Embed Size (px)
Citation preview

Section IIB Omics
Improving Crop Resistance to Abiotic Stress, First Edition.Edited by Narendra Tuteja, Sarvajeet Singh Gill, Antonio F. Tiburcio, and Renu Tuteja� 2012 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2012 by Wiley-VCH Verlag GmbH & Co. KGaA.
j241

10Functional Genomics of Drought Tolerance in Crops: EngineeringTranscriptional Regulators and PathwaysBala Rathinasabapathi
Drought stress is a major limitation for crop productivity worldwide, especially inirrigated agriculture. One solution to this problem is the use of cultivars that aretolerant to drought. However, conventional breeding to improve drought tolerancehas proven difficult because drought and dehydration tolerance in plants are not fullyunderstood. As a result of research during the past two decades, networks of genesparticipating in plant adaptations to drought stress have been identified. In thischapter, I present this research progress and indicate the use of specific genes toengineer crops for improved drought tolerance.
10.1Introduction
Drought stress causes severe devastation to crop productivity worldwide. Economiclosses due to the occurrence of a combination of drought andhigh-temperature stresson crops are enormous. For example, one estimate for economic losses caused bydrought and high-temperature stress for a 5-year period (2005–2009) totaled up toUS$19 billion in the United States [1].
Because water resources are becoming limited, genetic improvement of cropsfor increased tolerance to drought has become an important aim in agriculturalresearch. Drought and high-temperature tolerance in plants are complex traitswith various interacting components. These include plant adaptations related tocellular tolerance to oxidative, osmotic, and high-temperature stress. Otheradaptations relate to physiological features that enhance water uptake by theroots, and adaptations that decrease the loss of water through transpiration.Developmental adaptations related to flowering time and partitioning of biomassbetween leaves and grain are also important for crop productivity under stress.Some progress has been made by the use of conventional breeding to improvedrought tolerance in important crops such as rice [2], corn [3], cowpea [4], andchickpea [5], but the use of recombinantDNA and genomicmethods are likely to becrucial in the near future.
j243
Improving Crop Resistance to Abiotic Stress, First Edition.Edited by Narendra Tuteja, Sarvajeet Singh Gill, Antonio F. Tiburcio, and Renu Tuteja� 2012 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2012 by Wiley-VCH Verlag GmbH & Co. KGaA.

With the availability of genome sequence data for plants, microbes, and otherorganisms, and technologies to transfer and express foreign genes in crop plants,metabolic engineering has become an important tool for improving crops fortolerance to drought and high-temperature stress. Much of our current understand-ing of the genes playing roles in drought tolerance has come from the studies ofmodel species such as Arabidopsis thaliana and rice. High-throughput functionalgenomic tools to assess gene and protein expression in response to stress have greatlycontributed to rapid advances in this area. The objective of this chapter is to highlightsome of the most successful metabolic engineering attempts in improving droughtand high-temperature tolerance in crops by using rice as a crop model and suggestfuture opportunities with the greatest promise.
10.2Transcriptional Regulation of Stress Signaling Networks
Upon stress, specific networks of plant genes are upregulated leading to improvedplant tolerance to stress. In a metabolic engineering strategy termed �regulonengineering,� ectopic expression of specific transcription factors (TFs) at the earlypart of a signaling cascade is used to upregulate the transcription of a number ofgenes resulting in stress tolerance [6, 7]. An estimated 1300 TFs are known in ricegenome and about 45% are from gene families specific to plants [8]. Ray et al. [9]showed that in rice 5611 transcripts were modulated by water-deficit stress condi-tions [8]. Among about 50 families of TFs, several have been identified to havepossible roles in drought-stress tolerance. Members of the zinc finger TFs wereupregulated under drought and NAC family transcription factors were upregulatedby multiple abiotic stresses such as water deficit, salt, and cold stress [9].
