Embed Size (px)
Citation preview

HSP90 manages the endsDiane C. DeZwaan and Brian C. Freeman
Department of Cell and Developmental Biology, University of Illinois, Urbana-Champaign, 601 S. Goodwin Avenue, Urbana,
IL 61801, USA
Review
The telomere environment requires an efficient means toassemble and disassemble a multitude of structures tooperate correctly and to help achieve cellular homeosta-sis. Telomeres are challenged by a common bindingspecificity displayed by many of the protein componentsfor telomeric DNA, which could result in competitiveDNA interactions, and by a cell cycle-restricted timingof events, which enforces a narrow working period inwhich to perform numerous tasks. In this review, wediscuss how the HSP90 molecular chaperone networkavoids these obstacles and facilitates an effectiveoperation of the telomere system.
A dynamic cellular environment enables multi-steppathwaysTo achieve and maintain physiological balance a cellrequires numerous biological pathways to operate har-moniously, including paths functioning at or near theDNA. The variety of processes working along the genomeis considerable, yet the number of factors employedeclipses this total. Typically, individual componentsassemble into distinct complexes that operate seriallyalong a pathway to perform a cellular task. Adding tothe complexity is a multitude of fluctuating internal andexternal signals that must be monitored to initiate, con-tinue or halt a particular action. Although cooperativeinteractions between proteins of the same process providea mechanism for rapid and precise assembly, the inherentstability of such organized structures could interfere withthe proper timing of biological systems [1]. Hence, cellularfactors are needed to destabilize the various proteinassemblies to allow efficient transition between struc-tures or to terminate pathway activity [2]. We suggestthat molecular chaperones have evolved, in part, to fostera general and continuous mobile protein environmentwithin the cell, including events that occur along thegenome [3].
To illustrate the potential complexities and obstaclesfaced by most, if not all, biological pathways, we focus onthe impact of one molecular chaperone network on a singlecellular pathway. Specifically, we delve into the potentialpoints of influence that the heat shock protein 90 (HSP90)chaperone machinery has on telomere protein biology. Bycoordinating the assembly and disassembly of numeroustelomere complexes, we speculate that HSP90 and itsassociated co-chaperones facilitate a precise and efficientworking environment that is highly beneficial for telomerefunction.
Corresponding author: Freeman, B.C. ([email protected]).
384 0968-0004/$ – see front matter . Published by Elsev
The telomere protein systemTelomeres consist of a tract of G-rich DNA motifs culmi-nating in a 3’ single-stranded overhang (telomeric DNA)that is recognized by a collection of associated factors(telomere-binding proteins) that maintain the terminalDNA at an appropriate length to preserve genomestability and cell viability [4]. A multitude of events mustoccur at telomeres to achieve function and all of theactivities are challenged by a need to work preciselyand efficiently within a limited time-frame. Notably, asingle shortened telomeric end is sufficient to inducecellular senescence, whereas an unregulated telomeresystem can lead to malignant growth [5,6]. Hence, telo-mere biology presents an ideal molecular model to furtherour understanding of the cellular mechanisms that arerequired to proficiently direct and drive a pathway, whichaffects homeostasis.
Multiple protein complexes assemble at telomeres toexecute specific functions that are required for protectionand extension of the terminal DNA [4,7]. In general, thecomplexity of the telomere varies in different stages of thecell cycle; however, specific cellular needs can also directmolecular decisions. For instance, telomeric DNA isextended only in S-phase, but not all telomeres areextended in every cell cycle [8]. In a short temporal window(<15 min) during late S-phase, a decision is made on thebasis of the length of the double-stranded telomeric tract(i.e. the number of telomeric repeats are essentiallycounted) to either generate a telomerase-extendable stateor form an unextendable structure [9].
If a telomeric end is approaching a critically shortenedstate, which would trigger cellular senescence if reducedfurther, then the 3’ overhang will be extended [6]. Exten-sion of the DNA requires several different aspects ofS-phase-specific activity, including DNA resectioning,extending and fill-in replicating, which are all mediatedby distinct protein complexes (Figure 1). Interestingly,components of each assemblage are capable of binding tothe single-stranded, G-rich overhang, which presents theopportunity for inhibitory competitive DNA interactions.Yet, all three events operate efficiently within the allottednarrow time window following DNA replication in S phase.If, however, the telomeric DNA tract is sufficiently long,the structure will remain in an unextendable or cappedstate, although the composition of the cap will vary as thecell cycle progresses (Figure 1) [4,7]. Hence, the telomereenvironment must be maintained in a dynamic state inorder to respond to the signals that dictate function and toperform the work that is required to maintain the chro-mosomal termini. But what mechanism(s) are used toproperly assemble (i.e. avoid competitive binding) and
ier Ltd. doi:10.1016/j.tibs.2010.02.005 Trends in Biochemical Sciences 35 (2010) 384–391
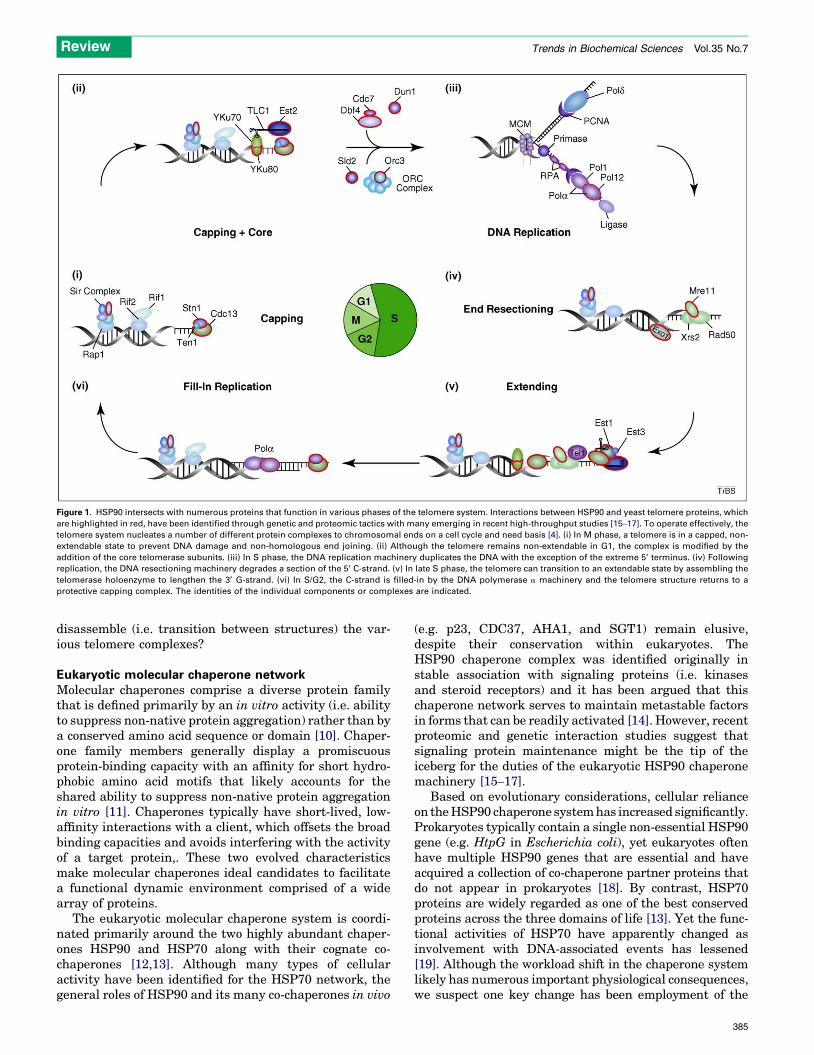
Figure 1. HSP90 intersects with numerous proteins that function in various phases of the telomere system. Interactions between HSP90 and yeast telomere proteins, which
are highlighted in red, have been identified through genetic and proteomic tactics with many emerging in recent high-throughput studies [15–17]. To operate effectively, the
telomere system nucleates a number of different protein complexes to chromosomal ends on a cell cycle and need basis [4]. (i) In M phase, a telomere is in a capped, non-
extendable state to prevent DNA damage and non-homologous end joining. (ii) Although the telomere remains non-extendable in G1, the complex is modified by the
addition of the core telomerase subunits. (iii) In S phase, the DNA replication machinery duplicates the DNA with the exception of the extreme 5’ terminus. (iv) Following
replication, the DNA resectioning machinery degrades a section of the 5’ C-strand. (v) In late S phase, the telomere can transition to an extendable state by assembling the
telomerase holoenzyme to lengthen the 3’ G-strand. (vi) In S/G2, the C-strand is filled-in by the DNA polymerase a machinery and the telomere structure returns to a
protective capping complex. The identities of the individual components or complexes are indicated.
Review Trends in Biochemical Sciences Vol.35 No.7
disassemble (i.e. transition between structures) the var-ious telomere complexes?
Eukaryotic molecular chaperone networkMolecular chaperones comprise a diverse protein familythat is defined primarily by an in vitro activity (i.e. abilityto suppress non-native protein aggregation) rather than bya conserved amino acid sequence or domain [10]. Chaper-one family members generally display a promiscuousprotein-binding capacity with an affinity for short hydro-phobic amino acid motifs that likely accounts for theshared ability to suppress non-native protein aggregationin vitro [11]. Chaperones typically have short-lived, low-affinity interactions with a client, which offsets the broadbinding capacities and avoids interfering with the activityof a target protein,. These two evolved characteristicsmake molecular chaperones ideal candidates to facilitatea functional dynamic environment comprised of a widearray of proteins.
The eukaryotic molecular chaperone system is coordi-nated primarily around the two highly abundant chaper-ones HSP90 and HSP70 along with their cognate co-chaperones [12,13]. Although many types of cellularactivity have been identified for the HSP70 network, thegeneral roles of HSP90 and its many co-chaperones in vivo
(e.g. p23, CDC37, AHA1, and SGT1) remain elusive,despite their conservation within eukaryotes. TheHSP90 chaperone complex was identified originally instable association with signaling proteins (i.e. kinasesand steroid receptors) and it has been argued that thischaperone network serves to maintain metastable factorsin forms that can be readily activated [14]. However, recentproteomic and genetic interaction studies suggest thatsignaling protein maintenance might be the tip of theiceberg for the duties of the eukaryotic HSP90 chaperonemachinery [15–17].
Based on evolutionary considerations, cellular relianceon theHSP90 chaperone systemhas increased significantly.Prokaryotes typically contain a single non-essential HSP90gene (e.g. HtpG in Escherichia coli), yet eukaryotes oftenhave multiple HSP90 genes that are essential and haveacquired a collection of co-chaperone partner proteins thatdo not appear in prokaryotes [18]. By contrast, HSP70proteins are widely regarded as one of the best conservedproteins across the three domains of life [13]. Yet the func-tional activities of HSP70 have apparently changed asinvolvement with DNA-associated events has lessened[19]. Although the workload shift in the chaperone systemlikely has numerous important physiological consequences,we suspect one key change has been employment of the
385

Review Trends in Biochemical Sciences Vol.35 No.7
HSP90 machine to foster homeostasis by modulatingprotein–DNA dynamics.
Historically, molecular chaperones have been recog-nized as factors that mediate the association and disas-sociation of protein complexes [20]. Recent studies indicatethat certain chaperones serve comparable roles withprotein–DNA structures [21–28]. For instance, severalmolecular chaperones, including HSP90, HSP70 andp23, can promote protein–DNA dynamics of diverse tar-gets, including components involved in RNA transcription,telomere maintenance, DNA repair and DNA replication[3,29–31]. Here, we focus on the role of the HSP90 chaper-one system inmodulating the assembly and disassembly oftelomere structures.
The HSP90 machine associates with the telomeraseenzymeThe initial connection between HSP90 and telomere com-ponents was made while attempting to identify cellularfactors required for telomerase holoenzyme function [32].The authors discovered that HSP90 and the p23 co-cha-perone interact with the protein subunit of human telo-merase (hTERT) through a yeast two-hybrid screen.Telomerase is a specialized reverse transcriptase (RT) inwhich the core enzyme comprises a protein module (TERTin humans and Est2 in yeast) and an RNA subunit (humanTR and yeast TLC1) [4,7]. Telomerase utilizes the RNA asa template to extend the 3’ chromosomal ends by 100nucleotides or more per cell cycle by reiteratively append-ing a short (6–8 bases) telomeric DNA repeat during eachbinding event [9,33]. Core human telomerase is sufficientfor robust DNA extension activity in vitro but core yeasttelomerase is not [34,35]. Nonetheless, both homologs relyon a multitude of additional proteins for proper control;HSP90 and p23 represent the first two proteins shown tointeract directly with telomerase and contribute to itsenzymatic activity [32].
The link between telomerase and molecular chaperoneswas extended by the finding that additional chaperones,including HSP70, HSP40 and HOP (HSP90/HSP70 orga-nizing protein), associate with the core human enzyme,presumably to assemble the RT and RNA template in vitro[36]. This observation fits with the classic steroid receptor–chaperone model in which chaperones are required togenerate andmaintain the hormone-binding state of recep-tors through a stable interaction that persists until thereceptor is hormone-activated [14]. Surprisingly, HSP90and p23 remain associated with telomerase even afterextending a telomeric DNA substrate, suggesting thatthese chaperones have a role beyond telomerase proteinfolding [36]. Notably, both HSP90 and p23 also functionwith DNA-associated steroid receptors to promote adynamic DNA-binding cycle that is required for propergene regulation [22,24]. If HSP90 and p23 regulate bothtelomerase and receptors at or near DNA, then perhapsthese chaperones have a general cellular function withdiverse DNA-binding proteins.
Unpredictably, an early yeast genetic study showed thatover-expression of either Hsp82 or Hsc82 (the two yeastHSP90 isoforms) led to telomeric DNA shortening, thussuggesting a negative effect of Hsp82/Hsc82 in telomere
386
DNA maintenance in vivo [37]. The cause of the decreasedtelomere DNA length was not apparent; however, thepotential for Hsp82 to affect telomere-binding proteinsbesides telomerase was revealed when HSP82 was ident-ified as a high-copy suppressor of two mutations (cdc13-1and stn1-157) known to alter telomere DNA length [37].Hence, HSP90 proteins might affect multiple telomere-associated proteins.
The observation that telomeric DNA length was essen-tially normal in a strain expressing limited Hsp82 levels(�10% of normal) further supported the contention thatHsp82 does not have a positive effect on telomerase [37].An Hsp82 protein reduction tactic was used because theeukaryotic HSP90 genes are essential and therefore it isnot possible to test for in vivo effects in the absence of anHSP90 protein. It should be noted, however, that normalyeast has �500,000 Hsp82 molecules per cell [38]. Thus, iftelomerase is a high-affinity substrate for Hsp82, a changein telomeric DNA length might not be expected in yeastexpressing �50,000 Hsp82 molecules.
At the very least, the two initial reports investigatingthe connection between HSP90 and telomere proteinssuggested that the relationship is complicated. Althoughthe discord between the in vitro and in vivo data might beunsatisfying, it is not entirely surprising given the natureof the cellular chaperone network (i.e. abundant and pro-miscuous proteins). For example, HSP70 is the fundamen-tal nascent chain-binding protein in vivo [39]. Yet, loss ofthe E. coli HSP70 gene DnaK results in only mild pheno-types [40]. However, in the absence of both DnaK and thetrigger factor (TF) molecular chaperone, cells die owing tomassive protein aggregation [41]. Nonetheless, based on invitro assays, a role for DnaK in protein folding had beenlong accepted before the discovery of the synthetic lethalphenotype between DnaK and TF. Therefore, in vitrostudies can identify important chaperone functions thatare not readily apparent by typical in vivo work given theredundant and/or compensatory nature of the cellularmolecular chaperone system.
To dissect the functional role of HSP90 with telomer-ase, suitable genetic conditions, in conjunction with amore detailed biochemical analysis, were necessary. Toavoid potential complications that might occur in thepresence of wild-type HSP90 proteins, Toogun et al. [27]screened yeast expressing select hsp82 or hsc82 alleles inan attempt to identify telomere-associatedmutants.Nota-bly, several mutants displayed both a decreased telomereDNA length in vivo and a decrease in telomerase DNAextension activity in vitro. Further mechanistic workrevealed that human and yeast HSP90 proteins canpromote DNA binding and nucleotide affinity for telomer-ase [27,42]. Hence, HSP90 chaperones contribute to telo-mere DNA lengthmaintenance by supporting telomerase.It is noteworthy that yeast Sba1 (the p23 ortholog) candissociate telomerase DNA complexes, suggesting thatHsp82 and Sba1 might coordinate a dynamic telomeraseDNA-binding cycle that parallels the chaperone receptormodel [2,26]. Whether direct communication betweenHsp82 and Sba1 is required to mediate the dynamics orif the chaperones function autonomously is yet to beaddressed. Nonetheless, if Hsp82 supports telomerase,

Table 1. Potential yeast Hsp90 telomere interacting partners
Gene Function Reference
SIR2 Chromatin silencing [15]
CTF4 DNA repair [16]
EXO1 DNA resectioning [16]
MRE11 DNA resectioning [16]
POL12 Fill-in replication [16]
RAD52 Telomere maintenance [16]
SLX8 Telomere maintenance [16]
SUM1 Chromatin silencing [16]
BUD32 Telomere maintenance [17]
CDC7 Chromatin silencing [17]
CDC13 Extending/Capping [17]
EST1 Extending [17]
MEC1 Extending [17]
RAD51 Telomere maintenance [17]
RAD59 Telomere maintenance [17]
STN1 Capping [17]
SWD3 Telomere maintenance [17]
YKU80 Capping + Core [17]
SBA1 Extending [26]
EST2 Extending [27]
Review Trends in Biochemical Sciences Vol.35 No.7
then why does Hsp82 over-expression lead to telomereDNA shortening [37]?
