Embed Size (px)
Citation preview

CLINICAL MICROBIOLOGY REVIEWS, Apr. 1992, p. 183-203 Vol. 5, No. 20893-8512/92/020183-21$02.00/0Copyright C) 1992, American Society for Microbiology
High-Frequency Switching in Candida albicansDAVID R. SOLL
Department of Biology, University of Iowa, Iowa City, Iowa 52242
INTRODUCTION ............................................................. 183DIMORPHISM AND SWITCHING ............................................................. 184HIGH-FREQUENCY VARIABILITY: EARLY OBSERVATIONS...................................................184HIGH-FREQUENCY SWITCHING: REDISCOVERY............................................................. 184ZINC AND COLONY PHENOTYPE IN STRAIN 3153A ............................................................. 188PLOIDY, MITOTIC RECOMBINATION, AND SWITCHING.......................................................188WHITE-OPAQUE TRANSITION............................................................. 188
Measuring the Frequency of Switching ............................................................. 190Environmental Parameters which Affect Switching ............................................................. 190Effects on Cellular Morphology ............................................................. 191Relationship between Hypha Formation and the White-Opaque Transition .....................................191Antigenic Variability............................................................. 193
GENE REGULATION AND SWITCHING............................................................. 193MECHANISM OF SWITCHING............................................................. 196EFFECTS OF SWITCHING ON PUTATIVE VIRULENCE TRAITS ..............................................198Hypha Formation............................................................. 198Acid Protease Secretion ............................................................. 198Adhesion to Epithelium and Plastic ............................................................. 198Drug Susceptibility ............................................................. 198Antigenic Change ............................................................. 199Sensitivity to Neutrophils and In Vitro Oxidants ............................................................. 199
SWITCHING IN INFECTING STRAINS............................................................. 199SWITCHING AND ADAPTATION ............................................................. 199CONCLUSION ............................................................. 200ACKNOWLEDGMENTS ............................................................. 200REFERENCES ............................................................. 201
INTRODUCTION
To be successful pathogens, many organisms have devel-oped the capacity to vary phenotype not only by a strictdevelopmental cycle, but also by spontaneously generatingvariants within infecting populations whose phenotypes ap-pear to have evolved primarily to escape threatening envi-ronmental changes such as the development of immunity bythe host. Procaryotes like Salmonella typhimurium (38),Borrelia hermsii (9), and Neisseria gonorrhoeae (125) andeucaryotes like Trypanosoma brucei (29) have developedswitching systems that spontaneously generate antigenicvariants in an infecting population. Presumably, these minorvariants enrich when the host immune system targets thepredominant antigenic state. Several factors appear to haveaffected the evolution of these systems. First, becausevariable antigenic states can be based in functional mole-cules, like flagellin in S. typhimurium (38), changes cannotcompromise function. Second, because these organismspersist as pathogens, variant offspring will continue to bechallenged by the same environmental changes and thereforemust retain the capacity for phenotypic variability. There-fore, switching must be reversible or relatively unlimited inthe repertoire of variability. Third, changes must occurspontaneously and at relatively high frequency to assurerapid enough enrichment after an environmental change. Formany of these high-frequency variability systems, either aprecise molecular mechanism has been elucidated (e.g., S.typhimurium [38]) or a general notion of mechanism has beenformulated (e.g., B. hernsii [9], N. gonorrhoeae [125], and
183
T. brucei [29]) at the level of DNA reorganization (13, 17,98).The fungal pathogen Candida albicans is also capable of
high-frequency, reversible phenotypic switching (82, 94,101, 102, 108, 110, 111, 116). Cells emanating from a singleprogenitor are capable of expressing very different generalphenotypes reflected by an assortment of variant colonymorphologies, and they are capable of passing on theirvariant phenotypes to their progeny in a heritable fashion.These variant progeny appear at frequencies well above thatof point mutation and are in turn capable of generatingprogeny with different phenotypes or with the phenotype ofthe original progenitor. As is the case in other switchingsystems (9, 29, 38, 125), switching in C. albicans can affectantigenicity in a reversible fashion (1, 2, 4, 34), but it can alsoaffect many other aspects of cellular physiology and mor-phology, including a number of putative virulence traits (110,111). Switching has been demonstrated to occur at sites ofinfection (116) and to occur between recurrent episodes ofinfection in select cases (113). It has been assumed, althoughnot definitively demonstrated, that switching in Candidaspp. plays a role similar to that in other infectious microor-ganisms. It has also been assumed, based on the paradigmgenerated from other switching pathogens (17), that themechanism of switching in C. albicans involves reversiblegenetic rearrangement, although no direct evidence exists tosupport this assumption. In the discussion that follows, theswitching process in C. albicans will be reviewed, withspecial emphasis placed on gene regulation, the phenotypic
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

CLIN. MICROBIOL. REV.
consequences of switching, possible molecular mechanismsthat may be involved in the switch event, and the role ofswitching in pathogenesis.
DIMORPHISM AND SWITCHING
C. albicans and related species are dimorphic, capable ofgrowing in the predominant budding form or in a specializedhyphal form (77, 105). Although relatively little is knownabout the regulatory events involved in the bud-hyphatransition, its characteristics are quite different from switch-ing, and a careful distinction must be made, at least concep-tually, between the two forms of phenotypic variability. Inthe budding form, C. albicans is very much like diploidSaccharomyces cerevisiae (12, 42, 107, 112). However,correct environmental cues will stimulate a mature Candidacell in the budding form to generate an elongate daughtercell, which then grows in a hyphal form (77, 105). The hyphahas a tubular morphology which includes a linear sequenceof unseparated elongate cellular compartments, each with anucleus, and with little or no constriction at the sites ofseptation. Intermediate pseudohyphal forms also occur witha continuum of compartment curvature from the ellipsoidalbud shape to the tubular hyphal shape. Pseudohypha forma-tion depends on both the strain and environmental condi-tions (70, 77). Hyphal growth differs from bud growth in anumber of subtle ways, including the position of the tran-sient filament ring (117), the dynamics of general and apicalwall expansion zones (115, 121), ionic currents (122), cellseparation, and gene expression (18, 32, 66). The elongatinghypha appears to play a fundamental role in effective tissuepenetration (104). The reversible bud-hypha transition rep-resents the basic phenotypic transition in C. albicans andrelated species, and mass conversion from one phenotype tothe other can be effected by a number of environmentalmanipulations including temperature (7, 21), pH (21, 31), andthe composition of the supporting medium (10, 58, 67, 99,126).
Switching, on the other hand, occurs spontaneously, atfrequencies above that of point mutation but usually farbelow that of mass conversion (14, 87, 101, 109, 111).Switching usually is discriminated by colony morphologyand in some cases by the phenotype of cells in the buddingphase (1, 2, 4, 102, 111). More importantly, cells expressingdifferent switch phenotypes in a single strain are usually stillcapable of undergoing the bud-hypha transition, although theenvironmental constraints on the transition may differ be-tween switch phenotypes (1). Therefore, the bud-hyphatransition represents reversible cellular differentiation thatcan be rapidly induced en masse by simple environmentalmanipulation. High-frequency switching, on the other hand,is usually spontaneous. It does not occur en masse, evenafter environmental stimulation (there is one exception tothis; see references 14, 87, and 102), and appears to repre-sent a more general variability system occurring spontane-ously at lower frequency and superimposed on the basicbud-hypha transition. In the case of the bud-hypha transi-tion, all cells in a single population will express alternativephenotypes under alternative environmental conditions. Inthe case of switching, a small number of cells in a singlepopulation will express a different phenotype under a singleset of environmental conditions.
HIGH-FREQUENCY VARIABILITY: EARLYOBSERVATIONS
Variants of C. albicans exhibiting aberrant colony mor-phology were reported as early as 1932 by Negroni (74). Hedescribed a variant rough colony morphology, the R-variant,in Mycotorula albicans, an early name for C. albicans.Subsequently, Mackinnon (64) described a process of grad-ual change, or "dissociation," from a normal to spikymorphology. However, it was J. Brown-Thomsen in 1968(19) who first demonstrated that colonial variation occurredin a number of strains of C. albicans and that it affected thebasic physiology of the cell. By streaking strains on maltagar, Brown-Thomsen was able to assess colonial phenotypeand the genesis of aberrant sectors (19). He found thatmorphological differences in streaks were the result ofdifferences in the proportions of buds, hyphae, and pseudo-hyphae, as well as the result of variations in pseudohyphalphenotype. Brown-Thomsen examined more than 300strains of C. albicans and identified 15 different morpholog-ical forms. Variants were obtained by examining platesstored at room temperature over a 6-month period. A few ofthese variants differed from parent strains in their capacitiesto assimilate specific sugars and to reduce tetrazolium salts.Although Brown-Thomsen did not measure the frequency ofvariant formation, or the frequencies of reversion and inter-conversion, he was taken by the limited number of morphol-ogies in the variant repertoire of each strain. Brown-Thom-sen's study of variability (19) and Mackinnon's study ofdissociation (64) received only marginal attention in the1970s (e.g., see reference 127) and were totally neglected inthe early 1980s.
HIGH-FREQUENCY SWITCHING: REDISCOVERY
In 1985, two reports of reversible high-frequency switch-ing in C. albicans rekindled interest in the potential forphenotypic variability other than the bud-hypha transition.Pomes et al. (82) demonstrated that, in strain 1001, UVirradiation gave rise to rough segregants. They found that alow dose of UV which killed roughly 13% of the cellpopulation increased the frequency of rough colonies from<10' (0 of 12,000 colonies screened) to 3 x 10-3 (6 of 1,982colonies). The spontaneous frequency of smooth coloniesgenerated by the rough variant 1001FR was <9 x 10-' (0 of1,136 colonies screened), but the spontaneous frequency ofsectored colonies was 3.5 x 10-3 (4 of 1,136 colonies).
In another report in 1985, Slutsky et al. (101) reported thatthe common laboratory strain 3153A was capable of high-frequency reversible switching. They discriminated severalcharacteristics of switching in strain 3153A which have sincebeen demonstrated for switching systems in other strains(102). When cells of the standard laboratory strain 3153Awere clonally plated on an amino acid-rich agar (59) that waslimiting for zinc (11) and then incubated at 25°C, the pre-dominant colony phenotype was "smooth" (Fig. 1A). How-ever, variant colonies appeared spontaneously at a fre-quency of 1.4 x 10-4 (7 of 50,500 colonies screened) andincluded several variant phenotypes: "star" (Fig. 1B),"ring" (Fig. 1C), and "irregular wrinkle" (Fig. 1D). Whencells of strain 3153A were irradiated with low doses of UVlight, which killed 8% of the population, variant coloniesappeared at a combined frequency of 2.5 x 10-2 (136 of5,600 colonies), and the repertoire of switch phenotypesexpanded from the original three to include "stipple" (Fig.1E), "hat" (Fig. 1F), and "fuzzy" (Fig. 1G). When cells
184 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 185
FIG. 1. Variant colony phenotypes in the switching repertoire of C. albicans 3153A. (A) Original smooth; (B) star; (C) ring; (D) irregularwrinkle; (E) stippled; (F) hat; (G) fuzzy (note aerial mycelia on colony dome); (H) revertant smooth. See Slutsky et al. (101) for details of thisswitching system; reprinted with permission of the publisher.
from a single colony of any variant were plated, the majoritymaintained their original variant phenotype, demonstratingheritability, but a significant minority exhibited other switchphenotypes, including the original smooth morphology (Fig.1H). Figure 2A illustrates diagrammatically the minimumfrequencies of switching between phenotypes in strain
3153A. Examples of a switch from ring to star, star to ring,star to irregular wrinkle, ring to irregular wrinkle, and ring tosmooth are presented in Fig. 2B to F, respectively. Sectoringwas also evident (Fig. 3A and B), demonstrating thatswitches occurring early in colony development were stableand continued to be expressed phenotypically. The majority
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

A
FIG. 2. Switching between variant colony phenotypes of C. albicans 3153A. (A) Switching map with minimum frequencies; (B) switchfrom ring to star; (C) switch from star to ring; (D) switch from star to irregular wrinkle; (E) switch from ring to irregular wrinkle; (F) switchfrom ring to revertant smooth. In each case, cells from a colony exhibiting a variant phenotype were plated and a region with a switchphenotype (arrow) was photographed. See Slutsky et al. (101) for details; reprinted with permission of the publisher.