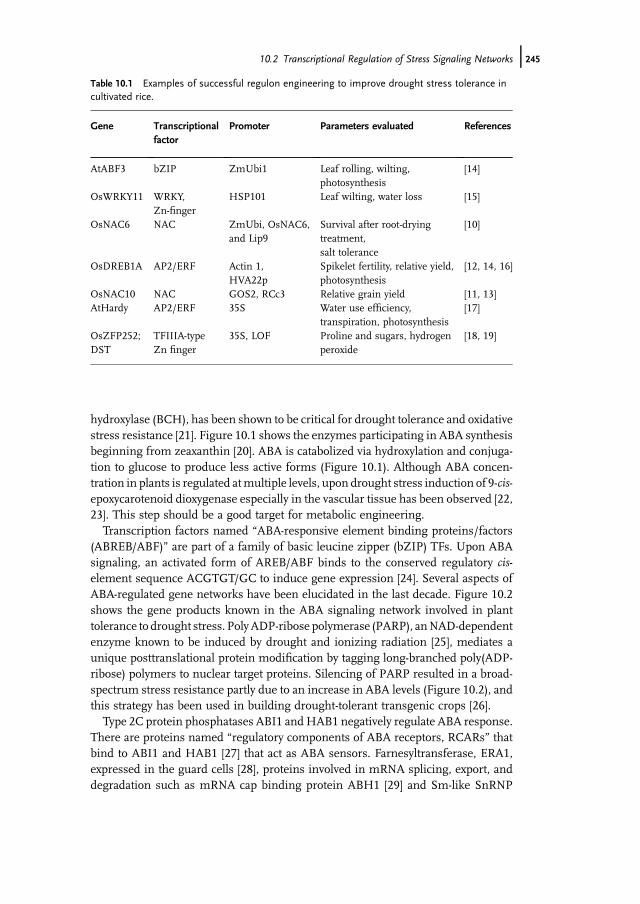
However, constitutive expression of the TFs often results in stunted growth andother developmental problems in the plant [10]. Expression of TFs under stress-inducible promoters such as RD29a, OsNAC6, and HSP101 have eliminated orreduced these problems [10]. Many early studies concentrated on evaluatingtransgenic plants ofmodel species for stress tolerance under laboratory conditions.These have suggested roles for specific transcription factors in drought tolerance.Recent studies, however, have evaluated transgenic crops expressing TFs under thecontrol of inducible or tissue-specific promoters, under field drought condi-tions [11–13]. Table 10.1 lists examples for successful regulon engineering toimprove drought and high-temperature stress tolerance in rice, although much ofthe basic information on the transcription factors emerged from studies in themodel plant A. thaliana.
10.2.1ABA-Inducible Transcription Factors
The growth regulator abscisic acid (ABA) is synthesized from the C40 carotenoidprecursor phytoene [20]. The step that leads to zeaxanthin, catalyzed by b-carotene
244j 10 Functional Genomics of Drought Tolerance in Crops
hydroxylase (BCH), has been shown to be critical for drought tolerance and oxidativestress resistance [21]. Figure 10.1 shows the enzymes participating in ABA synthesisbeginning from zeaxanthin [20]. ABA is catabolized via hydroxylation and conjuga-tion to glucose to produce less active forms (Figure 10.1). Although ABA concen-tration in plants is regulated atmultiple levels, upon drought stress induction of 9-cis-epoxycarotenoid dioxygenase especially in the vascular tissue has been observed [22,23]. This step should be a good target for metabolic engineering.
Transcription factors named �ABA-responsive element binding proteins/factors(ABREB/ABF)� are part of a family of basic leucine zipper (bZIP) TFs. Upon ABAsignaling, an activated form of AREB/ABF binds to the conserved regulatory cis-element sequence ACGTGT/GC to induce gene expression [24]. Several aspects ofABA-regulated gene networks have been elucidated in the last decade. Figure 10.2shows the gene products known in the ABA signaling network involved in planttolerance to drought stress. PolyADP-ribose polymerase (PARP), anNAD-dependentenzyme known to be induced by drought and ionizing radiation [25], mediates aunique posttranslational protein modification by tagging long-branched poly(ADP-ribose) polymers to nuclear target proteins. Silencing of PARP resulted in a broad-spectrum stress resistance partly due to an increase in ABA levels (Figure 10.2), andthis strategy has been used in building drought-tolerant transgenic crops [26].
