Embed Size (px)
Citation preview

In vivo Orientierung von Helicobacter
im Magenschleim von Maus und Gerbil
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie
der Ruhr-Universität Bochum
Angefertigt am Lehrstuhl für Physiologie
der Fakultät für Medizin
von
Manuela Konradt, geb. Stüben
aus Wuppertal
Bochum 2004

Für
meinen Mann Kai
und
unsere Tochter Anna Lena

Inhalt
1
INHALTSVERZEICHNIS
1. EINLEITUNG ............................................................................................. 3
Epidemiologie und Infektionsweg .................................................................. 4
Die Helicobacter pylori Erkrankung............................................................... 5
Therapie der Helicobacter Infektion .............................................................. 6
Die Virulenzfaktoren von Helicobacter.......................................................... 6
Die H. felis infizierte Maus und der H. pylori infizierte Gerbil als Tiermodelle ..................................................................................................... 10
Ziel der Arbeit ................................................................................................ 11
2. METHODEN ............................................................................................. 13
2.1 Versuchstiere und Haltung .................................................................... 13
2.2 Narkose ..................................................................................................... 14 2.2.1 Narkosetyp und Kreislaufsystem ............................................................................. 14 2.2.2 Messung der Atem- und Herzfrequenz .................................................................... 15
2.3 Operation .................................................................................................. 16 2.3.1 Eröffnung des Bauchraumes ................................................................................... 16 2.3.2 Fixierung des Magens ............................................................................................. 16 2.3.3 Peritoneale Spülung und Dialyse ............................................................................ 18 2.3.4 Eröffnung und Reinigung des Magens .................................................................... 18 2.3.5 Blutgasanalyse und Narkoseende............................................................................ 19
2.4 Beschreibung des Messfeldes .................................................................. 20
2.5 Entnahme von Schleimproben aus der Magenmucosa ........................ 21
2.6 Auswertung der Nanoschleimproben .................................................... 23 2.6.1 Ermittlung von Bakteriendichte und Abstand zur Gewebegrenze........................... 23 2.6.2 Motilitätsmessung.................................................................................................... 23
2.7 Versuchsbedingungen.............................................................................. 25 2.7.1 Zeitliche Einteilung ................................................................................................. 25 2.7.2 Kontrollbedingungen............................................................................................... 26 2.7.3 Phase veränderter Bedingungen ............................................................................. 27
2.8 Statistische Auswertung der Daten ........................................................ 28

Inhalt
2
3. ERGEBNISSE ........................................................................................... 29
3.1 Narkose ..................................................................................................... 29
3.2 Operation .................................................................................................. 30
3.3 Nanoprobennahme................................................................................... 30
3.4 Verteilung von Helicobacter im Magenschleim unter Kontrollbedingungen..................................................................................... 31
3.5 Orientierung von Helicobacter im Magenschleim unter veränderten Bedingungen ................................................................................................... 33
3.5.1 Der luminale pH und die luminalen Bicarbonat/CO2-Konzentrationen ................. 33 3.5.2 Der Harnstoff/Ammonium-Gradient ....................................................................... 34 3.5.3 Der pH/Bicarbonat/CO2-Gradient .......................................................................... 36
3.6 Messung des epithelnahen pH in vitro ................................................... 41
3.7 Zusammenfassung der Ergebnisse......................................................... 44
3.8 Ergänzungen............................................................................................. 46 3.8.1 Blutgaswerte............................................................................................................ 46
4. DISKUSSION ............................................................................................ 47
Der Zusammenhang von Bicarbonat, CO2 und pH.................................... 48
Wie registriert Helicobacter den pH? .......................................................... 49
Bakteriendichte in vivo .................................................................................. 50
Triple-Therapie und Orientierung von Helicobacter ................................. 51
LITERATURLISTE ......................................................................................... 52
ZUSAMMENFASSUNG .................................................................................. 64
SUMMARY........................................................................................................ 67
DANKSAGUNGEN........................................................................................... 69
LEBENSLAUF .................................................................................................. 70
ERKLÄRUNG ................................................................................................... 72

Einleitung
3
1. EINLEITUNG
Helicobacter ist ein gram-negatives,
mikroaerophiles, spiralförmiges Bakterium, welches
den Magenschleim besiedelt. Es besitzt 4-6 unipolare
Flagellen. Das 1,6 Mb große Genom von Helicobacter
pylori wurde 1997 als eines der ersten Genome
sequenziert (Tomb, 1997).
1983 gelang es erstmals Warren und Marshall, in
in vitro Kulturen Helicobacter pylori1 aus
Magenbiopsien anzuzüchten. Anhand dieser Kulturen
konnte gezeigt werden, dass H. pylori ein
entscheidender Pathogenitätsfaktor bei der Entstehung
von Magen- und Duodenalgeschwüren ist (Warren,
1983).
Die Helicobacter pylori Infektion ist heute als eine der häufigsten humanen
Infektionskrankheiten mit teils lebensbedrohlichen Folgeerkrankungen erkannt. Die Infektion
führt immer zu einer chronischen Gastritis, die in ca. 80 % der Fälle ohne subjektive
Beschwerden (Fiocca, 1989; Fiocca, 1992) verläuft. Typische Beschwerden einer Gastritis
sind wiederholt auftretende kolikartige Magenschmerzen sowie Unverträglichkeit
verschiedener Nahrungsmittel. Folgen der Langzeitinfektion können Magen- und
Duodenalgeschwüre sein (Dixon, 1990; Fiocca, 1989), außerdem besteht eine hohe Prävalenz
im Zusammenhang mit der Entstehung von MALT2-Lymphomen (Hussell, 1993;
Wotherspoon, 1993) sowie Magenkarzinomen (Parsonnet, 1991; Forman, 1991; The Eurogast
study group, 1993; Suerbaum, 2002). Dies hatte zur Folge, dass H. pylori 1994 von der World
Health Organisation (WHO) als definitives Karzinogen eingestuft wurde (IARC, 1999).
1 H. pylori wurde anfangs der Gattung Campylobacter zugeordnet, die Umgruppierung bzw. die Gattungsneubildung fand erst 1989 (Goodwin, 1989) statt. 2 MALT = Mucosa associated lymphoid tissue
1 µm
Abb. 1: Elektronenmikroskopische Aufnahmevon H. pylori. Mit freundlicher Genehmigung von PDDr. C. Josenhans (Institut für Hygieneund medizinische Mikrobiologie,Universität Würzburg).

Einleitung
4
Neben der humanpathogenen Spezies hat man inzwischen bei vielen Tierarten, vorwiegend
bei Wirbeltieren (Fox, 1990; Lee, 1988; Eaton, 1993; Bronsdon, 1991), selbst bei Vögeln
(Dewhirst, 1994) und Meeressäugern (Harper, 2002), artspezifisch infektiöse Helicobacter
Stämme nachweisen können. Zur Zeit sind mindestens 30 Spezies des Genus Helicobacter
bekannt und die Zahl weiterer Arten steigt kontinuierlich
an. Die einzelnen Helicobacter Spezies unterscheiden sich
mitunter stark in ihrer Morphologie und Biochemie, z.B.
besitzt H. felis, dessen Ursprungswirt Katzen und Hunde
sind, bipolare Flagellen und ist fast dreimal so groß wie
H. pylori.
Das Hauptaugenmerk der Forschung ruht jedoch auf
dem beim Menschen gefundenen Erreger H. pylori.
Obwohl H. pylori seit der ersten Kultivierung 1983 einer
der am gründlichsten untersuchten bakteriellen
Krankheitserreger ist, sind viele wesentliche Details des
Infektions- und des Krankheitsverlaufes nach wie vor noch
nicht ausreichend geklärt.
Epidemiologie und Infektionsweg
Weltweit sind mehr als eine Milliarde Menschen, also rund ein Fünftel der
Weltbevölkerung, mit Helicobacter pylori infiziert (The Eurogast study group, 1993). In
Industrieländern liegt die Durchseuchungsrate bei 20–50 %, während in Entwicklungsländern
die Infektionsdichte mitunter bei über 90 % der Bevölkerung liegt (Graham, 1991; Dixon,
1995; Megraud, 1989).
Die Ansteckung erfolgt meist im Kindesalter (Mitchell, 1992), wenn der soziale Kontakt
mit der Familie besonders eng ist. Die vergleichsweise hohe Durchseuchungsrate in den
Drittweltländern legt nahe, dass eine der wichtigsten Ursachen für ein erhöhtes
Infektionsrisiko das sozioökonomische Umfeld der Kindheit (Mendall, 1992) ist. Vor allem
beengte Wohnverhältnisse bei unzureichend hygienischen Verhältnissen sowie hohe
Geschwisterzahlen kennzeichnen die Risikofaktoren (Malaty, 1994; Malaty, 1999; Mendall,
1992).
1 µm
Abb. 2: Elektronenmikroskopische Aufnahmevon H. felis. Mit freundlicher Genehmigung von PDDr. C. Josenhans (Institut für Hygieneund medizinische Mikrobiologie,Universität Würzburg).

Einleitung
5
Die Übertragung von Helicobacter findet vorwiegend innerhalb der Familie
wahrscheinlich oral-oral von der Mutter auf das Kind (Rothenbacher, 2002) oder von Kind zu
Kind statt (Han, 2000). Im Mundraum sowie im Stuhl konnte genetisches Material von
Helicobacter nachgewiesen werden (Parsonnet, 1999), was zusätzlich eine fäkal-orale
Ansteckung nicht ausschließt.
Die orale Aufnahme erfordert, dass Helicobacter im Magenlumen dem Angriff durch
Säure und Pepsin widersteht, sich im Magenlumen orientiert und gezielt in den Magenschleim
eindringen kann.
Nach dem heutigen Kenntnisstand können andere Spezies, Nahrungsmittel oder Wasser als
Ansteckungsquellen weitgehend ausgeschlossen werden (Goodman, 1995).
Die Helicobacter pylori Erkrankung
Die im Kindesalter erworbene, akute Helicobacter pylori Primärinfektion, erzeugt meist
keine, bzw. uncharakteristische Krankheitssymptome, und ist daher im Vergleich zur
chronischen H. pylori Gastritis schwieriger zu diagnostizieren (Ernst, 1999).
Infolge einer chronischen H. pylori Infektion kommt es fast immer zu einer entzündlichen
Veränderung der Magenschleimhaut (Gastritis) (Marshall, 1986), welche unbehandelt
lebenslang oder bis zur Entwicklung einer vollständigen Mucosa-Atrophie bestehen bleibt, da
der Keim durch das Immunsystem des Wirtes nicht entfernt werden kann (Suerbaum, 2002).
Eine chronische Infektion kann wahrscheinlich zwei unterschiedliche Verläufe nehmen. Im
ersten Fall siedelt sich Helicobacter pylori vorwiegend im Antrum an. Dadurch kommt es
meist zu einer Hypergastrinämie mit erhöhter Säuresekretion, deren Folge Ulcera duodeni
oder ventriculi sein könnten (Evers, 1989). Im zweiten Fall erstreckt sich die Infektion
vorwiegend auf die Corpusregion des Magens. Nach langjähriger Infektion findet man
atrophische Metaplasien sowie eine bis zur Achlorhydrie verminderte Säuresekretion. Ein
solcher Verlauf kann zur Entwicklung eines Karzinoms führen (Bebelman, 1989).

Einleitung
6
Therapie der Helicobacter Infektion
Infizierte Patienten können mit einer Kombination zweier Antibiotika und einem
Säureblocker, über einen Zeitraum von 7 bis 10 Tagen verabreicht, behandelt werden (Triple-
Therapie) (Axon, 1996). Die alleinige Verabreichung der Antibiotika oder des Säureblockers
ist jedoch nahezu unwirksam. Deshalb geht man davon aus, dass erst die Gabe des
Säureblockers im Magenschleim das entsprechende Milieu schafft, welches eine
Antibiotikawirkung möglich macht. Bisher ist jedoch noch unklar, auf welchem Weg
Antibiotika und Säureblocker zusammenwirken.
Die Erfolgsquoten der Eradikation liegen bei 80-90 % (Labenz, 1995). Die Compliance der
Patienten spielt hierbei eine wichtige Rolle (Ruggiero, 2002), da die hohen Antibiotikadosen
mit Nebenwirkungen, vor allem Übelkeit, verbunden sind. Der Therapieerfolg wird außerdem
durch die ansteigende Resistenzentwicklung von Helicobacter gegen einige Antibiotika
(Debets-Ossenkopp, 1996) gefährdet.
Die Virulenzfaktoren von Helicobacter
Die Virulenzfaktoren von Helicobacter beschreiben die Eigenschaften des Bakteriums, die
wichtig sind für die initiale Kolonisation des Magens, für die Persistenz im Magenschleim
sowie für die gewebeschädigende Wirkung auf den Wirt. Helicobacter muss, um sich in
seiner ökologischen Nische ansiedeln zu können, welche die ersten 25 µm des epithelnahen
Magenschleims umfasst, zuerst das Magenlumen und die Schleimschicht durchdringen. Dabei
helfen eine ausgeprägte Motilität sowie die chemotaktischen Eigenschaften. Auch bei der
dauerhaften Besiedlung des Magenschleims, bei der sich das Bakterium fortlaufend
orientieren muss, sind Motilität und Chemotaxis essentiell. Ein weiterer essentieller Faktor für
die initiale und fortdauernde Besiedlung des Magenschleims ist die Urease. Motilität,
Chemotaxis und Urease werden im Folgenden genauer erläutert, da diese entscheidend für die
Orientierung der Bakterien im Magenschleim verantwortlich sind.

Einleitung
7
Motilität
Der kontinuierliche Schleimfluss im Magen erfordert von Helicobacter eine ausgeprägte
Motilität, um sich gezielt fortbewegen zu können. Die Ausbildung von 6-8 unipolaren
Geißeln (Flagellen), welche von einer membranartigen Hülle umgeben sind und sich an einem
Pol des Bakteriums finden, stellt ein charakteristisches morphologisches Merkmal von
H. pylori dar (Geis, 1993; Josenhans, 1995). Die Flagellenhülle, die aus einer
Lipopolysaccharidschicht besteht, schützt die säureempfindlichen Filamente vor dem
schädlichen Einfluss der Magensäure (Suerbaum, 1995) und ermöglicht es Helicobacter, sich
auch in der zähen Mucusschicht des Magenschleims fortzubewegen. Mutanten, bei denen die
Flagellen in ihrer Funktion eingeschränkt oder nicht vorhanden sind, verfügen über keinerlei
Möglichkeit zu infizieren (Eaton, 1996; Josenhans, 2002; Ottemann, 2002). Die Flagellen
erlauben es Helicobacter, sich rotierend um die Längsachse mit einer Geschwindigkeit von
30 µm pro Sekunde fortzubewegen (Karim, 1998).
Chemotaxis
In allen bekannten beweglichen Bakterien ist die Motilität eng an eine chemotaktische
Signalkaskade gebunden, welche es den Bakterien erlaubt, sich auf Lockstoffe hin zu
bewegen, oder sich von Reizstoffen zu entfernen.
Das chemotaktische Verhalten von Helicobacter ist zusammen mit seiner ausgeprägten
Motilität der wichtigste Faktor, welcher die Besiedlung des Magenschleims erlaubt.
Bisher wurden Untersuchungen zur Chemotaxis von Helicobacter nur in vitro durchgeführt
(Kihara, 1981; Pittman, 2001; Yoshiyama, 1999), wobei Harnstoff und Bicarbonat als
chemotaktische „attractants“ (Lockstoffe) für H. pylori beschrieben wurden (Mizote, 1997).
Über den genauen Verlauf der Signalkaskade besteht bisher noch Unklarheit. Aus
genetischen Homologiestudien weiß man, dass die chemotaktische Maschinerie von
Helicobacter, in weiten Teilen, der anderer gut untersuchter chemotaktischer Eubakterien
ähnelt (Taggart, 1979). Helicobacter besitzt drei klassische membrangebundene
Chemorezeptoren, sowie einen mutmaßlichen cytoplasmatischen Chemorezeptor (HP599)
(Tomb, 1997), welche das äußere Milieu analysieren. Über eine intrazelluläre, molekulare
Kettenreaktion werden die Informationen dann an den Flagellenmotor weiter vermittelt.
Einige an dieser Signalkaskade beteiligten Proteine sind beschrieben. Es handelt sich hierbei

Einleitung
8
um die rezeptorgekoppelte Autokinase CheA, das Verbindungsglied CheW, dessen genaue
Funktion nicht bekannt ist (Pittman, 2001), sowie CheY, welches direkt mit dem
Flagellenmotor interagiert (Rotter, 1979; Alm, 1999; Foynes, 2000; Bren, 2000).
Was genau von der Erkennung eines Lockstoffes hin bis zur gerichteten Bewegung bei
Helicobacter abläuft, ist bisher nicht geklärt, sicher weiß man aber, dass sich cheY- und
cheA-Mutanten durch eine andere Bewegungsart und Virulenz auszeichnen (Foynes, 2000).
Wie Helicobacter seinen natürlichen Lebensraum im Magenschleim erkennt, ist bisher
ebenfalls nicht bekannt. Helicobacter ist jedoch in der Lage, nicht nur seine ökologische
Nische zu erkennen, sondern diese trotz des dort herrschenden kontinuierlichen
Schleimflusses dauerhaft zu besiedeln. Das Bakterium orientiert sich also permanent neu im
Magenschleim und korrigiert seine Position mit Hilfe seiner ausgeprägten Motilität. Somit
muss die für Helicobacter notwendige Orientierungshilfe ein permanent im Magenschleim
vorliegender Gradient sein.
Urease
Essentiell für die Fähigkeit von Helicobacter pylori, sowie alle übrigen Helicobacter
Spezies, den Magenschleim zu kolonisieren, ist das von ihnen produzierte 550 kDa Enzym
Urease (Mobley, 1991; Mobley, 1995). Die Urease katalysiert die Spaltung von Harnstoff in
Ammoniak und Kohlendioxid und findet sich sowohl intrazellulär als auch im
periplasmatischen Raum. Die intrazelluläre Urease sorgt dort für einen neutralen pH (Scott,
2002; Ferrero, 1991). Urease-defiziente Mutanten können auch nach vorangegangener
Neutralisierung des Magenlumens nicht kolonisieren (Wirth, 1998). Somit müssen der Urease
noch weitere bisher nicht bekannte Funktionen zukommen. Dem bei der Harnstoff-Spaltung
entstehenden extrazellulären Ammonium wird zusätzlich ein zytotoxischer Effekt auf die
Magenepithelzellen zugesprochen, welcher bisher allerdings nur in hohen Dosen in vitro
nachgewiesen werden konnte (Tsujii, 1992; Sommi, 1996). Ferner wird der Urease eine bisher
unverstandene Rolle in der chemotaktischen Motilität von H. pylori zugesprochen
(Nakamura, 1998).
Der starke Umsatz von Harnstoff wird zur Diagnose einer H. pylori Infektion genutzt.
Beim C13-Harnstoff-Atemtest wird C13-markierter Harnstoff verabreicht und durch die Urease
umgesetztes C13O2 in der Ausatemluft der Patienten nachgewiesen (Graham, 1987).

Einleitung
9
Adhärenz
H. pylori ist in der Lage, engen Kontakt mit den Magenepithelzellen einzugehen. Das für
die Bindung an das Lewisb-Antigen der Epithelzellen notwendige bakterielle Adhäsin ist
bereits gut beschrieben (Wirth, 1996; Borén, 1993; Ilver, 1998). Verschiedenen anderen
Membranstrukturen von H. pylori wird ebenfalls eine Rolle bei der Adhärenz zugesprochen,
was aber noch nicht eindeutig geklärt ist (Odenbreit, 2002; Tomb, 1997). Die Adhärenz von
H. pylori an das Magenepithel ist nicht essentiell für eine Infektion, führt aber zu einer
spezifischen Immunantwort (Kavermann, 2003), wodurch eine Vielzahl von Signalkaskaden
ausgelöst werden. Andere Helicobacter Spezies wie zum Beispiel H. felis adhärieren nicht,
die Signalkaskaden bleiben somit größtenteils aus.
Cytotoxine
Das vacuolisierende Cytotoxin VacA wird von Helicobacter produziert und ruft in den
Epithelzellen eine Vakuolisierung des Zytoplasmas hervor, was im Tierexperiment
Schleimhautläsionen und Ulzera induzieren kann (Ghiara, 1995). Das 87 kDa große Protein
erfährt durch Säureaktivierung (Cover, 1997) eine Konformationsänderung, die es stabil
gegen Angriffe von Pepsin und Säure macht. Die Sekretion des Cytotoxins korreliert mit dem
Vorhandensein einer genetisch benachbarten Pathogenitätsinsel (PAI) cagA. Diese PAI wird
von einer Gruppe von 29 Genen gebildet und ist nicht bei allen Stämmen vorhanden (Censini,
1996). Die Entdeckung dieses Genabschnitts geht auf das stark immunogene Protein CagA
zurück, gegen das bei jedem Infizierten Antikörper gefunden werden.
Man teilt die H. pylori Stämme in CagA/VacA-positiven (Typ I) und CagA/VacA-
negative (Typ II) Stämme ein (Atherton, 1995; Blaser, 1996). Infektionen mit Typ I Stämmen
sind signifikant häufiger mit Ulkuskrankheiten, Karzinom und MALT-Lymphomen assoziiert
als Infektionen mit Typ II Stämmen (Parsonnet, 1997; Miehlke, 2001). Die genauen
Funktionen aller in diesem Abschnitt lokalisierten Gene sind noch nicht geklärt; einige Gene
kodieren wahrscheinlich für die Bildung eines bakteriellen Sekretionsapparates, der an der
Einschleusung von CagA sowie anderer bakterieller Proteine in die Wirtszellen beteiligt ist.
Die PAI ist nicht essentiell für eine Helicobacter Infektion. H. felis z.B. fehlt dieser
Genabschnitt komplett. Die mit H. felis infizierte Maus unterscheidet sich somit durch
geringere epitheliale Interaktion vom humanen Infektionsbild.

Einleitung
10
Die H. felis infizierte Maus und der H. pylori infizierte Gerbil als Tiermodelle
In der vorliegenden Arbeit wurden mit Helicobacter felis infizierte Mäuse und mit
Helicobacter pylori infizierte Mongolische Gerbils als Tiermodelle verwendet.
H. felis wurde aus Katzen isoliert (Lee, 1988) und kann auch in Mäusen eine stabile
Infektion hervorrufen (Lee, 1990; Enno, 1995), ist aber nicht in der Lage, an
Magenepithelzellen zu adhärieren. Außerdem fehlt H. felis die Pathogenitätsinsel cagA. Mit
H. felis infizierte Mäuse entwickeln nach ca. 8 Wochen eine leichte Gastritis. Eine Infiltration
der Mucosa mit neutrophilen Granulozyten ist ebenfalls zu beobachten. Bei einer
Langzeitinfektion wurde nach 22 bis 26 Monaten die Entstehung von lymphalen Tumoren, die
dem humanen MALT-Lymphom ähneln, beschrieben (Enno, 1995). Eine stabile Infektion
mit H. pylori in Mäusen wurde erstmals 1997 etabliert (Lee, 1997).
Mit H. pylori infizierte Gerbils zeigen schon
nach 2 Wochen eine deutliche Infiltration der
Mucosa mit neutrophilen Granulozyten
(Matsumoto, 1997; Hirayama, 1996), nach 3
Monaten können gastrische Geschwüre sowie
Atrophien in der antralen Mucosa nachgewiesen
werden (Honda, 1998; Ikeno, 1999). Nach langer
Infektion wurde die Entstehung von Karzinomen
beschrieben (Watanabe, 1998).
Der luminale pH, die Histologie des Magens sowie der Verlauf der chronischen Infektion
mit H. pylori, sind der des Menschen sehr ähnlich. Es wurde mit dem H. pylori infizierten
Gerbil somit ein Tiermodell etabliert, welches eine gute Übertragbarkeit von bestimmten
Resultaten auf den Menschen erlaubt.
Abb. 3: Mongolischer Gerbil (Meriones unguiculatus).

Einleitung
11
Ziel der Arbeit
Helicobacter besiedelt die Magenschleimhaut. Man findet ihn in der Schleimregion
oberhalb des Epithels, dem epithelnahen Schleimsegment (Schreiber, 1999), in welchem ein
pH zwischen 5 und 7 herrscht (Ross, 1981; Schreiber, 1997).
1999 wurde von Schreiber und Stüben et al. für die mit H. felis infizierte Maus in vivo
beschrieben (Schreiber, 1999), dass sich 95 % der Bakterien in einer epithelnahen
Schleimschicht von 25 µm Dicke aufhalten. Sie nehmen dort einen mittleren Abstand von
16 µm zum Epithel ein. Die Magenschleimschicht der Maus umfasst etwa 100 µm und lässt
sich aufgrund ihrer unterschiedlichen mikroskopischen Transparenz in einen luminalen und
einen epithelnahen Anteil gliedern. Die ersten 25 µm oberhalb des Epithels kann man als die
epithelnahe Region bezeichnet.
H. felis ist im epithelnahen Schleim hoch motil und bewegt sich mit einer mittleren
Geschwindigkeit von 30 µm/sec parallel zur Gewebeoberfläche. In luminalen
Schleimbereichen, in denen der pH unter 4,5 absinkt, findet man keine motilen Bakterien.
Helicobacter lebt in seinem Habitat zwischen zwei Fronten. Das Bakterium kann weder
durch das Epithel dringen, wahrscheinlich weil ihm dort die Immunzellen gefährlich werden
würden, noch kann es in den lumenseitigen Schleim wandern, weil dort die Einwirkung von
Säure und Pepsin tödlich wären.
Bisher ist nicht bekannt, wie es Helicobacter gelingt, den epithelnahen Schleim als sein
Habitat zu erkennen und diesen dauerhaft zu besiedeln. Helicobacter muss, hat er den Magen
erst einmal kolonisiert, gegen die Immunantwort des Magens sowie gegen die Sekretion des
Magenschleims, die zu einem kontinuierlichen Schleimfluss Richtung Lumen führt,
„schwimmen“.
Um seine Position im Magenschleim zu halten, benötigt er also neben seiner hohen
Motilität auch einen ständig gleichbleibenden chemischen Gradienten zur Orientierung.

Einleitung
12
Drei chemische Gradienten sind im Magenschleim beschrieben und kämen für die
chemotaktische Orientierung von Helicobacter in Frage:
• der pH-Gradient
Er zeichnet sich durch eine niedrigen pH im Magenlumen und einen fast neutralen pH
(5-7) am Gewebe aus (Schreiber, 1997; Talley, 1992; Schreiber, 2000).
• der Bicarbonat/CO2-Gradient
Er zeichnet sich durch äquimolare Konzentrationen von Bicarbonat und CO2 (bei
einem pH um 6) am Gewebe und hohe CO2- sowie niedrige
Bicarbonatkonzentrationen (bei saurem pH) im Lumen aus (Rees, 1982; Rune, 1969).
• der Harnstoff/Ammonium-Gradient
Der Harnstoff/Ammonium-Gradient wird durch die bakterielle Urease generiert, die
im epithelnahen Schleim Harnstoff in CO2 und Ammoniak umwandelt, was zur
Bildung von Ammonium führt (Kim, 1990). Man findet somit Richtung Lumen eine
abnehmende Harnstoff- und eine zunehmende Ammoniumkonzentration.
Bisher wurde das chemotaktische Verhalten von Helicobacter ausschließlich in vitro
(Kihara, 1981; Pittman, 2001) untersucht. In dieser Arbeit soll das chemotaktische Verhalten
von Helicobacter in vivo beschrieben werden.
Dazu werden die oben beschriebenen Gradienten bei H. felis infizierten Mäusen und
H. pylori infizierten Gerbils gezielt verändert. Aus dem Magen der narkotisierten Tiere
werden Schleimproben entnommen und untersucht, ob und inwieweit sich die Änderungen
auf die Orientierung von Helicobacter im Magenschleim auswirken. Die Probenentnahme
erfolgt mit einem von Schreiber und Stüben et al. entwickelten Nanoprobennahmesystem
(Schreiber, 1998), welches es ermöglicht, während eines Experimentes mehrere
Schleimvolumina im Nanoliterbereich zu entnehmen, ohne die kontinuierliche Schleimschicht
des Magens zu zerstören. Die Mucus und Mucosa enthaltenden Proben werden innerhalb von
Sekunden gewonnen und sofort abgekühlt. Auf diese Weise lässt sich die Position von
Helicobacter in Bezug auf die Gewebeoberfläche zum Zeitpunkt der Probennahme genau
bestimmen.

Methoden
13
2. METHODEN
2.1 Versuchstiere und Haltung
Für die Versuchsserie zur Orientierung von Helicobacter felis im Magenschleim der Maus
(Mus musculus) wurden 65 mit H. felis infizierte weibliche Mäuse verwendet. Die Infektion
mit dem Bakterienstamm ATCC 49179 (R. L. Ferrero, Institute Pasteur, Paris (Lee, 1988;
Paster, 1991)) wurde nach standardisierten Methoden (Mohammadi, 1996) von der
Forschungsabteilung der Firma Bayer AG, Wuppertal, im Rahmen dieses Projektes
durchgeführt. Den Mäusen, deren mittleres Gewicht 30 g betrug, wurde eine Infektionsdosis
von 1010 Keimen/ml oral verabreicht, der Nachweis der gelungenen Infektion erfolgte an 2
von 50 Mäusen histopathologisch.
Zur Untersuchung des chemotaktischen Verhaltens von Helicobacter pylori im
Magenschleim des Mongolischen Gerbils (Meriones unguiculatus) in vivo wurden 43 mit
H. pylori infizierte weibliche Gerbils, im Rahmen dieses Projektes, mittels standardisierter
Techniken (Hirayama, 1996), mit dem Bakterienstamm SS1 von der Firma Altana AG,
Konstanz, infiziert. