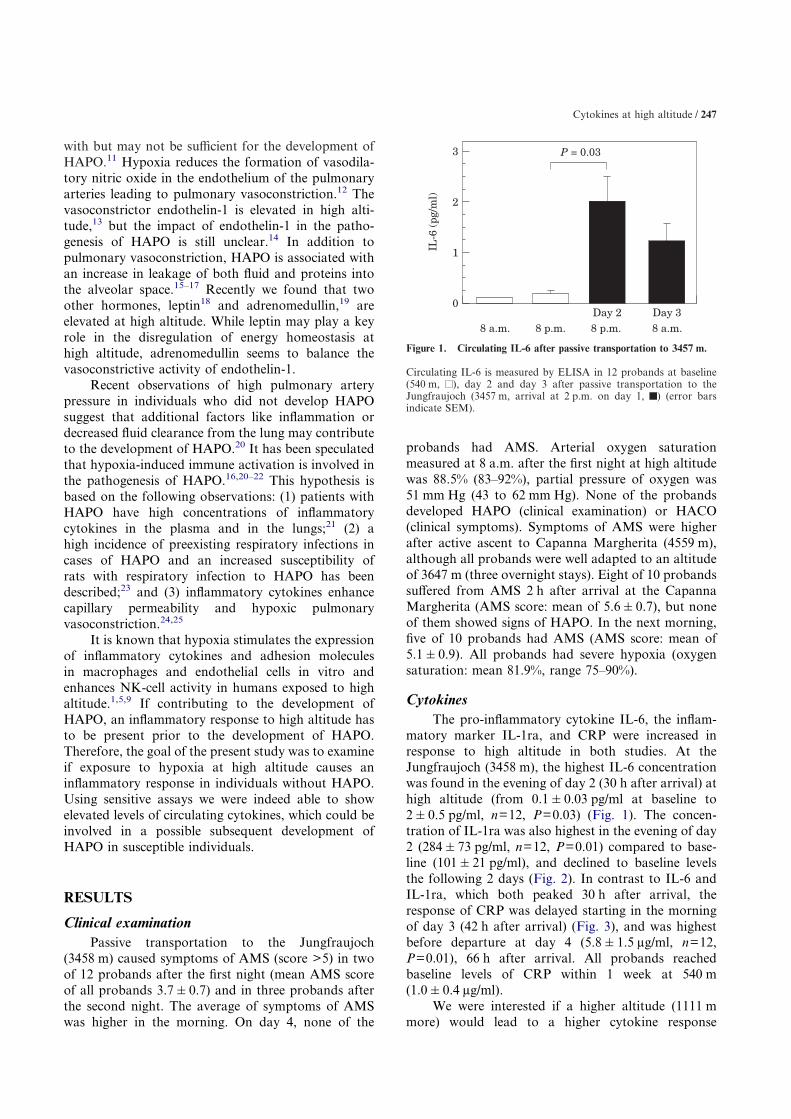
Figure 1. Circulating IL-6 after passive transportation to 3457 m.
Circulating IL-6 is measured by ELISA in 12 probands at baseline(540 m, ), day 2 and day 3 after passive transportation to theJungfraujoch (3457 m, arrival at 2 p.m. on day 1, ) (error barsindicate SEM).