186
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 187
FIG. 3. Examples of sectoring in C. albicans 3153A. (A) Original smooth colony with irregular wrinkled sector; (B) ring with star sector.
of variants in the 3153A switching repertoire were in ahigh-frequency mode of switching. Some smooth revertantsmimicked the colony phenotype of the parent strain but werestill in a high-frequency mode of switching, while other
revertants exhibited both the original smooth phenotype andthe lower spontaneous switching frequency characteristic ofthe parent strain. The studies by Pomes et al. (82) andSlutsky et al. (101) demonstrate that UV stimulates switch-
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

CLIN. MICROBIOL. REV.
ing and that switching is reversible at relatively high frequency.In addition, the study by Slutsky et al. (101) demonstrates thatthere can be more than two phenotypes in a switching reper-toire, that there is interconvertibility (switching from onevariant phenotype to another) as well as reversibility, and thatthere are different frequencies of switching, which in somecases are characteristic of particular colony phenotypes.
ZINC AND COLONY PHENOTYPE IN STRAIN 3153A
Before realizing that strain 3153A possessed a high-fre-quency, reversible switching system, Bedell and Soll (11)reported that a variant of strain 3153A, M10, was aberrant inhypha induction. When the parental wild-type strain enteredthe stationary phase in liquid medium, cells accumulated asunbudded singlets, homogeneous in size and blocked in Gl ofthe cell cycle. In contrast, when M10 cells entered thestationary phase in the same medium, more than 50% formedhyphae. Hypha formation in this case was inhibited when theconcentration of zinc in the original medium was increased10-fold (11). M10 proved to be the star phenotype (Fig. 1B) inthe 3153A switching repertoire, resolvable on agar containingthe defined nutrient composition of the amino acid-rich me-dium of Lee et al. (59) supplemented with arginine and alimiting concentration of zinc (11). If the concentration of zincin this agar medium was increased 10-fold, the star phenotypewas suppressed, and the star variant formed smooth colonies.However, if cells from these high-zinc plates were once againplated on low-zinc plates, they expressed the star phenotype,demonstrating that zinc suppressed expression of the starphenotype but did not alter the star "genotype" of the cells.
This point is extremely important in the investigation ofswitching in new strains since it shows that a switch pheno-type is not a fixed state, but rather is the way in which aparticular variant generates a colony in a specific environ-ment after a particular period of time. Switch phenotypes ofstrain 3153A were assessed in the original study of Slutsky etal. (101) after 7 days at 24°C on zinc-limited, defined nutrientagar. In many cases, the phenotype of a colony changed withprolonged incubation (e.g., in excess of 7 days) or whenincubated at another temperature. Media like yeast-peptoneD-dextrose agar [8]), used in many yeast studies, are rich inzinc and suppress expression of particular colony pheno-types, but they may not affect the switching mechanism (78).Therefore, when a strain is described as a low-frequency"switcher" or a "nonswitcher," one must assess whetherthe strain was analyzed on an agar which properly allowedexpression of the switch phenotypes.
PLOIDY, MITOTIC RECOMBINATION, ANDSWITCHING
In the early 1980s, several investigators attempted toassess the ploidy of C. albicans. Olaiya and Sogin (79)demonstrated by a number of indirect determinations that C.albicans is most likely diploid, and results in support of thisidea have continued to accumulate (112). Riggsby et al. (86)confirmed this interpretation by demonstrating that DNA-DNA hybridization kinetics were consistent with diploidy,and Whelan and colleagues (129, 131) demonstrated thatstrains were heterozygous and that mitotic recombinationbetween heterozygous alleles led to homozygosity. It wasfurther demonstrated in at least one strain that balancedlethal alleles existed on homologous chromosomes (132),suggesting, at least in this case, that haploidization would belethal. Such a condition is more likely the result of no sexual
cycle rather than the reason why no sexual, or meiotic, cyclehas been demonstrated in any bona fide C. albicans strain.There have been several suggestive, but ultimately uncon-firmed, reports of haploid strains (e.g., see reference 80).The discovery that C. albicans is diploid and heterozygous
for particular genes and the demonstration of mitotic recom-bination (130, 131) led to initial suggestions that high-fre-quency variant phenotypes may in some cases be due tomitotic recombination. Pomes et al. (82) suggested that therough variant 1001FR was the result of a mitotic recombina-tion leading to homozygosity of a recessive allele for therough phenotype, and Soll initially suggested that the veryfrequently occurring variant phenotype M10 (11) was theresult of mitotic recombination leading to homozygosity of arecessive allele for the M10 phenotype (106). Mitotic recom-bination could indeed result in the genesis of morphologicalmutants at high frequency, but these mutants, by definition,would be homozygous for genes responsible for the variantphenotype and would therefore be extremely stable, i.e.,capable of reverting to the original phenotype only through adominant point mutation or through a recessive point muta-tion followed by mitotic recombination. The high frequen-cies of reversion, the existence of multiple phenotypes, andthe high frequency of interconvertibility observed for the C.albicans switching systems are therefore inconsistent with amechanism of mitotic recombination.
WHITE-OPAQUE TRANSITION
Immediately after discovering reversible high-frequencyswitching in C. albicans 3153A, Soll and coworkers tested anumber of commensal strains isolated from the oral cavities ofhealthy young college students (118), pathogenic strains iso-lated from the vaginal canals of patients suffering from vaginalcandidiasis (116), and strains isolated from the blood of bonemarrow transplant patients (102, 119). When large numbers ofcells from each strain were clonally plated on agar, themajority of strains exhibited high-frequency reversibleswitching, but surprisingly, the repertoire of variant colonyphenotypes differed between strains on the same zinc-limit-ing, amino acid-rich agar used to characterize switching instrain 3153A (101). It soon became clear from these studiesthat there were several switching systems which differ in therepertoire of switch phenotypes (variant colony morpholo-gies) and that each strain appeared to be capable of expressingonly one of these systems. A switching system that includeda predominant transition between a relatively unmyceliatedcolony and a colony surrounded by a massive halo of myceliain the agar was found to be common in vaginal strains (116).Another switching system included a predominant transitionbetween normal-size and petite colonies (98). However, themost extraordinary switching system was observed in strainWO-1, which was isolated from a bone marrow transplantpatient at the University of Iowa Hospitals and Clinics inIowa City in 1985 (102).When the original cultures from the blood and lungs of this
patient were clonally replated on defined amino acid-richnutrient agar and incubated at 25°C for 7 days, two distinctmorphologies were apparent: a predominant smooth whitecolony and, less frequently, a flat grey colony, which wasoriginally labeled "opaque" because of the way in which itrefracted light under the lighting conditions used in theoriginal study (Fig. 4A). Believing that white and opaquecolonies from the original sample represented differentstrains, individual cells from representative white andopaque colonies were plated. Both white and opaque cells
188 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 189
FIG. 4. Variant colony phenotypes in the switching repertoire of C. albicans WO-1, which exhibits the white-opaque transition. (A) Switchfrom white to opaque (Op); (B) switch from opaque to white (W); (C) white colony with an opaque sector; (D) opaque colony with a white sector;(E) irregular wrinkle colony; (F) irregular wrinkled colony with a white sector; (G) irregularwrinkled colony next to white colony with an opaquesector; (H) medusa colony; (I) opaque colony with a medusa sector; (J) switch from opaque to white and medusa (M); (K) fried-egg and opaquecolonies; (L) fried-egg (FE) sector in an opaque colony. See Slutsky et al. (102) for details; reprinted with permission of the publisher.
gave rise to a minority of colonies of the opposite phenotype(see Fig. 4B for an example of an opaque to white switch) aswell as to an irregularly wrinkled phenotype (Fig. 4E, F, andG), a "medusa" phenotype (Fig. 4H, I, and J), and a "friedegg" phenotype (Fig. 4K and L). When cells from whitecolonies were plated, 87.10% (21,549 of 24,740) gave rise tocolonies with the original white phenotype; 6.40% (1,583 of24,740), to colonies with the opaque phenotype; 6.40%(1,584 of 24,740), to sectored colonies (Fig. 4C and D); and0.06% (15 of 24,740), to colonies with either the medusa orthe fried-egg phenotype (102). When cells from opaque
colonies were plated, 77.90% (13,502 of 17,332 coloniesscreened) gave rise to colonies with the opaque phenotype;17.80% (3,085 of 17,332), to colonies with the white pheno-type; 4.20% (728 of 17,332), to sectored colonies; and 0.13%(23 of 17,332), to colonies with either the medusa or thefried-egg phenotype (102). When cells from wrinkled colo-nies were plated, 0.30% (49 of 16,300) gave rise to colonieswith the white phenotype; 0.70% (114 of 16,300), to colonieswith the opaque phenotype; 0.13% (45 of 34,600), to colonieswith either the medusa or the fried-egg phenotype; and 0.89%(308 of 34,600), to sectored colonies. These results demon-
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

CLIN. MICROBIOL. REV.
strated that this strain, subsequently named WO-1 for white-opaque 1, possessed a switching system dominated by a phasetransition between a white and opaque colony-forming phe-notype, with three additional minor phenotypes in its switch-ing repertoire. As in the case of the 3153A switching system,the white-opaque transition is perfectly reversible (4) and thedifferent variant phenotypes are interconvertible.The white-opaque transition is an extremely stable and
manageable switching system. Unlike the switching systemsinvolving complex patterning in the colony dome (e.g., thesystem in strain 3153A), the white and opaque colony mor-phologies are easily distinguishable even after short periods ofincubation on many different nutrient agars, and the differentcell types can be differentially stained in the colony by anumber of different vital dyes including fast green, methylenered, bismuth indicator medium (87), and phloxine B (4).
Measuring the Frequency of Switching
In the first studies of C. albicans switching published in themid-1980s (82, 101, 102), data on the frequency of variantcolony formation were presented, but these data must not beconstrued as measurements of rates of switching. In theseoriginal experiments, cells from either a parental or a variantcolony were clonally plated, and the resulting colonies withthe original and alternative phenotypes, or sectors, werecounted. These measurements provided only the proportionsof cells in the original colony that gave rise to other pheno-types in the switching repertoire. Such a measure is affectedby (i) differential rates of switching, (ii) differential rates ofreversion or interconversion, (iii) differential viability in thecolony dome, (iv) differential rates of cell division, and (v)differential rates of spreading on agar by the different switchphenotypes. To obtain a more direct measure of rate, Rik-kerink et al. (87) isolated single cells on agar blocks and theninoculated each cell into 100 ,ul of nutrient medium. After 24h at 25°C, cells from each microculture were plated on agarand incubated for 4 days, and the number of original culturesthat did not generate a colony of opposite type (i.e., withouta switch) was measured. Using the Luria-Delbruck fluctua-tion formula, they estimated the frequency of spontaneousswitching at 25°C in the white to opaque direction to be lessthan 10- and that in the opaque to white direction to be 5 x10-4. In another approach to determining switching rates,Bergen et al. (14) employed the distinctive difference in whiteand opaque cell shapes (102) to measure the frequency ofswitching at the single cell level at 25°C. Individual white andopaque cells were placed on nutrient agar films either on thebottom of petri dishes or on glass slides so that they could bemonitored at relatively high magnification with an invertedmicroscope equipped with a long-working-distance lens andcondenser. As each microcolony developed, daughter cellswere counted and cellular phenotypes were scored. In a totalof 17,441 white microcolonies containing 2,083,160 cells, theyobserved 73 opaque cells. In a total of 645 opaque microcol-onies containing 9,624 cells, they observed 1,148 white cells.Bergen et al. (14) developed a mathematical model for switch-ing at the cellular level that subtracts mother cells andswitched daughter cells from the pool of switching candi-dates. The probability of a mother cell producing a daughtercell of opposite phenotype was determined by the formula:
/2 T(T S l/ks a = 2 -
where T is the final number of cells in the two phases, S is the
final number of switched cells, M is the number of mothercells, and k is the calculated number of generations. In asingle experiment, the probability aEXE. was calculated fromthe individual a values for analyzed microcolonies weightedfor the number of cells in each microcolony. The formulafor OtEXp was OtEXp = (0LlT1 + ... + atnTn)/(T1 + * * * + T),where n is the number of microcolonies, ai (1 s i c n) is theprobability of switching in each microcolony, and Ti (1 < i <n) is the final number of cells in each microcolony. Finally,the mean probability, axM, was determined for a number ofexperiments by computing the mean of OEXp values weightedfor the final number of cells in each experiment. The aoM foran opaque to white switch was calculated to be 1.0 x 10-1,and the oLM for a white to opaque switch was 1.7 x 10-5 (14).
