Embed Size (px)
Citation preview

52Mustard: Approaches for Crop Improvementand Abiotic Stress ToleranceSarvajeet Singh Gill, Ritu Gill, Gautam Kumar, Ashwani Pareek,Prabodh C. Sharma, Naser A. Anjum, and Narendra Tuteja
Adverse environmental conditions seriously affect crop growth, productivity,and genome stability. These adverse environmental factors are a menace forplants that prevent them from reaching their full genetic potential and thereforelimit the crop productivity worldwide. Stress conditions, such as extremetemperatures, water availability, and ion toxicity, represent abiotic stresses,which cause massive loss of crop yield. Global climatic pattern is also becomingmore unpredictable with increased occurrence of drought, flood, storms, heatwaves, and seawater intrusion. The oleiferous Brassica is the third mostimportant source of vegetable oil in the world after palm and soybean oil andgrown as an edible or an industrial oil crop that is used as a source of edibleprotein, in much the same way as soybean protein. According to a report ofthe United States Department of Agriculture (USDA), the world oilseed pro-duction was 397 metric ton in 2006–2007 and Indian agriculture contributedabout 15 and 8% to the world total acreage under oilseed cultivation andproduction, respectively. However, the average productivity in India is only791 kg ha�1 compared to the world average of 1718 kg ha�1. Despite a large areaunder cultivation of mustard, the productivity of the crop has dropped in recentyears because plant growth and development are affected by various abioticstress factors. Protecting crop productivity under unfavorable environmentalconditions is a major challenge for modern agriculture. In this review, we haveattempted to provide an overview of success obtained in raising germplasm withimproved salinity tolerance through breeding methods. The recent �Omics�approaches and their applications in abiotic stress research on mustard crop arealso presented.
52.1Introduction
Environmental stress is referred to the extreme environmental conditions that leadto alterations in plant metabolism resulting in decreased rate of plant�s growth
Improving Crop Resistance to Abiotic Stress, First Edition.Edited by Narendra Tuteja, Sarvajeet Singh Gill, Antonio F. Tiburcio, and Renu Tuteja� 2012 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2012 by Wiley-VCH Verlag GmbH & Co. KGaA.
j1351

processes, loss in productivity, or inducing damaging effect on any of plantorgans/parts besides alteration in anatomical, biochemical, or molecular regulation.Abiotic stresses such as extreme temperatures (freezing, cold, and heat), wateravailability (drought and flooding), and ion toxicity (salinity, alkalinity, and heavymetals) are the major causes that adversely affect the plant growth and productivityworldwide [1–5]. It has been found that the relative decreases in potential yieldsassociated with abiotic stress factors vary between 54 and 82% [2, 6]. It is estimatedfrom the comparison of record yields and average yields for various crop plants thatcrops mainly attain only 20% of their genetic potential for yield due to various bioticand abiotic stress factors. Therefore, it is clear that there is an urgent need to increaseabiotic stress tolerance in plants. Increasing crop production is now the highestagricultural priority worldwide because of increasing world population. According tothe United Nation�s World Population Prospects Report, the world population isincreasing at an alarming rate of approximately 74 million people per year and isexpected to reach more than 9 billion by 2050 (http://www.un.org/esa/population/unpop.htm), whereas global food productivity is declining due to the negative effectsof various environmental stress factors.
Plants respond to various stress factors such as salinity, heat, cold, drought,excess water, heavy metal toxicity, wounding, excess light, nutrient loss, anaerobicconditions, and radiations through multifaceted molecular signaling pathways byperceiving and transducing the stress signal(s) through a cascade of molecularnetworks eventually leading to the expression of stress-related genes. Theseresponses at the molecular, cellular, physiological and biochemical levels enablethe plants to survive [7, 8]. These responses include effects of abiotic stresses onoverall growth and development, electrolyte leakage, osmolyte accumulation, andthe ion homeostasis [8]. The new molecular �omics� tools have opened up newperspectives in stress biology. The �omics� approaches such as genomics, prote-omics, metabolomics, and transcriptomics have direct potential for improvingabiotic stress tolerance in crop plants that require knowledge of stress responseat molecular level, which includes gene expression to protein or metabolite andits phenotypic effects. Integration of phenotypic, genomic, transcriptomic, prote-omic, and metabolomic data will enable accurate and detailed gene networkreconstruction.
The oleiferous Brassica play an important role in global agriculture and horti-culture. These crops contribute both to the economy and to the health of popula-tions around the world. Brassica is the third most important source of vegetable oilin the world after palm and soybean oil and is grown as an edible or an industrial oilcrop that is used as a source of edible protein, in much the same way as soybeanprotein [9]. Brassica is the Latin name of a genus that is taxonomically placed withinthe Brassicaceae (Cruciferae), which is one of the 10th most economicallyimportant plant families in the world. The major mustard oil-producing countriesinclude Canada, China, France, Germany, India, and the United Kingdom.According to a report of the United States Department of Agriculture (USDA),the world oilseed production was 397 metric ton in 2006–2007. Indian agriculture
1352j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance

contributes about 15 and 8% to the world�s total acreage under oilseed cultivationand production, respectively. However, the productivity in India is only 791 kg ha�1
compared to the world average of 1718 kg ha�1 [10]. About 90% of the total landunder oilseed cultivation in India is occupied by Brassica juncea [11]. Indianmustard (B. juncea L. Czern & Coss.), belonging to the same genus Brassica inthe Cruciferae family, is an agriculturally and economically important crop widelycultivated in Asia and Europe, and abiotic stress is a major constraint to itsproductivity. B. juncea is a dominant crop grown for human consumption ascondiments, fodder, and seed. The Brassica plant is characterized by a largenumber of broad, oblong-shaped leaves present in the lower layers of the plantaxis [12]. B. juncea is a more productive oilseed than B. napus in hot regions ofRussia, India, China, and Canada with somewhat unreliable rainfalls, whereas B.napus is the higher yielding species in more temperate, wetter regions. Oil of B.juncea is consumed in large quantity and the production ranks second among alloilseeds in India [13]. In India, B. juncea is used as oil-bearing crop and has beenstudied extensively [14, 15]. However, due to the presence of undesirable long-chain fatty acids such as ecosenoic acid (10%) and erucic acid (50%) in the seed oil,it becomes detrimental to human health. Erucic acid increases blood cholesterol,interferes with myocardial conductance, and shortens coagulation time [16].European Economic Commission has restricted cultivation of Brassica crop thatcontains more than 10% erucic acid content in their oil [17]. The discovery of thegenes for low erucic acid oil production in the seeds of Indian mustard began theconversion of this ancient crop to a canola-type oilseed for dry areas.
52.1.1Description of Indian Mustard
B. juncea (Indian mustard) belongs to the family Cruciferae. There are nearly 40different varieties of this yellowflowering plant that are classified by the botanists intothe genusBrassica ofwhichB. juncea is one of themajor oilseed crops of India. The oilcontent varies between 35 and 45% and the protein content is between 20 and 24%.The seed residue is used as cattle feed and in fertilizers. It is a high biomass cropcharacterized by oblong-shaped leaves.
52.1.2Origin of Indian Mustard
Studies suggest that Brassica evolved from the genus Sinapidendron of the Mioceneage through Diplotaxis–Erucastrum complex [18]. B. juncea is generally thought tohave originated in theMiddle East, whereasB. rapa andB. nigra species overlapped inthe wild, but Central Asia and China are suggested as the sites of primary origin [19].Hemingway [20], however, considers that B. juncea (L.) may also have arisen byindependent hybridization at secondary centers in India, China, and Caucasus, as B.nigra was widely used as the commercial spice from early times.
52.1 Introduction j1353