CytokinesThe pro-inflammatory cytokine IL-6, the inflam-
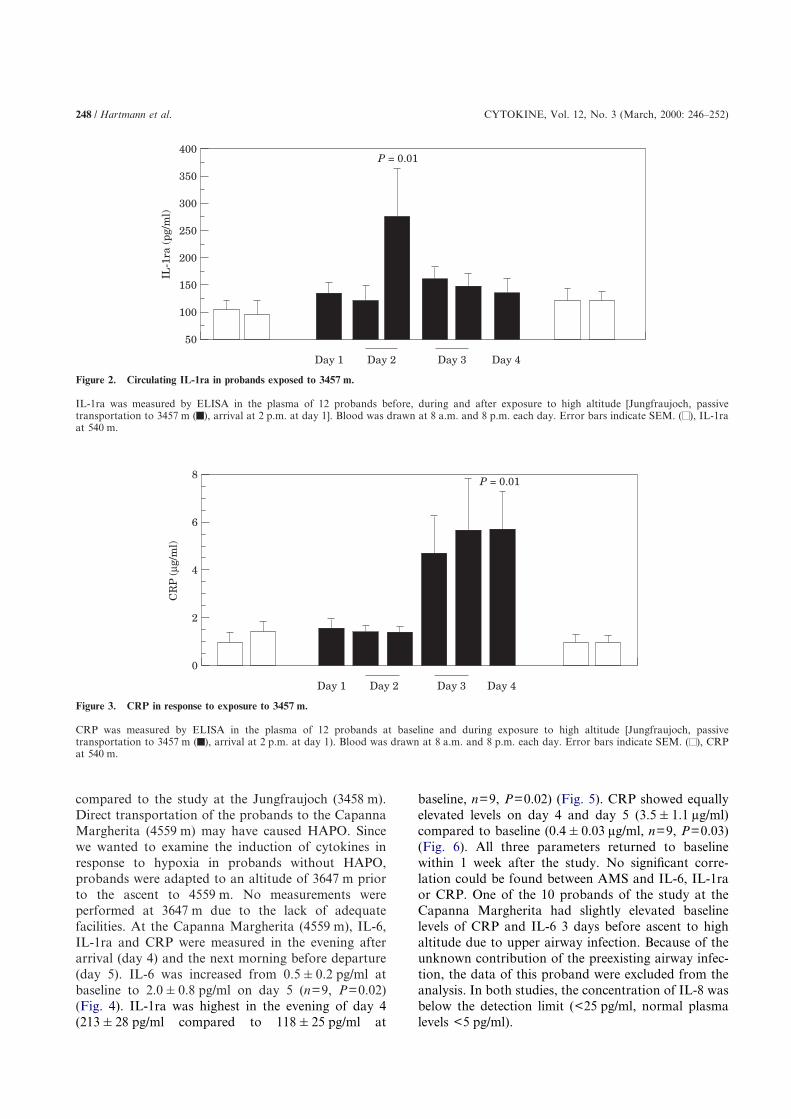
matory marker IL-1ra, and CRP were increased inresponse to high altitude in both studies. At theJungfraujoch (3458 m), the highest IL-6 concentrationwas found in the evening of day 2 (30 h after arrival) athigh altitude (from 0.1�0.03 pg/ml at baseline to2�0.5 pg/ml, n=12, P=0.03) (Fig. 1). The concen-tration of IL-1ra was also highest in the evening of day2 (284�73 pg/ml, n=12, P=0.01) compared to base-line (101�21 pg/ml), and declined to baseline levelsthe following 2 days (Fig. 2). In contrast to IL-6 andIL-1ra, which both peaked 30 h after arrival, theresponse of CRP was delayed starting in the morningof day 3 (42 h after arrival) (Fig. 3), and was highestbefore departure at day 4 (5.8�1.5 �g/ml, n=12,P=0.01), 66 h after arrival. All probands reachedbaseline levels of CRP within 1 week at 540 m(1.0�0.4 �g/ml).
We were interested if a higher altitude (1111 mmore) would lead to a higher cytokine response
248 / Hartmann et al. CYTOKINE, Vol. 12, No. 3 (March, 2000: 246–252)
400
50
IL-1
ra (
pg/m
l)
Day 2Day 1
P = 0.01
350
300
250
200
150
100
Day 3 Day 4
Figure 2. Circulating IL-1ra in probands exposed to 3457 m.
IL-1ra was measured by ELISA in the plasma of 12 probands before, during and after exposure to high altitude [Jungfraujoch, passivetransportation to 3457 m ( ), arrival at 2 p.m. at day 1]. Blood was drawn at 8 a.m. and 8 p.m. each day. Error bars indicate SEM. ( ), IL-1raat 540 m.
8
0
CR
P (
µg/m
l)
Day 2Day 1
P = 0.01
6
4
2
Day 3 Day 4
Figure 3. CRP in response to exposure to 3457 m.
CRP was measured by ELISA in the plasma of 12 probands at baseline and during exposure to high altitude [Jungfraujoch, passivetransportation to 3457 m ( ), arrival at 2 p.m. at day 1). Blood was drawn at 8 a.m. and 8 p.m. each day. Error bars indicate SEM. ( ), CRPat 540 m.