Hsp82 intersects with numerous telomere proteinsGenetic studies have suggested that Hsp82/Hsc82 affectother telomere proteins, given that yeast expressing anHsc82 S481Y mutant had shortened telomeric DNA butnormal telomerase activity in vitro [27] and, more notably,that HSP82 is a high-copy suppressor of the two mutantswith a telomere defect, cdc13-1 and stn1-157 [37]. BothCdc13 (cell division cycle 13) and Stn1 (suppressor ofCdc13) are crucial for telomere DNA maintenance:Cdc13 is a central recruitment factor required for nucleat-ing both extension (i.e. telomerase) and protection of tel-omere structures, and Stn1 is an essential component ofthe protection (capping) complex [43,44]. In addition toCdc13 and Stn1, recent high-throughput studies haveidentified numerous telomere-associated factors as poten-tial Hsp82 targets either through genetic or proteomicinteractions [15–17] (Table 1).
The telomere protein system must have a variety ofactivities, which include protecting, extending and repli-cating the terminal DNA in order to maintain the chro-mosomal ends (Figure 1). Although telomeres from alleukaryotes perform similar functions, detailed studies inbudding yeast make it a particularly useful model system.During each cell cycle the telomere protein compositiontransitions from one complex to the next depending uponphase and marked need. For example, at telomeres withrelatively short telomeric DNA tracts, the organizationwillfluctuate from an M-phase capping complex (minimallyCdc13–Stn1–Ten1) to a G1 capping structure that includesYKu70–YKu80-stabilized telomerase (core subunits) to theDNA replication machinery in S-phase to a DNA resection-ing assemblage (involving the MRX complex and the ExoIand SaeI nucleases) that degrades a segment of the C-strand in late S phase to a DNA-extending structure(telomerase–Est3–Est1–Cdc13) for the G-rich 3’-overhangstrand to a DNA polymerase a complex (Pol1, Pol12 andprimase) that fills in the complementary C-strand duringS/G2 and then returns to the capping structure found in M
phase (Figure 1). In addition to this route, the telomerestructure can remain capped throughout the cell cycle. Thedecision to follow an extendable or unextendable path isdetermined, in part, through a counting mechanism invol-ving the number of double-stranded telomeric DNA motifs(i.e. length of terminal DNA), the Rap1–Rif1–Rif2 proteincomplex, and the telomere-associated kinases Tel1 andMec1 [7]. Intriguingly, Hsp82 interacts with at least oneoperational component required for each of these telomereevents (highlighted in red in Figure 1). But what functionalrole does Hsp82 have with these various factors?
The telomere system is confronted with difficulties thatmight be alleviated with the assistance of a molecularchaperone. Specifically, telomeres face significant proteinmotility challenges; there is a need to transition betweenmultiple structures and so individual telomere assembliesrequire mobility to function efficiently (e.g. telomerelengthening requires telomerase repositioning) and thesystem must avoid potential competitive DNA-bindingevents that would otherwise sterically interfere with func-tion (e.g. Cdc13, Est1 (ever shorter telomeres), MRX, Stn1–
Ten1 and telomerase are all single-stranded, G-rich DNA-binding proteins) [4,7]. Although there are a number ofpossible explanations to account for the various challenges,we favor a model in which the telomere environment ismaintained in a dynamic state by the HSP90machinery. Iftelomeres were governed by dynamic remodeling, then theindividual components would exchange rapidly, fosteringefficient transitions between complexes and avoiding com-petitive binding events through transient DNA inter-actions. Notably, a recent report showed that Hsp82could solve these issues for at least one telomere com-ponent, Cdc13 [28].
Cdc13 plays a central role in yeast telomere biology, as itis required to nucleate both capping and extending com-plexes to the DNA ends [43]. Genetic studies demonstratethat Cdc13 protects chromosomal termini alongwith Stn1–
Ten1 and is required to extend the telomeric DNA inconjunction with the telomerase holoenzyme [4,7]. Todetermine whether Hsp82 influences Cdc13-mediatedevents, an in vitro system was established in whichCdc13 alone could stimulate telomerase DNA extensionactivity, but an unextendable telomeric DNA structureformed in the co-presence of Stn1–Ten1 [28]. Significantly,addition of Hsp82 dissociated the Cdc13–Stn1–Ten1–DNAcomplex but did not impede the ability of Cdc13 to stimu-late telomerase. Hence, Hsp82 permitted a transition be-tween the capping and extending assemblies withoutinterfering with the positive telomerase regulatory eventsprovided by Cdc13. Whether this disassembly activityexplains why HSP82 is a high-copy suppressor of thecdc13-1 and stn1-157 mutations remains unknown. How-ever, the ability of Hsp82 to promote DNA dissociation (i.e.Cdc13) and association (i.e. telomerase) of different telo-mere proteins might account for the extensive interactionnetwork between Hsp82 and telomere proteins, as it wouldcreate a self-organizing telomere environment (Box 1).
To illustrate, we expand upon the conceivableevents that occur in late S phase when telomeres aredirected to either extendable or unextendable states [7].Initial expectations posited that all telomeric DNA would
387

Box 1. Molecular chaperone-mediated protein self-
organization
By modulating the DNA-bound state of a broad array of telomere
proteins, HSP90 could create a local telomere environment function-
ing through the principles of self-organization. Self-organizing
systems operate by continuously breaking down into simpler
components and either remaining in a more elementary state
(termination of activity) or reassembling if the appropriate compo-
nents and energy resources are available (transition between
complexes) [68]. Essentially, if telomeres were maintained in a
self-organizing state, then the system would readily shift between
different functional states upon the integration of either additional
proteins or signals. The capacity to rapidly assimilate information
and generate the correct action is likely very relevant for telomeres
given the cell cycle-restricted timing of events and the number of
factors and complexes that must be assembled and disassembled in
order to function correctly [4,7].