Type 2C protein phosphatases ABI1 andHAB1 negatively regulate ABA response.There are proteins named �regulatory components of ABA receptors, RCARs� thatbind to ABI1 and HAB1 [27] that act as ABA sensors. Farnesyltransferase, ERA1,expressed in the guard cells [28], proteins involved in mRNA splicing, export, anddegradation such as mRNA cap binding protein ABH1 [29] and Sm-like SnRNP
Table 10.1 Examples of successful regulon engineering to improve drought stress tolerance incultivated rice.
Gene Transcriptionalfactor
Promoter Parameters evaluated References
AtABF3 bZIP ZmUbi1 Leaf rolling, wilting,photosynthesis
[14]
OsWRKY11 WRKY,Zn-finger
HSP101 Leaf wilting, water loss [15]
OsNAC6 NAC ZmUbi, OsNAC6,and Lip9
Survival after root-dryingtreatment,salt tolerance
[10]
OsDREB1A AP2/ERF Actin 1,HVA22p
Spikelet fertility, relative yield,photosynthesis
[12, 14, 16]
OsNAC10 NAC GOS2, RCc3 Relative grain yield [11, 13]AtHardy AP2/ERF 35S Water use efficiency,
transpiration, photosynthesis[17]
OsZFP252;DST
TFIIIA-typeZn finger
35S, LOF Proline and sugars, hydrogenperoxide
[18, 19]
10.2 Transcriptional Regulation of Stress Signaling Networks j245

protein, SAD1, are negative regulators of ABA signaling [30]. PYR1 and other ABAreceptors PYL1 and PYL2 bind to ABA to inhibit type 2C protein phosphatases, thuscontrolling SnRK2 kinase activity [30–32]. PYR1�s role in negative regulation on ABAsignaling was discovered by the use of a synthetic growth inhibitor pyrabactin [33]employing a chemical genetics approach.
Other inhibitory factor DOR, a putative F-box protein interacting with ASK14 andCUL1, is a negative regulator of ABA-mediated stomatal closure [34]. When twonegative regulators of ABA signaling were mutated simultaneously [35], the doublemutants showed strong hypersensitivity to ABA, and decreased water loss, suggest-ing this approach to be a way to improve crop performance under drought.
ABA signaling is also regulated via protein modifications and degradations. Iso-prenylated proteins contain an isoprenylcysteine methyl ester at the C-terminus. Anisoprenylcysteine methyltransferase (ICMT) is a negative regulator of ABA signal-ing [36]. Recently, an E3 ubiquitin ligase (AIRP1), a cytosolic protein involved indegradation of ubiquitinated proteins, was demonstrated to function as a positiveregulator of ABA-dependent response to drought stress [37]. In contrast, E3 ligase KEG(KEEPONGOING),whichubiquitinatesABI5, negatively regulatesABAsignaling [38].ABA promotes ABI5 accumulation by inducing the ubiquitination and degradation ofKEG [39]. A multitude of regulatory controls on ABA-mediated signaling has given anumber of new technologies to engineer crops for improved drought tolerance [40].
Zeaxanthin
AntheraxanthinZEP
ZEP
VDE
VDE
Violaxanthin
NSY
Neoxanthin
ISO? ISO?
9’-cis-Neoxanthin 9-cis-Violaxanthin
XanthoxinNCED NCED
H d l i and
Xanthoxin
Abscisic aldehydeABA2
Hy roxylationconjugation to glucoseAbscisic acid
AAO3
Figure 10.1 Synthesis and catabolism ofabscisic acid. The enzymes are ZEP: zeaxanthinepoxidase; VDE: violaxanthin deepoxidase;NSY: neoxanthin synthase; ISO: isomerase;
NCED: 9-cis-epoxycarotenoid dioxygenase;ABA2: short-chain alcohol dehydrogenase; andAAO3: aldehyde oxidase.
