Embed Size (px)
Citation preview

Biochimica et Biophysics Acta, 1124 (1992) 199-202 0 1992 Elsevier Science Publishers B.V. All rights reserved OOOS2760/92/$05.00
BBALIP 50336 Rapid Report
199
Identification of pristanoyl-CoA oxidase as a distinct, clofibrate non-inducible enzyme in rat liver peroxisomes
R.J.A. Wanders ‘, S. Denis ‘, C. Jakobs 2 and H.J. ten Brink 2
’ Department of Clinical Biochemistry, University Hospital Amsterdam, Amsterdam (The Netherlands) and ’ Department of Pediatrics, Free Unicersity Amsterdam, Amsterdam (The Netherlands)
(Received 20 December 1991)
Key words: Peroxisome; Fatty acid; Acyl-CoA oxidase; Zellweger syndrome
In this paper we describe the identification of pristanoyl-CoA oxidase activity in rat liver peroxisomes. This activity was not stimulated by clofibrate feeding. Furthermore, the activity was found in multiple tissues. These results show that pristanoyl-CoA oxidase is different from any of the known oxidases which include a clofibrate-inducible acyl-CoA oxidase and the recently identified cholestanoyl-Cob oxidase. Gelfiltration and chromatofocusing experi- ments provide conclusive evidence that we are dealing with a novel acyl-CoA oxidase with a unique function in peroxisomal p-oxidation.
In recent years there has been a renewed interest in the degradation of phytanic and pristanic acid, espe- cially since these two fatty acids have been found to accumulate in plasma from patients affected by one of a variety of peroxisomal disorders ([l-31; for reviews see Refs. 4 and 5). Studies in the early 1960s have shown that pristanic acid (2,6,10,14-tetramethylpenta- decanoic acid) is formed from phytanic acid (3,7,11,15- tetramethylhexadecanoic acid) via an c-u-oxidation pro- cess [7], although the individual steps of the pathway remain to be elucidated [81. Apart from endogenous synthesis from phytanic acid, pristanic acid is also derived from dietary sources such as butterfat [9]. Studies by Steinberg and co-workers [7] have shown that pristanic acid can be P-oxidized efficiently, alter- nately giving rise to acetyl-CoA and propionyl-CoA. Recent studies suggest that this occurs in peroxisomes rather than in mitochondria. Indeed, Singh et al. [lo] reported that pristanic acid &oxidation is deficient in fibroblasts from Zellweger patients which lack peroxi- somes. Furthermore, we have identified pristanoyl-CoA oxidase activity in human liver peroxisomes [ll]. We now report the identification of pristanoyl-CoA oxidase as a distinct, clofibrate non-inducible oxidase in rat liver peroxisomes. Different tissues from male Wistar rats (200-250 g> fed a standard laboratory diet plus or minus 0.5% (w/v) clofibrate (7-10 days), were finely
Correspondence: R.J.A. Wanders, Dept. Clinical Biochemistry, Uni-
versity Hospital Amsterdam, Meibergdreef 9, 1105 AZ, Amsterdam, The Netherlands.
minced in a medium containing 250 mM sucrose, 2 mM EDTA and 2 mM Mops-NaOH (final pH 7.4). After several washings the minced pieces were stored dry at - 80°C or used immediately. On the day of the experi- ment the minced pieces were thawed in buffer A containing 10 mM sodium pyrophosphate, 0.1% (w/v> Triton X-100, 1 mM EDTA and 0.01 mM FAD (final pH 9.0), subjected to sonication (three cycles of 15s at 80 W with time intervals of 45 s) at 4°C and used for enzyme assays.
Differential and density gradient centrifugation Rat livers were finely minced and homogenized in a
medium containing 250 mM sucrose, 2 mM EDTA and 2 mM Mops-NaOH (final pH 7.4). The resulting ho- mogenate was subjected to differential centrifugation to prepare a light mitochondrial fraction (see Ref. 12 for centrifugation conditions). This fraction was subse- quently subjected to equilibrium density gradient cen- trifugation in a linear Nycodenz gradient as described before [131. After unloading of the gradient enzyme activities were measured in each of the fractions (see below).