Environmental Parameters which Affect Switching
Both Rikkerink et al. (87) and Bergen et al. (14) found thatthe frequency for switching from white to opaque was lowerthan that for switching from opaque to white. However, theirmeasurements differed and should not be construed asabsolute, not only because the former was based on finalcolony phenotype while the latter was based on early cellularphenotype, but also because it has been demonstrated that anumber of environmental factors can affect frequency (14,55, 73, 82, 87, 101, 102). In C. albicans 1001 (82) and 3153A(101), low doses of UV stimulate switching. The same is truefor the white-opaque transition in strain WO-1 (73). Morrowet al. (73) found that, when white cells from suspensioncultures were plated on nutrient agar, the frequency ofopaque colonies was less than 4.6 x 10'. When cells fromsuspension cultures were treated with low doses of UV,killing between 11 and 17% of cells, the frequency of opaquecolonies was stimulated to a mean of 4.4 x 10', roughly a10-fold increase. In addition, the proportion of white colo-nies with opaque sectors increased dramatically.UV stimulation has been demonstrated to be bidirectional
(73). When opaque cells from suspension cultures were platedon nutrient agar, the frequency of white colonies was 2.1 x10-2. When cells from suspension cultures were treated withlow UV irradiation, killing between 35 and 60% of cells, thefrequency of white colonies was 1.3 x 10-1, roughly a 20-foldincrease. Again, the proportion of opaque colonies with whitesectors increased dramatically. Stimulation of sectoring dem-onstrates that the perturbation caused by the short period ofUV irradiation persists through many cell divisions. In otherwords, an instability caused by irradiating a single cell isexpressed many generations later as an increase in the prob-ability of a switch to the alternative phenotype.
In addition to UV irradiation, temperature also affects thefrequency of switching in the white-opaque transition.Slutsky et al. (102), when first describing the white-opaquetransition, presented evidence that both high and low incu-bation temperatures affected the proportion of white CFU inopaque colonies. They found that, when opaque colonieswere incubated at 4°C for 7 days, the proportion of whiteCFU (tested at 24°C) increased from 0.9 to 44.3%, and whenopaque colonies were incubated for 1 day at 370C, theproportion of white colonies increased to 91.3%. In a sub-sequent study, Rikkerink et al. (87) found that incubation ofopaque cells in nutrient medium for >5 h at 37°C stimulatedmass conversion to white CFU. These results demonstratedunequivocally that high temperature stimulated opaque cellsto switch to white CFU. Bergen et al. (14) subsequentlydemonstrated the same minimum 5-h induction period when
190 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 191
individual white cells were incubated on agar at 37°C andswitching was assessed at the cellular level.
In addition to UV and temperature, colony age alsoappears to affect switching. When white colonies are incu-bated for >10 days on defined nutrient agar containingphloxine B to visualize sectoring, there is a definite increasein the number of opaque sectors at a colony periphery (111).However, it has not been determined in this case whetherthe increase in sectors at the colony periphery is due to anincrease in switching frequency or to the differential rate ofgrowth of opaque cells at the colony periphery. Wrapping adish with Parafilm also stimulates sectoring at the whitecolony periphery (111). Finally, Kolotila and Diamond (55)have demonstrated that neutrophils and oxidants such asH202 stimulate switching in the white to opaque direction,an observation that will be returned to when the role ofswitching in pathogenesis is considered.
Effects on Cellular Morphology
The variant colony phenotypes in the switching reper-toires of strains 3153A and 1001 are due at least in part to theproportion and distribution of budding cells, hyphae,pseudohyphae, and branched hyphae in the colony dome(82, 101). However, in the white-opaque transition, thedifference between the white and the opaque colony pheno-type is due to a most unusual difference in the basicphenotype of the budding cell (102). Budding white cells areround to slightly ellipsoidal (Fig. 5A), and the buddingpattern is similar to that of other standard strains of C.albicans (12, 105, 112) and diploid strains of Saccharomycescerevisiae (33). In marked contrast, budding opaque cells areelongate and bean shaped, with one end sometimes blunt andwider than the other (Fig. 5B). The pattern of budding isaltered in opaque cells, with new daughter cells formingusually eccentrically at one end, and in many cases in abipolar fashion (Fig. 5C). In addition to a budding opaquecell population exhibiting very different shapes, its mean cellvolume is 114 ,um3 and the average roundness measurement(width divided by length) is 0.6, while the mean cell volumeof a budding white cell population is 33 ,m3 and the averageroundness measurement is 0.9 (4,102). The dry weight ofbudding opaque cells is also roughly twice that of buddingwhite cells (102). However, the DNA contents of opaque andwhite cells are roughly similar. Opaque cells in the mid-logphase of growth contain 6.4 ,ug of DNA per 108 cells, whilewhite cells contain 5.7 ,ug of DNA per 108 cells (102). Bothwhite and opaque cell populations are predominantly mono-nucleate and exhibit identical chromosome karyotypes re-solved by transverase alternating field electrophoresis (seelater discussion and Fig. 9). Therefore, the difference in sizeand shape between budding white and opaque cells are notthe result of a change in ploidy.
Perhaps the most extraordinary difference in cellular phe-notypes was observed when the surface of white and opaquecells were compared by scanning electron microscopy (1, 2,4). The surface of budding white cells was relatively smooth(Fig. 5D), like that of other C. albicans strains, but thesurface of opaque cells exhibited a unique punctate morphol-ogy (Fig. SE to H) which, as far as we are aware, has neverbefore been observed in yeast cells. The surfaces of matureopaque cells possess pimples with pits from which blebsfrequently emerge (Fig. SE to H). The average number ofpimples per cell has been estimated to be 141 (+39 [standarddeviation]; n = 6) (2). It is not clear in what state the surface
morphology actually exists in living cells, since it is possiblethat fixation may alter the natural punctate surface morphol-ogy. However, there is no question that some form ofpunctate pattern exists on the surface of mature buddingopaque cells that is absent on the surface of budding whitecells. Interestingly, the punctate pattern is not evident untilan opaque cell has grown to more than half of its final volume(4).The presence of "pimples" in the budding opaque cell
wall was also demonstrated by transmission electron micros-copy (TEM) (2). Cross sections of the white cell wall (Fig.6A) are similar to those of other budding strains of C.albicans. No indication of pimpling is evident. In contrast,cross sections of the opaque cell wall (Fig. 6B) containpimple structures, in some cases with channels traversingthe pimple wall and in many cases with membrane-boundvesicles emerging from the pimple apex (Fig. SI, J, and K).The mean diameter of emerging vesicles was computed fromTEMs of three different cells to be 146 nm (+ 16.5 [standarddeviation]) (2). In rare instances, flask-shaped vesicles wereobserved just under pimple structures (Fig. SL).
In addition to the difference in wall morphology, TEMcomparisons also demonstrated a difference in cytoplasmicmorphology. TEM profiles of budding white cells (Fig. 6A)were indistinguishable from those previously published forother strains of C. albicans (16, 49) and Saccharomycescerevisiae (22). However, profiles of budding opaque cellscontained giant vacuoles with spaghettilike material fillingthe vacuole interior (Fig. 6B). The giant vacuoles in opaquecells were initially misinterpreted as expanded cell nuclei(82). However, it was subsequently demonstrated that thesevacuoles pressed the remaining cytoplasm and other organ-elles, including the real cell nucleus, into a cytoplasmiccortex, and that the spaghettilike material in the vacuoleprobably represented vesicles which had collapsed duringfixation (2). The function of fungal vacuoles is extraordinar-ily complex (54), and the significance of an expanded vacu-ole with resident vesicles in opaque cells is as much amystery as the function of the wall pimples.
Relationship between Hypha Formation and theWhite-Opaque Transition
As noted above, switching affects the environmental con-straints on the bud-hypha transition in strains 3153A and1001 (82, 101), and the same is true for strain WO-1 (4).However, there is reason to believe that hypha-specificfunctions and hypha-specific gene expression may also playa direct role in the genesis of the unique opaque cellphenotype. To begin with, Anderson and Soll (3) firstdemonstrated that the pattern of actin granule distributiondiffered between growing buds and growing hyphae in strain3153A. In expanding buds, actin granules are distributedevenly throughout the cortex, while in elongating hyphae,actin granules are localized in the apex. Anderson and Soll(4) subsequently demonstrated that the pattern of actingranule distribution is budlike during early expansion of anopaque bud and then hyphalike during later expansion.These results suggest that, in the genesis of a mature buddingopaque bud, budding and hypha-forming mechanisms func-tion in temporal sequence (111). As we shall see later in thisreview, select hyphal genes may indeed be expressed inbudding opaque cells. However, opaque cells do make bonafide hyphae even though the constraints on the bud-hyphatransition differ markedly from those for white cells (1).
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

192 SOLL CLIN. MICROBIOL. REV.
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 193
More interesting, opaque cell hyphae do not possess pimples(Fig. 5H) and in all morphological respects are indistinguish-able from hyphae formed by white budding cells. Opaquecell hyphae are still in the opaque phase since, when opaquehyphae are induced to form buds, the phenotype of thedaughter cells is opaque, not white (1).Hypha formation also appears to play a fundamental role
in the transition from the white to the opaque cell phenotype.To describe the white to opaque and opaque to whitetransitions at the cellular level, Bergen et al. (14) separatedlarge numbers of individual opaque and white cells on agarand then followed their early budding patterns as theyproliferated into microcolonies. When opaque cells gave riseto white daughter cells at 24°C, the transition was direct. Anelongate opaque cell gave rise directly to an ellipsoidal whitecell. In contrast, when white cells gave rise to opaque cells,a pseudohyphal or hyphalike transition phenotype was firstgenerated by the white cell, and these transition cells thengave rise to morphologically legitimate opaque cells. Theseresults suggest that, in the switch from white to opaque,hypha-specific genes may be transiently recruited in thephenotypic transition.
Antigenic Variability
As noted in the introduction, most of the switching sys-tems which have been uncovered in infectious microorgan-isms involve antigenic variability at the cell surface (9, 13,17, 29, 38, 92, 125), and the white-opaque transition in C.albicans WO-1 is no exception (2, 4). When a rabbit antise-rum generated against opaque cell extract was used to stainthe surfaces of cells fixed but still impermeable to antibody,the antiserum stained the surfaces of budding white cells(Fig. 7A), hypha-forming white cells (Fig. 7B), buddingopaque cells (Fig. 7C), and hypha-forming opaque cells,demonstrating that opaque cells contain one or more anti-gens common to the surfaces of buds and hyphae in both thewhite and the opaque phases (1, 4). When this antiserum wasabsorbed with nonpermeabilized budding white cells, theabsorbed antiserum was no longer capable of staining thesurfaces of nonpermeabilized white budding cells or whitebudding cells giving rise to hyphae, but it was still capable ofstaining the surfaces of white hyphae (Fig. 7D) and buddingopaque cells, demonstrating that the surfaces of opaque cellsalso contain one or more hyphal antigens (1, 4). When thisantiserum was finally absorbed with nonpermeabilized whitehypha-forming cells, the absorbed antiserum was still capa-ble of staining opaque cells and did so in a punctate fashion(Fig. 7E and F), demonstrating that an opaque-specificantigen was present on the opaque cell surface and that thisantigen was localized in a punctate fashion similar to thepimple pattern discerned by scanning electron microscopyand TEM.When the original anti-opaque serum was absorbed with
nonpermeabilized budding white cells and used to stainWestern blots (immunoblots) of budding white and buddingopaque whole-cell extracts, three major antigens were found
to be opaque specific (2) (Fig. 7K2). The molecular masses ofthe three antigens were estimated to be 14,500, 21,000, and31,000 daltons. To determine which of the three opaque-specific antigens was located on the cell, the antiserum wasalso absorbed with nonpermeabilized opaque cells, render-ing it noninteractive with the opaque cell surface (Fig. 7Gand H). This absorbed antiserum was used to stain aWestern blot containing budding white and budding opaquewhole-cell extracts (Fig. 7K3). Only antibodies to the 14.5-kDa antigen were removed (Fig. 7K3), demonstrating thatonly the 14.5-kDa opaque-specific antigen is localized on theopaque cell surface. To demonstrate that the 14.5-kDaantigen is localized in pimples, Anderson et al. (2) firsttreated nonpermeabilized budding opaque cells with anti-opaque serum which had been absorbed with budding whitecells and then treated the opaque cells with goat anti-rabbitserum conjugated to colloidal gold. When these cells wereprocessed for TEM, immunogold was found localized exclu-sively at pimple apices (Fig. 7I and J), demonstrating that theopaque-specific 14.5-kDa antigen is localized in the apex ofthe unique opaque wall pimple (2), although not excludingthe possibility of intracellular localization as well.
