Embed Size (px)
Citation preview

THE JOURNAL OF BPX.OWXL CHEMISTRY Vol. 253, No. 11, Issue of June 10, pp. 4016-4021, 1978
Printed in U.S.A.
Immunochemical Studies on Cholecystokinin I. DEVELOPMENT OF SEQUENCE-SPECIFIC RADIOIMMUNOASSAYS FOR PORCINE
TRIACONTATRIAPEPTIDE CHOLECYSTOKININ*
(Received for publication, September 13, 1977)
JENS F. REHFELD
From the Institute of Medical Biochemistry, University of Aarhus, DK-8000 Aarhus C, Denmark
Highly sensitive, sequence-specific radioimmunoassays for the triacontatriapeptide hormone cholecystokinin (CCK,,) have been developed. CCK,, was labeled through one of its lysyl side chains with ‘2”I-hydroxyphenylpropionic acid-succinimide ester. The immunoreactivity of this tracer was indistinguishable from that of native CCK,, and stable for more than 3 months. High specific radioactivities of 350 to 600 $X/nmol were obtained by separation of labeled from unlabeled CCK by gel chromatography. Conventional iodi- nation techniques using chloramine-T lactoperoxidase were inapplicable, since even mild oxidation reduced the immu- noreactivity of CCK by 70 to 100%. Antisera were raised toward partially purified porcine CCK,, either free (injected into 30 guinea pigs) or coupled to bovine serum albumin (injected into 10 rabbits). All animals produced antisera in dilutions ranging from 500 to 50,000. The binding affinity ranged from lo8 to lOI* M-’ as expressed by the “effective” equilibrium constant, K,,. Accordingly, the most sensitive assays could detect less than 0.2 fmol of CCK,,. With respect to specificity, three sorts of antisera with high binding affinities (i.e. K eff 2 10”’ M-I) were produced. (a) Antisera from CCK-immunized guinea pigs (4478, 4479, and 4489) reacted with the midportion of CCK,,, presumably the sequence of amino acids from 20 to 25. (b) Antisera from CCK-immunized rabbit 4698 and from guinea pigs 4465 and 4488 reacted with the sequence 25 to 30. These antisera bound CCK,, and its COOH-terminal octapeptide with equi- molar potency, whereas they bound the triacontanonapep- tide (CCK,,,) with a potency varying from 15 to 60%. They did not react with the COOH-terminal tetrapeptide sequence common to gastrins and CCK. Neither did these antisera bind the nonsulfated COOH-terminal octapeptide analogue. (c) Three antisera from gastrin-immunized rabbits, 2609, 2716, and 2717, cross-reacted with CCK through the common COOH-terminal sequence 29 to 33 of CCK,, when 12”1-CCK,, was used as tracer. The results show that CCK,, is an excellent antigen, particularly in guinea pigs, and that
sensitive radioimmunoassays can be established provided CCK is not exposed to oxidation during the labeling proce-
* This study was supported by grants from the Danish Medical Research Council, j.nr. 512-5273, X2-5675, and 512-7230. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “‘uduer- tisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
dure. The sequence-specific radioimmunoassays have proved useful for elucidation of the molecular heterogeneity of cholecystokinin.
Cholecystokinin (CCK) together with gastrin and secretin comprise the three classical and established gastrointestinal hormones. CCK’ regulates gallbladder emptying (“cholecys- tokinin activity”) and secretion of enzymes from the pancreas (“pancreozymin activity”). CCK has been isolated from hog intestines as a triacontatriapeptide, CCK,, (11, the sequence of which is now determined (2) and confirmed in part by synthesis (3, 4). Recently, a CCK variant (CCK,,) with a further 6 residues linked to the NH, terminus of the triacon- tatriapeptide was described (5). A striking feature of both CCKs is that the COOH-terminal pentapeptide sequence, which constitutes the biologically active region of the mole- cules, is identical with the COOH-terminal region of the four characterized molecular forms of gastrin (6-8). The similarity in structure and function of CCK and gastrin suggests a common ancestry, a suggestion for which we recently found evidence (9).
In contrast to the remaining gastrointestinal hormones, sensitive and reliable radioimmunoassays for CCK have so far not been developed, even though CCK is known to be a very important hormone. Three factors have made the development of CCK radioimmunoassays difficult: 1) conventional isotope labeling techniques involve oxidation, which, however, has deleterious effects on the structure of CCK (10). It is, thus, not possible to use these conventional techniques for isotope labeling of CCK. 2) The COOH-terminal sequence of CCK, common also to gastrin, is strongly immunogenic, which renders many antisera raised against CCK unspecific. 3) Only limited amounts of pure CCK have been available.
The present report details the development of highly sensi- tive and sequence-specific radioimmunoassays for CCK. An essential step in this work was to label CCK with preservation of the immunoreactivity by the use of 12”I-hydroxyphenylpro- pionic acid-succinimide ester.
’ The abbreviations used are: CCK, cholecystokinin; CCKsS, tria- contanonapeptide cholecystokinin; CCK,,, triacontatriapeptide cho- lecystokinin; CCKs, COOH-terminal octapeptide of CCK,,; gastrin,,, heptadecapeptide gastrin, gastrin 34, triacontatetrapeptide gastrin.
4016
by guest on February 8, 2020http://w
ww
.jbc.org/D
ownloaded from

Radioimmunoassay for Cholecystohinin 4017
TABLE I
Effect of oxidation on immunoreactivity of various cholec,ystokinins (data in picomoles and percentage)
CCK, CC&, CWx Oxidant Antibody no.
