Embed Size (px)
Citation preview

Hydrogels preserve native phenotypes of valvularfibroblasts through an elasticity-regulatedPI3K/AKT pathwayHuan Wanga,b, Mark W. Tibbittb,c, Stephen J. Langera,b, Leslie A. Leinwanda,b, and Kristi S. Ansethb,c,d,1
aDepartment of Molecular, Cellular, and Developmental Biology, bBioFrontiers Institute, cDepartment of Chemical and Biological Engineering, and dHowardHughes Medical Institute, University of Colorado, Boulder, CO 80309
Edited by Rakesh K. Jain, Harvard Medical School and Massachusetts General Hospital, Boston, MA, and approved October 21, 2013 (received for reviewApril 17, 2013)
Matrix elasticity regulates proliferation, apoptosis, and differenti-ation of many cell types across various tissues. In particular,stiffened matrix in fibrotic lesions has been shown to promotepathogenic myofibroblast activation. To better understand theunderlying pathways by which fibroblasts mechano-sense matrixelasticity, we cultured primary valvular interstitial cells (VICs)isolated from porcine aortic valves on poly(ethylene glycol)-basedhydrogels with physiologically relevant and tunable elasticities.We show that soft hydrogels preserve the quiescent fibroblastphenotype of VICs much better than stiff plastic plates. Wedemonstrate that the PI3K/AKT pathway is significantly up-regulated when VICs are cultured on stiff gels or tissue culturepolystyrene compared with freshly isolated VICs. In contrast,myofibroblasts de-activate and pAKT/AKT decreases as early as2 h after reducing the substrate modulus. When PI3K or AKT isinhibited on stiff substrates, myofibroblast activation is blocked.When constitutively active PI3K is overexpressed, the myofibro-blast phenotype is promoted even on soft substrates. These datasuggest that valvular fibroblasts are sensing the changes in matrixelasticity through the PI3K/AKT pathway. This mechanism may beused by other mechano-sensitive cells in response to substrate mod-ulus, and this pathway may be a worthwhile target for treating ma-trix stiffness-associated diseases. Furthermore, hydrogels can bedesigned to recapitulate important mechanical cues in native tissuesto preserve aspects of the native phenotype of primary cells forunderstanding basic cellular responses to biophysical and biochemicalsignals, and for tissue-engineering applications.
mechanosensing | tissue stiffening |phosphatidylinositide 3-kinase signaling
Every cell has its distinct microenvironment. Fibroblasts residein the interstitial mesenchyme, maintaining the balance and
structure of the matrix; muscle satellite cells reside between thebasal lamina and muscle fibers, waiting to be activated uponinjury. In a basic sense, the cellular microenvironment is definedas the extracellular space surrounding and supporting the cell.This microenvironment is often comprised of extracellular ma-trix (ECM), soluble chemical factors, and neighboring cells. TheECM in particular does not just act as a passive scaffold, butinstructs cell fate through the coordinated and dynamic pre-sentation of biochemical and biophysical cues. For example, infibrotic tissues stiffened by excessive collagen deposition, resi-dent fibroblasts acquire an activated myofibroblast phenotypewith α-smooth muscle actin (αSMA) stress fibers (1). The in-tegrity and signaling of the ECM are essential for regulatingnormal cellular function and maintaining organ homeostasis.The aortic valve, which controls the unidirectional flow of blood
during heart contractions, is composed of elastin-, proteoglycan-and collagen-rich layers of ECM (2) and supports survival andmetabolism of valvular interstitial cells (VICs). VICs are the mainfibroblast population in cardiac valves that secrete ECM-relatedproteins for tissue development and homeostasis (3). Normal
valves maintain a bulk elastic modulus (E, a measure of the de-formation of a material to an external force) around 0.8–8 kPa (4).As valve stenosis develops, characterized by fibrotic stiffening andnodule-like calcification, the tissue can become as stiff as collag-enous bone (E, ∼27 ± 10 kPa) (5, 6). VICs have been shown todifferentiate from quiescent fibroblasts in normal valves intopathogenic myofibroblasts and osteoblast-like cells in diseasedvalves (7). Consistently, VICs are activated to myofibroblasts invitro in response to a high substrate modulus (E > 15 kPa), butthis phenotype can be reverted when the substrate modulus isreduced (8, 9). To better mimic the ECM environment of aorticvalves, we examined poly(ethylene glycol) (PEG)-based hydrogelsas culture substrates, which have physiologically relevant moduli(E, ∼7 kPa to ∼32 kPa), spanning the range of normal and dis-eased valves. Furthermore, the moduli of these PEG hydrogelscan be reduced in situ with light from 32 kPa to 7 kPa in ∼5 min(10), enabling close monitoring of changes in intracellular signal-ing in response to real-time modulus reduction.Despite the recognized importance of elasticity in regulating
the function and differentiation of many cell types, includingfibroblasts (1), mesenchymal stem cells (11), and myoblasts (12),the effect of matrix stiffness is still debated (13, 14) and manyquestions remain about how the mechanical cue of elasticity istranslated into intracellular signaling. Cells often bind to thematrix via integrins, mechano-sensitive transmembrance proteins
Significance
Valvular interstitial cells (VICs) are the principal cellular compo-nent of cardiac valves and maintain normal valve homeostasis.During valvular fibrosis, VICs differentiate into myofibroblastsand stiffen the valve matrix. The results in this report demon-strate that standard techniques of culturing VICs on supra-physiologically stiff, tissue-culture polystyrene cause a dramaticinduction of myofibroblast differentiation. In contrast, culturingVICs on soft, poly(ethylene glycol)-based hydrogels preservesthe native, quiescent phenotype. A detailed study of VICmechano-sensing reveals that matrix elasticity elicits patho-logic changes in VICs through PI3K/AKT signaling. A morecomplete understanding of the molecular mechanisms of VICmechano-biology may facilitate development of novel thera-peutics targeting downstream signaling in matrix-stiffness–as-sociated diseases, and may be applicable to fibrotic diseases indifferent tissues.
Author contributions: H.W., L.A.L., and K.S.A. designed research; H.W., M.W.T., and S.J.L.performed research; H.W., M.W.T., L.A.L., and K.S.A. analyzed data; and H.W., M.W.T.,L.A.L., and K.S.A. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The data reported in this paper have been deposited in the Gene Ex-pression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no. GSE48839).1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1306369110/-/DCSupplemental.