52.1.3Nomenclature
The oleiferous Brassica grown in India are divided into the following four groups:
1) Brownmustard:Commonlyknownas rai, raya, or laha (B. junceaL.Czern&Coss.)2) Sarson
a) Yellow sarson: B. campestris L. var. Sarson Prainb) Brown sarson: B. campestris L. var. Dichotoma Watt
3) Toria: lahi or maghi lahi (B. campestris L. var. Toria Duth)4) Taramira or Tara (Eruca sativa Mill.)
In addition, there are two other species, namely,B. nigraKoch. (Banarasi rai) andB.juncea var. Rugosa (Pahadi rai). These two species do not fall under any of the fourgroups. These are,moreover, grown to a limited extent.Mustard (B. juncea L. Czern&Coss.) is the dominant species grown in India [19].
52.1.4Botanical Description
Rape and mustard include annual herbs. Roots, in general, are long and tapering.Toria is more or less a surface feeder but Brown sarson bears long roots with limitedlateral spread enabling its successful cultivationunder drier conditions. The height ofthe stem varies from 45 cm (in some varieties of Toria) to 190 cm (in Yellow sarson).In Toria andBrown sarson, the branches arise at an angle of 30–40�. In Yellow sarson,the branches arise laterally at an angle of about 10–20� giving the plant a narrow andpyramidal shape. The inflorescence is a corymbose raceme. In the case of yellowsarson, the four petals are spread apart, whereas, in brown sarson and Toria, thepetals overlap ormay be placed apart, depending upon the cultivar. Theflowers bear ahypogynous ovary. In Brown sarson and Toria, the ovary is bicarpellary, whereas inYellow sarson, itmay also be tri- or tetracarpellary. The fruit is sliliqua. The sliliqua aretwo-valved, three-valved, or four-valved, depending upon the number of carpels in theovary. The flowers begin to open from 8:00 h and continue up to 12:00 h noon.
52.1.5Genomic Relationships of B. juncea
The relationships among the cultivated Brassica species were largely clarified bycytological work. There are six species of Brassica that merit attention for theireconomic importance. Among the six species, three are diploid, B. campestris(2n¼ 20, AA), B. nigra (2n¼ 16, BB), and B. oleraceae (2n¼ 18, CC), and the otherthree are amphidiploids,B. juncea (2n¼ 36, AABB),B.napus (2n¼ 38, AACC), andB.carinata (2n¼ 34, BBCC). The botanical and genomic relationships between these sixspecies may be represented in the form of a triangle usually known as triangle ofU [21] (Figure 52.1).
1354j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance

In this chapter, we have tried to explore the available literature on �omics�approaches such as genomics, transcriptomics, proteomics, and metabolomics inBrassica, themost important oilseed crop, for possible crop improvement and abioticstress tolerance.We also present the success obtained so far in improving the salinitytolerance in brassicas employing the traditional breeding approaches.
52.2Indian Mustard: Breeding Aspects under Salinity Stress
Limited efforts have been made for improving crop varieties for various abioticstresses following conventional selection and breeding programs. The complexmechanisms of abiotic stress tolerance, characterized by the expression of severalminor genes limit the rate and extent of progress in improving stress tolerance incrops through conventional breeding program. Furthermore, techniques employedfor selecting tolerant plants are time consumable and consequently expensive.Drought and salinity are the serious impediments in decreasing the growth andyield performance of different crop varieties. The availability of genetic resources
Figure 52.1 �Triangle of U� representing thegenomic relationships among six crop speciesof Brassica. Three diploid species are shown (B.rapa, B. nigra, and B. oleracea), which represent
the AA, BB, and CC genomes, respectively. Alsoshown are three tetraploid species (B. carinata,B. juncea, and B. napus), which are hybridcombinations of the basic genomes.
52.2 Indian Mustard: Breeding Aspects under Salinity Stress j1355

besides the creation and exploitation of novel variability forms the basis of conven-tional breeding approaches. Cultivars with improved performance under differentabiotic stresses have been developed through selection and evaluation in the targeteddrought-and salinity-affected areas. Concerted breeding efforts at Central SoilSalinity Research Institute have resulted in the development and release of salinitytolerant cultivars such as CS52, CS54, and CS56. These varieties have consistentlyperformed better in the salt-affected soils than the national checks, Varuna andKranti. Further studies have shown that amphipolyploid species, namely,B. napus,B.carinata, andB. juncea aremore tolerant toward salinity than the diploid brassicas. B.napus followed by B. carinata have been characterized as more salt tolerant,particularly in climatic zones with long cool periods. B. juncea, probably due to itsshorter growth period compared to B. napus and B. carinata, has proved morepromising in semiarid and arid climateswith shortwinters. Brassicas are by and largemore vulnerable to alkalinity than to salinity. Mustard genotypes have showntremendous variability in plant types. Vigorous growth during vegetative stage,medium plant height, early maturity, and better response to added inputs are someof the desired characteristics, which are associatedwith consistent yield performanceover an array of environments. Genotypes with medium plant height appear to bemore stable in varied environments. The exploitation of hybrid vigor, through the useofCMSand appropriate restorer, will go a longway in raising the production potentialof rapeseed and mustard in India.
Besides the yield potential and stability, germplasm is improved for traits con-tributing to increased productivity. Physiological characters such as nutrient useefficiency and harvest index need attention. The rapeseed–mustard crops producelarge biomass but fail to translocate it to the sink, resulting in low harvest index.Attempts should also be made to breed varieties for better response to low nitrogenapplication, which needs tailoring of the new plant type having less secondary andtertiary branching, reduced height, longer main shoot, more siliqua on main shoot,more number of seeds/siliqua, and higher 1000-seed weight with better nutrient useefficiency and capacity to transport assimilate to seeds.
Salinity stress causes yield decline in all the species and the genotypes. Thechallenge is to improve the genotypes with salt tolerance characteristics retaining theeconomic potential of the particular genotype. Stressful environments are often saidto increase the expansion of inbreeding depression. Plant�s behavior also varies withintensity of stress. Inbreeding depression occurs for survival under high stressconditions and for growth under lower stress levels. The workable strategy lies inselection for tolerant genotypes paying lesser penalty in terms of yield decline. A largenumber of germplasm needs to be evaluated under target environments to identifythe breeding material.
52.2.1Screening Methodology for Seedling Emergence
Under field salinity conditions, it has been observed that once a seedling emerges, itcontinues to growwith relatively lesser reductions in growth and yield. Accordingly, it
1356j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance

is imperative to know the salt tolerance potential of different Indianmustard genotypesfor seedling emergence by some rapid screening methodology under laboratoryconditions. For this, different genotypes have been evaluated under solution culturein laboratory, sand culture, and soil culture in pots to know their interrelationships.Variability in different genotypes with respect to their behavior under salinity stressmay be attributed to the differences in uptake of toxic ions alongwith the absorption ofwater by the seeds. Increase inNaþ uptake alongwith the decline in Kþ uptake undersalinity stress during germination has been reported by Sharma [22]. Furthermore,seedling emergence under solution culture and sand culture declined only due to theionic stress caused by salinity of the soil solution, whereas in soil culture, osmoticeffects play an associative role along with ionic effects. Accordingly, the higher declinein seedling emergence under soil culture may be associated with aggravation in ionicaccumulation caused by the increasing osmotic stress along with matric stress.Seedling emergence under soil culture was also observed to decline through post-germination salt injuries to hypocotyls at the time of their protruding through soil.
Application of piece-wise linear response model to the pooled data of sevengenotypes in different types of growing medium showed that threshold salinitylevel was highest in solution culture experiments (Ct¼ 14.15). It declined to 7.84 insand culture and further to 5.96 in soil culture experiments [23]. Similarly, the salinitylevel at which 50% reduction in seedling emergence occurred was also highest insolution culture (C50¼ 21.06), which declined to 15.96 in sand culture and to 12.31 insoil culture experiments. Along with the decline in Ct and C50 upon shifting fromsolution to sand and soil culture, the slope values increased from 7.25 in solutionculture to 9.30 in soil culture experiments. The regression equation for the pooleddata in three types of growing medium was computed to be Y¼� 3.8789 Xþ 111.417 (r2¼ 0.69). Furthermore, the regression equation for seedling emergenceunder different salinity levels in solution culture was Y¼� 2.1712 (X) þ 104.1922(r2¼ 0.38), while in sand culture, it was Y¼� 4.2505 (X) þ 114.4129 (r2¼ 0.87) andin soil culture the equation was Y¼� 4.6805 (X) þ 106.7743 (r2¼ 0.86), where X issalinity level and Y is seedling emergence.
Salinity levels in which the results for seedling emergence under solution or sandculture are typical representative of the results under soil culture were calculated.These values were computed where the two types of growingmedia recorded around50% of seedling emergence (C50). A significant positive correlation (r¼ 0.81) wasobserved between seedling emergence at ECiw 22 dSm
�1 in solution culture and ECe
12.8 dSm�1 in soil culture. Similar correlation between seedling emergence at ECiw
26 dSm�1 in solution culture with ECe 12.8 dSm�1 in soil culture was recorded to be
r¼ 0.92. Thus, using this method, a large number of genotypes can be tested forsalinity tolerance in a short period of time.
52.2.2Genetic Studies under Salt Stress
Genetic variability is the key to any crop improvement program, and the extent towhich the desirable characters are heritable is also important. For improving yield
52.2 Indian Mustard: Breeding Aspects under Salinity Stress j1357

and yield component characters, information on their genetic variability and theirinterrelationships in different characters is necessary. Partitioning the genotypiccorrelation coefficients of yield components into direct and indirect effects may helpestimate the actual contribution of an attribute and its influence through othercharacters. Sixty genotypes were evaluated for their adaptation under semiarid salinesoil conditions [24]. High variability was recorded for secondary branches/plant,siliqua/plant, 1000-seed weight, seed yield/plant, and seed yield/siliqua. Further lowvariability was recorded for seeds/siliqua and primary branches per plant. Seeds/plant were least affected by salinity. Seeds bearing siliqua/plant and seed yield perplot showed high genetic coefficients of variation (GCV). Heritability was very highfor the number of seeds/siliqua (99.74) and 1000-seed weight (61.44) and moderatefor siliqua length (39.72). Genetic advance was high for seed-bearing siliqua/plant(15.02) and plant height (11.89), followed by seeds/siliqua (5.69) and sterile siliqua/plant (8.33). Genetic advance as percent of mean was high for seeds/siliqua (45.73)and 1000-seed weight (42.91), followed by seed yield per plant. High heritabilityassociated with increased genetic advance indicated additive gene effects and greaterpossibility for improvement. Variability studies in 30 genotypes of Indian mustardindicated higher estimates of genetic and phenotypic coefficient of variation (PCV),heritability, and genetic advance for the number of primary branches and siliquae/plant on normal soil; and for seedling emergence, the number of siliquae on mainshoot, the number of secondary branches/plant, and seed yield on saline soil. Thenumber of siliquae/plant on normal soil and seedling emergence and plant height onsaline soil had significant positive association with seed yield. Seedling emergenceand plant height exhibited maximum direct, positive effects on seed yield on salinesoil.
Furthermore, genetic parameters for variability were also studied under alkalistress conditions in 19 genotypes of Indianmustard [25]. The GCV, PCV, heritability,and genetic advance as percent of mean were high for 1000-seed weight on normalsoil and for plant height, secondary branches per plant, the number of siliqua permain shoot length, and seed yield per plant on alkali soils. Correlation analysisrevealed that seed yield per plant has positive and significant association for primarybranches per plant and main shoot length on normal soil, and for plant height,secondary branches per plant and number of siliqua per main shoot length in alkalistress conditions. The path analysis showed that secondary branches per plant hadmaximum direct effect on seed yield under alkali stress conditions. These studiespoint out that the framing of selection criteria could be based on the number ofprimary branches per plant, main shoot length, and 1000-seed weight for normalconditions and secondary branches per plant, number of siliqua per main shootlength, and seed yield per plant under alkali stress conditions.
52.2.3Salt-Tolerant Varieties
Among different Brassica species, Indian mustard (raya) showed higher tolerance tosalinity/alkalinity stresses compared toGobhi sarson,Karan rai, and yellow and brown
1358j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance

sarson. Different genotypes of Indian mustard also showed differential tolerance tosaline and alkali stresses. Breeding efforts following conventional breedingappraoches at CSSRI, Karnal, led to the development of high-yield salt-tolerantgenotypes of Indianmustard, namely, CS52 in 1997, CS54 in 2005, andCS56 in 2008.These varieties were released by the Central Varietal Release Committee (CVRC),Government of India, for their cultivation in Punjab, Haryana, Rajasthan, andGujarat States of India (Table 52.1).
The salt-tolerant variety CS52 yields 20% higher in salt-affected soils compared tothe high-yield released varieties of Indian mustard at the national level. However, itsmaturity is longer by 1week compared to thewell-knownhigh-yield released varietiesof Indian mustard, that is, Varuna, Kranti, and Pusa Bold. Plant height of CS52 isaround 170–180 cm. This variety is more tolerant to insects and pests� attackcompared to other released varieties of Indian mustard. This variety accumulatesand compartmentalizes the toxic ions in the root part, hence restricting the concen-tration of toxic ions in leaves and stem. Further, the toxic ions get accumulated inlower leaves, which are shed ultimately reducing the effects of toxic ions on the plant.These processes help the plant to survive better under salt stress conditions. Anothercharacteristic of this variety is better adaptability under high temperature conditionsduring germination and seedling emergence stages.
These varieties can groweconomically in saline soils up to a soil salinity level of ECe
9 dSm�1 and in alkali soils up to pH 9.3.Within the salt-tolerant limits of this variety,CS52 and CS56 yield more than 1.6 ton ha�1, whereas CS54 can yield upto 1.9 tonha�1. By growing in saline soils and even irrigating with saline waters, 39% oilcontent has been determined from the seeds of these varieties. The latest releasedvariety CS56 has the additional advantage of delayed sowing upto 10th Novemberwithout significant reduction in seed yield besides the salt tolerance characteristics.
Table 52.1 Characteristics of Indian mustard varieties CS52, CS54, and CS56 released by CVRC forsalt-affected soils of India.
Variety CS52 CS54 CS56
Year of release 1997 2005 2008Plant height 170 cm 160 cm 202 cmMaturity duration 135 days 121 days 132 daysGrain type Medium Bold Medium1000-seed weight 4.0 g 5.3 g 4.4 gSalinity tolerance ECe 6–9 dS m�1 ECe 6–9 dS m�1 ECe 6–9 dS m�1
Alkali tolerance Upto pH 9.3 Upto pH 9.3 Upto pH 9.3Average seed yield 15–16 q ha�1 16–19 q ha�1 16–18 q ha�1
Recommendeddate of sowing
First fortnightof October
First fortnightof October
First fortnight of October(normal conditions) and upto10th November (late sown)
Recommended for Punjab, Haryana,and Rajasthan
Punjab, Haryana,and Rajasthan
Late-sown areas of the country(Punjab, Haryana, and Rajasthan)
52.2 Indian Mustard: Breeding Aspects under Salinity Stress j1359

52.3�Omics� Approaches for Abiotic Stress Tolerance
Plant research has the potential to contribute significantly to solve several of themostdaunting problems that our planet andwe face in the twenty-first century. Developingcrop plants with ability to tolerate abiotic stresses is need of the day that demandsmodern, novel strategies for a thorough understanding of plant�s response to abioticstresses. In particular, an array of innovative �omics� approaches such as genomics,proteomics, transcriptomics, and metabolomics are evolving at rapid pace, which isempowering the scientists to systematically analyze the genome at various levels andtheir effect on phenotypic variability. In contrast to the traditional approaches thatmostly focus on one or a few genes at a time, omics tools particularly genomicsallowed the use of important strategies such as genome-wide expression profilingthat is useful to identify genes associated with stress response. Furthermore,proteomics helped a lot to analyze the relationship between gene expression(transcriptomics) andmetabolism (metabolomics). Metabolomic studies, thus alongwith transcriptomics and proteomics, and their integration with systems biology, willlead to strategies to alter cellular metabolism for adaptation to abiotic stress condi-tions.Overall, the �omics� research approaches have produced copious data for livingsystems, which have necessitated the development of systems biology to integratemultidimensional biological information into networks and models. Applications ofsystems biology to plant science have been rapid and have increased our knowledgeabout circadian rhythms,multigenic traits, stress responses, and plant defenses, andhave advanced the virtual plant project. In the proceeding pages we will discuss indetail the �omics� approaches such as genomics, proteomics, and metabolomics forabiotic stress tolerance in mustard.
52.3.1Genomics Approaches and Abiotic Stress Tolerance in B. juncea
The application of genomics-type technologies is beginning to have an impact,enhancing our understanding of plant responses to abiotic stresses that interferewith the normal growth andmetabolism of plants. The ongoing genome-sequencingproject will give access to the required genes and will facilitate the analysis of theirexpression.
Abiotic stresses such as low temperature and high salinity adversely affect theseedling vigor and fertility ofB. juncea and consequent reduction in yield [26]. Studieson B. juncea [27, 28] have demonstrated the coordinated transcriptional regulation ofgenes involved in sulfur transport and assimilation and inGSHbiosynthesis. The Cdexposure, consequently, induces phytochelatin synthesis from glutathione (GSH).Zhu et al. [29, 30] overexpressed the c-glutamylcysteine synthetase gene fromEscherichia coli in B. juncea, resulting in increased biosynthesis of GSH and PCsand an increased tolerance to cadmium (Cd). A similar approach was taken withArabidopsis; c-glutamylcysteine synthetasewas expressed in both sense and antisenseorientations, resulting in plants with a wide range of GSH levels [31]. Flocco et al. [32]
1360j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance

tested the hypothesis that transgenic Indian mustard plants overexpressing ECS orGS, which have twofold levels of GSH and total nonprotein thiols, would haveenhanced tolerance to organic pollutants, including the PAH phenanthrene. Expo-sure of plants to organic pollutants significantly enhanced total nonprotein thiollevels in both wild-type and transgenic plants [32]. Flocco et al. [32] concluded thatGSH could be important for detoxification via conjugation to GSH, of many organicxenobiotics including phenanthrene, and that overexpression of enzymes involved inGSH biosynthesis offers a promising approach to create plants with the enhancedcapacity to tolerate not only heavy metals but also certain organic pollutants. Besidesthe genes mentioned above, a range of other stress-related proteins have beenemployed in raising transgenic plants with improved tolerance toward variousabiotic stresses as mentioned in Table 52.2.
52.3.2Proteomic Approaches and Abiotic Stress Tolerance in B. juncea
The simple paradigm, gene to mRNA to protein, belies the complexity of therelationship between a stretch of genomic DNA and its cognate protein products.The transcriptome analyses of gene expression at the mRNA level have contributedgreatly to our understanding of abiotic stress tolerance in plants.However, the level ofmRNA does not always correlate well with the level of protein, the key player in thecell [33]. Therefore, it is insufficient to predict protein expression level fromquantitative mRNA data. This is mainly due to posttranscriptional regulationmechanisms such as nuclear export and mRNA localization, transcript stability,translational regulation, and protein degradation. Proteome studies aim at thecomplete set of proteins encoded by the genome and thus complement the tran-scriptome studies. Aproteome constitutes all different proteins in a particular �body.�The body may be an organism, as in yeast proteomics [34], and can also be asubcellular particle such as an organelle [35, 36] or a protein complex [37].
52.3.3Metabolomics Approaches and Abiotic Stress Tolerance in B. juncea
Undoubtedly, transcriptomic and proteomic data are important steps in decipheringa complex biological process, but they are still insufficient since most biologicalprocesses are ultimatelymediated by cell metabolites. Metabolomics is considered toprovide a direct �functional readout of the physiological state� of an organism.Besides, alternative mRNA splicing, protein turnover rates, and posttranslationalmodifications that modulate protein activity imply that changes in the transcriptomeor proteome do not always correspond to alterations in the cell metabolome [38].Target analysis, metabolite profiling, and metabolic fingerprinting are differentconceptual approaches in metabolomics that can be used for a large range ofapplications, including phenotyping of genetically modified plants, substantialequivalence testing, determination of gene function, and monitoring responses tobiotic and abiotic stress. Metabolomics can therefore be seen as bridging the gap
52.3 �Omics� Approaches for Abiotic Stress Tolerance j1361
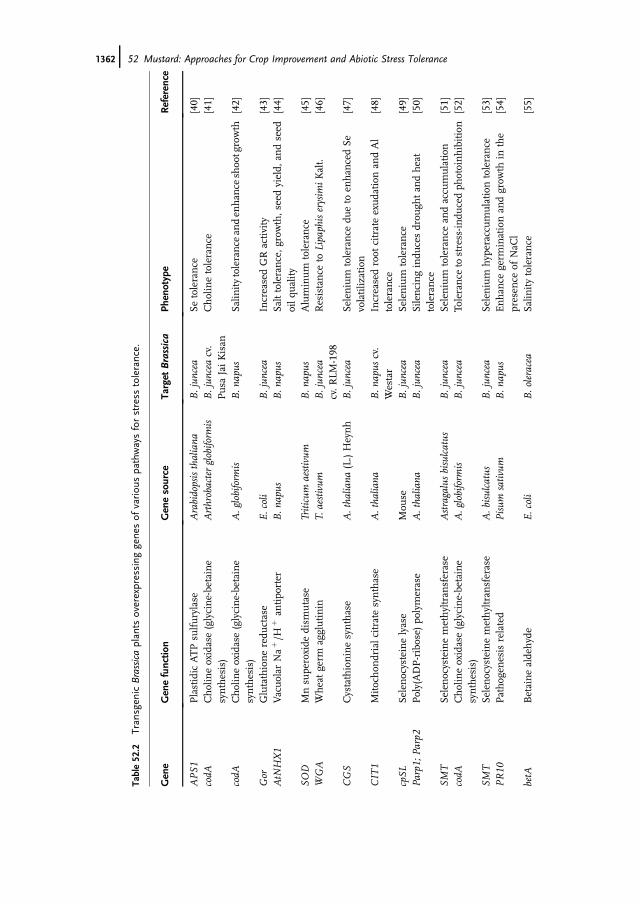
Table52.2
Tran
sgen
icBrassicaplan
tsoverexpressing
gene
sof
variou
spa
thwaysforstress
tolerance.
Gene
Genefunctio
nGenesource
Target
Brassica
Phenotype
Reference
APS1
Plastidic
ATPsulfurylase
Arabidopsisthaliana
B.jun
cea
Setolerance
[40]
codA
Cholineoxidase(glycine-betaine
synthesis)
Arthrobacterglobiform
isB.jun
ceacv.
Pusa
JaiKisan
Cholinetolerance
[41]
codA
Cholineoxidase(glycine-betaine
synthesis)
A.globiform
isB.n
apus
Salin
itytolerance
anden
han
ceshootgrowth
[42]
Gor
Glutathioneredu
ctase
E.coli
B.jun
cea
IncreasedGRactivity
[43]
AtN
HX1
Vacuolar
Naþ/H
þan
tipo
rter
B.n
apus
B.n
apus
Salttolerance,g
rowth,seedyield,
andseed
oilq
uality
[44]
SOD
Mnsuperoxide
dism
utase
Triticum
aestivum
B.n
apus
Aluminum
tolerance
[45]
WGA
Wheatgerm
agglutinin
T.aestivum
B.jun
cea
cv.R
LM-198
Resistance
toLipaphiserysim
iKalt.
[46]
CGS
Cystathioninesynthase
A.tha
liana
(L.)Heynh
B.jun
cea
Selenium
tolerance
dueto
enha
ncedSe
volatiliz
ation
[47]
CIT1
Mitochon
drialcitrate
synthase
A.tha
liana
B.n
apus
cv.
Westar
Increasedroot
citrateexudation
andAl
tolerance
[48]
cpSL
Selenocysteinelyase
Mou
seB.jun
cea
Selenium
tolerance
[49]
Parp1;P
arp2
Poly(ADP-ribose)
polymerase
A.tha
liana
B.jun
cea
Silencingindu
cesdrou
ghtan
dheat
tolerance
[50]
SMT
Selenocysteinemethyltran
sferase
Astragalusbisulcatus
B.jun
cea
Selenium
tolerance
andaccumulation
[51]
codA
Cholineoxidase(glycine-betaine
synthesis)
A.globiform
isB.jun
cea
Tolerance
tostress-in
ducedph
otoinhibition
[52]
SMT
Selenocysteinemethyltran
sferase
A.b
isulcatus
B.jun
cea
Selenium
hyperaccumulation
tolerance
[53]
PR10
Patho
genesisrelated
Pisum
sativum
B.n
apus
Enhan
cegerm
inationan
dgrow
thin
the
presen
ceof
NaC
l[54]
betA
Betainealdehyde
E.coli
B.oleracea
Salin
itytolerance
[55]
1362j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance
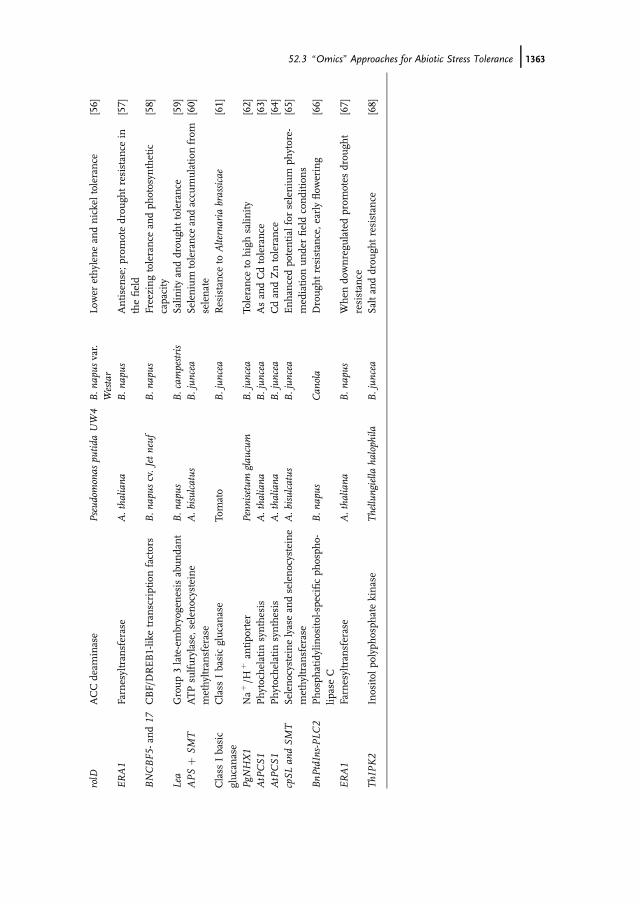
rolD
ACCdeam
inase
Pseud
omonas
putida
UW4
B.n
apus
var.
Westar
Lower
ethylen
ean
dnickeltoleran
ce[56]
ERA1
Farnesyltran
sferase
A.tha
liana
B.n
apus
Antisense;p
romotedrou
ghtresistan
cein
thefield
[57]
BNCBF5
-an
d17
CBF/DREB1-lik
etran
scriptionfactors
B.n
apus
cv.Jetneuf
B.n
apus
Freezingtolerance
andph
otosyn
thetic
capacity
[58]
Lea
Group3late-embryogenesisabundant
B.n
apus
B.cam
pestris
Salin
ityan
ddrou
ghttolerance
[59]
APSþ
SMT
ATPsulfurylase,selen
ocysteine
methyltran
sferase
A.b
isulcatus
B.jun
cea
Selenium
tolerance
andaccumulationfrom
selenate
[60]
Class
Ibasic
glucanase
Class
Ibasicglucanase
Tomato
B.jun
cea
Resistance
toAlternaria
brassicae
[61]
PgN
HX1
Naþ/H
þan
tipo
rter
Pennisetum
glau
cum
B.jun
cea
Tolerance
tohighsalin
ity
[62]
AtPCS1
Phytochelatin
synthesis
A.tha
liana
B.jun
cea
Asan
dCdtolerance
[63]
AtPCS1
Phytochelatin
synthesis
A.tha
liana
B.jun
cea
Cdan
dZntolerance
[64]
cpSL
andSM
TSe
lenocysteinelyasean
dselenocysteine
methyltran
sferase
A.b
isulcatus
B.jun
cea
Enhan
cedpo
tential
forselenium
phytore-
mediation
underfieldcondition
s[65]
BnP
tdIns-PLC
2Phosph
atidylinositol-specificph
osph
o-lip
aseC
B.n
apus
Can
ola
Droug
htresistan
ce,e
arlyflow
ering
[66]
ERA1
Farnesyltran
sferase
A.tha
liana
B.n
apus
When
downregu
latedprom
otes
drou
ght
resistan
ce[67]
ThIPK2
Inositol
polyph
osph
atekinase
Thellu
ngiella
halophila
B.jun
cea
Saltan
ddrou
ghtresistan
ce[68]
52.3 �Omics� Approaches for Abiotic Stress Tolerance j1363