246j 10 Functional Genomics of Drought Tolerance in Crops

However, antagonistic crosstalk between ABA signaling and systemic acquired resis-tance (SAR),mediated by salicylic acid [41], suggests that protecting crops fromdroughtstress damage via modulating ABA-signaling could have unintended consequences oncrop tolerance to biotic stress. More research is needed to examine this point.
10.3ABA-Independent Signaling Networks
10.3.1NAC Transcription Factors
Plants also regulate stress tolerance-related genes via ABA-independent pathways.Figure 10.3 shows the gene products involved in ABA-independent response todrought stress. NAM, ATAF, and CUC (NAC) transcription factors contain a highlyconserved N-terminal DNA binding domain and a variable C-terminal domain [42].Stress-related NAC TFs (SNACs) belong to group III, phylogenetically distinct fromfour other groups [43]. Specific NAC TFs induced by stress were identified indifferent plant species. SNAC1 inArabidopsis is expressed in the stomata. Transgeniccrops overexpressing SNAC1 protein had improved tolerance to drought [11].
DROUGHT
PARP LOS5, AIRP1
ABA
,
PYR1 chaperone
DownregulationER1, ABH1, SAD1, DOR, KEG, ICMT
ABI1
SnRK2SLAC1
Upregulation
ModificationPLD
AREBROS
SNAC
ABRE sites of promoters in ABA-responsive genesStomatal closure
RD29, LEA proteins, HSP, ZAT10
Synthesis of protectants
Figure 10.2 ABA-dependent signalingpathway leading to drought tolerance in plants.AIRP1: ABA-insensitive RING protein1; PARP:poly(ADP-ribose) polymerase; LOS5:molybdenum cofactor sulfurase; PYR1: ABAreceptor; PLD: phospholipase D; SLAC1: slowanion channel-associated 1; ABI1: type 2Cprotein phosphatase; ER1: farnesyltransferase
b-subunit; ABH1: mRNA cap-binding protein;ICMT: isoprenylcysteine methyltransferase;SAD1: Sm-like SnRNP protein; SnRK2: kinase;AREB: ABA-responsive element bindingprotein; LEA: dehydrin; HSP: heat shockprotein; ZAT10: salt tolerance zinc finger;RD29A: responsive to desiccation 29A.
10.3 ABA-Independent Signaling Networks j247

10.3.2DREB/CBF Transcription Factors
Yamaguchi-Shinozaki and Shinozaki identified TFs that are induced by drought butare not under ABA control. Some of these ABA-independent TFs (DREB1/CBF) areinvolved in cold tolerance andDREB2 (dehydration-responsive element binding 2) isimportant in drought tolerance [24, 43]. DREB2s interact with a cis-acting DRE/CRT(C-repeat) sequence to activate the expression of downstream genes. Overexpressionof DREB2-type transcription factors has been used as a strategy to achieve improvedtolerance to drought in many crop species [43–46]. In rice, and in other cereals, twotypes of transcripts of DREB2B – active and inactive – are produced by alternativesplicing. Upon stress, the relative amounts of active transcripts increase, likely by aregulation on the splicing machinery [46].
MicroRNAs are small noncoding single-stranded RNAs that regulate gene expres-sion by target mRNA cleavage and translational repression. Zhao et al. [47] showedthat in rice miR-169g was the only member induced by drought in the miR-169family [47] and they suggested that miR-169g could transiently inhibit NF-YAtranscription factor [48].
DROUGHT
PLC
Ca2+
CDPK
AREB
DREBNFs
iR169AREB
Stomatal
Binding to cis-elements
miR169
Transcriptional
Induction of LEA, ZAT10, HSPSynthesis of Osmoprotectants
closure regulation
Figure 10.3 ABA-independent signalingpathway for drought stress tolerance. PLC:phospholipase C; CDPK: Ca-dependent proteinkinase; AREB: ABA-responsive element binding
protein; DREB: dehydration-responsive elementbinding; SNAC: stress-responsive NAC;miR169: microRNA169; NFs: nuclear factor Yproteins.