19336–19341 | PNAS | November 26, 2013 | vol. 110 | no. 48 www.pnas.org/cgi/doi/10.1073/pnas.1306369110
Dow
nloa
ded
by g
uest
on
Mar
ch 1
3, 2
020
that stimulate intracellular biochemical signaling, including FAK,ERK, and JNK (15). Dupont et al. showed that YAP and TAZ,two Hippo-related transcription factors, can serve as sensors ofECM stiffness in mesenchymal stem cells (11). Huang et al.showed that lung fibroblasts initiate nuclear translocation ofmegakaryoblastic leukemia factor-1 and RhoA/Rock activation onstiff substrata (16). All of these results suggest that matrix elasticitycan elicit complex cellular signaling and transcriptional responses,but further characterization is needed. In this study, we set out todevelop a tissue-mimicking culture platform that better preservesaspects of the native VIC phenotype to study how VICs respondto matrix elasticity changes via the PI3K/AKT pathway.
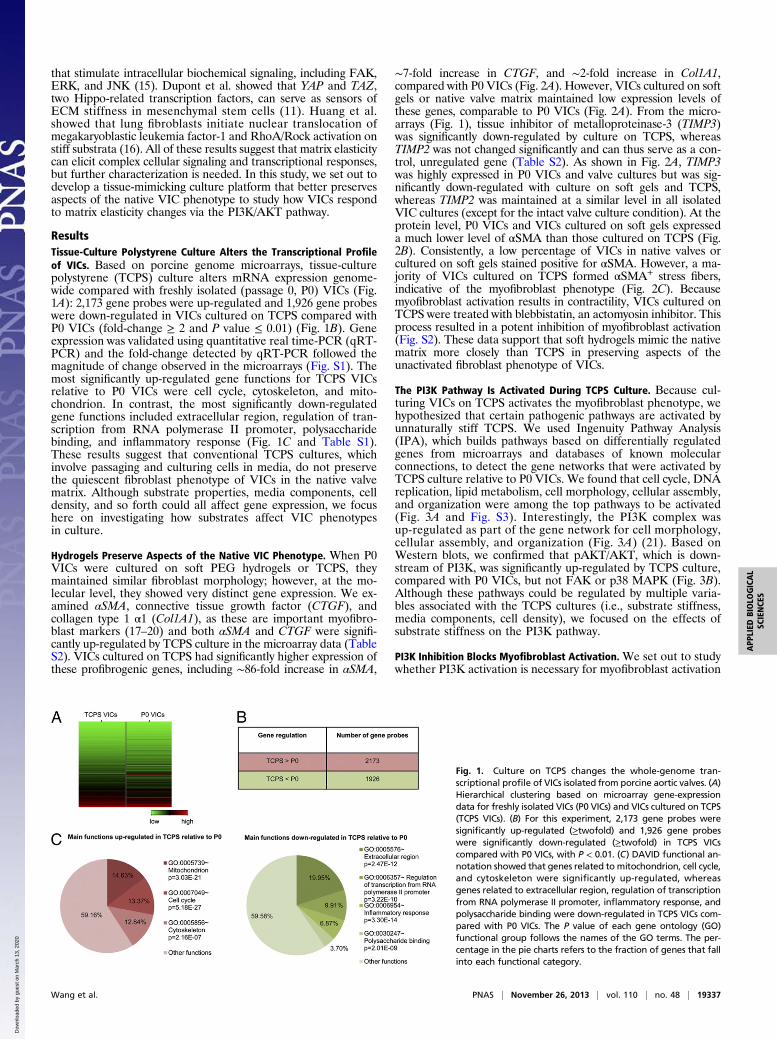
ResultsTissue-Culture Polystyrene Culture Alters the Transcriptional Profileof VICs. Based on porcine genome microarrays, tissue-culturepolystyrene (TCPS) culture alters mRNA expression genome-wide compared with freshly isolated (passage 0, P0) VICs (Fig.1A): 2,173 gene probes were up-regulated and 1,926 gene probeswere down-regulated in VICs cultured on TCPS compared withP0 VICs (fold-change ≥ 2 and P value ≤ 0.01) (Fig. 1B). Geneexpression was validated using quantitative real time-PCR (qRT-PCR) and the fold-change detected by qRT-PCR followed themagnitude of change observed in the microarrays (Fig. S1). Themost significantly up-regulated gene functions for TCPS VICsrelative to P0 VICs were cell cycle, cytoskeleton, and mito-chondrion. In contrast, the most significantly down-regulatedgene functions included extracellular region, regulation of tran-scription from RNA polymerase II promoter, polysaccharidebinding, and inflammatory response (Fig. 1C and Table S1).These results suggest that conventional TCPS cultures, whichinvolve passaging and culturing cells in media, do not preservethe quiescent fibroblast phenotype of VICs in the native valvematrix. Although substrate properties, media components, celldensity, and so forth could all affect gene expression, we focushere on investigating how substrates affect VIC phenotypesin culture.