GENE REGULATION AND SWITCHING
The identification of opaque-specific antigens (1, 2, 4)suggested that switching in C. albicans involves differentialgene expression, and a number of recent studies, bothindirect and direct, support this conclusion. A switch fromwhite to opaque in strain WO-1 results in the loss of thecapacity to assimilate four sugars, and a switch from opaqueto white reestablishes this capacity (109), suggesting thatswitching may have a dramatic effect on gene regulation.When mid-log-phase cultures of budding cells from the whiteand opaque phenotypes of strain WO-1 were pulse-labeledwith [3 S]methionine and polypeptides were extracted andseparated by two-dimensional polyacrylamide gel electro-phoresis, 5 of more than 200 labeled and reproduciblyresolvable polypeptides differed between the two pheno-types (111). Two were specific to the opaque phase, one wasspecific to the white phase, and two changed isoelectricfocusing points between the two phases (111). Unfortu-nately, two-dimensional-polyacrylamide gel electrophoresisis by no means a definitive test of differential gene expres-sion, since (i) only the most abundantly synthesized and/orstable polypeptides are resolvable and (ii) differential syn-thesis in many cases cannot be distinguished from differen-tial posttranslational modification.A more definitive proof that differential gene expression
occurs in the white-opaque transition has been establishedwith the cloning of cDNAs for messengers which are phasespecific. Morrow et al. (73a) performed a differential screenof an opaque cDNA library and isolated several opaque-specific cDNAs distinguishable by differences in transcriptsize. The first opaque-specific gene identified, Op27a, wastranscribed in opaque but not in white cell cultures grown in
FIG. 5. Differences in cell morphology of the white budding and opaque budding cells of C. albicans WO-1. (A) Budding white cells fromthe dome of a colony; (B) budding opaque cells from the dome of a colony; (C) opaque cell exhibiting bipolar budding; (D) scanning electronmicrograph of white budding cell with daughter bud and bud scar (arrowhead); (E, F, G) scanning electron micrographs of budding opaquecells with pimple structures (pimple apices are indicated by arrows); (H) opaque cell with hypha (H); (I, J, K) small vesiclelike structurespresumably emerging from or located at the apices of pimples (noted by arrows); (L) a vesicle positioned under a pimple with a shaft traversingthe pimple center. See Anderson et al. (1, 2) for details; reprinted with permission of the publisher.
VOL. S, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

CLIN. MICROBIOL. REV.
A
FIG. 6. TEMs of white (A) and opaque (B) budding cells. Note the relatively constant morphology of the white cell wall and the pimpledwall of opaque cells. (B) A pimple with an emerging vesicle and complex intrapimple morphology (arrow) and a vesiclelike structure on thewall (arrowheads) are indicated. Also note the giant vacuole filled with spaghettilike material (presumably collapsed vesicles) in the opaquecell in panel B. See Anderson et al. (2) for details; reprinted with permission of the publisher.
defined liquid medium (Fig. 8). It was also not transcribedduring hypha formation in white cell cultures (Fig. 8). Thesequence of the Op27a cDNA exhibited 99.1% homologywith the excreted acid protease gene cloned by Hube et al.(48) from strain ATCC 10231 and contained (i) all but the first15 bases of the open reading frame; (ii) the hydrophobic
signal sequence at the 5'-end, including the lysine residue atamino acid position 36, a characteristic of many pepsinogens(25); and (iii) two catalytic aspartyl domains which combinein the folded molecule to generate the active site (25). Theaddition of serum proteins to opaque cells resulted in littleadditional stimulation of Op27a transcription (73a).
194 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 195
H
-c31kd
21l4kd
FIG. 7. An opaque cell-specific antigen resides in the pimple apex. An antiserum was generated against total opaque cell extract. (A)Nonpermeabilized budding white cell (MC) with medium-sized bud (B) immunostained with unabsorbed antiserum; (B) nonpermeabilizedhypha-forming white cell with long hypha (H) immunostained with unabsorbed antiserum; (C) nonpermeabilized opaque cell (OC) with smallband (OB) immunostained with unabsorbed antiserum; (D) nonpermeabilized budding white cells (MC) with hyphae (H) immunostained withantiserum absorbed with nonpermeabilized budding white cells; (E) nonpermeabilized budding opaque cell immunostained with antiserumabsorbed with nonpermeabilized hypha-forming white cells; (F) phase-contrast image of cell in panel E; (G) nonpermeabilized budding opaquecell stained with antiserum absorbed with nonpermeabilized opaque cells; (H) phase-contrast image of cell in panel G; (I, J) indirectimmunogold staining of opaque cell pimples with absorbed antiserum; (K) Western blot demonstrating that the 14.5-kDa antigen is presenton the budding opaque cell surface and that the opaque cell-specific 21- and 31-kDa antigens are absent from the surface. Lanes 1,Amido-black-stained blots of white cell (W) and opaque cell (Op) extracts; lanes 2, blots of white and opaque cell extracts stained withantiserum absorbed with nonpermeabilized white budding cells; lanes 3, blots of white and opaque cell extracts stained with antiserumabsorbed first with nonpermeabilized budding white cells and then with nonpermeabilized opaque cells (note that only antibodies to the14.5-kDa antigen are removed). See Anderson et al. (2, 4) for details; reprinted with permission of the publisher.
Srikantha and Soll (120a) performed a differential screen vation and deactivation of phase-specific genes in both theof a white cDNA library subjected to subtraction hybridiza- white to opaque and the opaque to white directions. Regu-tion and cloned a white-specific cDNA. These results dem- lation appears to be at the level of transcription, but nuclearonstrate that the white-opaque transition involves the acti- mun-on experiments will have to be performed to substantiate
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

196SOLL~~~~~~~~~~~~~~~~~~~~~CLIN.MICROBIOL. REV.
A. flfl~c B. .0 m
FIG. 8. Demonstration of opaque-specific transcription of gene
0p27a, which encodes for the excreted acid protease. A Northern
(RNA) blot of opaque budding (Op.b.), white budding (Wt.b.) and
white hypha-forming (Wt.h.) cells, stained with ethidium bromide
(A) was probed with the opaque-specific 0p27a cDNA (B). Note
that only a single transcript hybridized with 0p27a cDNA. st.,
molecular weight standards. See Morrow and Soll (73a) for details.
this conclusion (24). These results also underscore the need
to distinguish between genes involved in the actual switch
event and genes regulated by the switch event.
MECHANISM OF SWITCHING
A number of general characteristics have been determined
for C. albicans switching, including (i) very high frequen-
cies, (ii) heritability, (iii) reversibility and interconvertibility,
(iv) environmental stimulants (e.g., UV and temperature),and (v) high and low modes of switching (14, 82, 87, 101, 102,
110, 111). Unfortunately, none of these characteristics pro-
vides clues to the actual mechanism. In a first attempt to
obtain some insights into the switching mechanism, South-
ern blots of EcoRI-digested DNA of C. albicans were probedwith the mating-type locus Mata (52) and the transposableelement Ty of Saccharomyces cerevisiae (15). The rationale
for this was the observed homology between a number of C.
albicans and Saccharomyces cerevisiae genes (56). Indeed,
cloning of the majority of C. albicans genes to date has been
accomplished through Saccharomyces cerevisiae mutant
rescue (56). Therefore, it was hypothesized that C. albicans
may have recruited a variability system already demon-
strated in Saccharomyces cerevisiae to accomplish high-
frequency switching in its evolution to a pathogenic state.
However, no homologous sequences were observed even at
low stringency to either Mata or Ty (109). In a second
approach, moderately repetitive genomic sequences were
cloned and tested for reversible reorganization correlatingwith phenotypic switching. Scherer and Stevens (95) cloned
and characterized a sequence which they named 27A, and
Hicks, Soll, and coworkers (93, 96, 113, 116) cloned and
characterized a sequence which they named Ca3. Both
turned out to contain sequences of a moderately repetitivegene family distributed among most of the chromosomes ofC. albicans (93, 95). Reorganization of the Ca3, or 27A,sequences were observed at reasonable frequencies (95), butthese changes did not correlate with rapid phenotypicswitching (95, 113). A second moderately repetitive se-quence, Ca, was found to be subtelomeric (93) and highlyrecombinational in some strains (93, 116), but again genomicrearrangements of the Ca7 sequences did not correlate withphenotypic switching.
It is clear from the results thus far accumulated onswitching in C. albicans that, whatever the mechanism, it ismost likely reversible, in a fashion similar to the conserva-tive phase transitions in S. typhymurium (38) and Esche-richia ccli (38) and mating-type switching in Saccharomycescerevisiae (52). Indeed, switching has been followed in strainWO-1 through more than 50 sequential transitions betweenwhite and opaque with no noticeable effect on subsequentswitching frequencies, colony morphology, or cellular phe-notype. Rustchenko-Bulgac et al. (91) suggested that spon-taneous variant phenotypes in C. albicans may be the resultof chromosomal rearrangements identifiable by orthagonalfield alternating gel electrophoresis and transverse alternat-ing field electrophoresis. However, in this study no experi-ments were performed in which reversible chromosomalrearrangements were demonstrated in a sequence in which astrain switched back and forth several times between twoalternative colony morphologies, as in the white-opaque(102) or unmyceliated-heavily myceliated (116) phase transi-tions. Chromosomal rearrangements do occur in C. albicansat frequencies high enough to cause significant karyotypePolymorphisms between strains (65, 71, 72, 103), but there isno convincing evidence that such rearrangements are thebasis of reversible phenotypic switching or that they canoccur over and over again in a perfectly reversible fashion, arequirement for a phase transition.