between genotype and phenotype. Metabolic changes underpin plant developmentand responses to applied stresses, and that metabolic information reflects biologicalendpointsmore accurately than transcript or protein analysis. Hence, the only way tothe complete understanding of both gene function and molecular events controllingcomplex plant processes is to analyze the transcriptome, the proteome, and themetabolome in an integrative manner [39].
Acknowledgment
AP would like to thank the support of DBT, Govt. of India, for funding Brassicaresearch in his laboratory.
References
1 Mahajan, S. and Tuteja, N. (2005) Cold,salinity and drought stresses: an overview.Arch. Biochem. Biophys., 444, 139–158.
2 Khan, N.A. and Singh, S. (2008) PlantResponses to Abiotic Stresses, IKInternational, New Delhi.
3 Tuteja, N. (2007) Mechanisms of highsalinity tolerance in plants. MethothsEnzymol., 428, 419–438.
4 Purty, R.S., Kumar, G., Singla-Pareek,S.L., and Pareek, A. (2008) Invitedcontribution towards salinity tolerance inBrassica: an overview. Physiol. Mol. Biol.Plants, 14, 15–22.
5 Gill, S.S., Khan, N.A., Anjum, N.K., andTuteja, N. (2011) Amelioration ofcadmium stress in crop plants bynutrients management: morphological,physiological and biochemical aspects, inPlant Nutrition and Abiotic Stress ToleranceIII. Plant Stress 5 (Special Issue 1) (edsN.A.Anjum and F. Lopez-Lauri), pp. 1–23.
6 Bray, E.A., Bailey-Serres, J., andWeretilnyk, E. (2000) Responses to abioticstresses, in Biochemistry and MolecularBiology of Plants (eds W. Gruissem, B.Buchannan, and R. Jones), AmericanSociety of Plant Physiologists, Rockville,MD, pp. 1158–1249.
7 Nakashima, K., Ito, Y., and Shinozaki, K.Y.(2009) Transcriptional regulatorynetworks in response to abiotic stresses inArabidopsis and grasses. Plant Physiol.,149, 88–95.
8 Kumar, G., Purty, R.S., Sharma, M.P.,Singla-Pareek, S.L., and Pareek, A.(2009) Physiological responses amongBrassica species under salinity stressshow strong correlation withtranscript abundance for SOSpathway-related genes. J. PlantPhysiol., 166, 507–520.
9 Zhang, G.Q., Tang, G.X., and Zhou, W.J.(2003) A preliminary study on theinterspecific hybridization of B. compestrisand B. oleracea through ovary culture. Sci.Agric. Sin., 36, 1409–1413.
10 Damodaran, T. and Hegde, T.M. (2002)Oilseeds Situation: A StatisticalCompendium, DOR, Hyderabad, pp. 210–229.
11 Khan, N.A., Singh, S., Nazar, R., and Lone,P.M. (2007) The source–sink relationshipin mustard. Asian Aust. J. Plant Sci.Biotechnol., 1, 10–18.
12 Weiss, E.A. (1983)Oilseed Crops, LongmanInc, New York.
13 Chopra, V.L. and Prakash, S. (1991)Taxonomy, distribution and cytogenetics,in Brassica Oilseed in Indian Agriculture(eds V.L. Chopra and S. Prakash), VikasPub. House Pvt. Ltd., New Delhi,pp. 257–304.
14 Lionneton, E., Aubert, G., Ochatt, S., andMerah, O. (2004) Genetic analysis ofagronomic and quality traits in mustard(Brassica juncea). Theor. Appl. Genet., 109,792–799.
1364j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance

15 Oram, R.N., Kirk, J.T.O., Veness, P.E.,Hurlstone, C.J., Edlington, J.P., andHalsall, D.M. (2005) Breeding Indianmustard [Brassica juncea (L.) Czern.] forcold-pressed, edible oil production; areview. Aust. J. Agric. Res., 56, 581–596.
16 Renard, S. and Mcgregor, S. (1992)Antithrombogenic effect of erucic acidpoor rapeseed oil in the rats. Rev. Fr. CropCros., 23, 393–396.
17 Dhillon, S.S., Kumar, P.R., and Gupta, N.(1992) Breeding objectives andmethodology, in Breeding Oilseed Brassicas(eds K.S. Labana, S.S. Banga, and S.K.Banga), Narosa Publishing House,New Delhi, pp. 10–17.
18 G�omez-Campo, C. and Tortosa, M.E.(1974) The taxonomic and evolutionarysignificance of some juvenile characters inthe tribe Brassiceae (Cruciferae). Bot. J.Linn. Soc., 69, 105–112.
19 Prakash, S. (1980) Cruciferous oilseeds inIndia, in Brassica Crops and Wild Allies:Biology and Breeding (eds S. Tsunoda, K.,Hinata and C. Gomez-Campo), JapanScience Society Press, Tokyo, pp. 151–163.
20 Hemingway, J.S. (1976) Mustards, inEvolution of Crop Plants (eds N.W.Simmonds and J. Smart), Longman,London, pp. 82–86.
21 UN (1935) Genomic-analysis in Brassicawith special reference to the experimentalformation of B. napus and peculiar modeof fertilization. Jpn. J. Bot., 7, 389–452.
22 Sharma, P.C. (2003) Salt tolerance ofIndian mustard (Brassica juncea L.):factors affecting growth and yield. IndianJ. Plant Physiol., 8 (Special issue (NS)),368–372.
23 Sinha, T.S., Singh, D., Sharma, P.C., andSharma, H.B. (2003) Rapid screeningmethodology for salt tolerance duringgermination and seedling emergencein Indian mustard (Brassica juncea L.).J. Plant Physiol., 8 (Special issue (NS)),363–367.
24 Sinha, T.S. (1991) Genetic adaptation ofIndian mustard (Brassica juncea) to semi-arid saline soil conditions. Ind. J. Agric.Sci., 61, 251–254.
25 Sinha, T.S., Singh, D., Sharma, P.C., andSharma, H.B. (2002) Genetic variability,correlation and path coefficient studies
and their implications of selections of highyielding genotypes in Indian mustard(Brassica juncea L) under normal and sodicsoil conditions. Ind. J. Coastal Agric. Res.,20, 31–36.
26 Zhang, Y.X., Xu, J., Han, L., Wei, W.,Guan, Z.Q., Cong, L., andChai, T.Y. (2006)Highly efficient shoot regeneration andAgrobacterium-mediated transformationprotocol of Brassica juncea. Plant Mol. Biol.Rep., 24, 255–255.
27 Sch€afer, H.J., Haag-Kerwer, A., andRausch, T. (1998) cDNA cloning andexpression analysis of genes encodingGSH synthesis in roots of the heavy metalaccumulator Brassica juncea L: evidencefor Cd-induction of a putativemitochondrial c-glutamylcysteinesynthetase isoform. Plant Mol. Biol., 37,87–97.
28 Lee, S. and Leustek, T. (1999) The effect ofcadmiumon sulfate assimilation enzymesinBrassica juncea.Plant Sci., 141, 201–207.
29 Zhu,O.Y.L., Pilon-Smits, E.A.H., Jouanin,L., and Terry, N. (1999) Overexpression ofglutathione synthetase in Indian mustardenhances cadmium accumulation andtolerance. Plant Physiol., 119, 73–79.
30 Zhu, Y.L., Pilon-Smits, E.A.H., Jouanin,L., and Terry, N. (1999) Cadmiumtolerance and accumulation in Indianmustard is enhanced by overexpressingc-glutamylcysteine synthetase. PlantPhysiol., 121, 1169–1177.
31 Xiang, C., Werner, B.L., Christensen,E.M., andOliver,D.J. (2001) Thebiologicalfunctions of glutathione revisited inArabidopsis transgenic plants with alteredglutathione levels. Plant Physiol., 126,564–574.
32 Flocco, C.G., Lindblom, S.D., and Pilon-Smits, E.A.H. (2004) Overexpression ofenzymes involved in glutathione synthesisenhances tolerance to organic pollutantsin Brassica juncea. Int. J. Phyto., 6,289–304.
33 Yan, S.P., Zhang, Q.Y., Tang, Z.C., Su,W.A., and Sun, W.N. (2006) Comparativeproteomic analysis provides new insightsinto chilling stress responses in rice.Mol. Cell Proteom., 5, 484–496.
34 Washburn, M.P., Wolters, D., and Yates,J.R., 3rd (2001) Large-scale analysis of the
References j1365

yeast proteome by multidimensionalprotein identification technology. Nat.Biotechnol., 2001, 19, 242–247.
35 Prime, T.A., Sherrier, D.J., Mahon, P.,Packman, L.C., and Dupree, P. (2000) Aproteomic analysis of organelles fromArabidopsis thaliana. Electrophoresis, 21,3488–3499.
36 Peltier, J.B., Friso, G., Kalume, D.E.,Roepstorff, P., Nilsson, F., Adamska, I.,and van Wijk, K.J. (2000) Proteomics ofthe chloroplast: systematic identificationand targeting analysis of lumenal andperipheral thylakoid proteins. Plant Cell,12, 319–341.
37 Verma,R., Chen, S., Feldman,R., Schieltz,D., Yates, J., Dohmen, J., and Deshaies,R.J. (2000) Proteasome proteomics:identification of nucleotide sensitiveproteasome-interacting proteins, by massspectrometric analysis of affinity-purifiedproteasomes. Mol. Biol. Cell, 11,3425–3439.
38 Sumner, L.W., Mendes, P., and Dixon,R.A. (2003) Plant metabolomics: large-scale phytochemistry in the functionalgenomics era. Phytochem., 62, 817–836.
39 Dixon, R.A. (2001) Natural productsand plant disease resistance. Nature, 411,843–847.
40 Pilon-Smits, E.A.H., Hwang, S., Lytle,C.M., Zhu, Y., Tai, J.C., Bravo, R.C., Chen,Y., Leustek, T., and Terry, N. (1999)Overexpression of ATP sulfurylase inIndian mustard leads to increasedselenate uptake, reduction, and tolerance.Plant Physiol., 119, 123–132.
41 Prasad, K.V.S.K., Sharmila, P., Kumar,P.A., and Pardha Saradhi, P. (2000)Transformation of Brassica juncea (L.)Czern with bacterial codA gene enhancesits tolerance to salt stress. Mol. Breed., 6,489–499.
42 Huang, J., Hirji, R., Adam, L.,Rozwadowski, K.L., Hammerlindl, J.K.,Keller, W.A., and Selvaraj, G. (2000)Genetic engineering of glycinebetaineproduction toward enhancing stresstolerance in plants: metabolic limitations.Plant Physiol., 122, 747–756.
43 Pilon-Smits, E.A., Zhu, Y.L., Sears, T., andTerry, N. (2000) Overexpression ofglutathion reductase in Brassica juncea:
effects on cadmium accumulation andtolerance. Physiol. Plant., 110, 455–460.
44 Zhang,H.X.,Hodson, J.N.,Williams, J.P.,and Blumwald, E. (2001) Engineering salt-tolerant Brassica plants: characterizationof yield and seed oil quality in transgenicplants with increased vacuolar sodiumaccumulation. Proc. Natl. Acad. Sci. USA,98, 12832–12836.
45 Basu, U., Good, A.G., and Taylor, G.J.(2001) Transgenic Brassica napus plantsoverexpressing aluminium-inducedmitochondrial manganese superoxidedismutase cDNA are resistant toaluminium. Plant Cell Environ., 24,1269–1278.
46 Kanrar, S., Venkateswari, J., Kirti, P.B.,andChopra, V.L. (2002) Transgenic Indianmustard (Brassica juncea) with resistanceto the mustard aphid (Lipaphis erysimiKalt.). Plant Cell Rep., 20, 976–981.
47 Van Huysen, T., Abdel-Ghany, S., Hale,K.L., LeDuc, D., Terry, N., and Pilon-Smits, E.A.H. (2003) Overexpression ofcystathionine-synthase enhancesselenium volatilization in Brassica juncea.Planta, 218, 71–78.
48 Anoop, V.M., Basu,U.,McCammon,M.T.,McAlister-Henn, L., andTaylor, G.J. (2003)Modulation of citrate metabolism altersaluminum tolerance in yeast andtransgenic canola overexpressing amitochondrial citrate synthase. PlantPhysiol., 132, 2205–2217.
49 Garifullina, G.F., Owen, J.D., Lindblom,S.D., Tufan,H., Pilon,M., and Pilon-Smit,E.A.H. (2003) Expression of a mouseselenocysteine lyase in Brassica junceachloroplasts affects selenium toleranceand accumulation. Physiol. Plant., 118,538–544.
50 Block, M.D., Christoph, V., De, B.D., andMarc, C. (2005) Poly(ADP-ribose)polymerase in plants affects energyhomeostasis, cell death and stresstolerance. Plant J., 41, 95–106.
51 LeDuc, D.L., Tarun, A.S., Montes-Bayon,M., Meija, J., Malit, M.F., Wu, C.P.,AbdelSamie, M., Chiang, C.Y., Tagmount,A., deSouza, M., Neuhierl, B., Bock, A.,Caruso, J., and Terry, N. (2004)Overexpression of selenocysteinemethyltransferase in Arabidopsis and
1366j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance

Indian mustard increases seleniumtolerance and accumulation. PlantPhysiol., 135, 377–383.
52 Prasad, K.V.S.K. and Pardha Saradhi, P.(2004) Enhanced tolerance tophotoinhibition in transgenic plantsthrough targeting of glycinebetainebiosynthesis into the chloroplasts. PlantSci., 166, 1197–1212.
53 Danika, L.L.D., Tarun, A.S., Montes-Bayon,M.,Meija, J.,Malit,M.F.,Wu, C.P.,AbdelSamie, M., Chiang, C.Y., Tagmount,A., deSouza, M., Neuhierl, B., Bock, A.,Caruso, J., and Terry, N. (2004)Overexpression of selenocysteinemethyltransferase in Arabidopsis andIndian mustard increases seleniumtolerance and accumulation. PlantPhysiol., 135, 377–383.
54 Srivastava, S., Fristensky, B., and Kav,N.N.V. (2004) Constitutive expression of aPR10protein enhances the germination ofBrassica napus under saline conditions.Plant Cell Physiol., 45, 1320–1324.
55 Bhattacharya, R.C., Maheswari, M.,Dineshkumar, V., Kirti, P.B., Bhat, S.R.,and Chopra, V.L. (2004) Transformation ofBrassica oleracea var. capitatawith bacterialbetA gene enhances tolerance to saltstress. Sci. Horticul., 100, 215–227.
56 Stearns, J.C., Shah, S., Greenberg, B.M.,Dixon, D.G., and Glick, B.R. (2005)Tolerance of transgenic canola expressing1-aminocyclopropane-1-carboxylic aciddeaminase to growth inhibition by nickel.Plant Physiol. Biochem., 43, 701–708.
57 Yang,W., Jifeng, Y.,Monika, K.,Maryse, C.,Angela, S., Charlene,M., Tina,U., Carlene,S., Jiangxin, W., David, D.T., Peter, M., andHuang, Y. (2005) Molecular tailoring offarnesylation for plant drought toleranceand yield protection. Plant J., 43, 413–424.
58 Savitch, L.V., Allard, G., Seki, M., Robert,L.S., Tinker, N.A., Huner, N.P.A.,Shinozaki, K., and Singh, J. (2005) Theeffect of overexpression of two brassicaCBF/DREB1-like transcription factors onphotosynthetic capacity and freezingtolerance in Brassica napus. Plant CellPhysiol., 46, 1525–1539.
59 Park, B-.J., Liu, Z., Kanno, A., Kameya T.(2005) Increased tolerance to salt- andwater-deficit stress in transgenic lettuce
(Lactuca sativa L.) by constitutiveexpressionof LEA.PlantGrowthRegul.,45,165–171.
60 LeDuc, D.L., Abdelsamie, M., M�ontes-Bayon, M., Wu, C.P., Reisingerand, S.J.,and Terry, N. (2006) Overexpressing bothATP sulfurylase and selenocysteinemethyltransferase enhances seleniumphytoremediation traits in Indianmustard. Environ. Pollut., 144, 70–77.
61 Mondal, K.K., Bhattacharya, R.C.,Koundal, K.R., and Chatterjee, S.C. (2007)Transgenic Indian mustard (Brassicajuncea) expressing tomato glucanase leadsto arrested growth of Alternaria brassicae.Plant Cell Rep., 26, 247–252.
62 Rajagopal, D., Agarwal, P., Tyagi, W.,Singla-Pareek, S.L., Reddy, M.K., andSopory, S.K. (2007) Pennisetum glaucumNaþ/Hþ antiporter confers high level ofsalinity tolerance in transgenic Brassicajuncea. Mol. Breed., 19, 137–151.
63 Gasic,K.andKorban,S.S. (2007)TransgenicIndianmustard (Brassica juncea) plantsexpressing anArabidopsis phytochelatinsynthase (AtPCS1) exhibit enhancedAsandCd tolerance. PlantMol. Biol., 64, 361–369.
64 Gasic, K. and Korban, S.S. (2007)Transgenic Indian mustard (Brassicajuncea) plants expressing an Arabidopsisphytochelatin synthase (AtPCS1) exhibitenhanced As and Cd tolerance. Plant Mol.Biol., 64, 361–369.
65 Banuelos, G., LeDuc, D.L., Pilon-Smits,E.A.H., and Terry, N. (2007) TransgenicIndian mustard overexpressingselenocysteine lyase or selenocysteinemethyltransferase exhibit enhancedpotential for selenium phytoremediationunder field conditions. Environ. Sci.Technol., 41, 599–605.
66 Georges, F., Das, S., Ray, H., Bock, C.,Nokhrina, K., Kolla, V.A., and Keller, W.(2009) Over-expression of Brassica napusphosphatidylinositol-phospholipase C2 incanola induces significant changes ingene expression and phytohormonedistribution patterns, enhances droughttolerance and promotes early floweringand maturation. Plant Cell Environ., 32,1664–1681.
67 Wang, Y., Beaith, M., Chalifoux, M., Ying,J., Uchacz, T., Sarvas, C., Griffiths, R.,
References j1367

Kuzma, M., Wan, J., and Huang, Y. (2009)Shoot-specific down-regulation of proteinfarnesyltransferase (-subunit) for yieldprotection against drought in canola.Mol.Plant, 2, 191–209.
68 Zhu, J.Q., Zhang, J.T., Tang, R.J., Lv, Q.D.,Wang, Q.Q., Yang, L., and Zhang, H.X.
(2009) Molecular characterization ofThIPK2, an inositol polyphosphate kinasegene homolog from Thellungiellahalophila, and its heterologous expressionto improve abiotic stress tolerance inBrassica napus. Physiol. Plant., 136,407–425.
1368j 52 Mustard: Approaches for Crop Improvement and Abiotic Stress Tolerance





