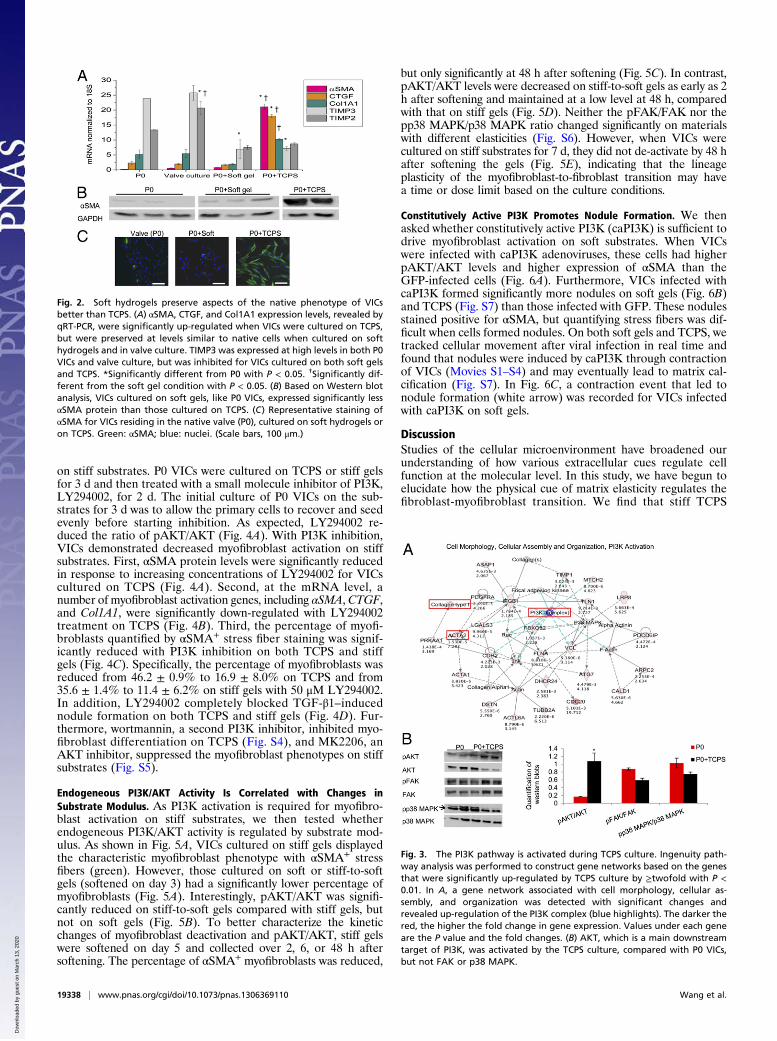
Hydrogels Preserve Aspects of the Native VIC Phenotype. When P0VICs were cultured on soft PEG hydrogels or TCPS, theymaintained similar fibroblast morphology; however, at the mo-lecular level, they showed very distinct gene expression. We ex-amined αSMA, connective tissue growth factor (CTGF), andcollagen type 1 α1 (Col1A1), as these are important myofibro-blast markers (17–20) and both αSMA and CTGF were signifi-cantly up-regulated by TCPS culture in the microarray data (TableS2). VICs cultured on TCPS had significantly higher expression ofthese profibrogenic genes, including ∼86-fold increase in αSMA,
∼7-fold increase in CTGF, and ∼2-fold increase in Col1A1,compared with P0 VICs (Fig. 2A). However, VICs cultured on softgels or native valve matrix maintained low expression levels ofthese genes, comparable to P0 VICs (Fig. 2A). From the micro-arrays (Fig. 1), tissue inhibitor of metalloproteinase-3 (TIMP3)was significantly down-regulated by culture on TCPS, whereasTIMP2 was not changed significantly and can thus serve as a con-trol, unregulated gene (Table S2). As shown in Fig. 2A, TIMP3was highly expressed in P0 VICs and valve cultures but was sig-nificantly down-regulated with culture on soft gels and TCPS,whereas TIMP2 was maintained at a similar level in all isolatedVIC cultures (except for the intact valve culture condition). At theprotein level, P0 VICs and VICs cultured on soft gels expresseda much lower level of αSMA than those cultured on TCPS (Fig.2B). Consistently, a low percentage of VICs in native valves orcultured on soft gels stained positive for αSMA. However, a ma-jority of VICs cultured on TCPS formed αSMA+ stress fibers,indicative of the myofibroblast phenotype (Fig. 2C). Becausemyofibroblast activation results in contractility, VICs cultured onTCPS were treated with blebbistatin, an actomyosin inhibitor. Thisprocess resulted in a potent inhibition of myofibroblast activation(Fig. S2). These data support that soft hydrogels mimic the nativematrix more closely than TCPS in preserving aspects of theunactivated fibroblast phenotype of VICs.
The PI3K Pathway Is Activated During TCPS Culture. Because cul-turing VICs on TCPS activates the myofibroblast phenotype, wehypothesized that certain pathogenic pathways are activated byunnaturally stiff TCPS. We used Ingenuity Pathway Analysis(IPA), which builds pathways based on differentially regulatedgenes from microarrays and databases of known molecularconnections, to detect the gene networks that were activated byTCPS culture relative to P0 VICs. We found that cell cycle, DNAreplication, lipid metabolism, cell morphology, cellular assembly,and organization were among the top pathways to be activated(Fig. 3A and Fig. S3). Interestingly, the PI3K complex wasup-regulated as part of the gene network for cell morphology,cellular assembly, and organization (Fig. 3A) (21). Based onWestern blots, we confirmed that pAKT/AKT, which is down-stream of PI3K, was significantly up-regulated by TCPS culture,compared with P0 VICs, but not FAK or p38 MAPK (Fig. 3B).Although these pathways could be regulated by multiple varia-bles associated with the TCPS cultures (i.e., substrate stiffness,media components, cell density), we focused on the effects ofsubstrate stiffness on the PI3K pathway.
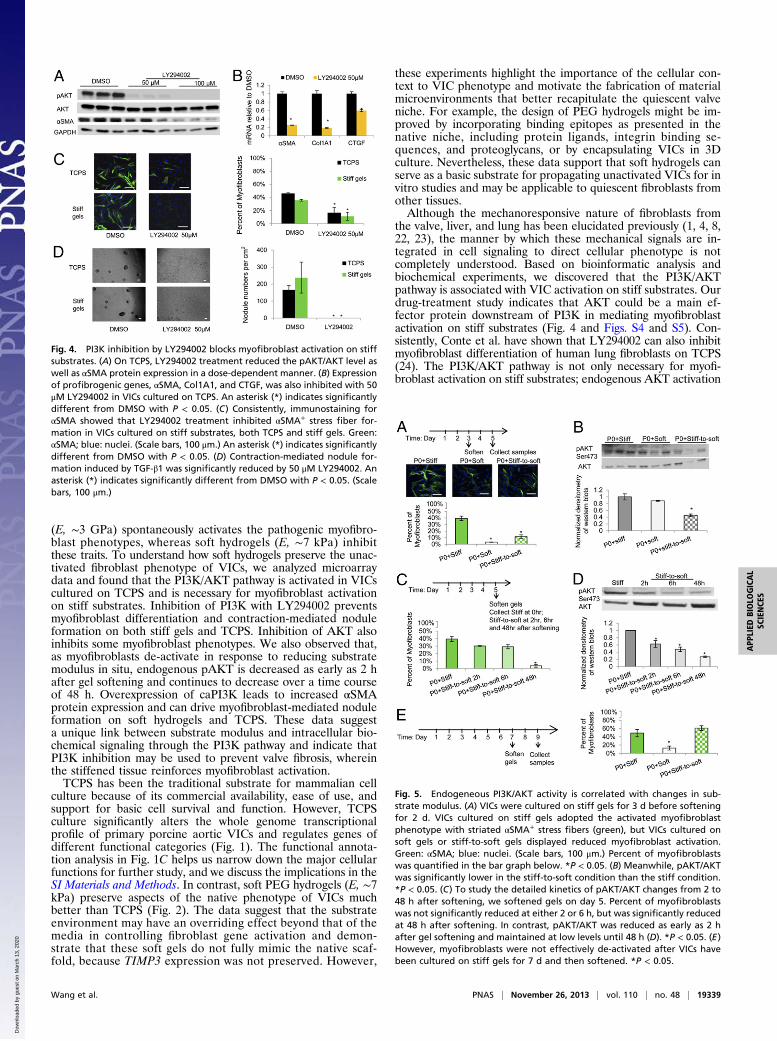
PI3K Inhibition Blocks Myofibroblast Activation. We set out to studywhether PI3K activation is necessary for myofibroblast activation
Fig. 1. Culture on TCPS changes the whole-genome tran-scriptional profile of VICs isolated from porcine aortic valves. (A)Hierarchical clustering based on microarray gene-expressiondata for freshly isolated VICs (P0 VICs) and VICs cultured on TCPS(TCPS VICs). (B) For this experiment, 2,173 gene probes weresignificantly up-regulated (≥twofold) and 1,926 gene probeswere significantly down-regulated (≥twofold) in TCPS VICscompared with P0 VICs, with P < 0.01. (C) DAVID functional an-notation showed that genes related to mitochondrion, cell cycle,and cytoskeleton were significantly up-regulated, whereasgenes related to extracellular region, regulation of transcriptionfrom RNA polymerase II promoter, inflammatory response, andpolysaccharide binding were down-regulated in TCPS VICs com-pared with P0 VICs. The P value of each gene ontology (GO)functional group follows the names of the GO terms. The per-centage in the pie charts refers to the fraction of genes that fallinto each functional category.
Wang et al. PNAS | November 26, 2013 | vol. 110 | no. 48 | 19337
APP
LIED
BIOLO
GICAL
SCIENCE
S
Dow
nloa
ded
by g
uest
on
Mar
ch 1
3, 2
020
on stiff substrates. P0 VICs were cultured on TCPS or stiff gelsfor 3 d and then treated with a small molecule inhibitor of PI3K,LY294002, for 2 d. The initial culture of P0 VICs on the sub-strates for 3 d was to allow the primary cells to recover and seedevenly before starting inhibition. As expected, LY294002 re-duced the ratio of pAKT/AKT (Fig. 4A). With PI3K inhibition,VICs demonstrated decreased myofibroblast activation on stiffsubstrates. First, αSMA protein levels were significantly reducedin response to increasing concentrations of LY294002 for VICscultured on TCPS (Fig. 4A). Second, at the mRNA level, anumber of myofibroblast activation genes, including αSMA, CTGF,and Col1A1, were significantly down-regulated with LY294002treatment on TCPS (Fig. 4B). Third, the percentage of myofi-broblasts quantified by αSMA+ stress fiber staining was signif-icantly reduced with PI3K inhibition on both TCPS and stiffgels (Fig. 4C). Specifically, the percentage of myofibroblasts wasreduced from 46.2 ± 0.9% to 16.9 ± 8.0% on TCPS and from35.6 ± 1.4% to 11.4 ± 6.2% on stiff gels with 50 μM LY294002.In addition, LY294002 completely blocked TGF-β1–inducednodule formation on both TCPS and stiff gels (Fig. 4D). Fur-thermore, wortmannin, a second PI3K inhibitor, inhibited myo-fibroblast differentiation on TCPS (Fig. S4), and MK2206, anAKT inhibitor, suppressed the myofibroblast phenotypes on stiffsubstrates (Fig. S5).
Endogeneous PI3K/AKT Activity Is Correlated with Changes inSubstrate Modulus. As PI3K activation is required for myofibro-blast activation on stiff substrates, we then tested whetherendogeneous PI3K/AKT activity is regulated by substrate mod-ulus. As shown in Fig. 5A, VICs cultured on stiff gels displayedthe characteristic myofibroblast phenotype with αSMA+ stressfibers (green). However, those cultured on soft or stiff-to-softgels (softened on day 3) had a significantly lower percentage ofmyofibroblasts (Fig. 5A). Interestingly, pAKT/AKT was signifi-cantly reduced on stiff-to-soft gels compared with stiff gels, butnot on soft gels (Fig. 5B). To better characterize the kineticchanges of myofibroblast deactivation and pAKT/AKT, stiff gelswere softened on day 5 and collected over 2, 6, or 48 h aftersoftening. The percentage of αSMA+ myofibroblasts was reduced,
but only significantly at 48 h after softening (Fig. 5C). In contrast,pAKT/AKT levels were decreased on stiff-to-soft gels as early as 2h after softening and maintained at a low level at 48 h, comparedwith that on stiff gels (Fig. 5D). Neither the pFAK/FAK nor thepp38 MAPK/p38 MAPK ratio changed significantly on materialswith different elasticities (Fig. S6). However, when VICs werecultured on stiff substrates for 7 d, they did not de-activate by 48 hafter softening the gels (Fig. 5E), indicating that the lineageplasticity of the myofibroblast-to-fibroblast transition may havea time or dose limit based on the culture conditions.
Constitutively Active PI3K Promotes Nodule Formation. We thenasked whether constitutively active PI3K (caPI3K) is sufficient todrive myofibroblast activation on soft substrates. When VICswere infected with caPI3K adenoviruses, these cells had higherpAKT/AKT levels and higher expression of αSMA than theGFP-infected cells (Fig. 6A). Furthermore, VICs infected withcaPI3K formed significantly more nodules on soft gels (Fig. 6B)and TCPS (Fig. S7) than those infected with GFP. These nodulesstained positive for αSMA, but quantifying stress fibers was dif-ficult when cells formed nodules. On both soft gels and TCPS, wetracked cellular movement after viral infection in real time andfound that nodules were induced by caPI3K through contractionof VICs (Movies S1–S4) and may eventually lead to matrix cal-cification (Fig. S7). In Fig. 6C, a contraction event that led tonodule formation (white arrow) was recorded for VICs infectedwith caPI3K on soft gels.
DiscussionStudies of the cellular microenvironment have broadened ourunderstanding of how various extracellular cues regulate cellfunction at the molecular level. In this study, we have begun toelucidate how the physical cue of matrix elasticity regulates thefibroblast-myofibroblast transition. We find that stiff TCPS
Fig. 2. Soft hydrogels preserve aspects of the native phenotype of VICsbetter than TCPS. (A) αSMA, CTGF, and Col1A1 expression levels, revealed byqRT-PCR, were significantly up-regulated when VICs were cultured on TCPS,but were preserved at levels similar to native cells when cultured on softhydrogels and in valve culture. TIMP3 was expressed at high levels in both P0VICs and valve culture, but was inhibited for VICs cultured on both soft gelsand TCPS. *Significantly different from P0 with P < 0.05. †Significantly dif-ferent from the soft gel condition with P < 0.05. (B) Based on Western blotanalysis, VICs cultured on soft gels, like P0 VICs, expressed significantly lessαSMA protein than those cultured on TCPS. (C) Representative staining ofαSMA for VICs residing in the native valve (P0), cultured on soft hydrogels oron TCPS. Green: αSMA; blue: nuclei. (Scale bars, 100 μm.)
Fig. 3. The PI3K pathway is activated during TCPS culture. Ingenuity path-way analysis was performed to construct gene networks based on the genesthat were significantly up-regulated by TCPS culture by ≥twofold with P <0.01. In A, a gene network associated with cell morphology, cellular as-sembly, and organization was detected with significant changes andrevealed up-regulation of the PI3K complex (blue highlights). The darker thered, the higher the fold change in gene expression. Values under each geneare the P value and the fold changes. (B) AKT, which is a main downstreamtarget of PI3K, was activated by the TCPS culture, compared with P0 VICs,but not FAK or p38 MAPK.
19338 | www.pnas.org/cgi/doi/10.1073/pnas.1306369110 Wang et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
3, 2
020
(E, ∼3 GPa) spontaneously activates the pathogenic myofibro-blast phenotypes, whereas soft hydrogels (E, ∼7 kPa) inhibitthese traits. To understand how soft hydrogels preserve the unac-tivated fibroblast phenotype of VICs, we analyzed microarraydata and found that the PI3K/AKT pathway is activated in VICscultured on TCPS and is necessary for myofibroblast activationon stiff substrates. Inhibition of PI3K with LY294002 preventsmyofibroblast differentiation and contraction-mediated noduleformation on both stiff gels and TCPS. Inhibition of AKT alsoinhibits some myofibroblast phenotypes. We also observed that,as myofibroblasts de-activate in response to reducing substratemodulus in situ, endogenous pAKT is decreased as early as 2 hafter gel softening and continues to decrease over a time courseof 48 h. Overexpression of caPI3K leads to increased αSMAprotein expression and can drive myofibroblast-mediated noduleformation on soft hydrogels and TCPS. These data suggesta unique link between substrate modulus and intracellular bio-chemical signaling through the PI3K pathway and indicate thatPI3K inhibition may be used to prevent valve fibrosis, whereinthe stiffened tissue reinforces myofibroblast activation.TCPS has been the traditional substrate for mammalian cell
culture because of its commercial availability, ease of use, andsupport for basic cell survival and function. However, TCPSculture significantly alters the whole genome transcriptionalprofile of primary porcine aortic VICs and regulates genes ofdifferent functional categories (Fig. 1). The functional annota-tion analysis in Fig. 1C helps us narrow down the major cellularfunctions for further study, and we discuss the implications in theSI Materials and Methods. In contrast, soft PEG hydrogels (E, ∼7kPa) preserve aspects of the native phenotype of VICs muchbetter than TCPS (Fig. 2). The data suggest that the substrateenvironment may have an overriding effect beyond that of themedia in controlling fibroblast gene activation and demon-strate that these soft gels do not fully mimic the native scaf-fold, because TIMP3 expression was not preserved. However,
these experiments highlight the importance of the cellular con-text to VIC phenotype and motivate the fabrication of materialmicroenvironments that better recapitulate the quiescent valveniche. For example, the design of PEG hydrogels might be im-proved by incorporating binding epitopes as presented in thenative niche, including protein ligands, integrin binding se-quences, and proteoglycans, or by encapsulating VICs in 3Dculture. Nevertheless, these data support that soft hydrogels canserve as a basic substrate for propagating unactivated VICs for invitro studies and may be applicable to quiescent fibroblasts fromother tissues.Although the mechanoresponsive nature of fibroblasts from
the valve, liver, and lung has been elucidated previously (1, 4, 8,22, 23), the manner by which these mechanical signals are in-tegrated in cell signaling to direct cellular phenotype is notcompletely understood. Based on bioinformatic analysis andbiochemical experiments, we discovered that the PI3K/AKTpathway is associated with VIC activation on stiff substrates. Ourdrug-treatment study indicates that AKT could be a main ef-fector protein downstream of PI3K in mediating myofibroblastactivation on stiff substrates (Fig. 4 and Figs. S4 and S5). Con-sistently, Conte et al. have shown that LY294002 can also inhibitmyofibroblast differentiation of human lung fibroblasts on TCPS(24). The PI3K/AKT pathway is not only necessary for myofi-broblast activation on stiff substrates; endogenous AKT activation
Fig. 4. PI3K inhibition by LY294002 blocks myofibroblast activation on stiffsubstrates. (A) On TCPS, LY294002 treatment reduced the pAKT/AKT level aswell as αSMA protein expression in a dose-dependent manner. (B) Expressionof profibrogenic genes, αSMA, Col1A1, and CTGF, was also inhibited with 50μM LY294002 in VICs cultured on TCPS. An asterisk (*) indicates significantlydifferent from DMSO with P < 0.05. (C) Consistently, immunostaining forαSMA showed that LY294002 treatment inhibited αSMA+ stress fiber for-mation in VICs cultured on stiff substrates, both TCPS and stiff gels. Green:αSMA; blue: nuclei. (Scale bars, 100 μm.) An asterisk (*) indicates significantlydifferent from DMSO with P < 0.05. (D) Contraction-mediated nodule for-mation induced by TGF-β1 was significantly reduced by 50 μM LY294002. Anasterisk (*) indicates significantly different from DMSO with P < 0.05. (Scalebars, 100 μm.)
Fig. 5. Endogeneous PI3K/AKT activity is correlated with changes in sub-strate modulus. (A) VICs were cultured on stiff gels for 3 d before softeningfor 2 d. VICs cultured on stiff gels adopted the activated myofibroblastphenotype with striated αSMA+ stress fibers (green), but VICs cultured onsoft gels or stiff-to-soft gels displayed reduced myofibroblast activation.Green: αSMA; blue: nuclei. (Scale bars, 100 μm.) Percent of myofibroblastswas quantified in the bar graph below. *P < 0.05. (B) Meanwhile, pAKT/AKTwas significantly lower in the stiff-to-soft condition than the stiff condition.*P < 0.05. (C) To study the detailed kinetics of pAKT/AKT changes from 2 to48 h after softening, we softened gels on day 5. Percent of myofibroblastswas not significantly reduced at either 2 or 6 h, but was significantly reducedat 48 h after softening. In contrast, pAKT/AKT was reduced as early as 2 hafter gel softening and maintained at low levels until 48 h (D). *P < 0.05. (E)However, myofibroblasts were not effectively de-activated after VICs havebeen cultured on stiff gels for 7 d and then softened. *P < 0.05.