Suzuki et al. (124) described a strain of C. albicans,NUM961, which generated a variant semirough colony onyeast-peptone D-dextrose agar at a frequency of 3 x 1O'.The DNA content of the NUM961 parental strain wasroughly one-third higher than that of a standard strain of C.albicans, and the DNA content of a semirough variant(SRT-1) was roughly twice that of a standard strain. Suzukiet al. (124) concluded that the parent strain was near-triploidand that SRT-1 was near-tetraploid. Pulse field gel electro-phoresis suggested differences in banding patterns bothbetween the parent and SRT mutant and between differentrevertant smooth strains. Again, these results suggest thatchromosomal rearrangements occur frequently in C. albi-cans and seem to be associated in a random fashion withmorphological variants and revertants, but they fail to pro-vide a cause-effect relationship. A standard transverse alter-nating field electrophoresis gel is presented in Fig. 9A for awhite-opaque-white-opaque switching sequence, and a sec-ond gel with expanded lanes run for a longer period forhigher resolution is presented in Fig. 9B for a white-opaquesequence. Both gels possess a lane with separated Saccha-romyces cerevisiae chromosomes for reference. No changesin karyotype are evident in the C. albicans lineages.What, then, is the mechanism for switching in C. albi-
cans? All of the switching mechanisms in pathogenic organ-isms which have been elucidated involve genomic rearrange-ments (13, 17, 98). Although the search for mobile DNAsequences has by no means been exhausted, it has not yetprovided us with a switching mechanism. In addition, South-ern blots probed with opaque-specific cDNAs have not
196 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 197
A. B._¢o 3 o
FIG. 9. Transverse alternating field electrophoresis separation ofchromosomes of C. albicans WO-1 in switching sequences. (A)Separation in a white (Wt.)-opaque (Op.)-white-opaque sequence;(B) separation in a white-opaque transition. S.c., saccharomycescerevisiae.
uncovered associated genomic rearrangement, and Southernblots of white and opaque DNA digested with 14 differentendonucleases and probed with the opaque-specific Op27acDNA demonstrated atbsolutely no polymorphism (73a). Ittherefore seems more prudent at this point in the search fora mechanism to identify genes involved in the actual switchevent. Goshorn and Scherer (39) fused cells of strain 655, a
high-frequency switcher with a wrinkled colony morphologyon yeast-peptone D-dextrose agar, with cells of stains 1006and 981, smooth strains with apparently low switchingfrequency on yeast-peptone D-dextrose agar. They foundfusants with the origihal wrinkled 655 phenotype, with theoriginal smooth 1006 phenotype, and with an altered wrin-kled phenotype. They also obtained fusants with "mixed"phenotype, an unstable phenotype producing both smoothand wrinkled colonies. They concluded from these experi-ments that the wrinkled phenotype is recessive to smoothbecause they obtained only one 655-type wrinkled pheno-type out of 34 fusants, While the rhajority of fusants fell intothe mixed category of high-frequency switchers (39). How-ever, if Goshorn and Scherer had combined wrinkled andaltered wrinkled (intermediate) phenotypes into one cate-gory, they may not have made this conclusion since thenumber of fuSants in the combined category would havebeen greater than the category of smooth phenotypes.Scherer and Magee (94) subsequently used these data toconclude that switching was recessive, but in fact half of allfusants (17 of 34) were placed in a mixed category of strainsproducing wrinkled and smooth colonies at high frequency.Although Goshorn and Scherer (39) suggest that the mixedphenotype is the result of selective chromosome loss, it isjust as reasonable to conclude that fusion may have in factstimulated switching. Subsequent plating experiments atidgenetic analysis of the different phenotypes generated inmixed strains which would have distinguished between thesealternatives were not performed. However, experiments byChu and Magee were described in a review by Scherer andMagee (94) in which fusants obtained from strain WO-1 andstable strains (i.e., unrelated strains which appeared to below-frequency switchers on zinc-containing yeast extract-peptone-dextrose medium) were stable and did not generateopaque colonies and heat shock-induced loss of chromo-somes resulted in the return of the white-opaque transition.
These results suggest that strains with stable phenotypes onyeast extract-peptone-dextrose medium contain a genewhich represses either the white-opaque transition (theability to switch) or the expression of the opaque phenotype.The gene necessary for expression of the opaque phenotypeand the URA3 gene appear linked, and repressor functionhas been excluded from chromosome 3, which contains theURA3 gene (94). Finally, Gil et al. (37) identified threemorphological mutant phenotypes which were suppressed infusions with wild-type strains. These three sets of fusionexperiments represent the first published attempts to identifygenes involved in phenotypic switching, but the results ofsuch experiments must be interpreted with some caution.First, definition of a "nonswitching" strain can be suspect ifswitching has been assessed on agar high in zinc, as dis-cussed in a previous section. Second, many of the fusionexperiments have, been performed with genetically differentstrainis, so controls are essential in which a switching strainwith the same genetic background as the nonswitching strainis used as a fusant partner to the switching strain beinganalyzed. Finally, it should be kept in mind that there is verylikely a difference between the switch event and the capacityto express switch phenotypes. Mutations or suppression ofeither would give rise to the same incapacity to express aparticular switch phenotype.Although most speculation on the mechanism of the
switch event still focuses on reversible genomic rearrange-ments, epigenetic mechanisms must also be considered(108). Sonneborn first demonstrated that the complex asym-metric pattern of ciliary rows on Pardmecium was some-times altered by aberrant cytokinesis and that these alter-ations were heritable even though no changes had occurredin the micronucleus (120). Such a non-nuclear, hereditarychange appears to depend on the role of an establishedpattern in the genesis of a subsequent pattern during dupli-cation of ciliates. Since yeasts do not possess the complexcortical organization of protozoans, it seems unlikely thatcortical inheritance plays a major role in yeast morphogen-esis or that this form of heritable change provides a modelfor switching. A more applicable epigenetic model may befound in heterochromatization. In female mammalian em-bryos, heterochromatization of one of the two X chromo-somes occurs randomly, and this random decision remainsfixed through subsequent cell divisions (62). Recently, heri-table epigenetic changes have been demonstrated in Saccha-romyces cerevisiae (81). The SIR (silent information regula-tor) genes in Saccharomyces cerevisiae are involved inestablishing the silent state and the SIR2, -3, and -4 genes areinvolved in maintaining the silent state (81). The mutationsirl-1 identified the SIRI gene and was found to be leaky(88). The sirl-i mutation resulted in a cell population geno-typically homogeneous but phenotypically heterogeneousfor the repression of HMLct, the silent locus for mating-typea. By single-cell analysis, it was demonstrated that alterna-tive phenotypes persisted through multiple generations,demonstrating heritability of the alternative epigenetic states(81). The probability of a switch from the derepressed totepressed state has been estimated to be roughly 4 x l0-3per cell division, or once per 250 cell divisions (81). It hasalso been demonstrated that the SIR2 gene is involved in thesuppression of recombination both for the silent mating-typeloci and between ribosomal genes (40, 53). Perhaps the mostrelevant observations involving epigenetic states and theSIR genes in Saccharomyces cerevisiae were recently re-ported by Gottschling and coworkers (5, 41). They foundthat, when the ADE2, URA3, TRPI, or HIS3 gene was
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

CLIN. MICROBIOL. REV.
placed close to a telomeric region, transcription was re-pressed, but expression switched between repressed andexpressed states, and these states were heritable throughmitosis. When clonal colonies were stained for expression ofthe ADE2 gene, sectoring was observed (5, 41). Therefore,genetically equivalent cells expressed alternative and herita-ble phenotypes. These positional effects were found to bedependent on functional SIR2, -3, and -4 genes. It has beensuggested that silencing results from heterochromatization(5, 41, 81), and this provides a very interesting and testablemodel for the white-opaque or the unmyceliated-heavilymyceliated transition in C. albicans. In this model, a regu-latory gene is positioned next to a heterochromatic regionwhich can exist in two alternative states, a silencing stateand a relaxed state. Transition between the two states resultsin the deactivation-activation of the associated regulatorygene, which in turn leads to repression or expression of a setof phenotype-specific genes (e.g., opaque-specific genes).
EFFECTS OF SWITCHING ON PUTATIVEVIRULENCE TRAITS
C. albicans is an extremely successful pathogen, but it haslong been realized that no single phenotypic trait is solelyresponsible for pathogenesis (76). The phenotypic character-istics which have been considered putative virulence traitsinclude the capacity to form a hypha for dissemination intissue (104), excretion of acid protease (57, 63, 90) andphospholipase (84) for tissue penetration, selective adhesionto both epithelium (23, 69) and plastic (68), and responsive-ness to complement (45, 60) and perhaps female hormones(61, 83). Other characteristics which may enhance the patho-genic potential of C. albicans include the capacity to sup-press the immune system (28), drug resistance (26, 27, 128),and the capacity to live as a commensal in several bodylocations of healthy individuals (76, 114). High-frequencyphenotypic switching has been demonstrated to affect manyof these traits in a reversible fashion.
Hypha Formation
As noted, the differences in colony morphology betweenthe switch phenotypes of C. albicans 3153A and 1001 resultfrom differences in the proportions of buds, hyphae, andpseudohyphae in the colony domes (82, 101), suggesting thatthe environmental constraints or cues for the bud-hyphatransition differ between the different switch phenotypes.Anderson et al. (1) demonstrated that opaque cells differedfrom white cells in strain WO-1 in not readily forminghyphae in suspension cultures under the regimen of pH-regulated dimorphism. Opaque cells did, however, formhyphae en masse when anchored to the wall of a perfusionchamber or to a monolayer of human epithelial cells. In someswitching systems, particular switch phenotypes were char-acterized by perfuse hypha formation. The fuzzy phenotypein strain 3153A (101) and the medusa and fried-egg pheno-types in strain WO-1 (102, 111) were characterized byperfuse mycelium formation, and the major phenotypictransition in 40% of vaginitis strains was between a weaklymyceliated and a heavily myceliated CFU (116).
Acid Protease Secretion
C. albicans secretes an aspartyl proteinase which is be-lieved to be involved in tissue penetration. In two studies, itwas demonstrated that variants which secreted less protein-
ase were less virulent in models (57, 63). There have alsobeen a number of studies in which correlations between thein vitro level of acid protease activity and strain pathogene-sis have been made (20, 36, 85), but as Odds points out in acareful review of the subject (76), there is also one studywhich demonstrates no correlation (75) and a study suggest-ing that, at least in the oral cavity, the pH of saliva wouldsubstantially reduce protease activity (35). This ambiguityseems to surface in virulence studies related to almost everyputative virulence factor for C. albicans pathogenesis, and ittherefore seems reasonable to suggest that virulence of C.albicans is multifaceted; therefore, no single characteristic isexclusively requisite (77). In the white-opaque transition,transcription of the excreted acid proteinase gene is underthe regulation of switching (73a). This is true for not onlytranscription of the gene, but also excretion of the enzyme.Serum proteins in the supporting growth medium have beendemonstrated to stimulate excretion in the white phase, butthe stimulated level is still 10-fold less than than in unin-duced opaque cells, suggesting that maximal expression ofthe excreted acid protease gene may be primarily under theregulation of switching. Dutton and Penn (30) also demon-strated that a switching variant obtained after passagethrough a mouse differed from the parent strain in the levelof excreted acid protease activity. It is therefore possiblethat the variation observed between strains for acid protein-ase excretion is in part a reflection of the switch phenotypeexpressed at the time a strain is tested.
Adhesion to Epithelium and Plastic
Selective adhesion to the epithelium of the oral cavity andvaginal canal have long been considered virulence traits ofC. albicans and related species (23, 69), and adhesion toplastic probably represents one route of invasion in cathe-terized patients (68). Kennedy et al. (51) found that adhesionto epithelium and cohesion differed dramatically betweenwhite and opaque budding cells of strain WO-1. Whitebudding cells were more adhesive than opaque budding cellsto buccal epithelium, but were less cohesive, probably as aresult of increased surface hydrophobicity. On the otherhand, white and opaque cells exhibited the same level ofadhesion to plastic.
Drug SusceptibilityDifferent strains of C. albicans exhibit different suscepti-
bilities to the commonly used antifungal drugs, including thepolyenes (6, 27), 5-fluorocytosine (97, 123), and the azoles(46, 50, 92). There are, no doubt, a number of ways in whichC. albicans achieves drug resistance, including mutation (43,47) and mitotic recombination (130). High-frequency switch-ing can now be added to this list of mechanisms. In a detailedstudy of a single recurrent vaginitis patient, it was demon-strated by DNA fingerprinting that the same strain wasresponsible for three sequential infections and that pheno-typic switching occurred between episodes (113). At the timeof the first vaginal infection, the infecting strain exhibited amedium-sized smooth white colony phenotype. Eighty daysafter clotrimazole treatment, a second infection ensued, andthe infecting strain now exhibited a small-sized, irregular-edge colony phenotype. Seven days after butoconazoletreatment, a third infection ensued, and the infecting strainhad reverted to the medium-sized smooth white phenotypeobserved in the first infection. In this scenario, one wouldhave expected the small-sized, irregular-edge phenotype in
198 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 199
the second infection to be clotrimazole resistant, since clot-rimazole was used to suppress the first infection, but thereverse was true (113). In the third infection, the strainreverted to the phenotype and level of clotrimazole suscepti-bility observed in the first infection. Therefore, reversiblechanges in drug susceptibility can accompany switching at thesite of infection, but these changes may not result in resis-tance to the drug used for treatment, suggesting that enrich-ment of a switch phenotype after drug therapy may be due toa mechanism for escaping drug action other than drug resis-tance. For instance, application of an imidazole to the vaginalcanal may eradicate cells exhibiting the major switch pheno-type at the time of treatment, but this treatment may missanother switch phenotype of the same strain with a selectiveadvantage in colonizing the skin of the groin and inner thigh.After treatment, this switch phenotype may then recolonizethe vaginal canal. The alternating switch phenotypes in thestudy just described (113) exhibited different profiles of drugsusceptibility which included a number of drugs not used intherapy, suggesting that each phenotype in the switchingrepertoire of the infecting strain exhibits a specific drugsusceptibility profile. This conclusion is supported by studiesof drug susceptibility for switch phenotypes in the 3153Asystem (100) and the white-opaque transition (111).