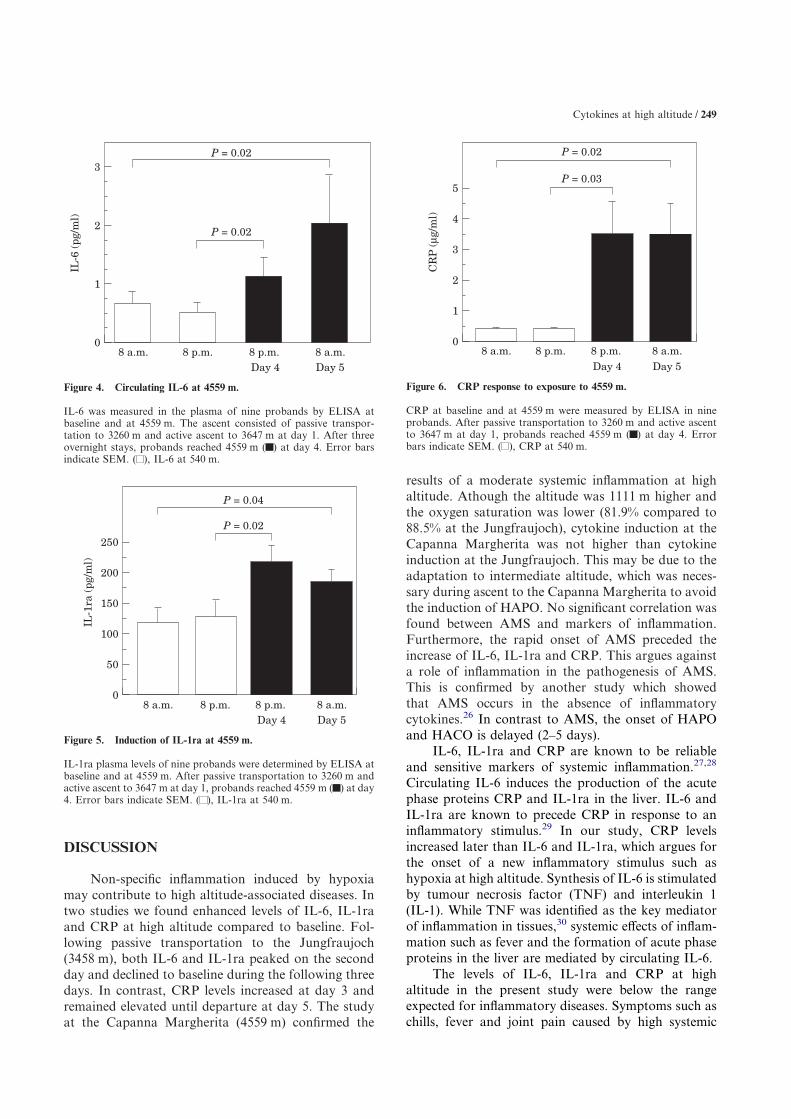
compared to the study at the Jungfraujoch (3458 m).Direct transportation of the probands to the CapannaMargherita (4559 m) may have caused HAPO. Sincewe wanted to examine the induction of cytokines inresponse to hypoxia in probands without HAPO,probands were adapted to an altitude of 3647 m priorto the ascent to 4559 m. No measurements wereperformed at 3647 m due to the lack of adequatefacilities. At the Capanna Margherita (4559 m), IL-6,IL-1ra and CRP were measured in the evening afterarrival (day 4) and the next morning before departure(day 5). IL-6 was increased from 0.5�0.2 pg/ml atbaseline to 2.0�0.8 pg/ml on day 5 (n=9, P=0.02)(Fig. 4). IL-1ra was highest in the evening of day 4(213�28 pg/ml compared to 118�25 pg/ml at
baseline, n=9, P=0.02) (Fig. 5). CRP showed equallyelevated levels on day 4 and day 5 (3.5�1.1 �g/ml)compared to baseline (0.4�0.03 �g/ml, n=9, P=0.03)(Fig. 6). All three parameters returned to baselinewithin 1 week after the study. No significant corre-lation could be found between AMS and IL-6, IL-1raor CRP. One of the 10 probands of the study at theCapanna Margherita had slightly elevated baselinelevels of CRP and IL-6 3 days before ascent to highaltitude due to upper airway infection. Because of theunknown contribution of the preexisting airway infec-tion, the data of this proband were excluded from theanalysis. In both studies, the concentration of IL-8 wasbelow the detection limit (<25 pg/ml, normal plasmalevels <5 pg/ml).