Review Trends in Biochemical Sciences Vol.35 No.7
be lengthened during the duplication of each chromosome[45,46]. However, direct testing showed that individualtelomere size is not increased in every cell cycle [9].Furthermore, the extended telomeres displayed a sizeablevariability in the amount of DNA added, as the interquar-tile range in extension length was 14–80 nucleotides. Toaccount for the distribution in extension lengths, telomeresmust be governed by a system that reacts continuously toregulatory signals and can control telomerase action beforeand after each added telomeric DNA repeat. To be able torespond to the incoming signals, we suspect that the
Figure 2. HSP90 maintains the telomere environment in a dynamic state that permits r
telomeres are guided to either a telomerase extendable state or maintained in a protective
order to transition to a required structure during the narrow time-frame afforded in late S ph
into various complexes depending upon the available components (e.g. Stn1–Ten1 versus
fosters both dissociation (protection components) and association (extension subunits) of
extension complex, telomeres are able to transition efficiently between the different func
388
HSP90 chaperone systemmaintains the telomere environ-ment in a dynamic state (Figure 2). In our model, Hsp82promotes the continual dissociation of the unextendablecomplex (Cdc13–Stn1–Ten1), which will reform unless therequisite components and signals for constructing theextendable structure are present. For example, assemblyof the telomerase holoenzyme is restricted to S-phasebecause the essential Est1 subunit is expressed only atthis point of the cell cycle [47]. In addition, the actions ofcertain kinases (e.g. cyclin-dependent kinase 1 (Cdk1) andTel1) appear to guide telomeres towards the requisite stateby minimally modifying Cdc13. Cdk1-mediated phos-phorylation likely disfavors Cdc13 interactions withStn1 and fosters association with Est1 [48]. In addition,the Tel1 kinase is required for telomere DNAmaintenance,and its potential phosphorylation targets include Cdc13and the MRX complex subunit Xrs2 [49,50]. However, theprecise mechanistic contributions of Tel1 signaling areincompletely understood.
After nucleation of the modified telomerase holoenzymeat a telomere, Hsp82 and its co-chaperone Sba1 are likelyto promote a cyclical telomerase action, as Hsp82 supportsDNA binding by telomerase and Sba1 dissociates telomer-ase DNA complexes [26,27]. How Hsp82 promotes bindingwhile Sba1 facilitates release is not clear, though it mightoccur through a chaperone-mediated opening and closingof the DNA-binding cleft. Nevertheless, rapid cycling oftelomerase on and off the telomeric DNA would be
apid and efficient reactions to incoming signals and components. In late S phase,
unextendable form depending upon the length of the adjacent telomeric DNA [7]. In
ase, telomere structures are likely to be continuously disassembled and reassembled
telomerase–Est1–Est3) and local signals (e.g. Cdk1 and Tel1 kinases). In yeast, Hsp82
telomere structures. In conjunction with the Sba1 co-chaperone that destabilizes the
tional states in order to maintain the chromosomal termini properly.

Box 2. HSP90 and telomere therapeutics
The most common HSP90 inhibitors are based upon the natural
benzoquinone ansamycins (e.g. geldanamycin) that are bound in
the ATP-binding pocket of HSP90 and inhibit both nucleotide
hydrolysis and chaperone functions [69–71]. Geldanamycin deriva-
tives, including 17AAG and IPI504, that have increased hydrophili-
city and lowered hepatotoxicity, are currently in Phase I-III trials
(http://clinicaltrials.gov/ct2/results?term=hsp90). Although the clin-
ical trials for various HSP90 inhibitors (from benzoquinones to
purine-scaffold compounds) are promising, the single compound
studies appear to have limited efficacy [72,73]. Fortunately, the lack
of significant toxicity and an ability to trace certain HSP90 inhibitors
position these compounds as ideal agents for combinatorial
treatments [74].
Telomere therapeutics has been approached from several angles,
although the two most promising avenues have focused on the
telomerase RNA template and the telomeric DNA [75,76]. Mamma-
lian telomeric DNA repeats can form G-quartet structures and small
molecules designed to foster the four-stranded quartet fold cause
telomeric DNA shortening by uncapping the termini, which
eventually leads to cellular senescence [65]. Compounds targeting
telomerase are often designed as RNA antagonists that hybridize
with the template region and block DNA extension activity. The
most promising antagonist is GRN163L, which causes progressive
telomere shortening and inhibits cell growth. Unfortunately,
telomere therapies typically have a substantial effective lag time,
which depends on the length of the initial telomeric DNA in the
treated cells, in order to induce replicative senescence and prevent
tumor growth [65]. Hence, telomere disruption, either through
deprotection of the ends or by blocking telomerase activity, is likely
insufficient for cancer treatment given the time-scale necessary for
an effective response. However, given the long-term potential of
preventing tumor regrowth, the telomere inhibitors might prove
advantageous in conjunction with other cytotoxic agents [74].
Review Trends in Biochemical Sciences Vol.35 No.7
beneficial for effective extension of the chromosomal ter-mini. Following sufficient telomeric DNA lengthening,Sba1-mediated dissociation of the telomerase complexand action by unidentified phosphatases (which wouldreverse the influence of the Cdk1 and Tel1 kinases), arelikely to promote the reformation of the Cdc13–Stn1–Ten1unextendable structure. Hence, by maintaining the telo-mere components in a dynamic state, the Hsp82 chaperonemachine would permit the system to transition efficientlybetween different operative states, which are guided bypost-translational modifications and the presence of therequired subunits; the chaperone-mediated interchangewould also foster effective action for complexes that requiremovement (e.g. telomerase-mediated DNA extension).
HSP90, telomere proteins and cancerThe HSP90 chaperone network serves a wide variety ofclient proteins operating in a number of pathways requiredfor cell viability, including telomere maintenance, signaltransduction, transcription, protein transport, chromatinremodeling, and protein stabilization [12,14]. The influ-ence of the HSP90 system on these cellular functions ismagnified during periods of stress [17]. Normally, theHSP90 system aids cell survival; however, during periodsof chronic physiological shock (e.g. cancer) the beneficialinfluence of HSP90 might shift to a detrimental role, asHSP90 appears to be central for supporting a broad rangeof cancers [51]. As might be anticipated, components of theHSP90 machinery, including HSP90 and p23, increase inexpression with the development of metastatic growth [52–
55]. In a comparable manner, telomerase activity appearsto play a central role in general cancer progression [5].