Enzyme activity measurements The activity of pristanoyl-CoA oxidase and palmi-
toyl-CoA oxidase was measured in a medium contain- ing 50 mM Mops-NaOH (pH 7.61, 1 mM homovanillic acid, 20 U/ml horseradish peroxidase, 0.1 mM NaN,, 0.01 mM FAD, 0.02% (w/v) Triton X-100, and 0.1 mM pristanoyl-CoA or palmitoyl-CoA. In the latter case 0.6 mg/ml bovine serum albumin (fatty acid free) was
200
included as well. Fluorescence was followed in time using a Cobas-Bio Centrifugal Analyzer (excitation wavelength: 3’27 nm; emission filter: 410-490 nm), Hoffman-La Roche (Base& Switzerland). The activities of glutamate dehydrogenase, p-amino acid oxidase and esterase were measured as described before [12].
Chromatofocusing and gelfiltration experiments A light mitochondrial fraction enriched in peroxi-
somes was prepared from rat liver as described above. The final pellet was taken up in buffer A containing 0.1% (w/v) Triton X-100 followed by sonication (three cycles of 15 s with 45 s intervals, 4°C). An aliquot of the resulting supernatant (0.5 ml, 3 mg protein) was ap- plied to a PBE 94 gel (Pharmacia, Uppsala, Sweden) which was equilibrated with 25 mM ethanolamine/ acetic acid (pH 9.51, containing 0.1% (w/v> Triton X-100. The column was subsequently eluted with 25 mM Polybuffer 96HCl (pH 7.0) containing 0.1% (w/v> Triton X-100. Fractions of 1 ml were collected fol- lowed by analysis of pH and palmitoyl-CoA and pris- tanoyl-CoA oxidase activity.
In the experiment of Fig. 2B an aliquot of the Triton X-100 extract of a peroxisome-enriched fraction (see above) was applied to gelfiltration using a XK 16/70 (Pharmacia, Uppsala, Sweden) column (d 16 mm> con- taining Sephacryl S-300 HR. The column was eluted with buffer A containing 10 mM sodium pyrophos- phate, 1 mM EDTA, 0.01 mM FAD and 0.1% (w/v) Triton X-100 (pH 9.0). The column was calibrated
using a gelfiltration calibration kit (Pharmacia, Upp- sala, Sweden). ~omovaniIlic acid, palmitoyl~CoA, bovine serum albumin (fatty acid free) and nucleotides were obtained from Sigma (St. Louis, MO, USA). Horseradish peroxidase (grade II> was from Boehringer (Mannheim, Germany). Pristanic acid was synthesized as described before [ll]. Pristanoyl-CoA was synthe- sized enzymatically using a partially purified acyl-CoA synthetase preparation [I41 followed by purification on HPLC. Its identity was checked by gas chromatog- raphy/mass spectrometry. All other chemicals were of the purest grade available.
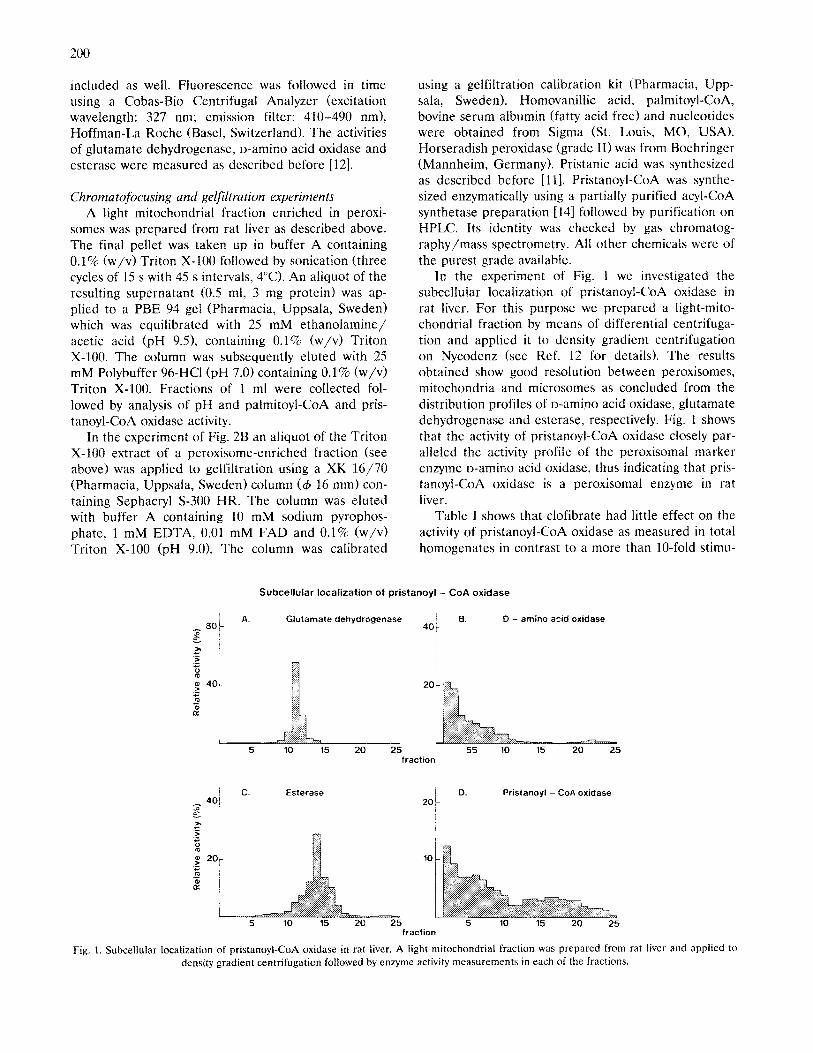