Cytokines at high altitude / 249
3
0
IL-6
(pg
/ml)
2
1
P = 0.02
P = 0.02
8 a.m.Day 5Day 4
8 p.m. 8 p.m. 8 a.m.
Figure 4. Circulating IL-6 at 4559 m.
IL-6 was measured in the plasma of nine probands by ELISA atbaseline and at 4559 m. The ascent consisted of passive transpor-tation to 3260 m and active ascent to 3647 m at day 1. After threeovernight stays, probands reached 4559 m ( ) at day 4. Error barsindicate SEM. ( ), IL-6 at 540 m.
250
08 a.m.
IL-1
ra (
pg/m
l)
Day 5Day 4
200
50
8 p.m. 8 p.m. 8 a.m.
P = 0.02
150
100
P = 0.04
Figure 5. Induction of IL-1ra at 4559 m.
IL-1ra plasma levels of nine probands were determined by ELISA atbaseline and at 4559 m. After passive transportation to 3260 m andactive ascent to 3647 m at day 1, probands reached 4559 m ( ) at day4. Error bars indicate SEM. ( ), IL-1ra at 540 m.
5
08 a.m.
CR
P (
µg/m
l)
Day 5Day 4
3
1
8 p.m. 8 p.m. 8 a.m.
P = 0.03
P = 0.02
4
2
Figure 6. CRP response to exposure to 4559 m.
CRP at baseline and at 4559 m were measured by ELISA in nineprobands. After passive transportation to 3260 m and active ascentto 3647 m at day 1, probands reached 4559 m ( ) at day 4. Errorbars indicate SEM. ( ), CRP at 540 m.
DISCUSSION
Non-specific inflammation induced by hypoxiamay contribute to high altitude-associated diseases. Intwo studies we found enhanced levels of IL-6, IL-1raand CRP at high altitude compared to baseline. Fol-lowing passive transportation to the Jungfraujoch(3458 m), both IL-6 and IL-1ra peaked on the secondday and declined to baseline during the following threedays. In contrast, CRP levels increased at day 3 andremained elevated until departure at day 5. The studyat the Capanna Margherita (4559 m) confirmed the
results of a moderate systemic inflammation at highaltitude. Athough the altitude was 1111 m higher andthe oxygen saturation was lower (81.9% compared to88.5% at the Jungfraujoch), cytokine induction at theCapanna Margherita was not higher than cytokineinduction at the Jungfraujoch. This may be due to theadaptation to intermediate altitude, which was neces-sary during ascent to the Capanna Margherita to avoidthe induction of HAPO. No significant correlation wasfound between AMS and markers of inflammation.Furthermore, the rapid onset of AMS preceded theincrease of IL-6, IL-1ra and CRP. This argues againsta role of inflammation in the pathogenesis of AMS.This is confirmed by another study which showedthat AMS occurs in the absence of inflammatorycytokines.26 In contrast to AMS, the onset of HAPOand HACO is delayed (2–5 days).
IL-6, IL-1ra and CRP are known to be reliableand sensitive markers of systemic inflammation.27,28
Circulating IL-6 induces the production of the acutephase proteins CRP and IL-1ra in the liver. IL-6 andIL-1ra are known to precede CRP in response to aninflammatory stimulus.29 In our study, CRP levelsincreased later than IL-6 and IL-1ra, which argues forthe onset of a new inflammatory stimulus such ashypoxia at high altitude. Synthesis of IL-6 is stimulatedby tumour necrosis factor (TNF) and interleukin 1(IL-1). While TNF was identified as the key mediatorof inflammation in tissues,30 systemic effects of inflam-mation such as fever and the formation of acute phaseproteins in the liver are mediated by circulating IL-6.