Wang et al. PNAS | November 26, 2013 | vol. 110 | no. 48 | 19339
APP
LIED
BIOLO
GICAL
SCIENCE
S
Dow
nloa
ded
by g
uest
on
Mar
ch 1
3, 2
020

is correlated with substrate modulus reduction and cell pheno-types. The pAKT/AKT level is increased when P0 VICs are cul-tured on TCPS (Fig. 3B) and decreased as early as 2 h after gelsoftening, which happens before decreases in αSMA+ stress fibers(Fig. 5 C and D). The time-course experiments suggest a potentialdirect link between de-activation of AKT and reduction in sub-strate modulus. However, we did not observe a significant differ-ence in pAKT/AKT between stiff gels and static soft gels (Fig. 5B).We hypothesize that pAKT/AKT may be responding to dynamicelasticity changes in a time-dependent manner. It is possible that,as the cells have been cultured on static soft gels for 5 d, theyadapt to their matrix environment and show a sustained level ofpAKT/AKT. Because our photodegradable-PEG hydrogels havebeen extensively characterized to ensure that light-mediated deg-radation does not lead to significant changes in ligand density orsubstrate topography (9, 25–27), VICs should be primarily sensingthe reduction in substrate modulus.PI3K is a phospholipid kinase that specifically phosphorylates
the 3-OH group of the inositol ring and generates secondarymessengers, such as phosphatidylinositol (3-5)-trisphosphate(PIP3) that is bound to the inner plasma membrane (28).Downstream effector proteins then bind to PIP3 through thepleckstrin-homology (PH) domain and activate signaling eventsregulating actin polymerization, cell growth, cell cycle, and ap-optosis, among other functions (21). Interestingly, the PI3Kpathway has been shown to be mechano-sensitive in muscle tis-sues. Elevated AKT phosphorylation has been observed in ratskeletal muscles in response to exercise or passive stretch and inthe mouse diaphragm in response to tissue stretch and aging (29).In addition, cardiac-specific overexpression of active AKT hasbeen shown to enhance myocardial contractility (30). Our studyshows that PI3K in fibroblasts is mechano-responsive to elasticitychanges of culture substrates. When softening gels from ∼32 kPato ∼7 kPa, there was reduced PI3K activity measured by pAKT/AKT, indicating that PI3K can respond bidirectionally to me-chanical forces (i.e., in response to substrate modulus reduction orincreased mechanical load during muscle stretch and contraction).Interestingly, we also observed that overexpressing caPI3K can
overcome some effects of soft substrates and promote myofibro-blast phenotypes on soft gels (Fig. 6 A and B). It has been shownthat cell nodules form via contraction in an αSMA-dependentmanner when VICs are cultured on TCPS (31), and blebbistatin,which inhibits myosin II-mediated actomyosin contraction, signifi-cantly blocks myofibroblast differerntiation (Fig. S2). We observedsimilar patterns of nodule formation for VICs infected with caPI3Kadenoviruses on soft gels and TCPS (Movies S2 and S4). This
finding indicates that caPI3K may have increased αSMA-mediatedcontractility of VICs. Reif et al. have shown that activation of PI3Kpromotes Rac- and Rho-dependent stress fiber formation, focal-adhesion development and membrane ruffling in Swiss 3T3 cells(32). It is possible that PI3K may activate small GTPases or othermyosin-related proteins in these fibroblasts to increase their con-tractility and tension with their substrates. These data point toa potential role of the PI3K/AKT pathway in transmitting externalbiomechanical cues of elasticity to intracellular cytoskeleton re-modeling, gene regulation, and cell fate determination.One intriguing question is how VICs mechano-sense the ma-
trix elasticity changes and transmit that into PI3K/AKT signaling.Substrate modulus could be acting through multiple mechano-sensitive cell surface proteins and converge on the downstreamPI3K signaling (Fig. S8). For example, focal adhesions are themain connection points between cells and their matrix, and serveas sensors to various mechanical cues (33, 34). Various proteinsin the focal-adhesion machinery could affect PI3K activation,such as integrins (35) and focal-adhesion kinase (36). Further-more, a large screening study has shown that siRNA-mediatedknockdown of 85 human protein tyrosine kinases induced dis-tinct alterations in the focal-adhesion morphology on differentstiffnesses (37), and tyrosine phosphorylation levels are linkedto mechanically induced changes controlling different cellularfunctions (38). As protein tyrosine kinases are common up-stream receptors which activate PI3K (39), it is possible thattheir regulation, which is linked with focal-adhesion formation,could affect PI3K/AKT phosphorylation. Another importantgroup of mechano-senstive proteins are mechano-sensitive ionchannels. Our preliminary data support that they may be involvedin the elasticity-regulated myofibroblast activation, as treatmentwith their inhibitor reduced VIC myofibroblast differentiation onTCPS (Fig. S8). For example, the TRPC family cation channelscan be induced by plasma membrane deformation (40) and lead toCa2+ influx into the cells, which can then affect a number of sig-naling events intracellularly, including PI3K (41). This resultsupports some basic hypotheses about potential upstream mechano-sensing events and provides a future candidate to explore as thefield seeks to better understand the continuum of mechanicalsignaling from the ECM to the nucleus, especially related to theupstream mechano-sensing machinery in VICs.During normal wound healing of mesenchymal tissues, the
fibroblast-to-myofibroblast transition is an essential process, be-cause myofibroblasts contract and secrete ECM proteins to repairthe tissue. Toward the end of healing, apoptosis of myofibroblastshas been observed to put a brake on active tissue remodeling (42,43). When activated cells fail to undergo apoptosis, chronic fibrosiscan occur. Therefore, temporal regulation of myofibroblast apo-ptosis and survival is crucial in determining healthy and diseasedtissue regeneration. PI3K/AKT signaling antagonizes apoptosisand promotes cell survival and proliferation. This process islargely mediated by the effector protein AKT, which inhibitsapoptosis by directly phosphorylating BAD and Forkhead tran-scription factors (44). Two independent reports have shown thatAKT activation confers an apoptosis-resistant phenotype to myo-fibroblasts (45, 46). In fibrotic tissues, elevated AKT activity inmyofibroblasts, induced either by stiffened matrix or proinflam-matory cytokines, could be an important cause of persistentactivation. Therefore, targeting the PI3K/AKT pathway may accel-erate the turnover of myofibroblasts through apoptosis.Unlike a decade ago, tissue engineering and regenerative
medicine have not been restricted to the laboratory, but havebeen implemented in the clinic (47). It is realized that the scaf-fold does not need to completely recapitulate the native tissuestructures and functionalities before implantation, but can beinstructive because cells and scaffolds are plastic and mutuallyresponsive. However, important properties of these cell-culturescaffolds can be informed by basic cell biology research. Forexample, the elasticity of the materials can control key cell sig-naling events and it may be important for the elasticity to matchthe changing environment in vivo (48, 49). Over the years, PEG
Fig. 6. Constitutively active PI3K promotes contraction-mediated noduleformation by VICs on soft substrates. (A) caPI3K increased phosphorylationof AKT, one of the main downstream effectors of PI3K. Similarly, the αSMAprotein level was increased by caPI3K overexpression. (B) Four days afterviral infection, increased nodule formation was observed for VICs treatedwith caPI3K but cultured on soft gels. Cell nodules induced by caPI3K stainedpositive for αSMA. *P < 0.05. (Scale bars, 100 μm.) (C) Real-time tracking ofnodule formation in caPI3K-infected VICs revealed that nodule formationwas mediated by cell contraction. White arrows highlight a contractionevent (Movie S2).