4698 2609 4418 4698 2609 4478 4698 2609
Control (No oxidant)
Chloramine-T
(5 CLP) Chloramine-T
GO I.%) Lactoperoxidase
(4 Pd Lactoperoxidase
(20 pg)
pm01 %
pm01 %
pm01 %
pm01 %
pm01 %
836 865 837 728 825 740 780 819 100 100 100 100 100 100 100 100
42 68 0 56 101 250 187 72
5 8 0 8 12 29 24 9 0 0 0 0 17 0 23 15
0 0 0 0 2 0 3 2
167 147 259 166 156 163 125 146
20 17 31 23 19 22 16 18
34 61 67 36 33 22 0 16
4 7 8 5 4 3 0 2
MATERIALS AND METHODS
Preparation of CCK Antigen -Two CCK preparations were used. I) Porcine CCK,, (30 mg, 20% purified CCK, a generous gift from V. Mutt, Gastrointestinal Hormone Laboratory, Karolinska Institutet, Stockholm) was dissolved in 1.0 ml of N,N-dimethylformamide and conjugated to 30 mg of bovine serum albumin (the Danish Serum Institute, Copenhagen) dissolved in 2.5 ml of 0.05 M sodium phos- phate, pH 7.5, by addition of 135 mg of l-ethyl-3(3-dimethylamino- propyl)carbodiimide-HCl (Lot. 400119 (Sigma)) as described by Good- friend et al. (111, but with a molar ratio between hapten, carrier, and carbodiimide as used for gastrin (12). The reagents were mixed for 20 h at 20°C and then dialyzed exhaustively for 48 h at 4°C against 0.05 M sodium phosphate, pH 7.5. The conjugate was divided into six portions, stored at -20°C. 2) Porcine CCK,, (30 mg, 20% pure, from V. Mutt, Stockholm) was dissolved in 12 ml of 0.05 M
sodium phosphate, pH 7.5, and divided into four portions which were stored at - 20°C.
say (16, 17). 1 nmol of octa-, triacontatria-, and triacontanonapeptide CCK in 10 ~1 of 0.05 M sodium phosphate, pH 7.5, was mixed with either 4 pg or 20 pg of lactoperoxidase and 2 nmol of H,O, (17) or 5 or 50 pg of chloramine-T for 30 s. After addition of 50 pg of sodium metabisulfite the material was gel filtered on Sephadex G-10 col- umns using 0.05 M sodium phosphate, pH 7.5, as eluant. The immunoreactivity of the labeled CCKs was assessed with the ra- dioimmunoassay as described below, using three antisera with different region specificities (4478, 4698, and 2609).
Immunization -The first portion of the conjugate was suspended in 0.85% saline to a total volume of 5 ml and emulsified with an equal volume of Freund’s complete adjuvant (the Danish Serum Institute). Multiple subcutaneous injections of the conjugate were administered in the back of 10 Danish white rabbits (2 to 4 kg weight) totaling approximately 1 mg of conjugate and 100 fig of CCK/rabbit. Five booster injections using Freund’s incomplete ad- juvant were given subcutaneously with 2 to 2.5 months interval. The rabbits were bled from the ear vein 10 to 15 days after the last four boostings. The unconjugated CCK was dissolved in 0.85% saline to a total volume of 8 ml and emulsified with an equal volume of Freund’s complete adjuvant. Multiple subcutaneous injections were administered into the back of 30 guinea pigs (0.5 to 1 kg weight) totaling approximately 250 pg of the porcine gut extract or 50 wg of CCK/guinea pig. Three booster injections using Freund’s incomplete adjuvant were given subcutaneously with 2 to 2.5 months’ interval. The guinea pigs were bled by cardiac puncture 10 to 15 days after the last two boostings. Serum from the bleedings was separated and stored at -20°C. The antisera were designated by the number of the animal.
Radioiodination -The approach used for preparation of 12ZI-CCK was to obtain maximal immunoreactivity by avoiding oxidation and subsequently to obtain a high specific radioactivity by separation of 1251-labeled from unlabeled CCK. Oxidation was avoided by conju- gating 12”I-hydroxyphenylpropionic-succinimide ester to one of the free NH, groups of CCK (lysyl side chains in positions 1 and 11) as described for human thyroid-stimulating hormone (hTSH), human growth hormone (hGH), and human.luteinizing hormone (hLH1 (18). Separation of the tracer from unlabeled CCK was achieved by elution on Sephadex G-50 columns. Samples of 99% pure porcine CCK,, (from V. Mutt, GIH Laboratory, Karolinska Institutet, Stock- holm) were dissolved in 0.2 M CH,COOH and distributed in portions of 5 pg, which were lyophilized and stored at -80°C. At the time of labeling the CCK was dissolved in 4 ~1 of 5% 0.05 M CH,COOH and mixed with 20 ~1 of 0.05 M sodium borate, pH 8.5. The mixture was added to 1 mCi of dried ‘““I-N-succinimidyl-3-(4-hydroxyphenyl) propionate (New England Nuclear). After 30-min vigorous stirring in an ice bath, 0.2 M glycine in 250 ~1 of 0.05 M sodium borate, pH 8.5, was added to the reaction tube, which was stirred for a further 5 min at 0°C. The solution was mixed with 1.5 ml of 0.1 M sodium phosphate, pH 7.5, plus 0.2% gelatin. Fractions of 1.0 ml were collected at 4°C and a flow rate of 5 ml/h. The immunoreactivity and specific radioactivity of the radiolabeled CCK were assessed by its binding to surplus of antiserum (4698) and self-displacement under conditions as previously described for ““I-gastrin (19).