248j 10 Functional Genomics of Drought Tolerance in Crops

Although manipulation of TFs would continue to be an important strategy forachieving drought-tolerant plants, its limitation in its present form would be thatonly a level of stress tolerance already availablewithin the species could be achieved.While further intricate details on the roles of genes involved in these stressregulatory pathways will continue to emerge, we also need studies searching forstructural genes involved in pathways leading to stress tolerance in naturally stress-tolerant species. Future metabolic engineering strategies should combine themanipulation of transcriptional network and introduce novel pathways from stresstolerant species.
10.4Pathways for Osmoprotectant Synthesis
Many plants synthesize small molecules in response to stress. Osmoprotectants arecompatible solutes usually derived fromamino acids and sugars. Amino acid proline;the quaternary ammonium compounds glycine betaine, proline betaine, and b-ala-nine betaine; sugar alcohols mannitol, sorbitol, and trehalose; and raffinose familyoligosaccharides have been identified as osmoprotectants in plants [49]. Accumu-lation of osmoprotectants could have multiple benefits for the plant includingosmotic adjustment, turgor maintenance, and protection of proteins, DNA, andmembranes from damage by stress [50]. A series of studies on engineering plants forglycine betaine overproduction has identified constraints in the availability of theprecursor choline and its import into chloroplasts [51]. Engineering plants foroverproduction of glycine betaine has resulted in plants with improved droughttolerance [52, 53]. In another study, simultaneous stress-inducible expression ofcholine oxidase (for glycine betaine synthesis) and superoxide dismutase andascorbate peroxidase (to improve oxidative stress tolerance) resulted in transgenicpotato plants with increased tolerance to drought [54]. InArabidopsis, raffinose familyoligosaccharides galactinol and raffinose may function as osmoprotectants againstdrought stress [55] and oxidative damage [56]. Transgenic plants overexpressingDREB genes had increased levels of proline suggesting that proline synthesismay beunder the control of DREB. A comparative functional genomics study on maizegenotypes differing in drought tolerance concluded that sugar synthesis, sugartransport, and cell injury prevention are key factors in determining drought toler-ance [57]. Studies are needed to link the signaling pathways for stress to understandhow osmoprotectant synthesis, accumulation, and degradation are regulated both atthe transcriptional and at the translational levels.
10.5Transporters
Drought stress tolerance can be improved if solute concentration in the vacuole couldbe increased, thus favoring water uptake. Apse et al. [58] reported a strategy to increase
10.5 Transporters j249

the activity of a vacuolar sodium/proton antiporter, thus improving plant tolerance tosalinity stress [58]. Similarly,when theactivity ofHþ pumpon the vacuolarmembrane(vacuolar Hþ pyrophosphatase) is increased, the plants gain improved tolerance toboth salinity and drought [59]. This strategy has now been applied to improve droughttolerance in a number of crops including cotton [60, 61] and maize [62].
Aquaporins (or major intrinsic proteins, MIPs) facilitate water transport overcellular membranes. In plants, they occur as a family of 30–35 proteins classified astonoplast intrinsic proteins (TIPs), plasma membrane intrinsic proteins (PIPs),NOD26-like intrinsic proteins (NIPs), and small basic intrinsic proteins (SIPs). InA.thaliana, many of them were downregulated by drought stress at the transcriptionallevel, but two PIPs whose promoters had drought stress response elements wereupregulated [63], suggesting these two PIPs with possible roles in drought tolerance.Constitutive overexpression of PIPs in transgenic plants improved vigor underfavorable growth conditions, but not under drought or salt stress [64]. More researchis needed to engineer stress-regulated PIPs.
10.6Combination of Drought and High-Temperature Stress and Oxidative Stress
In the field, drought often occurs together with high-temperature stress. The biologyof the combination of these two stress factors on plants has not been examined at themolecular level, although it is well known that both drought and high-temperaturestress induce oxidative damage. Ectopic expression of single transgenes involved inosmoprotection or oxidative stress tolerance has resulted in plants with improvedtolerance to high-temperature stress [65, 66]. Plant breeders have recognized thatgenotypes that are resistant to oxidative stress are also resistant to drought and high-temperature stress [67], providing a correlating evidence for the importance ofoxidative stress tolerance. Functional genomic studies on naturally dehydrationtolerant species and the effects of simultaneous application of drought and hightemperature on plants have revealed key roles for antioxidant enzymes in bothdrought and high-temperature stress tolerance [68, 69]. Future work is needed toidentify differences, similarities, and overlaps between signaling pathways leading tohigh-temperature stress and those leading to drought tolerance.
10.7Conclusions
Although Arabidopsis and model crop rice have contributed to our understanding ofhow plants adapt to drought and high-temperature stress, time is ripe for beginningfunctional genomic studies on species that are naturally tolerant to stress. Wildrelatives of major crops could be a starting point [70], although they have not beenused extensively in conventional breeding. Drought tolerance has been identified inwild emmer (Triticum dicoccoides) [71], certain barley genotypes from Tibet [72], wild
250j 10 Functional Genomics of Drought Tolerance in Crops

Oryza spp. and accessions of cultivated rice [73], and pearl millet (Pennisetumglaucum) [74]. Analysis of orthologues of Arabidopsis and rice genes with key rolesin drought stress tolerance pathways in these wild grasses should be useful both touncover novelties and to improve cereal crops for drought tolerance.
Understanding drought tolerance in plants is a long-term goal as this is a valuablebut complex trait. Genome sequences, high-throughput technologies such as micro-arrays, metabolomics, and proteomics have provided us unprecedented tools withcapabilities to identify genes and gene products with potential roles in droughttolerance. However, functional tests on specific genes using mutant and overexpres-sion lines are still laborious and time consuming. To facilitate these tests, new rapidtechnologies to create and analyze stable transgenic lines in major crops are needed.Despite that, as illustrated above, transgenic crops improved for drought tolerancewith great promise are tested under field conditions and perhaps represent the firststeps toward our next green revolution.
Acknowledgment
Funding support from the Consortium for Plant Biotechnology Research Inc. to theauthor�s research program is gratefully acknowledged.
References
1 NESDIS (2010) BillionDollarUSWeatherDisasters. NOAA Satellite andInformation Service, US Department ofCommerce. Available at http://www.ncdc.noaa.gov/oa/reports/billionz.html#chron. (Accessed 15 Feb 2011).
2 Serraj, R., Kumar, A., McNally, K.L. et al.(2009) Adv. Agron., 103, 41–98.
3 Bruce,W.B., Edmeades, G.O., and Barker,T.C. (2002) J. Exp. Bot., 53, 13–25.
4 Agbicodo, E.M., Fatokun, C.A.,Murannaka, S. et al. (2009)Euphytica, 167,353–370.
5 Singh, K.B., Malhotra, R.S., Haliala, M.H.et al. (1994) Euphytica, 73, 137–149.
6 Jaglo-Ottosen, K.R., Gilmour, S.J., Zarka,D.G. et al. (1998) Science, 280, 104–106.
7 Kasuga, M., Liu, Q., Miura, S. et al. (1999)Nat. Biotechnol., 17, 287–291.
8 Kikuchi, S., Satoh, K., Nagata, T. et al.(2003) Science, 301, 376–379.
9 Ray, S., Dansana, P.K., Giri, J. et al. (2010)Funct. Integr. Genomics, 11, 157–178.
10 Nakashima, K., Tran, L.S., VanNguyen,D.et al. (2007) Plant J., 51, 617–630.
11 Hu, H., Dai, M., Yao, J. et al. (2006) Proc.Natl. Acad. Sci. USA, 35, 12987–12992.
12 Xiao, B.Z., Chen, X., Xiang, C.B. et al.(2009) Mol. Plant, 2, 73–83.
13 Jeong, J.S., Kim, S.Y., Baek, K.H. et al.(2010) Plant Physiol., 153, 185–197.
14 Oh, S.J., Song, S.I., Kim, Y.S. et al. (2005)Plant Physiol., 138, 341–351.
15 Wu, X., Shiroto, Y., Kishitani, S. et al.(2009) Plant Cell Rep., 28, 21–30.
16 Wang, Q., Guan, Y., Wu, Y. et al. (2008)Plant Mol. Biol., 67, 589–602.
17 Karaba, A., Dixit, S., Greco, R. et al. (2007)Proc. Natl. Acad. Sci. USA, 104, 15270–15275.
18 Xu,D.Q., Huang, J., Guo, S.Q. et al. (2008)FEBS Lett., 582, 1037–1043.
19 Huang, X.Y., Chao, D.Y., Gao, J.P. et al.(2009) Genes Dev., 23, 1805–1817.
20 Nambara, E. and Marion-Poll, A. (2005)Plant Biol., 56, 165–185.
21 Du,H.,Wang,N., Cui, F. et al. (2010)PlantPhysiol., 154, 1304–1318.
22 Iuchi, S., Kobayashi, M., Taji, T. et al.(2001) Plant J., 27, 325–333.
References j251

23 Endo, A., Sawada, Y., Takahashi, H. et al.(2008) Plant Physiol., 147, 1984–1993.
24 Yamaguchi-Shinozaki, K. and Shinozaki,K. (2006) Plant Biol., 57, 781–803.
25 Doucet-Chabeaud, G., Godon, C.,Brutesco, C. et al. (2001) Mol. Genet.Genomics, 265, 954–963.
26 Vanderauwera, S., Block, D.M., andSteene, N.V. (2007) Proc. Natl. Acad. Sci.USA, 104, 15150–15155.
27 Ma, Y., Szostkiewicz, I., Korte, A. et al.(2009) Science, 324, 1064–1068.
28 Pei, Z.M., Ghassemian, M., Kwak, C.M.et al. (1998) Science, 282, 287–290.
29 Hugouvieux, V., Murata, Y., Young,J.J. et al. (2002) Plant Physiol., 130,1276–1287.
30 Xiong, L.,Gong,Z., Rock,C.D. et al. (2001)Dev. Cell, 1, 771–781.
31 Santiago, J., Dupeux, F., Round, A. et al.(2009) Nature, 462, 665–669.
32 Weiner, J.J., Peterson, F.C., Volkman, B.F.et al. (2010) Curr. Opin. Plant Biol., 13,495–502.
33 Park, S.Y., Fung, P., Nishimura, N. et al.(2009) Science, 324, 1068–1071.
34 Zhang, Y., Xu, W., Li, Z. et al. (2008) PlantPhysiol., 148, 2121–2133.
35 Saez, A., Robert, N., Maktabi, M.H. et al.(2006) Plant Physiol., 141, 1389–1399.
36 Huizinga, D.H., Omosegbon, O., Omery,B. et al. (2008) Plant Cell, 20, 2714–2728.
37 Ryu,M.Y., Cho, S.K., and Kim,W.T. (2010)Plant Physiol., 154, 1983–1997.
38 Liu, H. and Stone, S.L. (2010) Plant Cell,22, 2630–2641.
39 Somvanshi, V.S. (2009) Recent Pat. DNAGene Seq., 3, 16–25.
40 Yasuda, M., Ishikawa, A., Jikumaru, Y.et al. (2008) Plant Cell, 20, 1678–1692.
41 Olsen, A.N., Ernst, H.A., Leggio, L.L. et al.(2005) Trends Plant Sci., 10, 79–87.
42 Fang, Y., You, J., Xie, K. et al. (2008) Mol.Genet. Genomics, 280, 547–563.
43 Sakuma, Y., Maruyama, K., Qin, F. et al.(2006) Proc. Natl. Acad. Sci. USA, 103,18822–18827.
44 Morran, S., Eini, O., Pyvovarenko, T. et al.(2011) Plant Biotechnol. J., 9, 230–249.
45 Chen, J.Q., Meng, X.P., Zhang, Y. et al.(2008) Biotechnol. Lett., 30, 191–198.
46 Matsukura, S., Mizoi, J., Yoshida, T.et al. (2010) Mol. Genet. Genomics, 283,185–196.
47 Zhao, B., Liang, R., Ge, L. et al. (2007)Biochem. Biophys. Res. Commun., 354,585–590.
48 Zhao, B., Ge, L., Liang, R. et al. (2009)BMC Mol. Biol., 10, 29–39.
49 Rathinasabapathi, B. and Kaur, R. (2006)Metabolic engineering for stresstolerance, in Physiology and MolecularBiology of Stress Tolerance (eds K.V.M. Rao,A.S. Raghavendra, and J.K. Reddy),Springer, The Netherlands, pp. 255–299.
50 Rontein, D., Basset, G., and Hanson, A.D.(2002) Metab. Eng., 4, 49–56.
51 Nuccio, M.L., McNeil, S.D., Ziemak, M.J.et al. (2000) Metab. Eng., 2, 300–311.
52 Quan, R., Shang, M., Zhang, H. et al.(2004) Plant Biotechnol. J., 2, 477–486.
53 Shen, B., Hohmann, S., Jensen, R.G., andBohnert, H.J. (1999) Plant Physiol., 121,45–52.
54 Ahmad, R., Kim, Y.H., Kim, M.D. et al.(2010) Physiol. Plant., 138, 520–533.
55 Taji, T., Ohsumi, C., Iuchi, S. et al. (2002)Plant J., 29, 417–426.
56 Nishizawa, A., Yabuta, Y., and Shigeoka, S.(2008) Plant Physiol., 147, 1251–1263.
57 Li, Y., Sun, C., Huang, Z. et al. (2009)PlantCell Physiol., 50, 2092–2111.
58 Apse, M.P., Aharon, G.S., Snedden, W.A.et al. (1999) Science, 285, 1256–1258.
59 Gaxiola, R.A., Li, J., Undurraga, S. et al.(2001) Proc. Natl. Acad. Sci. USA, 98,11444–11449.
60 Lv, S., Zhang, K., Gao, Q. et al. (2008)PlantCell Physiol., 49, 1150–1164.
61 Pasapula, V., Shen, G., Kuppu, S. et al.(2011) Plant Biotechnol., 9, 88–99.
62 Li, B., Wei, A., Song, C. et al. (2008) PlantBiotech. J., 6, 146–159.
63 Alexandersson, E., Danielson, J.A.H.,Rade, J. et al. (2010) Plant J., 61, 650–660.
64 Aharon, R., Shahak, Y., Wininger, S. et al.(2003) Plant Cell, 15, 439–447.
65 Fouad, W.M. and Rathinasabapathi, B.(2006) Plant Mol. Biol., 60, 495–505.
66 Sundaram, S. and Rathinasabapathi, B.(2010) Planta, 231, 361–369.
67 Altinkut, A., Kazan, K., Ipekci, Z., andGozukirmizi, N. (2001) Euphytica, 121,81–86.
68 Oliver, M.J., Dowd, S.E., Zargoza, J. et al.(2004) BMC Genomics, 5, 89.
69 Rizhsky, L., Daveletova, S., Liang, H. et al.(2004) Plant Physiol., 134, 1683–1696.
252j 10 Functional Genomics of Drought Tolerance in Crops

70 Hahhar, H. and Hodgkin, T. (2007)Euphytica, 156, 1–13.
71 Xie,W. andNevo, E. (2008)Euphytica, 164,603–614.
72 Zhao, J., Sun, H., Dai, H. et al. (2010)Euphytica, 172, 395–403.
73 Liu, L., Lafitte, R., and Guan, D. (2004)Euphytica, 138, 149–161.
74 Yadav, R.S., Sehgal, D., and Vadez, V.(2010) J. Exp. Bot., 62, 397–408.
References j253





