19340 | www.pnas.org/cgi/doi/10.1073/pnas.1306369110 Wang et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
3, 2
020

hydrogels have been studied to incorporate different functionaland dynamic modules to serve as a better surrogate scaffold forcell culture and tissue regeneration (50). Even though one canchange intracellular signaling by chemical cues and transcriptionfactors easily, it is still necessary to learn how matrix mechanicalcues can be used to regulate cellular functions to understandbasic cell biology and for clinical applications. This outside-inprogramming may reveal unique manners to intervene duringdisease progression.
Materials and MethodsPrimary VICs were harvested from porcine aortic valves based on a sequentialcollagenase digestion. These cells were cultured either on TCPS or PEGhydrogels, which were manufactured as previously published (8). Hydrogelswere made with two different moduli, ∼7 kPa (soft) and ∼32 kPa (stiff), or
degraded in situ from ∼32 kPa to ∼7 kPa (stiff-to-soft) over time. Cells cul-tured on different substrates were then fixed for immunocytochemistryanalysis or lysed to collect RNA or protein for gene expression quantificationbased on real-time PCR and microarrays or protein analysis based on West-ern blot. Porcine genome microarrays (Affymetrix) were performed basedon standard hybridization methods and analyzed using multiple bio-informatics tools. Experimental data were based on at least three biologicalrepeats and analyzed using a one-way ANOVA statistical test. Please refer toSI Materials and Methods for detailed methods.
ACKNOWLEDGMENTS. We thank Dr. Daniel Alge, Chun Yang, and Dr. MalarAzagarsamy for help with peptide synthesis; Dr. Jennifer Leight for help inobtaining adenoviruses; Dr. Kristen Barthel for editing the manuscript; andDr. Nicholas Farina and Dr. Bradley Olwin for scientific discussions regardingmicroarray analysis. This study was supported in part by National Institutesof Health Grant R01 HL089260 and the Howard Hughes Medical Institute.
1. Liu F, et al. (2010) Feedback amplification of fibrosis through matrix stiffening andCOX-2 suppression. J Cell Biol 190(4):693–706.
2. Hinton RB, Yutzey KE (2011) Heart valve structure and function in development anddisease. Annu Rev Physiol 73:29–46.
3. Liu AC, Joag VR, Gotlieb AI (2007) The emerging role of valve interstitial cell phe-notypes in regulating heart valve pathobiology. Am J Pathol 171(5):1407–1418.
4. Chen J-H, Chen WLK, Sider KL, Yip CYY, Simmons CA (2011) β-catenin mediates me-chanically regulated, transforming growth factor-β1-induced myofibroblast differ-entiation of aortic valve interstitial cells. Arterioscler Thromb Vasc Biol 31(3):590–597.
5. Mohler ER, 3rd, et al. (2001) Bone formation and inflammation in cardiac valves.Circulation 103(11):1522–1528.
6. Engler AJ, Sen S, Sweeney HL, Discher DE (2006) Matrix elasticity directs stem celllineage specification. Cell 126(4):677–689.
7. Rajamannan NM, et al. (2011) Calcific aortic valve disease: Not simply a degenerativeprocess: A review and agenda for research from the National Heart and Lung andBlood Institute Aortic Stenosis Working Group. Executive summary: Calcific aorticvalve disease-2011 update. Circulation 124(16):1783–1791.
8. Wang H, Haeger SM, Kloxin AM, Leinwand LA, Anseth KS (2012) Redirecting valvularmyofibroblasts into dormant fibroblasts through light-mediated reduction in sub-strate modulus. PLoS ONE 7(7):e39969.
9. Kloxin AM, Benton JA, Anseth KS (2010) In situ elasticity modulation with dynamicsubstrates to direct cell phenotype. Biomaterials 31(1):1–8.
10. Kloxin AM, Tibbitt MW, Kasko AM, Fairbairn JA, Anseth KS (2010) Tunable hydrogelsfor external manipulation of cellular microenvironments through controlled photo-degradation. Adv Mater 22(1):61–66.
11. Dupont S, et al. (2011) Role of YAP/TAZ in mechanotransduction. Nature 474(7350):179–183.
12. Engler AJ, et al. (2004) Myotubes differentiate optimally on substrates with tissue-likestiffness: Pathological implications for soft or stiff microenvironments. J Cell Biol166(6):877–887.
13. Trappmann B, et al. (2012) Extracellular-matrix tethering regulates stem-cell fate. NatMater 11(7):642–649.
14. Khetan S, et al. (2013) Degradation-mediated cellular traction directs stem cell fate incovalently crosslinked three-dimensional hydrogels. Nat Mater 12(5):458–465.
15. Katsumi A, Orr AW, Tzima E, Schwartz MA (2004) Integrins in mechanotransduction.J Biol Chem 279(13):12001–12004.
16. Huang X, et al. (2012) Matrix stiffness-induced myofibroblast differentiation is me-diated by intrinsic mechanotransduction. Am J Respir Cell Mol Biol 47(3):340–348.
17. Hinz B, Celetta G, Tomasek JJ, Gabbiani G, Chaponnier C (2001) Alpha-smooth muscleactin expression upregulates fibroblast contractile activity. Mol Biol Cell 12(9):2730–2741.
18. Ponticos M, et al. (2009) Pivotal role of connective tissue growth factor in lung fi-brosis: MAPK-dependent transcriptional activation of type I collagen. Arthritis Rheum60(7):2142–2155.
19. Gressner OA, Gressner AM (2008) Connective tissue growth factor: A fibrogenicmaster switch in fibrotic liver diseases. Liver Int 28(8):1065–1079.