Mammalian telomerase is expressed only in highlyproliferative cell types such as germ and somatic stemcells [56]. Normal differentiated cells typically reach repli-cative senescence once the DNA length of even a singletelomere is reduced to a certain size [6]. In contrast toaverage cells, 85�90% of all malignant tissues displaytelomerase activity, and tumors without telomerase areoften diagnosed as benign [57]. Telomerase up-regulationminimally averts senescence and allows the altered cell toperpetuate and generate additional cancerous cells (e.g.tumor formation). Intriguingly, the human protooncogenec-Myc can activate transcription of both HSP90A andTERT [58]. Although a rise in telomerase expression is acommon route to increased activity, other mechanismsare apparent. For example, experiments using a prostatecancer model showed that increased telomerase activityis correlated with a rise in HSP90 and p23 levels, butnot in telomerase protein expression, as cells progresstowards malignancy [52]. Modulation of telomeraseactivity through HSP90-dependent mechanisms has beensuggested to occur in numerous cancer types, includinghepatomas, adrenalomas, glioblastomas and melanomas[59–62]. Thus, telomerase activity and molecular chaper-one levels correlate with cancer cell progression and thesephenomena appear to be linked. The high prevalence ofHSP90 and telomere defects in a broad range of cancersmakes these ideal candidates for medicinal purposes andboth have been pursued asmolecular targets for anticancertherapeutics in clinical trials (Box 2).
Several reports exploiting cell culture models indicatethat a combinatorial tactic incorporating HSP90 and tel-omere inhibitors might be rewarding. Given the reliance oftelomerase on HSP90, it was anticipated that reducingHSP90 activity would foster telomerase inhibition. Indeed,the combination of the template antagonist GRN163L and17-(allylamino)-17-demethoxygeldanamycin (17AAG), andHSP90 inhibitor, greatly increased the induced death ofmyeloma cells [63]. As telomerase has not been shown tosupport HSP90 activity, it was perhaps surprising to notethat reducing the basal levels of the telomerase proteinsubunit TERT by anRNAi strategy led to increased 17AAGpotency in culture [64]. Furthermore, the application of theG-quartet compound RHPS4 in conjunction with 17AAGhad a synergistic inhibitory effect on breast cancer cellgrowth [65]. Conceivably, the finding that HSP90 modu-lates telomere-capping components helps rationalize theobserved synergy with the telomere-uncapping drug [28].Together, the inhibitor studies support the premise thatcombinatorial targeting of HSP90 and telomeres will havea beneficial medicinal outcome.
In addition to reducing or preventing tumor growth,mixing HSP90 and telomere compounds might proveadvantageous for other reasons. As mentioned, telomeraseactivity is present in dividing cells, including stem andgerm lines, and therefore a means to avoid these cell typeswould be beneficial. In general, cancer cells display an�100-fold higher affinity for HSP90 inhibitors relativeto normal tissue [66]. Hence, if telomere compoundswere delivered with HSP90 inhibitors, a selectiveadvantage might be gained in targeting malignant tissues.
389

Review Trends in Biochemical Sciences Vol.35 No.7
Furthermore, as 17AAG can be labeled and traced withinwhole organisms, it might be possible to apply additionalcytotoxic agents locally [67]. Hence, the use of HSP90inhibitors and telomere drugs together could provide amulti-faceted therapeutic benefit.
Concluding remarksTelomere protein biology serves as an excellent molecularparadigm to understand how the efficiency of a singlesystem can impact homeostasis, as telomere dysfunctioncan lead to either cellular senescence or uncontrolledgrowth (i.e. cancer). Given the number of employedproteins with common binding specificity (Figure 1), adecrease in telomere efficiency is highly plausible due tocomplex misassembly or impaired structure disassembly.The fundamental properties of molecular chaperones(abundant proteins with promiscuous but weak bindingactivities) would help the telomere system avoid thesechallenges by cultivating a self-organizing environment(Box 1). In essence, the ability of the HSP90 chaperonenetwork to foster telomere protein dynamics parallels themore established molecular chaperone roles in proteinfolding in which a chaperone does not dictate the finalfolded structure (path direction) but rather helps the nas-cent chain (system) avoid off-pathway energy barriers thatcommonly occur in protein folding (multi-step) energylandscapes. Given the large number of HSP90 interactingtelomere targets (Figure 1), it will be interesting to learnwhether future studies continue to reveal a role for theHSP90 network in creating a dynamic telomere environ-ment.
AcknowledgementsWe apologize to all authors whose work could not be cited due to spacelimitations. We appreciate the helpful comments on the manuscript madeby F. Echtenkamp and E. Zelin. D.C.D. was supported by a CMB NIHtraining grant and B.C.F. was supported by Public Service grantDK074270.
References1 Misteli, T. (2001) The concept of self-organization in cellular
architecture. J. Cell Biol. 155, 181–1852 Freeman, B.C. and Yamamoto, K.R. (2001) Continuous recycling: a
mechanism for modulatory signal transduction. Trends Biochem. Sci.26, 285–290
3 DeZwaan, D.C. and Freeman, B.C. (2008) HSP90: the Rosetta stone forprotein dynamics? Cell Cycle 7, 1006–1012
4 Gilson, E. and Geli, V. (2007) How telomeres are replicated. Nat. Rev.Mol. Cell Biol. 8, 825–838
5 Shay, J.W. and Wright, W.E. (1996) Telomerase activity in humancancer. Curr. Opin. Oncol. 8, 66–71
6 Hemann,M.T. et al. (2001) The shortest telomere, not average telomerelength, is critical for cell viability and chromosome stability. Cell 107,67–77
7 Shore, D. and Bianchi, A. (2009) Telomere length regulation: couplingDNA end processing to feedback regulation of telomerase.EMBO J. 28,2309–2322
8 Marcand, S. et al. (2000) Cell cycle restriction of telomere elongation.Curr. Biol. 10, 487–490
9 Teixeira, M.T. et al. (2004) Telomere length homeostasis is achieved viaa switch between telomerase extendible and -nonextendible states.Cell117, 323–335
10 Ellis, R.J. and van der Vies, S.M. (1991) Molecular chaperones. Annu.Rev. Biochem. 60, 321–347
11 Hendrick, J.P. and Hartl, F.U. (1993) Molecular chaperone functions ofheat-shock proteins. Annu. Rev. Biochem. 62, 349–384
390
12 Wegele, H. et al. (2004) HSP70 and HSP90–a relay team for proteinfolding. Rev. Physiol. Biochem. Pharmacol. 151, 1–44
13 Mayer, M.P. and Bukau, B. (2005) HSP70 chaperones: cellularfunctions and molecular mechanism. Cell Mol. Life Sci. 62, 670–684
14 Pratt, W.B. and Toft, D.O. (2003) Regulation of signaling proteinfunction and trafficking by the HSP90/HSP70-based chaperonemachinery. Exp. Biol. Med. 228, 111–133
15 Millson, S.H. et al. (2005) A two-hybrid screen of the yeast proteome forHSP90 interactors uncovers a novel HSP90 chaperone requirement inthe activity of a stress-activated mitogen-activated protein kinase,Slt2p (Mpk1p). Euk. Cell 4, 849–860
16 Zhao, R. et al. (2005) Navigating the chaperone network: an integrativemap of physical and genetic interactions mediated by the HSP90chaperone. Cell 120, 715–727
17 McClellan, A.J. et al. (2007) Diverse cellular functions of the HSP90molecular chaperone uncovered using systems approaches. Cell 131,121–135
18 Johnson, J.L. and Brown, C. (2009) Plasticity of the HSP90 chaperonemachine in divergent eukaryotic organisms.Cell Stress Chaperones 14,83–94
19 Ziemieowicz, A. et al. (2001) Calf thymus Hsc70 and Hsc40 cansubstitute for DnaK and DnaJ function in protein renaturation butnot in bacteriophage DNA replication. FEBS Lett. 507, 11–15
20 Ellis, R.J. (2007) Protein misassembly: macromolecular crowding andmolecular chaperones. Adv. Exp. Med. Biol. 594, 1–13
21 Shaknovich, R. et al. (1992) Conformational activation of a basic helix-loop-helix protein (MyoD1) by the C-terminal region of murine HSP90(HSP84). Mol. Cell Biol. 12, 5059–5068
22 Freeman, B.C. and Yamamoto, K.R. (2002) Disassembly oftranscriptional regulatory complexes by molecular chaperones.Science 296, 2232–2235
23 Muller, L. et al. (2004) HSP90 regulates the activity of wild type p53under physiological and elevated temperatures. J. Biol. Chem. 279,48846–48854
24 Stavreva, D.A. et al. (2004) Rapid glucocorticoid receptor exchange at apromoter is coupled to transcription and regulated by chaperones andproteasomes. Mol. Cell Biol. 24, 2682–2697
25 Walerych, D. et al. (2004) HSP90 chaperones wild-type p53 tumorsuppressor protein. J. Biol. Chem. 279, 48836–48845
26 Toogun, O.A. et al. (2007) The p23 molecular chaperone promotesfunctional telomerase complexes through DNA dissociation. Proc.Natl. Acad. Sci. U.S.A. 104, 5765–5770
27 Toogun, O.A. et al. (2008) The HSP90 molecular chaperone modulatesmultiple telomerase activities. Mol. Cell Biol. 28, 457–467
28 DeZwaan, D.C. et al. (2009) The Hsp82 molecular chaperone promotesa switch between unextendable and extendable telomere states. Nat.Struct. Mol. Biol. 16, 711–716
29 Konieczny, I. and Zylicz, M. (1999) Role of bacterial chaperones in DNAreplication. Genet. Eng. 21, 95–111
30 Richter, K. et al. (2007) The cellular world according to HSP90. Nat.Struct. Mol. Biol. 14, 90–94
31 Hager, G.L. et al. (2009) Transcription dynamics.Mol. Cell 35, 741–75332 Holt, S.E. et al. (1999) Functional requirement of p23 and HSP90 in
telomerase complexes. Genes Dev. 13, 817–82633 Britt-Compton, B. et al. (2009) Short telomeres are preferentially
elongated by telomerase in human cells. FEBS Lett. 583, 3076–308034 Cohn, M. and Blackburn, E.H. (1995) Telomerase in yeast. Science 269,
396–40035 Morin, G.B. (1989) The human telomere terminal transferase enzyme is
a ribonucleoprotein that synthesizes TTAGGG repeats.Cell 59, 521–52936 Forsythe, H.L. et al. (2001) Stable association of HSP90 and p23, but
Not HSP70, with active human telomerase. J. Biol. Chem. 276, 15571–
1557437 Grandin, N. and Charbonneau, M. (2001) HSP90 levels affect telomere
length in yeast. Mol. Genet. Genomics 265, 126–13438 Ghaemmaghami, S. et al. (2003) Global analysis of protein expression
in yeast. Nature 425, 737–74139 Beckmann, R.P. et al. (1990) Interaction of HSP70 with newly
synthesized proteins: implications for protein folding and assembly.Science 248, 850–854
40 Bukau, B. and Walker, G.C. (1989) Cellular defects caused by deletionof the Escherichia coli dnaK gene indicate roles for heat shock proteinin normal metabolism. J. Bacteriol. 171, 2337–2346