Evaluution of Anitsera -Sera from the immunized animals were evaluated in four respects: 1) titer (defined as the dilution of the serum which can bind 50% of 2.5 fmol of ‘““I-CCK:,, at equilibrium); 2) binding affinity (expressed by the “effective” equilibrium con- stant, K,,,, which is defined as the slope of the curve at zero hormone concentration in a Scatchard diagram (13, 14)). K,,, governs the detection limit of the assay (14). 3) Specificity (expressed in percent- age as the molar ratio between the inhibition dose-50 of CCK,,, and inhibition dose-50 of the CCK congener); and 4) heterogeneity of the antibodies with respect to binding affinities (expressed by the index of heterogeneity as derived from Sips (15)). The potency of CCK,, and CCK congeners to displace ‘““I-CCK,, from the antisera was tested in the following concentrations of the peptides: 0, 3, 10, 30. 100, 1000, 10,000, and 100,000 PM.
Oxidation of CCK -Since oxidation almost entirely removes the biological activity of CCK (10) and since preliminary iodinations of CCK,, and CCK,,2 using the chloramine-T technique (16) yielded tracers of very poor immunoreactivity, the effect of oxidation per se was investigated using chloramine-T and lactoperoxidase as used in conventional iodination of proteins and peptides for radioimmunoas-
Radioimmunoassay Procedure -Unless otherwise indicated, the radioimmunoassay was an equilibrium system carried out at pH 8.4 in disposable flint glass tubes (10 x 75 mm) employing 0.02 M
Verona1 buffer, pH 8.4, containing 0.1% bovine serum albumin (Ortho) as diluent. The 2.4-ml incubation mixtures contained 2.0 ml of diluted anti-CCK serum, 250 ~1 of ““I-CCK,, containing approxi- mately 2000 cpm, and 150 ~1 of the standard CCK (99% pure porcine CCK,,,,) or of the unknown. The standards for approximately 100 experiments were aliquoted into sets of assay tubes which were kept at -20°C until use. The assay was set up at room temperature using an automatic diluter-dispenser (LKB, Sweden). The mixtures were incubated at 4°C for 44 to 48 h, which was necessary to obtain equilibrium. The “bound” (B) and “free” (F) tracers were separated by addition of 0.5 ml of a suspension of activated charcoal (20 mg, Merck, Darmstadt) and plasma (50%, outdated human plasma generously supplied by the Blood Bank, Aarhus County Hospital) in 0.02 M sodium phosphate, pH 7.4, to each tube. The tubes were centrifuged for 10 min at 2000 rpm, and the supernatant (B) and sedimented charcoal (F) were counted with an efficiency of approxi- mately 45% in automaticy scintillation counters (LKB-Wallac) for 5 min and the binding percentage was calculated from the equation
B - ((El + F) x D) ~ x 100
(B + zq ~ ((B + F) x D)
p J. F. Rehfeld, unpublished observation. where D represents the “damage” defined as B x lOO/(B + F) in the
by guest on February 8, 2020http://w
ww
.jbc.org/D
ownloaded from
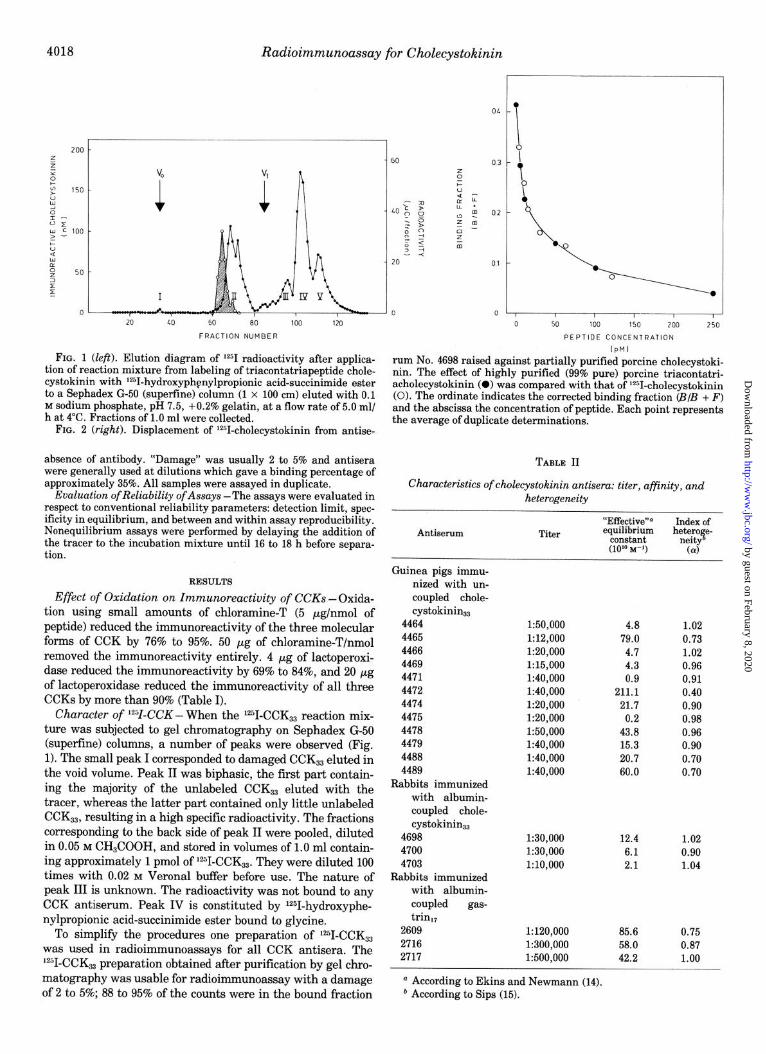
4018 Radioimmunoassay for Cholecystokinin
0
FRACTION NUMBER
FIG. 1 (left). Elution diagram of IsI radioactivity after applica- tion of reaction mixture from labeling of triacontatriapeptide chole- cystokinin with V-hydroxyphenylpropionic acid-succinimide ester to a Sephadex G-50 (superfine) column (1 x 100 cm) eluted with 0.1 M sodium phosphate, pH 7.5, +O.Z% gelatin, at a flow rate of 5.0 ml/ h at 4°C. Fractions of 1.0 ml were collected.