20. Hinz B, et al. (2007) The myofibroblast: One function, multiple origins. Am J Pathol170(6):1807–1816.
21. Vanhaesebroeck B, Stephens L, Hawkins P (2012) PI3K signalling: The path to dis-covery and understanding. Nat Rev Mol Cell Biol 13(3):195–203.
22. Olsen AL, et al. (2011) Hepatic stellate cells require a stiff environment for myofi-broblastic differentiation. Am J Physiol Gastrointest Liver Physiol 301(1):G110–G118.
23. Wipff P-J, Rifkin DB, Meister J-J, Hinz B (2007) Myofibroblast contraction activateslatent TGF-beta1 from the extracellular matrix. J Cell Biol 179(6):1311–1323.
24. Conte E, et al. (2011) Inhibition of PI3K prevents the proliferation and differentiationof human lung fibroblasts into myofibroblasts: The role of class I P110 isoforms. PLoSONE 6(10):e24663.
25. Tibbitt MW, Kloxin AM, Anseth KS (2013) Modeling controlled photodegradation inoptically thick hydrogels. J Polym Sci A Polym Chem 51(9):1899–1911.
26. Tibbitt MW, Kloxin AM, Sawicki LA, Anseth KS (2013) Mechanical properties anddegradation of chain and step-polymerized photodegradable hydrogels. Macro-molecules 46(7):2785–2792.
27. Kloxin AM, Kloxin CJ, Bowman CN, Anseth KS (2010) Mechanical properties of cell-ularly responsive hydrogels and their experimental determination. Adv Mater 22(31):3484–3494.
28. Cantley LC (2002) The phosphoinositide 3-kinase pathway. Science 296(5573):1655–1657.
29. Pardo PS, Lopez MA, Boriek AM (2008) FOXO transcription factors are mechano-sensitive and their regulation is altered with aging in the respiratory pump. Am JPhysiol Cell Physiol 294(4):C1056–C1066.
30. Condorelli G, et al. (2002) Akt induces enhanced myocardial contractility and cell sizein vivo in transgenic mice. Proc Natl Acad Sci USA 99(19):12333–12338.
31. Benton JA, Kern HB, Leinwand LA, Mariner PD, Anseth KS (2009) Statins block calcificnodule formation of valvular interstitial cells by inhibiting alpha-smooth muscle actinexpression. Arterioscler Thromb Vasc Biol 29(11):1950–1957.
32. Reif K, Nobes CD, Thomas G, Hall A, Cantrell DA (1996) Phosphatidylinositol 3-kinasesignals activate a selective subset of Rac/Rho-dependent effector pathways. Curr Biol6(11):1445–1455.
33. Hayakawa K, Tatsumi H, Sokabe M (2012) Mechano-sensing by actin filaments andfocal adhesion proteins. Commun Integr Biol 5(6):572–577.
34. Bershadsky AD, Balaban NQ, Geiger B (2003) Adhesion-dependent cell mechano-sensitivity. Annu Rev Cell Dev Biol 19(1):677–695.
35. Watabe H, Furuhama T, Tani-Ishii N, Mikuni-Takagaki Y (2011) Mechanotransductionactivates α₅β₁ integrin and PI3K/Akt signaling pathways in mandibular osteoblasts.Exp Cell Res 317(18):2642–2649.
36. Xia H, Nho RS, Kahm J, Kleidon J, Henke CA (2004) Focal adhesion kinase is upstreamof phosphatidylinositol 3-kinase/Akt in regulating fibroblast survival in response tocontraction of type I collagen matrices via a β 1 integrin viability signaling pathway.J Biol Chem 279(31):33024–33034.
37. Prager-Khoutorsky M, et al. (2011) Fibroblast polarization is a matrix-rigidity-dependentprocess controlled by focal adhesion mechanosensing. Nat Cell Biol 13(12):1457–1465.
38. Giannone G, Sheetz MP (2006) Substrate rigidity and force define form through ty-rosine phosphatase and kinase pathways. Trends Cell Biol 16(4):213–223.
39. Schlessinger J (2000) Cell signaling by receptor tyrosine kinases. Cell 103(2):211–225.40. Spassova MA, Hewavitharana T, Xu W, Soboloff J, Gill DL (2006) A common mecha-
nism underlies stretch activation and receptor activation of TRPC6 channels. Proc NatlAcad Sci USA 103(44):16586–16591.
41. Zanou N, et al. (2012) Trpc1 ion channel modulates phosphatidylinositol 3-kinase/Aktpathway during myoblast differentiation and muscle regeneration. J Biol Chem287(18):14524–14534.
42. Uhal BD (2008) The role of apoptosis in pulmonary fibrosis. European RespiratoryReview 17(109):138–144.
43. Desmoulière A, Redard M, Darby I, Gabbiani G (1995) Apoptosis mediates the de-crease in cellularity during the transition between granulation tissue and scar. Am JPathol 146(1):56–66.
44. Biggs WH, 3rd, Meisenhelder J, Hunter T, Cavenee WK, Arden KC (1999) Protein ki-nase B/Akt-mediated phosphorylation promotes nuclear exclusion of the wingedhelix transcription factor FKHR1. Proc Natl Acad Sci USA 96(13):7421–7426.
45. Horowitz JC, et al. (2007) Combinatorial activation of FAK and AKT by transforminggrowth factor-beta1 confers an anoikis-resistant phenotype to myofibroblasts. CellSignal 19(4):761–771.
46. Kulasekaran P, et al. (2009) Endothelin-1 and transforming growth factor-beta1 in-dependently induce fibroblast resistance to apoptosis via AKT activation. Am J RespirCell Mol Biol 41(4):484–493.
47. Bader A, Macchiarini P (2010) Moving towards in situ tracheal regeneration: the bi-onic tissue engineered transplantation approach. J Cell Mol Med 14(7):1877–1889.
48. Young JL, Engler AJ (2011) Hydrogels with time-dependent material properties en-hance cardiomyocyte differentiation in vitro. Biomaterials 32(4):1002–1009.
49. Kloxin AM, Kasko AM, Salinas CN, Anseth KS (2009) Photodegradable hydrogels fordynamic tuning of physical and chemical properties. Science 324(5923):59–63.
50. Peppas NA, Hilt JZ, Khademhosseini A, Langer R (2006) Hydrogels in biology andmedicine: From molecular principles to bionanotechnology. Adv Mater 18(11):1345–1360.
Wang et al. PNAS | November 26, 2013 | vol. 110 | no. 48 | 19341
APP
LIED
BIOLO
GICAL
SCIENCE
S
Dow
nloa
ded
by g
uest
on
Mar
ch 1
3, 2
020