Antigenic Change
The switching systems of Trypanosoma spp., Neissenaspp., Salmonella spp., and Borrelia spp. impact directly oncell surface antigenicity (9, 13, 17, 29, 38, 98, 125), and it hasbeen assumed that these surface changes occur, at least inpart, to escape the immune system. There is ample evidenceof antigenic change during the course of infection for some ofthese pathogens (9, 89). Antigenic differences have beendemonstrated between different strains of C. albicans, sepa-rating them into A and B serotypes (44), and there has beenmounting evidence that C. albicans might have the capacityto cycle through antigenic states during the course of aninfection. Antigenic variability has been demonstrated in thewhite-opaque transition and involves expression of anopaque-specific surface antigen (1, 2, 4). This antigenic switchdoes not appear to involve replacement of the opaque-specificantigen by an alternative white-specific antigen, as occurs inthe replacement systems in other pathogens (9, 29, 38, 108).The white-opaque transition is more analogous to the ON-OFF mechanism of type 1 fimbriae formation in E. coli (38).In addition, Fruit et al. (34) demonstrated that portions of aclonal colony of C. albicans could be differentially stainedwith a monoclonal antibody to a surface glycoprotein, sug-gesting differential expression resulting from switching.
Sensitivity to Neutrophils and In Vitro Oxidants
Since polymorphonuclear leukocytes appear to be respon-sible for containing C. albicans dissemination. Kolotila andDiamond (55) tested whether the white and opaque phases ofstrain WO-1 exhibited different sensitivities to the candi-dacidal activity of polymorphonuclear leukocytes. Althoughthere were equivalent levels of phagocytosis of white andopaque cells, white cells were more resistant to the candi-dacidal activity of intact polymorphonuclear leukocytes.Similar differences were observed in sensitivity to cell-freeoxidants. As previously noted, both intact polymorphonu-clear leukocytes and oxidants stimulated switching in thewhite-opaque direction (55).
SWITCHING IN INFECTING STRAINS
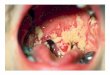
From the preceding review, there seems little doubt thatswitching affects putative virulence traits. However, ifswitching plays a role in pathogenesis, it must occur in vivo,and there are a number of observations which suggest thatthis is indeed the case. To demonstrate switching at the siteof C. albicans carriage or infection, cells must be clonallyplated onto zinc-limiting agar directly from the sampled site(110, 116). In such a study of 11 patients with vaginalcandidiasis (116), multiple colony morphologies were ob-served in four of the primary isolates. Fingerprinting withthe moderately repetitive probe Ca3 (referred to as JH3 inthe original study) demonstrated that the multiple pheno-types in each of these isolates represented the same strain ineach case, and subsequent plating experiments demon-strated that multiple phenotypes in primary isolates wereinterconvertible at high frequency (116). In two isolates, P3and P5, four different colony morphologies were identified inthe original plating, and in two other isolates, P6 and P8, twodifferent colony morphologies were identified. Nine of the 11original isolates were in high-frequency modes of switching,generating variant colonies in plating experiments at fre-quencies between 10-2 and 10-3. Switching also occurs incommensal strains, but it is somewhat harder to documentbecause of the usually lower intensity of commensal car-riage. However, in the vulvovaginal region, commensalcarriage can be quite intense, and in a minority of cases,multiple phenotypes have been observed in primary clonalplatings (112a). In Fig. 10A, an example is presented of aprimary clonal culture from the vaginal wall of a healthyasymptomatic woman. In this case, carriage was very highwith no sign of morbidity. The commensal population wascomposed of 96% smooth white colonies and 4% irregularwrinkled colonies with a heavily myceliated halo (Fig. 10).Cells from either phenotype switched to the alternativephenotype at relatively high frequency.As previously noted in this review, in an analysis of a
single recurrence in a patient, the colony morphology of theinfecting strain changed in each of the two successiveepisodes (113). However, an analysis of only one patientwith recurrence precludes any conclusion concerning theextent of switching in recurrent infections.
SWITCHING AND ADAPTATION
In contrast to the antigenic switching systems functioningin B. hermsii (9), T. brucei (29), N. gonorrhoeae (125), and S.typhimurium (38), switching in C. albicans results in com-plex changes in phenotype which can potentially affect morethan one aspect of cell physiology, morphology, or viru-lence. Therefore, the role of switching in C. albicans maynot be restricted to a single aspect of pathogenesis. Ittherefore seems more fruitful to consider what advantagemay be gained by C. albicans in possessing variabilitysystems such as the ones so far described. Help in answeringthis question may be provided by considering the process ofadaptation in evolution. Variability of offspring is basic tosuccessful propagation in a changing environment (133) andprovides a population with a vehicle for expanding into newenvironments. Variation usually is provided by mutationsleading to allelic differences which are shuffled into newcombinations by sexual cycles and meiotic recombination.This general scenario, with different details, may be the keyto understanding the role played by switching in C. albicansand related species. In each population of C. albicans,
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

CLIN. MICROBIOL. REV.
FIG. 10. Switching at the site of C. albicans carriage. (A) Pri-mary plating from the vaginal wall of an asymptomatic patient; (B)increased magnification of one heavily myceliated and two smoothcolonies. Cells replated from the two phenotypes switched to thealternative phenotypes at a high frequency.
roughly 1 in 1,000 cells may exist in an altered phenotypicform, providing it with a potential advantage in a differentecological niche represented by different tissues and bodylocations or a changing environmental niche involving a
change in host physiology or drug therapy. A recent studysuggests that adaptation may indeed occur within the con-
fines of a healthy individual (114). In this study, bothCandida carriage and strain relatedness were assessed in 52healthy women at 17 anatomical locations, using isolationprocedures which assess carriage intensity (110, 116) and a
computer-assisted DNA fingerprinting system which com-
putes genetic similarity between isolates on the basis of thepattern of Southern blots probed with the moderately repet-itive sequence Ca3 (96, 114). Seventy-three percent of testindividuals carried Candida spp. in at least one anatomicallocation, and 36% carried Candida spp. in more than one of
three major anatomical locations: the oral cavity, the anorec-tal region, and the vulvovaginal region. In 7 of 11 testindividuals in which Candida spp. were simultaneouslyisolated from the oral cavity and the vulvovaginal region, thepairs of isolates were genetically unrelated, and in the remain-ing four cases, the pairs were genetically similar but noniden-tical. In marked contrast, nine pairs of isolates from the vulvaand vaginal canal were identical, which was not surprisinggiven the contiguity of the two regions. These results suggestthat, in 7 of 11 cases, the disparate environments of the oralcavity and vaginal canal selected for unrelated vaginotropicand oral-tropic strains of Candida spp., respectively. How-ever, in 4 of the 11 cases, the isolate pairs were geneticallysimilar but not identical, suggesting that they had evolvedfrom a single progenitor. Scherer and Stephens (95) obtainedsimilar results demonstrating that isolates from different bodylocations were similar but nonidentical. If a single strain candiverge genetically in two body locations, one may concludethat the anatomically separated populations are not mixingand have therefore adapted to alternative environmentalniches. It seems reasonable to suggest that the phenotypicdiversity necessary for a single strain initially to inhabitdifferent anatomical niches, in this case the vaginal canal andoral cavity, may be provided by high-frequency phenotypicswitching. This suggestion can be tested.
CONCLUSION
High-frequency switching provides C. albicans with asecond level of phenotypic variability which is superimposedon the bud-hypha transition. Switching has been observed ina majority of commensal and pathogenic strains, as well as inthe related pathogenic species C. tropicalis. It thereforeappears to represent a general mechanism for populationvariability in this class of fungal pathogens. Switching inCandida spp. sometimes has dramatic effects on phenotype,as in the case of the white-opaque transition in C. albicansWO-1, and involves differential gene transcription. It canaffect many aspects of cellular physiology and morphology,including many putative virulence traits. Because C. albicanshas no identified sexual cycle and appears to be an obligatediploid, it lacks the avenue of variability provided by meiosis.Because of its broad success as both a commensal and apathogen, one may safely presume that C. albicans hasclaimed a number of other mechanisms for generating popu-lation variability fundamental to adaptation and that switchingrepresents one of them. Because C. albicans lacks a sexualcycle which would facilitate effective genetic studies, andbecause switching is a relatively recent discovery, or redis-covery, there have been few aggressive investigations into themolecular basis of the switching mechanism. It is hoped thatthis review will stimulate interest in this extraordinary pro-cess of phenotypic variability and lead to the elucidation ofthe molecular mechanism and the phenotypic consequencesof switching in this important fungal pathogen.
ACKNOWLEDGMENTS
I am indebted to B. Morrow, J. Anderson, T. Shrikantha, J.Schmid, and R. Galask for permission to report results which havenot yet been published, to J. Hicks, P. Magee, and S. Scherer forvaluable discussions related to switching, and to D. Wessels forhelping in the development of the manuscript.The work in my laboratory was supported by Public Health
Service grant A123922 from the National Institutes of Health.
200 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 201
REFERENCES
1. Anderson, J. M., L. Cundiff, B. Schnars, M. Gao, I. Mackenzie,and D. R. Soil. 1989. Hypha formation in the white-opaquetransition of Candida albicans. Infect. Immun. 57:458-467.
2. Anderson, J. M., R. Mihalik, and D. R. Soil. 1990. Ultrastruc-ture and antigenicity of the unique cell and pimple of theCandida opaque phenotype. J. Bacteriol. 172:224-235.
3. Anderson, J. M., and D. R. Soll. 1986. Differences in actinlocalization during bud and hypha formation in the yeastCandida albicans. J. Gen. Microbiol. 132:2035-2047.
4. Anderson, J. M., and D. R. Soll. 1987. Unique phenotype ofopaque cells in the white-opaque transition of Candida albi-cans. J. Bacteriol. 169:5579-5588.
5. Aparicho, 0. M., B. C. Billington, and D. E. Gottschling. 1991.Modifiers of position effects are shared between telomeric andsilent mating-type loci in S. cerevisiae. Cell 66:1279-1287.
6. Athar, M. A., and H. I. Winner. 1991. Development ofresistance by Candida species to polyene antibiotics in vitro. J.Med. Microbiol. 4:505-517.
7. Auger, P., and J. Joly. 1977. Factors influencing germ tubeproduction in Candida albicans. Mycopathologia 61:183-186.
8. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G.Seidman, J. A. Smith, and K. Struhi. 1989. Current protocols inmolecular biology. John Wiley and Sons, Inc., New York.
9. Barbour, A. 1989. Antigenic variation in relapsing fever Bor-relia species: genetic aspects, p. 783-790. In D. E. Berg andM. M. Howe (ed.), Mobile DNA. American Society for Micro-biology, Washington, D.C.
10. Barlow, A. J. E., and T. Aldersley. 1974. Factors present inserum and seminal plasma which format germ-tube formationand mycelial growth of Candida albicans. J. Gen. Microbiol.82:261-272.
11. Bedell, G. W., and D. R. Soil. 1979. Effects of low concentra-tions of zinc on the growth and dimorphism of Candidaalbicans: evidence for zinc-resistant and -sensitive pathwaysfor mycelium formation. Infect. Immun. 26:348-354.
12. Bedell, G. W., A. Werth, and D. R. Soll. 1980. The regulationof nuclear migration and division during synchronous budformation in released stationary phase cultures of the yeastCandida albicans. Exp. Cell. Res. 127:103-113.
13. Berg, D. E., and M. M. Howe (ed.). 1989. Mobile DNA.American Society for Microbiology, Washington, D.C.
14. Bergen, M., E. Voss, and D. R. Soll. 1990. Switching at thecellular level in the white-opaque transition of Candida albi-cans. J. Gen. Microbiol. 136:1925-1936.
15. Bocke, J. D. 1989. Transposable elements in Saccharomycescerevisiae. p. 335-374. In D. E. Berg and M. M. Howe (ed.),Mobile DNA. American Society for Microbiology, Washing-ton, D.C.