FIG. 2 (rig&). Displacement of ‘*1-cholecystokinin from antise-
absence of antibody. “Damage” was usually 2 to 5% and antisera were generally used at dilutions which gave a binding percentage of approximately 35%. All samples were assayed in duplicate.
Evaluation of Reliability ofAssays -The assays were evaluated in respect to conventional reliability parameters: detection limit, spec- ificity in equilibrium, and between and within assay reproducibility. Nonequilibrium assays were performed by delaying the addition of the tracer to the incubation mixture until 16 to 18 h before separa- tion.
RESULTS
Effect of Oxidation on Immunoreactivity of CCKs -Oxida- tion using small amounts of chloramine-T (5 I.l.g/nmol of peptide) reduced the immunoreactivity of the three molecular forms of CCK by 76% to 95%. 50 pg of chloramine-T/nmol removed the immunoreactivity entirely. 4 pg of lactoperoxi- dase reduced the immunoreactivity by 69% to 84%, and 20 pug of lactoperoxidase reduced the immunoreactivity of all three CCKs by more than 90% (Table I).
Character of V-CCK - When the 1251-CCK33 reaction mix- ture was subjected to gel chromatography on Sephadex G-50 (superfine) columns, a number of peaks were observed (Fig. 1). The small peak I corresponded to damaged CCKa3 eluted in the void volume. Peak II was biphasic, the first part contain- ing the majority of the unlabeled CCK, eluted with the tracer, whereas the latter part contained only little unlabeled CCKs3, resulting in a high specific radioactivity. The fractions corresponding to the back side of peak II were pooled, diluted in 0.05 M CH,COOH, and stored in volumes of 1.0 ml contain- ing approximately 1 pmol of 1251-CCK33. They were diluted 100 times with 0.02 M Verona1 buffer before use. The nature of peak III is unknown. The radioactivity was not bound to any CCK antiserum. Peak IV is constituted by 1251-hydroxyphe- nylpropionic acid-succinimide ester bound to glycine.
To simplify the procedures one preparation of 1251-CCK, was used in radioimmunoassays for all CCK antisera. The 1251-CCK, preparation obtained after purification by gel chro- matography was usable for radioimmunoassay with a damage of 2 to 5%; 88 to 95% of the counts were in the bound fraction
Guinea pigs immu- nized with un- coupled chole- cystokinin,
4464 4465 4466 4469 4471 4472 4474 4475 4478 4479 4488 4489
Rabbits immunized with albumin- coupled chole- cystokinin33
4698 4700 4703
Rabbits immunized with albumin- coupled gas- trin,,
2609 2716 om, y ’ L ‘ * According to Ekins and Newmann (14). b According to Sips (15).
20 01
0 0 0 50 100 150 200 250 7 I I I I I
PEPTIOE CONCENTRATION (PM)
rum No. 4698 raised against partially puritied porcine cholecystoki- nin. The effect of highly purified (99% pure) porcine triacontatri- acholecystokinin (0) was compared with that of L251-cholecystokinin (0). The ordinate indicates the corrected binding fraction (B/B + F) and the abscissa the concentration of peptide. Each point represents the average of duplicate determinations.
TABLE II
Characteristics of cholecystokinin antisera: titer, affinity, and heterogeneity
Antiserum Titer “Effective”” Index of equilibrium