In the experiment of Fig. 1 we investigated the subcellular localization of pristanoyl-CoA oxidase in rat liver. For this purpose we prepared a light-mito- chondrial fraction by means of differential centrifuga- tion and applied it to density gradient centrifugation on Nycodenz (see Ref. 12 for details). The results obtained show good resolution between peroxisomes, mitochondria and microsomes as concluded from the distribution profiles of p-amino acid oxidase, glutamate dehydrogenase and esterase, respectively. Fig. 1 shows that the activity of pristanoyl-CoA oxidase closely par- alleled the activity profile of the peroxisoma1 marker enzyme o-amino acid oxidase, thus indicating that pris- tanoyl-CoA oxidase is a peroxisomal enzyme in rat liver.
Table I shows that clofibrate had little effect on the activity of pristanoyl-CoA oxidase as measured in total homogenates in contrast to a more than IO-fold stimu-
Subcellular localization of pristanoyl - CoA oxidase
;; 8Ot A. Glutamate dehydrogenase
40t 13. D - amino acid oxidase
a- i I
fraction
’ c. Esterase
2 401
2
0.
t Pristaooyl - CoA cxidase
20
5 10 15 20 25 fraction
5 10 15 20 25
Fig. 1. Subcellular localization of pristanoyl-CoA oxidase in rat liver. A light mitochondrial fraction was prepared from rat liver and applied to
density gradient centrifugation followed by enzyme activity measurements in each of the fractions.
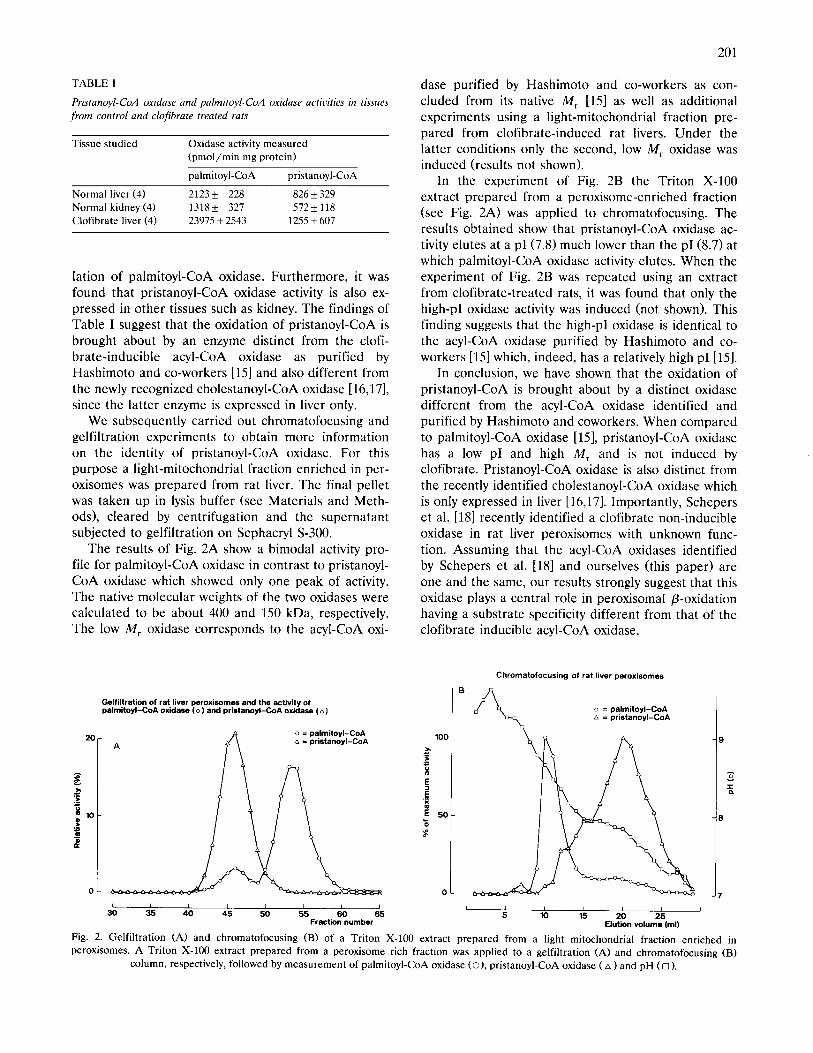
TABLE I
Pristanoyl-CoA oxidase and palmitoyl-CoA oxidase activities in tissues from control and clofibrate treated rats
Tissue studied Oxidase activity measured
(pmol/min mg protein)
Normal liver (4)
Normal kidney (4)
Clofibrate liver (4)
palmitoyl-CoA pristanoyl-CoA
2123+ 228 826? 329
1318+ 327 572k 118
23975+2543 1255k607
lation of palmitoyl-CoA oxidase. Furthermore, it was found that pristanoyl-CoA oxidase activity is also ex- pressed in other tissues such as kidney. The findings of Table I suggest that the oxidation of pristanoyl-CoA is brought about by an enzyme distinct from the clofi- brate-inducible acyl-CoA oxidase as purified by Hashimoto and co-workers [15] and also different from the newly recognized cholestanoyl-CoA oxidase [16,17], since the latter enzyme is expressed in liver only.
We subsequently carried out chromatofocusing and gelfiltration experiments to obtain more information on the identity of pristanoyl-CoA oxidase. For this purpose a light-mitochondrial fraction enriched in per- oxisomes was prepared from rat liver. The final pellet was taken up in lysis buffer (see Materials and Meth- ods), cleared by centrifugation and the supernatant subjected to gelfiltration on Sephacryl S-300.
The results of Fig. 2A show a bimodal activity pro- file for palmitoyl-CoA oxidase in contrast to pristanoyl- CoA oxidase which showed only one peak of activity. The native molecular weights of the two oxidases were calculated to be about 400 and 150 kDa, respectively. The low M, oxidase corresponds to the acyl-CoA oxi-
Gelfiltration of rat liver peroxisomes and the activity of palmitoyl-CoA oxidase (0) and pristanoyl-CoA oxidaaa (A)
zo- o = palmitoyl-CoA
A D = pristanoyl-CoA
0 I I I I I I I J
30 35 40 45 50 55 50 65 Fraction number
201
dase purified by Hashimoto and co-workers as con- cluded from its native M, 1151 as well as additional experiments using a light-mitochondrial fraction pre- pared from clofibrate-induced rat livers. Under the latter conditions only the second, low M, oxidase was induced (results not shown).
In the experiment of Fig. 2B the Triton X-100 extract prepared from a peroxisome-enriched fraction (see Fig. 2A) was applied to chromatofocusing. The results obtained show that pristanoyl-CoA oxidase ac- tivity elutes at a pI (7.8) much lower than the pI(8.7) at which palmitoyl-CoA oxidase activity elutes. When the experiment of Fig. 2B was repeated using an extract from clofibrate-treated rats, it was found that only the high-p1 oxidase activity was induced (not shown). This finding suggests that the high-p] oxidase is identical to the acyl-CoA oxidase purified by Hashimoto and co- workers [15] which, indeed, has a relatively high p1 [153.
In conclusion, we have shown that the oxidation of pristanoyl-CoA is brought about by a distinct oxidase different from the acyl-CoA oxidase identified and purified by Hashimoto and coworkers. When compared to palmitoyl-CoA oxidase [15], pristanoyl-CoA oxidase has a low pI and high M, and is not induced by clofibrate. Pristanoyl-CoA oxidase is also distinct from the recently identified cholestanoyl-CoA oxidase which is only expressed in liver [16,171. Importantly, Schepers et al. [18] recently identified a clofibrate non-inducible oxidase in rat liver peroxisomes with unknown func- tion. Assuming that the acyl-CoA oxidases identified by Schepers et al. [18] and ourselves (this paper) are one and the same, our results strongly suggest that this oxidase plays a central role in peroxisomal P-oxidation having a substrate specificity different from that of the clofibrate inducible acyl-CoA oxidase.