16. Borgers, M., and S. DeNollin. 1974. The preservation ofsubcellular organelles of Candida albicans with conventionalfixation. J. Cell. Biol. 62:574-581.
17. Borst, P., and D. R. Greaves. 1987. Programmed rearrange-ments altering gene expression. Science 235:658-667.
18. Brown, L. A., and W. L. Chaffin. 1981. Differential expressionof cytoplasmic proteins during yeast bud and germ tubeformation in Candida albicans. Can. J. Microbiol. 27:580-585.
19. Brown-Thomsen, J. 1968. Variability in Candida albicans(Robin) Berkhous. Hereditas 60:335-398.
20. Budtz-Jorgensen, E. 1971. Proteolytic activity of Candida spp.as related to the pathogenesis of denature stomatitis. Sab-ouraudia 12:266-271.
21. Buffo, J., M. A. Herman, and D. R. Soil. 1984. A characteri-zation of pH-regulated dimorphism in Candida albicans. My-copathologia 85:21-30.
22. Byers, B. 1981. Cytology of the yeast life cycle, p. 59-96. InJ. N. Strathern, E. W. Jones, and J. R. Broach (ed.), Themolecular biology of the yeast Saccharomyces: life cycle andinheritance. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
23. Calderone, R. A., R. L. Cihlar, D. D. Lee, K. Hoberg, andW. M. Scheld. 1985. Yeast adhesion in the pathogenesis of
endocarditis due to Candida albicans: studies with adherence-negative mutants. J. Infect. Dis. 152:710-715.
24. Chandrasekhar, A., M. Rotman, B. Kraft, and D. R. Soll. 1990.Developmental mechanisms regulating the rapid decrease in acohesion glycoprotein mRNA in Dictyostelium function pri-marily at the level of mRNA degradation. Dev. Biol. 141:262-269.
25. Davies, D. R. 1990. The structure and function of the asparticproteinases. Annu. Rev. Biophys. Biophys. Chem. 19:189-215.
26. Defever, K. S., W. L. Whelan, A. L. Rogers, E. S. Beneke,J. M. Veselenak, and D. R. Soll. 1982. Candida albicansresistance to 5-fluorocytosine: frequency of partially resistantstrains among clinical isolates. Antimicrob. Agents Chemo-ther. 22:810-815.
27. Dick, J. D., W. G. Merz, and R. Saral. 1980. Incidence ofpolyene-resistant yeasts recovered from clinical specimens.Antimicrob. Agents Chemother. 18:158-163.
28. Domer, J., K. Elkins, D. Enaist, and P. Baker. 1988. Modula-tion of immune responses by surface polysaccharides of Can-dida albicans. Rev. Infect. Dis. 10:419-422.
29. Donelson, J. E. 1989. DNA rearrangements and antigenicvariation in African trypanosomes, p. 763-782. In D. E. Bergand M. M. Howe (ed.), Mobile DNA. American Society forMicrobiology, Washington, D.C.
30. Dutton, S., and C. W. Penn. 1989. Biological attributes ofcolony-type variants of Candida albicans. J. Gen. Microbiol.135:3363-3372.
31. Evans, E. G., F. C. Odds, M. D. Richardson, and R. T.Holland. 1974. Effect of growth medium on filament productionin Candida albicans. Sabouraudia 12:112-119.
32. Finney, R., C. J. Langtimm, and D. R. Soil. 1985. Theprograms of protein synthesis accompanying the establishmentof alternative phenotypes in Candida albicans. Mycopatholo-gia 91:3-15.
33. Freifelder, D. 1960. Bud position in Saccharomyces cerevisiae.J. Bacteriol. 80:567-575.
34. Fruit, J., J. C. Cailliez, F. C. Odds, and D. Poulain. 1990.Expression of an epitope by surface glycoproteins of Candidaalbicans. Variability among species, strains and yeast cells ofthe genus Candida. J. Med. Vet. Mycol. 28:241-252.
35. Germaine, G. R., and L. M. Tellepson. 1981. Effect of pH andhuman saliva on protease production by Candida albicans.Infect. Immun. 31:323-326.
36. Ghannoum, M. A., and K. A. Elteen. 1986. Correlative rela-tionship between proteinase production, adherence and patho-genicity of various strains of Candida albicans. J. Med. Vet.Mycol. 24:407-413.
37. Gil, C., R. Pomes, and C. Nombela. 1988. A complementationanalysis by parasexual recombination of Candida albicansmorphological mutants. J. Gen. Microbiol. 134:1587-1595.
38. Glasgow, A. C., K. T. Hughes, and M. I. Simon. 1989. BacterialDNA inversion systems, p. 637-660. In D. E. Berg and M. M.Howe (ed.), Mobile DNA. American Society for Microbiology,Washington, D.C.
39. Goshorn, A. K., and S. Scherer. 1989. Genetic analysis ofprototropic natural variants of Candida albicans. Genetics123:667-673.
40. Gottlieb, S., and R. E. Esposito. 1989. A new role for a yeasttranscriptional silencer gene, SIR2, in regulation of recombi-nation in ribosomal DNA. Cell 56:771-776.
41. Gottschling, D. E., 0. M. Aparicio, B. L. Billington, and V. A.Zakian. 1990. Position effect at S. cerevisiae telomeres: revers-ible repression of Pol II transcription. Cell 63:751-762.
42. Gow, N. A. R., G. Henderson, and G. V. Gooday. 1986.Cytological interrelationships between the cell cycle and du-plication cycle of Candida albicans. Microbios 47:97-105.
43. Hamilton-Miller, J. M. T. 1972. Physiological properties ofmutagen-induced variants of Candida albicans resistant topolyene antibiotics. J. Med. Microbiol. 5:425-440.
44. Hasenclever, H. F., and W. 0. Mitchell. 1961. Antigenic studiesof Candida. I. Observation of two antigenic groups in Candidaalbicans. J. Bacteriol. 82:570-573.
45. Heidenreich, F., and M. P. Dierich. 1985. Candida albicans and
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

CLIN. MICROBIOL. REV.
Candida stellatoidea, in contrast to other Candida species,bind C3b and C3d but not C3b. Infect. Immun. 50:598-600.
46. Hitchcock, C. A., K. J. Barrett-Bee, and N.J. Russel. 1986. Thelipid composition of azole-sensitive and azole-resistant strainsof Candida albicans. J. Gen. Microbiol. 132:2421-2431.
47. Hsucher, C. C., and D. S. Feingold. 1974. Two types ofresistance to polyene antibiotics in Candida albicans. Nature(London) 251:656-659.
48. Hube, B., C. J. Turner, F. C. Odds, H. Eiffert, G. J. Boulnois,H. Kochel, and R. Ruschel. 1991. Sequence of the Candidaalbicans gene encoding the secretory aspartate proteinase. J.Med. Vet. Mycol. 29:129-132.
49. Ito-Kuwa, S. 1986. Ultrastructural changes in the cell wallduring germ tube and bud formation in the dimorphic fungusCandida albicans. Odontology 73:1586-1602.
50. Johnson, E. M., M. D. Richardson, and D. W. Warnock. 1984.In vitro resistance to imidazole antifungals in Candida albi-cans. J. Antimicrob. Chemother. 13:547-558.
51. Kennedy, M. J., A. L. Rogers, L. R. Hanselman, D. R. Soll, andR. J. Yancey. 1988. Variation in adhesion and cell surfacehydrophobicity in Candida albicans white and opaque pheno-types. Mycopathologia 102:149-156.
52. Klar, A. J. S. 1989. The interconversion of yeast mating type:Saccharomyces cerevisiae and Schizosaccharomyces pombe,p. 671-691. In D. E. Berg and M. M. Howe (ed.), Mobile DNA.American Society for Microbiology, Washington, D. C.
53. Klar, A. J. S., J. N. Strathern, and J. A. Abraham. 1984.Involvement of double-stranded chromosomal breaks for mat-ing type switching in Saccharomyces cerevisiae. Cold SpringHarbor Symp. Quant. Biol. 49:77-82.
54. Klionsky, D. J., P. K. Herman, and S. D. Ewer. 1990. Thefungal vacuole: composition, function, and biogenesis. Micro-biol. Rev. 54:266-292.
55. Kolotila, M. P., and R. D. Diamond. 1990. Effects of neutro-phils and in vitro oxidants on survival and phenotypic switch-ing of Candida albicans WO-1. Infect. Immun. 58:1174-1179.
56. Kurtz, M. B., R. Kelly, and D. R. Kirsch. 1990. Moleculargenetics of Candida albicans, p. 21-73. In D. R. Kirsch, R.Kelly, and M. B. Kurtz (ed.), The genetics of Candida. CRCPress, Boca Raton, Fla.
57. Kwon-Chung, K. J., D. Lehman, C. Good, and P. T. Magee.1985. Genetic evidence for the role of extracellular proteinase invirulence of Candida albicans. Infect. Immun. 49:571-575.
58. Land, G. A., W. C. MacDonald, R. L. Stijernholm, and L.Friedman. 1975. Factors affecting filamentation in Candidaalbicans: changes in respiratory activity of Candida albicansduring filamentation. Infect. Immun. 12:119-127.
59. Lee, K. L., H. R. Buckley, and C. C. Campbell. 1975. An aminoacid liquid synthetic medium for development of mycelial andyeast forms of Candida albicans. Sabouraudia 13:148-153.
60. Linchan, L., E. Wadsworth, and R. Calderone. 1988. Candidaalbicans C3d receptor, isolated by using a monoclonal anti-body. Infect. Immun. 56:1981-1986.
61. Loose, D. S., D. Schurman, and D. Feldman. 1981. A cortico-steroid binding protein and endogenous ligand in Candidaalbicans indicating a possible steroid-receptor system. Nature(London) 293:477-479.
62. Lyon, M. 1961. Gene action in the X-chromosome of the mouse(Mus musculus L.). Nature (London) 190:372-373.
63. MacDonald, F., and F. C. Odds. 1980. Inducible proteinaseof Candida albicans in diagnostic serology and in the pathogen-esis of systemic,candidiasis. J. Med. Microbiol. 13:423-436.
64. Mackinnon, J. E. 1940. Dissociation in Candida albicans. J.Infect. Dis. 66/67:59-77.
65. Magee, B. B., and P. T. Magee. 1987. Electrophoretic karyo-types and chromosome numbers in Candida species. J. Gen.Microbiol. 133:425-430.
66. Manning, M., and T. G. Mitchell. 1980. Analysis of cytoplas-mic antigens of the yeast and mycelial phases of Candidaalbicans by two-dimensional electrophoresis. Infect. Immun.30:484-495.
67. Mardon, D. W., E. Balish, and A. W. Phillips. 1969. Control ofdimorphism in a biochemical variant of Candida albicans. J.
Bacteriol. 100:701-707.68. McCourtie, J., and L. J. Douglas. 1981. Relationship between
cell surface composition of Candida albicans and adherence toacrylic after growth on different carbon source. Infect. Immun.32:1234-1241.
69. McCourtie, J., and L. J. Douglas. 1984. Relationship betweencell surface composition, adherence, and virulence of Candidaalbicans. Infect. Immun. 45:6-12.
70. Merson-Davies, L. A., and F. C. Odds. 1989. A morphologyindex for characterization of cell shape in Candida albicans. J.Gen. Microbiol. 135:3143-3152.
71. Merz, W. G. 1990. Candida albicans strain delineation. Clin.Microbiol. Rev. 3:321-334.
72. Merz, W. G., C. Connelly, and P. Hieter. 1988. Variation ofelectrophoretic karyotypes among clinical isolates of Candidaalbicans. J. Clin. Microbiol. 26:842-845.
73. Morrow, B., J. Anderson, E. Wilson, and D. R. Soil. 1989.Bidirectional stimulation of the white-opaque transition ofCandida albicans by ultraviolet irradiation. J. Gen. Microbiol.135:1201-1208.
73a.Morrow, B., T. Srikantha, and D. R. Soli. Submitted forpublication.
74. Negroni, P. 1935. Variacion bacia et tipo R de Mycotorulaalbicans. Rev. Soc. Argent. Biol. 11:449-453.
75. Niczyporuk, W., E. Malyszko, H. Szarmach, and J. Roszkiew-iez. 1986. In vitro-untersuchungen zur proteolytischen undlipolytischen Aktivitat hefeanalicher Pilze Sourie deren Viru-lenz geneuber Mausen. Mykosen 29:89-95.
76. Odds, F. C. 1985. Candida albicans proteinase as a virulencefactor in the pathogenesis of Candida infections. Zentralbl.Bakteriol. A 260:539-542.
77. Odds, F. C. 1988. Candida and candidosis: a review andbibliography. Bailliere Tindale, London.
78. Odds, F. C., and L. A. Merson-Davies. 1989. Colony variationsin Candida species. Mycoses 32:275-282.
79. Olaiya, A. F., and S. J. Sogin. 1979. Ploidy determination ofCandida albicans. J. Bacteriol. 140:1043-1049.
80. Olaiya, A. F., J. R. Steed, and S. J. Sogin. 1980. Deoxyribo-nucleic acid-deficient strains of Candida albicans. J. Bacteriol.141:1284-1290.
81. Pitlus, L., and J. Rine. 1989. Epigenetic inheritance of tran-scriptional states of S. cerevisiae. Cell 59:637-647.
82. Pomes, R., C. Gil, and C. Nombela. 1985. Genetic analysis ofCandida albicans morphological mutants. J. Gen. Microbiol.131:2107-2113.
83. Powell, B. L., C. L. Frey, and D. J. Drutz. 1984. Identificationof a 17-estradiol binding protein in Candida albicans andCandida (Torulopsis) glabrata. Exp. Mycol. 8:304-313.
84. Pugh, D., and R. A. Cawson. 1977. The cytochemical localiza-tion of phospholipase in Candida albicans infecting the chickchorio-allantoic membrane. Sabouraudia 15:29-35.
85. Reymold, H., H. Fasold, and F. Staib. 1968. Purification andcharacterization of a proteolytic enzyme from Candida albi-cans. Biochim. Biophys. Acta 167:399-406.
86. Riggsby, W. S., L. J. Torres-Bauza, J. W. Willis, and T. M.Townes. 1982. DNA content, kinetic complexity, and the ploidyquestion in Candida albicans. Mol. Cell. Biol. 2:853-862.
87. Rikkerink, E. H. A., B. B. Magee, and P. T. Magee. 1988.Opaque-white phenotype transition: a programmed morpho-logical transition in Candida albicans. J. Bacteriol. 170:895-899.
88. Rine, J., J. N. Strathern, J. Hicks, and I. Herskowitz. 1979. Asuppressor of mating-type locus mutations in Saccharomycescerevisiae: evidence for and identification of cryptic mating-type loci. Genetics 93:877-901.
89. Ross, R., and D. Thomsen. 1910. A case of sleeping sicknessstudied by precise enumerative methods. Regular periodicincrease in the parasites disclosed. Proc. R. Soc. London Ser.B. 82:411-415.
90. Ruchel, R. 1981. Properties of purified proteinase from theyeast Candida albicans. Biochim. Biophys. Acta 659:99-113.
91. Rustchenko-Bulgac, E. P., F. Sherman, and J. B. Hicks. 1990.Chromosomal rearrangements associated with morphological
202 SOLL
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

HIGH-FREQUENCY SWITCHING IN C. ALBICANS 203
mutants provide a means for genetic variation on Candidaalbicans. J. Bacteriol. 172:1276-1283.
92. Ryley, J. F., R. G. Wilson, and K. J. Barrett-Bee. 1984. Azoleresistance in Candida albicans. Sabouraudia 22:53-63.
93. Sadhu, C., M. J. McEachern, E. P. Rustchenko-Bulgac, J.Schmid, D. R. Soil, and J. B. Hicks. 1991. Telomeric anddispersed repeat sequences in Candida yeasts and their use instrain identification. J. Bacteriol. 173:842-850.
94. Scherer, S., and P. T. Magee. 1990. Genetics of Candidaalbicans. Microbiol. Rev. 54:226-241.
95. Scherer, S., and D. A. Stevens. 1988. A Candida albicansdispersed, repeated gene family and its epidemiologic applica-tions. Proc. Natl. Acad. Sci. USA 85:1452-1456.
96. Schmid, J., E. Voss, and D. R. Soil. 1990. Computer-assistedmethods for assessing Candida albicans strain relatedness bySouthern blot hybridization with repetitive sequence Ca3. J.Clin. Microbiol. 28:1236-1243.
97. Scholer, H. J. 1980. Fluorocytosine, p. 35-106. In D. C. E.Speller (ed.), Antifungal chemotherapy. John Wiley and Sons,Chichester, England.
98. Shapiro, J. 1983. Mobile genetic elements. Academic Press,Inc., Orlando, Fla.
99. Simonetti, N., V. Strippoli, and A. Cassone. 1974. Yeast-mycelial conversion induced by N-acetyl-D-glucosamine inCandida albicans. Nature (London) 250:344-346.
100. Slutsky, B. 1986. A characterization of two high frequencyswitching systems in the dimorphic yeast Candida albicans.Ph.D. thesis. University of Iowa, Iowa City.
101. Slutsky, B., J. Buffo, and D. R. Soll. 1985. High frequencyswitching of colony morphology in Candida albicans. Science230:666-669.
102. Slutsky, B., M. Staebell, J. Anderson, L. Risen, M. Pfaller, andD. R. Soil. 1987. "White-opaque transition": a second high-frequency switching system in Candida albicans. J. Bacteriol.169:189-197.
103. Snell, R. G., I. F. Hermans, R. J. Witkins, and B. E. Corner.1987. Chromosomal variation in Candida albicans. NucleicAcids Res. 15:3625.
104. Sobel, J. D., G. Muller, and H. R. Buckley. 1984. Critical roleof germ tube formation in the paihogenesis of candidal vagini-tis. Infect. Immun. 44:576-580.
105. Soil, D. R. 1984. The cell cycle and commitment to alternativecell fates in Candida albicans, p. 143-162. In P. Nurse and E.Streiblova (ed.), The microbial cell cycle. CRC Press, BocaRaton, Fla.
106. Soll, D. R. 1985. The role of zinc in Candida dimorphism. Curr.Top. Med. Mycol. 1:258-285.
107. Soll, D. R. 1986. The regulation of cellular differentiation in thedimorphic yeast Candida albicans. Bioessays 5:5-11.
108. Soil, D. R. 1989. High-frequency switching in Candida albi-cans, p. 791-798. In D. E. Berg and M. M. Howe (ed.), MobileDNA. American Society for Microbiology, Washington, D.C.
109. Soll, D. R. 1990. Dimorphism and high frequency switching inCandida albicans, p. 147-176. In D. R. Kirsch, R. Kelly, andM. B. Kurtz (ed.), The genetics of Candida. CRC Press, BocaRaton, Fla.
110. Soll, D. R. 1991. Current status of the molecular basis ofCandida pathogenicity, p. 503-540. In G. Cole and H. Hoch(ed.), The fungal spore and disease initiation in plants andanimals. Plenum Press Inc., New York.
111. Soil, D. R., J. Anderson, and M. Bergen. 1991. The develop-mental biology of the white-opaque transition in Candidaalbicans, p. 20-45. In R. Prasad (ed.), The molecular biology ofCandida albicans. Springer-Verlag, Berlin.
112. Soil, D. R., G. Bedell, J. Thiel, and M. Brummel. 1981. Thedependency of nuclear division on volume in the dimorphicyeast Candida albicans. Exp. Cell Res. 133:55-62.
112a.Soll, D. R., and R. Galask. Unpublished data.113. Soil, D. R., R. Galask, S. Isley, T. V. G. Rao, D. Stone, J.
Hicks, J. Schmid, K. Mac, and C. Hanna. 1989. "Switching" ofCandida albicans during successive episodes of recurrentvaginitis. J. Clin. Microbiol. 27:681-690.
114. Soil, D. R., R. Galask, J. Schmid, C. Hanna, K. Mac, and B.Morrow. 1991. Genetic dissimilarity of commensal strains ofCandida spp. carried in different anatomical locations of thesame healthy women. J. Clin. Microbiol. 29:1702-1710.
115. Soll, D. R., M. A. Herman, and M. A. Staebell. 1985. Theinvolvement of cell wall expansion in the two modes ofmycelium formation of Candida albicans. J. Gen. Microbiol.131:2367-2375.
116. Soil, D. R., C. J. Langtimm, J. McDowell, J. Hicks, and R.Galask. 1987. High-frequency switching in Candida strainsisolated from vaginitis patients. J. Clin. Microbiol. 25:1611-1622.
117. Soll, D. R., and L. H. Mitchell. 1983. Filament ring formation inthe dimorphic yeast Candida albicans. J. Cell Biol. 96:486-493.
118. Soil, D. R., B. Slutky, S. Matkenzie, C. Langtimm, and M.Staebell. 1987. Switching systems in Candida albicans andtheir possible roles in oral candidiasis, p. 52-59. In J. Mac-kenzie, C. Squier, and E. Dabelsteen (ed.), Oral mucosadiseases: biology, etiology and therapy. Laegeforeningens Fo-larg, Denmark.
119. Soil, D. R., M. Staebell, C. Langtimm, M. Pfaller, J. Hicks, andT. V. G. Rao. 1988. Multiple Candida strains in the course ofa single systemic infection. J. Clin. Microbiol. 26:1448-1459.
120. Sonneborn, T. M. 1963. Does preformed cell structure play anessential role in cell heredity?, p. 165-221. In J. M. Allen (ed.),The nature of biological diversity. McGraw-Hill Book Co.,New York.
120a.Srikantha, T., and D. R. Soll. Unpublished data.121. Staebell, M., and D. R. Soil. 1985. Temporal spatial differences
in cell wall expansion during bud and mycelium formation inCandida albicans. J. Gen. Microbiol. 131:1467-1480.
122. Stewart, M., N. A. R. Gow, and D. V. Bowen. 1988. Cytoplas-mic alkalinization during germ tube formation in Candidaalbicans. J. Gen. Microbiol. 134:1079-1087.
123. Stiller, R. L., J. E. Bennett, H. J. Scholer, M. Wall, A. Polak,and D. A. Stevens. 1982. Susceptibility to 5-fluorocytosine andprevalence of serotype in 402 Candida albicans isolates, fromthe United States. Antimicrob. Agents Chemother. 22:482-487.
124. Suzuki, T., I. Kobayashi, T. Kanbe, and K. Tanaka. 1989. Highfrequency variation of colony morphology and chromosomereorganization in the pathogenic yeast Candida albicans. J.Gen. Microbiol. 135:425-434.
125. Swanson, J., and J. M. Koomey. 1989. Mechanisms for varia-tion of pili and outer membrane protein II in Neisseria gonor-rhoea, p. 743-762. In D. E. Berg and M. M. Howe (ed.),Mobile DNA. American Society for Microbiology, Washing-ton, D.C.
126. Taschdjian, L. L., J. J. Burchall, and P. J. Kozinn. 1960. Rapididentification of Candida albicans by filamentation on serumand screen substitutes. Am. J. Dis. Child. 99:212-215.
127. Vogel, R. A., and R. S. Sponeler. 1970. The study and signifi-cance of colony dissociation in Candida albicans. Sabouraudia7:273-278.
128. Warnock, D. W., E. M. Johnson, M. D. Richardson, andC. F. H. Vickers. 1983. Modified response to ketoconazole ofCandida albicans from a treatment failure. Lancet i:642-643.
129. Whelan, W. L. 1987. The genetics of medically importantfungi. Crit. Rev. Microbiol. 21:99-170.
130. Whelan, W. L., E. S. Beneke, A. L. Rogers, and D. R. Soll.1981. Segregation of 5-fluorocytosine-resistant variants byCandida albicans. Antimicrob. Agents Chemother. 19:1078-1081.
131. Whelan, W. L., and P. T. Magee. 1981. Natural heterozygosityin Candida albicans. J. Bacteriol. 145:896-903.
132. Whelan, W. L., and D. R. Soll. 1982. Mitotic recombination inCandida albicans: recessive lethal alleles linked to a generequired for methionine biosynthesis. Mol. Gen. Genet. 187:477-485.
133. Williams, G. C. 1975. Sex and evolution. Princeton UniversityPress, Princeton, N.J.
VOL. 5, 1992
on July 2, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from