constant heterop-
(1O’O IO) neity
(a)
1:50,000 1:12,000 1:20,000 1:15,000 1:40,000 1:40,000 1:20,000 1:20,000 150,000 1:40,000 1:40,000 1:40,000
1:30,000 12.4 1.02 1:30,000 6.1 0.90 1:10,000 2.1 1.04
1:120,000 1:300,000 1:500,000
4.8 1.02 79.0 0.73 4.7 1.02 4.3 0.96 0.9 0.91
211.1 0.40 21.7 0.90
0.2 0.98 43.8 0.96 15.3 0.90 20.7 0.70 60.0 0.70
85.6 0.75 58.0 0.87 42.2 1.00
by guest on February 8, 2020http://w
ww
.jbc.org/D
ownloaded from
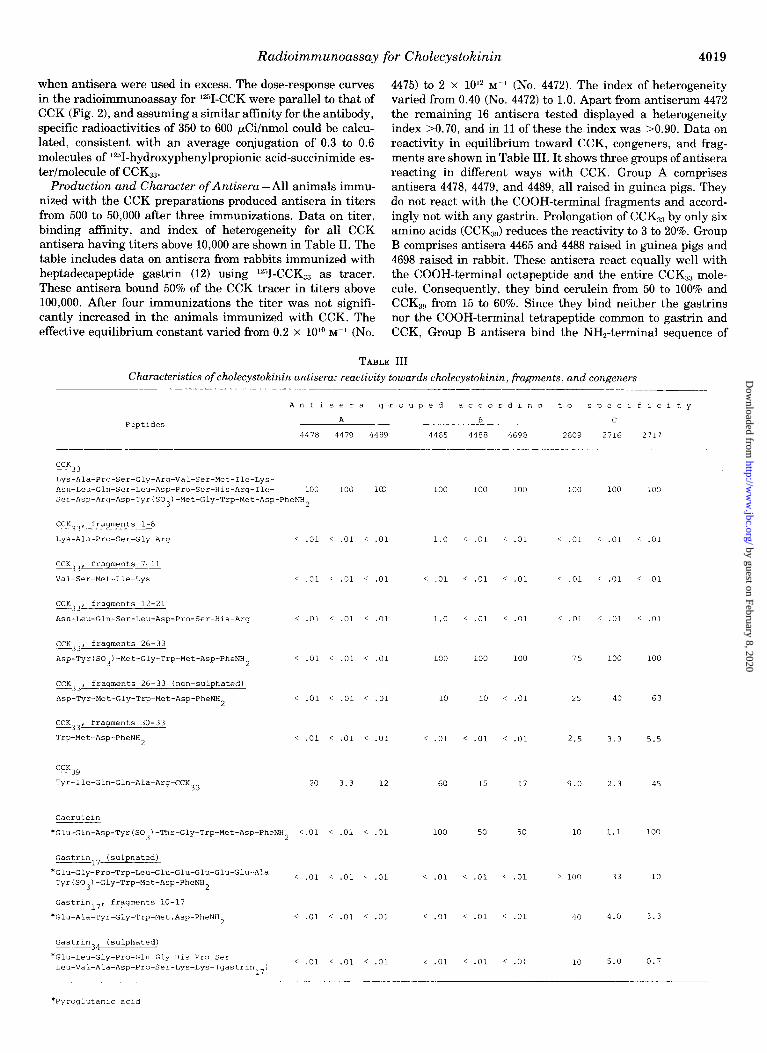
Radioimmunoassay for Cholecystokinin 4019
when antisera were used in excess. The dose-response curves 4475) to 2 x lOI M-’ (No. 4472). The index of heterogeneity in the radioimmunoassay for Y-CCK were parallel to that of varied from 0.40 (No. 4472) to 1.0. Apart from antiserum 4472 CCK (Fig. 2), and assuming a similar affinity for the antibody, the remaining 16 antisera tested displayed a heterogeneity specific radioactivities of 350 to 600 &!i/nmol could be calcu- index >0.70, and in 11 of these the index was BO.90. Data on lated, consistent with an average conjugation of 0.3 to 0.6 reactivity in equilibrium toward CCK, congeners, and frag- molecules of *2”I-hydroxyphenylpropionic acid-succinimide es- ments are shown in Table III. It shows three groups of antisera ter/molecule of CCKs3. reacting in different ways with CCK. Group A comprises
Production and Character of Antisera -All animals immu- antisera 4478, 4479, and 4489, all raised in guinea pigs. They nized with the CCK preparations produced antisera in titers do not react with the COOH-terminal fragments and accord- from 500 to 50,000 after three immunizations. Data on titer, ingly not with any gastrin. Prolongation of CCK,, by only six binding affinity, and index of heterogeneity for all CCK amino acids (CCK,,,) reduces the reactivity to 3 to 20%. Group antisera having titers above 10,000 are shown in Table II. The B comprises antisera 4465 and 4488 raised in guinea pigs and table includes data on antisera from rabbits immunized with 4698 raised in rabbit. These antisera react equally well with heptadecapeptide gastrin (12) using 12”I-CCK,, as tracer. the COOH-terminal octapeptide and the entire CCK,, mole- These antisera bound 50% of the CCK tracer in titers above cule. Consequently, they bind cerulein from 50 to 100% and 100,000. After four immunizations the titer was not signifi- CCK,,, from 15 to 60%. Since they bind neither the gastrins cantly increased in the animals immunized with CCK. The nor the COOH-terminal tetrapeptide common to gastrin and effective equilibrium constant varied from 0.2 x lOlo M-’ (No. CCK, Group B antisera bind the NH,-terminal sequence of
TABLE III Characteristics of cholecystokinin antisera: reactivity towards cholecystokinin, fragments, and congeners
PeptIdes
~-- -___-
AntlSera grouped according to speclflcity
A 8 c
4478 4479 4489 4465 4488 4698 2609 2716 2717
Lys-Ala-Pro-Ser-Gly-Arg-Val-Ser-Met-Ile-Ly~-
Asn-Leu-Gln-Ser-l,eu-Asp-Pro-Ser-His-Arg-Ile- 100 100 1cO Ser-Rsp-Arg-Asp-Tyr(SOj)-Met-Gly-Trp-Met-Asp-Ph~NH2
CCK33. fragments l-6 -__
Lys-Ala-Pro-Ser-Gly-Arq
CCK33, fragments 7-11
"al-Ser-Met-Ile-Lys
CCKjj. fragments 12-21