The levels of IL-6, IL-1ra and CRP at highaltitude in the present study were below the rangeexpected for inflammatory diseases. Symptoms such aschills, fever and joint pain caused by high systemic

250 / Hartmann et al. CYTOKINE, Vol. 12, No. 3 (March, 2000: 246–252)
levels of cytokines in inflammatory diseases are notcommon at high altitude. Thus, if elevated at highaltitude, systemic cytokine levels are expected to be inthe upper normal range. Detection of such minorchanges of cytokines at high altitude requires highsensitivity ELISAs as used in our study. A moderateincrease of circulating cytokine levels may reflect sub-stantial inflammation in specific tissues such as thelung, which might contribute to the pathogenesis ofHAPO in susceptible individuals. Higher levels ofinflammatory cytokines during infectious diseasesmight be responsible for the higher rate of HAPO inanimal models or in mountaineers with an infection.23
Different metabolic needs and microvascular flowreserve determine the tolerance of tissues to hypoxia.The brain is highly sensitive to hypoxia. Szarflarskiet al. showed a transient increase of IL-1 and TNFexpression in the brain of rats following hypoxia.31
Astrocytes, connective tissue cells of the brain, produceIL-6 in response to hypoxia.32,33
Oxygen is critical for the heart. Hypoxia stimu-lates IL-6 synthesis in cardiac myocytes.34 In vascularendothelium, low oxygen stimulates the expressionof both inflammatory cytokines and adhesionmolecules.5–9 Finally, immune cells are activated bylow oxygen. Hypoxia synergizes with different immunestimuli for the production of inflammatory cytokinesand adhesion molecules in human peripheral bloodmononuclear cells and alveolar macrophages.1–4
NK-cell activity is enhanced in humans exposed tohigh altitude.35
Thus, hypoxia functions as a danger signal for theimmune system by inducing the synthesis of inflamma-tory cytokines and the expression of adhesion mol-ecules in different cell types and tissues. Oxygensensitive elements in promotor regions of genes andhypoxia-induced RNA binding proteins33 have beenidentified to regulate cellular responses to low oxy-gen.36 The responses to low oxygen concentrationhave been evolved as a physiological mechanism todetect tissue injury and to improve tissue repair.However, these responses are potentially harmful ifenvironmental oxygen decreases at high altitude.
Several studies examined cytokines in high alti-tude. Klausen et al. reported a comparable increase ofIL-6 from 1.4 at baseline to 4.7 pg/ml at day 2 afterpassive transportation to 4350 m,37 and a second peakof IL-6 at day 4 probably due to exercise at day 3. Incontrast to our study, they were not able to detectelevated concentrations of IL-1ra and CRP. This mightbe due to a difference in sensitivity of the used ELISAs,however, information about the lower detection limitsof the ELISAs is not provided in their study. Klegeret al. examined capillary permeability following passiveand active ascent to 4559 m.16 They found a highincrease of both IL-6 and CRP in probands with
HAPO (IL-6 >20 pg/ml at day 2, CRP >40 �g/ml atday 4). Minor but no significant changes of IL-6 andCRP were found in probands with AMS but withoutHAPO. The lack of significant cytokine detection innon-HAPO individuals may be again due to lowersensitivity of the used ELISAs. In another study,increased IL-6 was found in the bronchoalveolar fluidof patients with HAPO. Plasma cytokine levels werenot examined in this study.21 Swenson et al. found noinduction of inflammatory cytokines during the first8 h of exposure to hypoxia.26 This is in accordancewith our finding of earliest detection of cytokines atday 2. These studies and the present study examine thecytokine response in non-adapted individuals exposedto a rapid decrease in environmental oxygen. To date,no immunological data are available for individualspermanently living at high altitude.
In conclusion, the present study demonstrates thepresence of moderate systemic inflammation inresponse to high altitude. The time course of thisinflammatory response argues against an involvementof inflammation in AMS. If hypoxia-induced inflam-mation is involved in the development of HAPO orHACO has to be addressed in further studies testinganti-inflammatory drugs as prophylactic therapy inindividuals susceptible to HAPO.
MATERIALS AND METHODS
Probands and study designTwo independent studies were performed, study A at
the Jungfraujoch (3458 m) and study B at the CapannaMargherita (4559 m).
Study ATwelve healthy probands (1 female, 11 male, age 28�2
years, range 19 to 41) reached the research facility at theJungfraujoch (3458 m), Switzerland, by passive transporta-tion on day 1 (2 p.m.). Probands stayed at the Jungfraujochfor a total of 68 h (day 1 to day 4), including three overnightstays. Probands were confined to moderate activities. Bloodsamples were drawn at 8 a.m. and 8 p.m. each day, clinicalevaluation was performed three times a day.
Study BTen healthy probands (3 female, 7 male, age 32�5
years, range 20 to 41) started their active ascent at an altitudeof 3260 m and reached 3647 m on day 1. After three over-night stays, they continued the ascent to the CapannaMargherita (4559 m) on day 4 (arrival at 1 p.m.) and stayedthere for 22 h. Clinical evaluation and collection of bloodsamples were performed at the Capanna Margherita at6 p.m. and at 8 a.m. In both studies, blood pressure, heartrate, blood gas analysis and clinical signs of the lung wereexamined to detect early signs of HAPO. Blood was collectedin EDTA-containing pyrogen-free tubes and plasma sampleswere frozen at �20�C. Baseline clinical parameters and

Cytokines at high altitude / 251
baseline cytokines were assessed within 1 week before andafter the study in Munich (540 m).
Acute mountain sickness scoreThe degree of acute mountain sickness was determined
by the Lake Louise score38 including the clinical symptomsheadache, dizziness, sleeplessness, loss of appetite, nauseaand vomiting. Acute mountain sickness was diagnosed whenthe Lake Louise score was higher than 5.
Detection of cytokinesThe ELISAs used for cytokine measurement had the
following lower detection limits: IL-6 ELISA (Biotrak)0.1 pg/ml, IL-1ra ELISA (R&D Systems) 22 pg/ml, CRPELISA (DRG) 0.1 �g/ml, and IL-8 ELISA (Biotrak)25 pg/ml.
Statistical evaluationData were expressed as means�SEM. Statistical sig-
nificance of differences was determined by the unpairedtwo-tailed Student’s t-test. Differences were consideredstatistically significant for P<0.05. Statistical analyseswere performed by using StatView 4.51 software (AbacusConcepts Inc., Calabasas, CA, USA).
Acknowledgement
This study was supported by the Max-PlanckGesellschaft and the Deutsche Gesellschaft fur Bergund Expeditionsmedizin. We thank the Italian AlpineClup (CAI Varallo) for hospitality at the CapannaRegina Margherita.
REFERENCES
1. Ghezzi P, Dinarello CA, Bianchi M, Rosandich ME,Repine JE, White CW (1991) Hypoxia increases production ofinterleukin-1 and tumor necrosis factor by human mononuclear cells.Cytokine 3:189–194.
2. Hempel SL, Monick MM, Hunninghake GW (1996) Effectof hypoxia on release of IL-1 and TNF by human alveolar macro-phages. Am J Respir Cell Mol Biol 14:170–176.
3. Scannell G (1996) Leukocyte responses to hypoxic/ischemicconditions. New Horiz 4:179–183.
4. VanOtteren GM, Standiford TJ, Kunkel SL, Danforth JM,Strieter RM (1995) Alterations of ambient oxygen tension modulatethe expression of tumor necrosis factor and macrophage inflamma-tory protein-1 alpha from murine alveolar macrophages. Am JRespir Cell Mol Biol 13:399–409.
5. Shreeniwas R, Koga S, Karakurum M, Pinsky D, Kaiser E,Brett J, Wolitzky BA, Norton C, Plocinski J, Benjamin W, BurnsDK, Goldstein A and Stern D (1992) Hypoxia-mediated inductionof endothelial cell interleukin-1 alpha. An autocrine mechanismpromoting expression of leukocyte adhesion molecules on the vesselsurface. J Clin Invest 90:2333–2339.
6. Yan SF, Ogawa S, Stern DM, Pinsky DJ (1997) Hypoxia-induced modulation of endothelial cell properties: regulation ofbarrier function and expression of interleukin-6. Kidney Int 51:419–425.
7. Ala Y, Palluy O, Favero J, Bonne C, Modat G, Dornand J(1992) Hypoxia/reoxygenation stimulates endothelial cells to pro-
mote interleukin-1 and interleukin-6 production. Effects of freeradical scavengers. Agents Actions 37:134–139.
8. Ginis I, Mentzer SJ, Faller DV (1993) Oxygen tensionregulates neutrophil adhesion to human endothelial cells via anLFA-1-dependent mechanism. J Cell Physiol 157:569–578.
9. Karakurum M, Shreeniwas R, Chen J, Pinsky D, Yan SD,Anderson M, Sunouchi K, Major J, Hamilton T, Kuwabara K et al.(1994) Hypoxic induction of interleukin-8 gene expression in humanendothelial cells. J Clin Invest 93:1564–1570.
10. Hamilton AJ, Cymmerman A, Black PM (1986) Highaltitude cerebral edema. Neurosurgery 19:841–849.
11. Bartsch P (1997) High altitude pulmonary edema. Respir-ation 64:435–443.
12. Scherrer U, Vollenweider L, Delabays A, Savcic M,Eichenberger U, Kleger GR, Fikrle A, Ballmer PE, Nicod P, BartschP (1996) Inhaled nitric oxide for high-altitude pulmonary edema.N Engl J Med 334:624–629.
13. Goerre S, Wenk M, Bartsch P, Luscher TF, Niroomand F,Hohenhaus E, Oelz O, Reinhart WH (1995) Endothelin-1 in pulmon-ary hypertension associated with high-altitude exposure. Circulation91:359–364.
14. Droma Y, Ge RL, Tanaka M, Koizumi T, Hanaoka M,Miyahara T, Yamaguchi S, Okada K, Yoshikawa S, Fujimoto K,Matsuzawa Y, Kubo K, Kobayashi T, Sekiguchi M (1996) Acutehypoxic pulmonary vascular response does not accompany plasmaendothelin-1 elevation in subjects susceptible to high altitudepulmonary edema. Intern Med 35:257–260.
15. Schoene RB, Swenson ER, Pizzo CJ, Hackett PH, RoachRC, Mills WJ Jr, Henderson WR Jr, Martin TR (1988) The lung athigh altitude: bronchoalveolar lavage in acute mountain sickness andpulmonary edema. J Appl Physiol 64:2605–2613.
16. Kleger GR, Bartsch P, Vock P, Heilig B, Roberts Ln,Ballmer PE (1996) Evidence against an increase in capillary per-meability in subjects exposed to high altitude. J Appl Physiol81:1917–1923.
17. Hansen JM, Olsen NV, Feldt-Rasmussen B, Kanstrup IL,Dechaux M, Dubray C, Richalet JP (1994) Albuminuria and overallcapillary permeability of albumin in acute altitude hypoxia. J ApplPhysiol 76:1922–1927.
18. Tschop M, Strasburger CJ, Hartmann G, Biollaz J, BartschP (1998) Elevated leptin levels at high altitude are associated withloss of appetite. Lancet 352:1119–1120.
19. Toepfer M, Hartmann G, Schlosshauer M, Hautmann H,Tschop M, Fischer R, Huber RM (1998) Adrenomedullin: a playerat high altitude? Chest 113:1428.
20. Bartsch P (1997) High altitude pulmonary edema. Respir-ation 64:435–443.
21. Kubo K, Hanaoka M, Yamaguchi S, Hayano T, HayasakaM, Koizumi T, Fujimoto K, Kobayashi T, Honda T (1996)Cytokines in bronchoalveolar lavage fluid in patients with highaltitude pulmonary oedema at moderate altitude in Japan. Thorax51:739–742.
22. Kaminsky DA, Jones K, Schoene RB, Voelkel NF (1996)Urinary leukotriene E4 levels in high-altitude pulmonary edema. Apossible role for inflammation. Chest 110:939–945.
23. Carpenter TC, Reeves JT, Durmowicz AG (1998) Viralrespiratory infection increases susceptibility of young rats tohypoxia-induced pulmonary edema. J Appl Physiol 84:1048–1054.
24. Ziesche R, Petkov V, Williams J, Zakeri SM, Mosgoller W,Knofler M, Block LH (1996) Lipopolysaccharide and interleukin 1augment the effects of hypoxia and inflammation in human pulmon-ary arterial tissue. Proc Natl Acad Sci USA 93:12 478–12 483.