Review Trends in Biochemical Sciences Vol.35 No.7
41 Deuerling, E. et al. (1999) Trigger factor and DnaK cooperate in foldingof newly synthesized proteins. Nature 400, 693–696
42 Keppler, B.R. et al. (2006) The biochemical role of the heat shockprotein 90 chaperone complex in establishing human telomeraseactivity. J. Biol. Chem. 281, 19840–19848
43 Pennock, E. et al. (2001) Cdc13 delivers separate complexes to thetelomere for end protection and replication. Cell 104, 387–396
44 Grandin, N. et al. (1997) Stn1, a newSacchromyces cerevisiae protein, isimplicated in telomere size regulation in association with Cdc13.GenesDev. 11, 512–527
45 Shampay, J. and Blackburn, E.H. (1988) Generation of telomere-lengthheterogeneity in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci.U. S. A. 85, 534–538
46 Blackburn, E.H. et al. (1989) Recognition and elongation of telomeresby telomerase. Genome 31, 553–560
47 Osterhage, J.L. et al. (2006) Proteasome-dependent degradation ofEst1p regulates the cell cycle-restricted assembly of telomerase inSaccharomyces cerevisiae. Nat. Struct. Mol. Biol. 13, 720–728
48 Li, S. et al. (2009) Cdk1-dependent phosphorylation of Cdc13coordinates telomere elongation during cell-cycle progression. Cell136, 50–61
49 Nakada, D. et al. (2003) ATM-related Tel1 associates with double-strand breaks through an Xrs2-dependent mechanism. Genes Dev. 17,1957–1962
50 Tseng, S.F. et al. (2006) The telomerase-recruitment domain of thetelomere binding protein Cdc13 is regulated by Mec1p/Tel1p-dependent phosphorylation. Nucleic Acids Res. 34, 6327–6336
51 Whitesell, L. and Lindquist, S.L. (2005) HSP90 and the chaperoning ofcancer. Nat. Rev. Cancer 5, 761–772
52 Akalin, A. et al. (2001) A novel mechanism for chaperone-mediatedtelomerase regulation during prostate cancer progression. Cancer Res.61, 4791–4796
53 Oxelmark, E. et al. (2006) The co-chaperone p23 differentially regulatesestrogen receptor target genes and promotes tumor cell adhesion andinvasion. Mol. Cell Biol. 26, 5205–5213
54 Elmore, L.W. et al. (2008) Overexpression of telomerase-associatedchaperone proteins in prostatic intraepithelial neoplasia andcarcinomas. Oncol. Rep. 20, 613–617
55 McDowell, C.L. et al. (2009) Expression of HSP90 chaperone [corrected]proteins in human timor tissue. Int. J. Biol. Macromol. 45, 310–314
56 Flores, I. et al. (2006) Telomerase regulation and stem cell behaviour.Curr. Opin. Cell Biol. 18, 254–260
57 Cech, T.R. (2004) Beginning to understand the end of the chromosome.Cell 116, 273–279
58 Teng, S.C. et al. (2004) Direct activation of HSP90A transcription by c-Myc contributes to c-Myc-induced transformation. J. Biol. Chem. 279,14649–14655
59 Lee, C.L. et al. (2003) Strategic shotgun proteomics approach forefficient construction of an expression map of targeted proteinfamilies in hepatoma cell lines. Proteomics 3, 2472–2486
60 Shervington, A. et al. (2006) Glioma: what is the role of c-Myc, HSP90and telomerase? Mol. Cell Biochem 283, 1–9
61 Becker, B. et al. (2004) Induction of HSP90 protein expression inmalignant melanomas and melanoma metastases. Exp. Dermatol.13, 27–32
62 Mitsiades, C.S. et al. (2006) Antimyeloma activity of heat shockprotein-90 inhibition. Blood 107, 1092–1100
63 Shammas, M.A. et al. (2008) Telomerase inhibitor GRN163L inhibitsmyeloma cell growth in vitro and in vivo. Leukemia 22, 1410–1418
64 Villa, R. et al. (2003) Inhibition of telomerase activity by geldanamycinand 17-allylamino, 17-demethoxygeldanamycin in human melanomacells. Carcinogenesis 24, 851–859
65 Cookson, J.C. et al. (2005) Pharmacodynamics of the G-quadruplex-stabilizing telomerase inhibitor 3,11-difluoro-6,8,13-trimethyl-8H-quinto[4,3,2,k1]acridinium methosulfate (RHPS4) in vitro; activityin human tumour cells correlates with telomere length and can beenhanced, or antagonized, with cytotoxic agents. Mol. Pharmacol. 68,1551–1558
66 Kamal, A. et al. (2003) A high-affinity conformation of HSP90 conferstumour selectivity on HSP90 inhibitors. Nature 425, 407–410
67 Daozhen, C. et al. (2007) Synthesis of (131)I-labeled-[(131)I]iodo-17-allylamino-17-demethoxy geldanamycin ([(131)I]iodo-17-AAG) and itsbiodistribution in mice. Cancer Biother. Radiopharm. 22, 607–612
68 Prigogine, I. and Nicolis, G. (1971) Biological order, structure andinstabilities. Q. Rev. Biophys. 4, 107–148
69 Whitesell, L. and Cook, P. (1996) Stable and specific binding of heatshock protein 90 by geldanamycin disrupts glucocorticoid receptorfunction in intact cells. Mol. Endocrinol. 10, 705–712
70 Prodromou, C. et al. (1997) Identification and structuralcharacterization of the ATP/ADP-binding site in the HSP90molecular chaperone. Cell 90, 65–75
71 Wandinger, S.K. et al. (2008) TheHSP90 chaperonemachinery. J. Biol.Chem. 283, 18473–18477
72 Kim, Y.S. et al. (2009) Update on HSP90 inhibitors in clinical trial.Curr. Top. Med. Chem. 9, 1479–1492
73 Powers, M.V. and Workman, P. (2007) Inhibitors of the heat shockresponse: biology and pharmacology. FEBS Lett. 581, 3758–3769
74 Hwang, M. et al. (2009) HSP90 inhibitors: multi-targeted antitumoreffects and novel combinatorial therapeutic approaches in cancertherapy. Curr. Med. Chem. 16, 3081–3092
75 Chen, H. et al. (2009) Strategies targeting telomerase inhibition. Mol.Biotechnol. 41, 194–199
76 Burger, A.M. (2007) Highlights in experimental therapeutics. CancerLett. 245, 11–21
391





