Asn-Leu-Gin-Ser-Leu-Asp-Pro-Ser-His-Arg
cCK33, fragments 26-33
Asp-Tyr(S03)-Met-Gly-Trp-Met-Asp-Ph.&W2
CCK33, fragments 26-33 (non-sulphated)
Asp-Tyr-Met-Gly-Trp-Met-Asp-PheNH2
CCK33, fragments 30-33
Trp-Met-Asp-PheNH2
CCK,q
Tyr-Ile-Gin-Gln-Ala-Arg-CCK 33
< .Ol < .Ol < .Ol
< .Ol < .Ol < .Ol
< .Ol C .Ol < .Ol
< .Ol < .Ol < .Ol
< .Ol ( .Ol < .Ol
< .Ol < .Ol < .Ol
20 3.3 12
*Glu-Gin-Asp-Tyr(S03)-Thr-Gly-Trp-Met-Asp-PheNH2 C.01 < .Ol < .Ol
Gastrln17 (sulphated) ____
'Glu-Gly-Pro-Trp-Leu-Glu-Glu-Glu-Glu-Glu-Ala Tyr(S03)-Gly-Trp-Met-Asp-PheNH2
< .Ol < .Ol < .Ol
Gastrin , fragments lo-17
'Glu-Ala-Tyr-Gly-Trp-Met.Asp-PheNH2 < .Ol < .Ol < .Ol
Gastru,34 (sulphated)
'Glu-Leu-Gly-Pro-Gin-Gly-His-Pro-Ser- Leu-Val-Ala-Asp-Pro-Ser-Lys-Lys-(gastrin~7)
< .Ol c .Ol < .Ol
100 100
1.0 < .Ol
< .Ol < .Ol
1.0 < .Ol
100 100
10 10
< .Ol < .Ol
60 15
100 50
< .Ol < .Ol
c .Ol < .Ol
c .Ol < .Ol
100
< .Ol
< .Ol
< .Ol
100
< .Ol
< .Ol
17
50
< .Ol
< .Ol
< .Ol
LOO
< .Ol
< .Ol
< .Ol
75
21
2.5
9.0
10
> 100
40
10
100
< .Ol
< .Ol
< .Ol
100
40
3.3
2.3
1.1
33
4.0
5.0
100
< .Ol
< .Ol
< .Ol
100
63
5.5
45
100
10
3.3
0.7
*Pyroqlutam~c acid
by guest on February 8, 2020http://w
ww
.jbc.org/D
ownloaded from
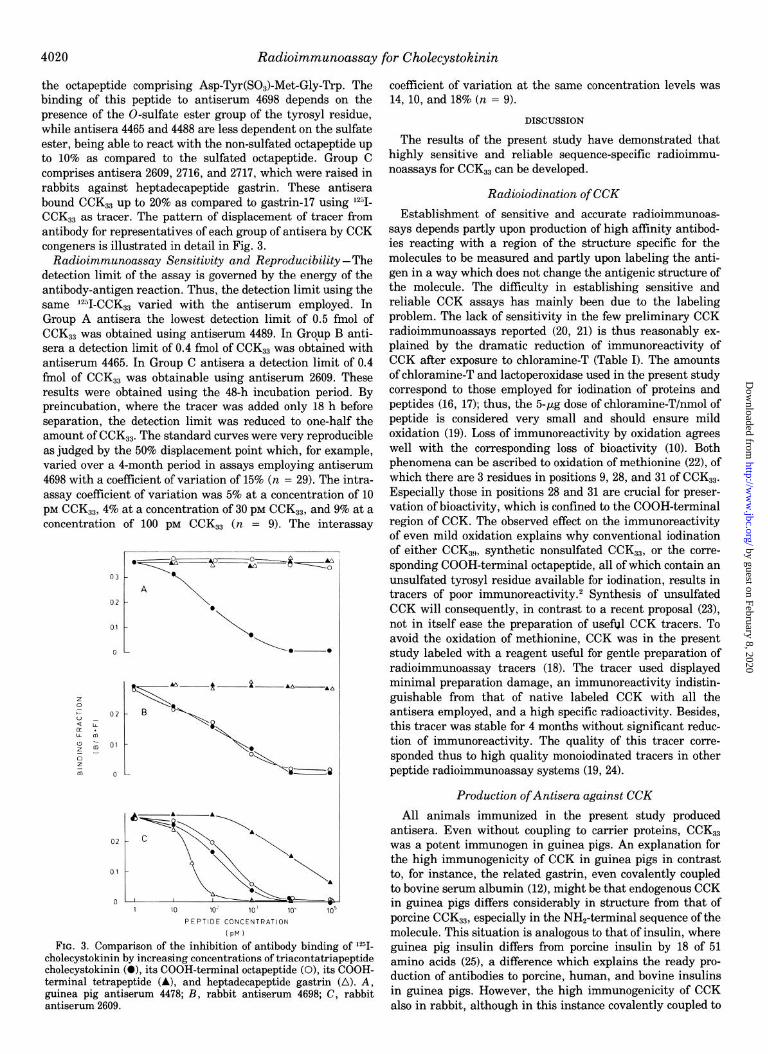
4020 Radioimmunoassay for Cholecystokinin
the octapeptide comprising Asp-Tyr(SO&Met-Gly-Trp. The binding of this peptide to antiserum 4698 depends on the presence of the O-sulfate ester group of the tyrosyl residue, while antisera 4465 and 4488 are less dependent on the sulfate ester, being able to react with the non-sulfated octapeptide up to 10% as compared to the sulfated octapeptide. Group C comprises antisera 2609, 2716, and 2717, which were raised in rabbits against heptadecapeptide gastrin. These antisera bound CCKs3 up to 20% as compared to gastrin-17 using lz51- CCKs3 as tracer. The pattern of displacement of tracer from antibody for representatives of each group of antisera by CCK congeners is illustrated in detail in Fig. 3.
Radioimmunoassay Sensitivity and Reproducibility-The detection limit of the assay is governed by the energy of the antibody-antigen reaction. Thus, the detection limit using the same ‘2”I-CCK,, varied with the antiserum employed. In Group A antisera the lowest detection limit of 0.5 fmol of CCK,, was obtained using antiserum 4489. In Group B anti- sera a detection limit of 0.4 fmol of CCKs3 was obtained with antiserum 4465. In Group C antisera a detection limit of 0.4 fmol of CCK,, was obtainable using antiserum 2609. These results were obtained using the 48-h incubation period. By preincubation, where the tracer was added only 18 h before separation, the detection limit was reduced to one-half the amount of CCKs3. The standard curves were very reproducible as judged by the 50% displacement point which, for example, varied over a 4-month period in assays employing antiserum 4698 with a coefficient of variation of 15% (n = 29). The intra- assay coefficient of variation was 5% at a concentration of 10 pM CCKs3, 4% at a concentration of 30 PM CCKz3, and 9% at a concentration of 100 PM CCKz3 (n = 9). The inter-assay
-8 o3 b~UfG===a”==z;n.-- -=..--A:
A
02 - \ .
01 - \
0 - .-.