Chromatofocusing of rat liver peroxisomes
o = palmitoyl-CoA a = pristanoyl-CoA
I I I 1 I 4
5 lo 15 20 25 Elution volume (ml)
3
B Ip
3
r
Fig. 2. Gelfiltration (A) and chromatofocusing (B) of a Triton X-100 extract prepared from a light mitochondrial fraction enriched in
peroxisomes. A Triton X-100 extract prepared from a peroxisome rich fraction was applied to a gelfiltration (A) and chromatofocusing (B)
column, respectively, followed by measurement of palmitoyl-CoA oxidase CO), pristanoyl-CoA oxidase (A ) and pH (0 ).

202
Acknowledgements
Mrs. B. Groendijk is gratefully acknowledged for expert preparation of the manuscript and The Princess Beatrix Fund (The Hague, The Netherlands) for finan- cial support.
References
Klenk, E. and Kahlke, W. (1963) Hoppe-Seyler’s Z. Physiol.
Chem. 333, 133-138.
Poulos, A., Sharp, P., Fellenberg, A.J. and Johnson, D.W. (1988)
Eur. J. Pediatr. 147, 143-147.
Ten Brink, H.J., Wanders, R.J.A., Stellaard, F., Schutgens, R.B.H.
and Jakobs, C. (1991) J. Inher. Metab. Dis. 14, 345-348.
Wanders, R.J.A., Heymans, H.S.A., Schutgens, R.B.H., Barth,
P.G., Van den Bosch, H. and Tager, J.M. (1988) J. Neurol. Sci.
88, l-39.
Steinberg, D. (1989) in The Metabolic Basis of Inherited Disease
(Striver, C.R., Beaudet, A.L., Sly, W.S. and Valle, D., eds.), pp.
1533-1550, McGraw-Hill, New York.
Lazarow, P.B. and Moser, H.W. (1989) in The Metabolic Basis of
Inherited Disease (Striver, CR., Beaudet, A.L., Sly, W.S. and
Valle, D., eds), pp. 1501-1532, McGraw-Hill, New York.
12
13
14
15
16
17
18
Tsai, S.C., Avigan, J. and Steinberg, D. (1969) J. Biol. Chem. 244,
2682-2692.
Skjeldal, O.H. and Stokke, 0. (1988) Scan. J. Clin. Invest. 48,
97-102.
Hanssen, R.P. and Morrison, J.D. (1964) Biochem. J. 93,225-229.
Singh, H., Usher, S., Johnson, D. and Poulos, A. (19901 J. Lipid
Res. 31, 217-225.
Wanders, R.J.A., Ten Brink, H.J., Van Roermund, C.W.T.,
Schutgens, R.B.H., Tager, J.M. and Jakobs, C. (1990) Biochem.
Biophys. Res. Commun. 172, 490-495.
Wanders, R.J.A., Romeijn, G.J., Schutgens, R.B.H. and Tager,
J.M. (1989) Biochem. Biophys. Res. Commun. 164, 550-555.
Wanders, R.J.A., Van Roermund, C.W.T., Van Wijland, M.J.A.,
Heikoop, J., Schutgens, R.B.H., Schram, A.W. and Tager, J.M.
(1987) J. Clin. Invest. 80, 1778-1783.
Tanaka, T., Hosaka, K., Hoshimara, M. and Numa, S. (19791 Eur.
J. Biochem. 98, 165-172.
Osumi, T., Hashimoto, T. and Ui, N. (1980) J. Biochem. 87,
1735-1746.
Casteels, M., Schepers, L., Van Eldere, J., Eyssen, H. and Man-
naerts, G.P. (1988) J. Biol. Chem. 263, 4654-4661.
Pedersen, J.I., Hvattum, E., Flatabo, T. and Bjorkhem, I. (1988)
Biochem. Int. 17, 163-169.
Schepers, L., Van Veldhoven, P.P., Casteels, M., Eyssen, H.J.
and Mannaerts, G.P. (1990) J. Biol. Chem. 265, 5242-5246.





