25. Liu SF, Dewar A, Crawley DE, Barnes PJ, Evans TW(1992) Effect of tumor necrosis factor on hypoxic pulmonary vaso-constriction. J Appl Physiol 72:1044–1049.
26. Swenson ER, MacDonald A, Vatheuer M, Maks C,Treadwell A, Allen R, Schoene RB (1997) Acute mountain sicknessis not altered by a high carbohydrate diet nor associated withelevated circulating cytokines. Aviat Space Environ Med 68:499–503.
27. Adamik B, Zimecki M, Wlaszczyk A, Kubler A (1997)Immunological status of septic and trauma patients: High tumornecrosis factor alpha serum levels in septic and trauma patients are

252 / Hartmann et al. CYTOKINE, Vol. 12, No. 3 (March, 2000: 246–252)
not responsible for increased mortality; a prognostic value of seruminterleukin 6. Arch Immunol Ther Exp 45:169–175.
28. Herrmann JL, Blanchard H, Brunengo P, Lagrange PH(1994) TNF alpha, IL-1 beta and IL-6 plasma levels in neutropenicpatients after onset of fever and correlation with the C-reactiveprotein (CRP) kinetic values. Infection 22:309–315.
29. Arend WP, Malyak M, Guthridge CJ, Gabay C (1998)Interleukin-1 receptor antagonist: role in biology. Annu Rev Immu-nol 16:27–55.
30. Eigler A, Sinha B, Hartmann G, Endres S (1997) TamingTNF: strategies to restrain this proinflammatory cytokine. ImmunolToday 18:487–492.
31. Szaflarski J, Burtrum D, Silverstein FS (1995) Cerebralhypoxia-ischemia stimulates cytokine gene expression in perinatalrats. Stroke 26:1093–1100.
32. Hori O, Matsumoto M, Kuwabara K, Maeda Y, Ueda H,Ohtsuki T, Kinoshita T, Ogawa S, Stern DM, Kamada T (1996)Exposure of astrocytes to hypoxia/reoxygenation enhances expres-sion of glucose-regulated protein 78 facilitating astrocyte release ofthe neuroprotective cytokine interleukin 6. J Neurochem 66:973–979.
33. Matsuo N, Ogawa S, Imai Y, Takagi T, Tohyama M, SternD, Wanaka A (,121995) Cloning of a novel RNA binding poly-peptide (RA301) induced by hypoxia/reoxygenation. J Biol Chem270:28 216–28 222.
34. Yamauchi TK, Ihara Y, Ogata A, Yoshizaki K, Azuma J,Kishimoto T (1995) Hypoxic stress induces cardiac myocyte-derivedinterleukin-6. Circulation 91:1520–1524.
35. Klokker M, Kharazmi A, Galbo H, Bygbjerg I, PedersenBK (1993) Influence of in vivo hypobaric hypoxia on function oflymphocytes, neutrocytes, natural killer cells, and cytokines. J ApplPhysiol 74:1100–1106.
36. Fandrey J (1995) Hypoxia-inducible gene expression.Respir Physiol 101:1–10.
37. Klausen T, Olsen NV, Poulsen TD, Richalet JP, PedersenBK (1997) Hypoxemia increases serum interleukin-6 in humans. EurJ Appl Physiol 76:480–482.
38. Bartsch P, Mueller A, Hofstetter D, Maggiorini M, Vock P,Oelz O (1993) AMS and HAPE scoring in the Alps. In Sutton JR,Houston CS, Coates G (eds) Hypoxia and Molecular Medicine,Queen City Printers, Burlington VT, pp 265–271.
![Automation of [ F]fluoroacetaldehyde synthesis ... · Automation of [18F]fluoroacetaldehydesynthesis:applicationtoarecombinanthuman interleukin-1 receptor antagonist (rhIL-1RA)†](https://img.pdfslide.net/doc/110x75/6011fd57f1072037ad7b66b8/automation-of-ffluoroacetaldehyde-synthesis-automation-of-18ffluoroacetaldehydesynthesisapplicationtoarecombinanthuman.jpg)




