PEPTIDE CONCENTRATION
I PM I
FIG. 3. Comparison of the inhibition of antibody binding of T- cholecystokinin by increasing concentrations of tr&contatr&peptide cholecvstokinin (0). its COOH-terminal OCtaDeDtide (0). its COOH- terminal tetrapeptide (A), and heptadecap$ide gas&in (A). A, guinea pig antiserum 4478; B, rabbit antiserum 4698; C, rabbit antiserum 2609.
coefficient of variation at the same concentration levels was 14, 10, and 18% (n = 9).
DISCUSSION
The results of the present study have demonstrated that highly sensitive and reliable sequence-specific radioimmu- noassays for CCKa3 can be developed.
Radioiodination of CCK
Establishment of sensitive and accurate radioimmunoas- says depends partly upon production of high affinity antibod- ies reacting with a region of the structure specific for the molecules to be measured and partly upon labeling the anti- gen in a way which does not change the antigenic structure of the molecule. The difficulty in establishing sensitive and reliable CCK assays has mainly been due to the labeling problem. The lack of sensitivity in the few preliminary CCK radioimmunoassays reported (20, 21) is thus reasonably ex- plained by the dramatic reduction of immunoreactivity of CCK after exposure to chloramine-T (Table I). The amounts of chloramine-T and lactoperoxidase used in the present study correspond to those employed for iodination of proteins and peptides (16, 17); thus, the 5-pg dose of chloramine-T/nmol of peptide is considered very small and should ensure mild oxidation (19). Loss of immunoreactivity by oxidation agrees well with the corresponding loss of bioactivity (10). Both phenomena can be ascribed to oxidation of methionine (22), of which there are 3 residues in positions 9,28, and 31 of CCK%. Especially those in positions 28 and 31 are crucial for preser- vation of bioactivity, which is confined to the COOH-terminal region of CCK. The observed effect on the immunoreactivity of even mild oxidation explains why conventional iodination of either CCK,!,, synthetic nonsulfated CCKz3, or the corre- sponding COOH-terminal octapeptide, all of which contain an unsulfated tyrosyl residue available for iodination, results in tracers of poor immunoreactivity.2 Synthesis of unsulfated CCK will consequently, in contrast to a recent proposal (231, not in itself ease the preparation of useful CCK tracers. To avoid the oxidation of methionine, CCK was in the present study labeled with a reagent useful for gentle preparation of radioimmunoassay tracers (18). The tracer used displayed minimal preparation damage, an immunoreactivity indistin- guishable from that of native labeled CCK with all the antisera employed, and a high specific radioactivity. Besides, this tracer was stable for 4 months without significant reduc- tion of immunoreactivity. The quality of this tracer corre- sponded thus to high quality monoiodinated tracers in other peptide radioimmunoassay systems (19, 24).
Production of Antisera against CCK
All animals immunized in the present study produced antisera. Even without coupling to carrier proteins, CCK,, was a potent immunogen in guinea pigs. An explanation for the high immunogenicity of CCK in guinea pigs in contrast to, for instance, the related gastrin, even covalently coupled to bovine serum albumin (121, might be that endogenous CCK in guinea pigs differs considerably in structure from that of porcine CCKs3, especially in the NH,-terminal sequence of the molecule. This situation is analogous to that of insulin, where guinea pig insulin differs from porcine insulin by 18 of 51 amino acids (25), a difference which explains the ready pro- duction of antibodies to porcine, human, and bovine insulins in guinea pigs. However, the high immunogenicity of CCK also in rabbit, although in this instance covalently coupled to
by guest on February 8, 2020http://w
ww
.jbc.org/D
ownloaded from

Radioimmunoassay for Cholecystokinin 4021
bovine serum albumin, caused antisera of high titers and preparations. I am grateful to Dr. M. Ondetti, The Squibb binding affinities. Recent postulates about CCK,, being a poor Institute of Medical Research, Princeton, N. J., for his gener- immunogen (26) are, thus, not correct and originate rather in ous gifts of synthetic cholecystokinin fragments. The expert the use of CCK tracers of low specific radioactivity and technical assistance of Bodil Basse and Gitte Hollose is grate- immunoreactivity, as discussed above. fully acknowledged.
Character of Antisera against CCK
The CCK antisera of high titers and avidities reacted with different regions of CCK,,.
Group A - Antisera (Table III and Fig. 3) did not react with any gastrin. Thus, they seemed to require the entire triacon- tatriapeptide. However, since these antisera did bind a molec- ular form of CCK in gut extracts smaller than CCKY3, and since this form was also measured by antiserum 4698 and cross-reacting gastrin antisera, as shown in the following article (27), it is likely that the antisera of Group A react with the COOH-terminal half of CCK,,, particularly the sequence 20 to 25, which was not available for testing.
Group B -Antisera (Table III and Fig. 3) reacted fully with both the entire triacontatriapeptide and with its COOH-ter- minal octapeptide. Since these antisera did not react with COOH-terminal tetrapeptide nor with any gastrins containing the same COOH-terminal pentapeptide as CCKs3, they are obviously directed against the NH,-terminal sequence of the octapeptide, region 25 to 30 of the amino acid sequence of CCL.
Group C- Antisera (Table III and Fig. 3) are composed of antisera raised against heptadecapeptide gastrin (121, react- ing avidly with the COOH-terminal pentapeptide common to both gastrin and cholecystokinin. They are, thus, specific for region 29 to 33 of cholecystokinin. Using ““I-CCK,, as tracer these gastrin antisera reacted in some instances better with CCK,, than with the gastrins. Using the antisera with the highest binding affinities, detection limits below 1 PM were encountered. This detection limit is of the same order of magnitude as obtained by the most sensitive gastrin (28-30) and secretin radioimmunoassays (31-33).
It is noteworthy that 11 out of 15 CCK antisera displayed heterogeneity indices ~0.9 (Table II). This observation de- serves two comments. First, most CCK antisera are appar- ently monodisperse with respect to binding energy, an unu- sual feature for antisera in general. A similar homogeneity in binding energy has, however, been observed also for antisera against heptadecapeptide gastrin (12, 34). It is consequently possible that the two related hormones, gastrin and CCK, pose a relatively homogenous antigenic stimulus in accord- ance with the theory of Haber et al. (35). The second implica- tion of the observed homogeneity in binding energy is that the tracer has to be structurally homogenous in order to result in the observed high indices of heterogeneity with antisera specific for different regions of CCK,,. The results of the present heterogeneity studies consequently provide further evidence that the CCK tracer is of high structural integrity.
Acknowledgments -I thank professor V. Mutt, Department of Biochemistry, The Karolinska Institute, Stockholm, for helpful discussions and generous gifts of many cholecystokinin
2. 3.
4.
5. 6. 7. 8.
9. 10. 11.
12.
13. Scatchard, G. (1949)Ann. N. Y. Acad. Sci. 51, 660-672 14. Ekins, R., and Newmann, B. (1970) Acta Endocrinol. (Cph.) 147
15. 16.
17.
18.
19.
20.
21.
22.
23.
24.
25. 26.
27. 28.
29. 30.
31.
32.
33.
34.
35
REFERENCES Mutt, V., and Jorpes, J. E. (1967) Biochem. Biophys. Res.
Commun. 26, 392-397 Mutt, V., and Jorpes, E. (1971) Biochem. J. 125, 57P-58P Ondetti, M. A., PlusEec, J., Sabo, E. F., Sheehan, J. T., and
Williams, N. (1970)5. Am. Chem. Sot. 92, 195-199 Bodanszky, M., Chaturvedi, N., Hudson, D., and Itoh, M. (1972)
J. Org. Chem. 37, 2303-2310 Mutt, V. (1976) C’lin. Endocrinol. 5, 175s-183s Gregory, R. A., and Tracy, H. J. (1964) Gut 5, 103-117 Rehfeld, J. F. (1972) Biochim. Biophys. Acta 285, 364-372 Gregory, R. A., and Tracy, H. J. (1975) in Gastrointestinal
Hormones (Thompson, J. C., ed) pp. 13-24, University of Texas Press, Austin
Larsson, L.-I., and Rehfeld, J. F. (1977) Nature, 269, 335-338 Mutt, V. (1964)Acta Chem. &and. 18, 2185-2186 Goodfriend, T. L., Levine, L., and Fasman, G. D. (1964) Science
144, 1344-1346 Rehfeld, J. F., Stadil, F., and Rubin, B. (1972) Stand. J. Clin.
Lab. Invest. 30, 221-232
(Suppl.), 11-36 Sins. R. (1948) J. Chem. Phvs. 16. 490-495 Hunter, W. M., and Greenwood, F. C. (1962) Nature 194, 495-
496 Thorell, J. I., and Johansson, B. G. (1971) Biochim. Biophys.
Acta 251, 363-369 Bolton, A. E., and Hunter, W. M. (1973) Biochem. J. 133, 529-
539 Stadil, F., and Rehfeld, J. F. (1972) Stand. J. Clin. Lab. Invest.
30, 361-368 Go, V. L. W., Ryan, R. J., and Summerskill, W. H. J. (1971) J.
Lab. Clin. Med. 77, 684-689 Harvey, R. F., Dowsett, L., Hartog, M., and Read, A. E. (1973)
Lancet II 826-827 Dedman, M. L., Farmer, T. H., and Morris, C. J. 0. R. (1961)
Biochem. J. 78, 348-352 Yajima, H., Mori, Y., Kiso, Y., Koyama, K., Tobe, T., Seto-
yama, M., Adachi, H., Kanno, T., and Saito, A. (1976) Chem. Pharmacol. Bull. 24 (51, 1110-1113
Damkjaer Nielsen, M., Jorgensen, M., and Giese, J. (1971) Acta Endocrinol. (Cph.) 67, 104-116
Smith, L. F. (1966) Am. J. Med. 40, 662-666 Bloom, S. R. (1977) in Gastrointestinal Physiology (Crane, R. K.,
ed) Vol. 3, PP. 71-103. Universitv Park Press. Baltimore Rehfeld, J. F: 11978) J. &ioZ. Chem. 253, 4022-4030 Rehfeld, J. F., and Stadil, F. (1973) Sand. J. Clin. Lab. Znuest.
31, 459-464 Yalow, R. S., and Berson, S. A. (1970) Gastroenterology 58, l-14 McGuigan, J. E., and Trudeau, W. L. (1970) Gastroenterology
58, 139-150 Tai, H. H., Korsch, B., and Chey, W. Y. (1975) Anal. B&hem.
69, 34-42 Bloom, S. R. (1975) in Gastrointestinal Hormones (Thompson, J.
C., ed) pp. 257-268, University of Texas Press, Austin Fahrenkrug, J., Schaffalitzky de Muckadell, 0. B., and Rehfeld,
J. F. (1976) Stand. J. Clin. Lab. Inuest. 36, 281-287 Goldenberg, K., Berkowitz, J. M., and Praissman, M. (1974) J.
Zmmunol. 112,1008-1018 Haber, E., Richards, F. F., Spragg, J., Austen, K. F., Valloton,
M., and Page, L. B. (1967) Cold Spring Harbor Symp. Quant. Biol. 32, 299-310
by guest on February 8, 2020http://w
ww
.jbc.org/D
ownloaded from

J F Rehfeldcholecystokinin.
sequence-specific radioimmunoassays for porcine triacontatriapeptide Immunochemical studies on cholecystokinin. I. Development of
1978, 253:4016-4021.J. Biol. Chem.
http://www.jbc.org/content/253/11/4016.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/253/11/4016.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 8, 2020http://w
ww
.jbc.org/D
ownloaded from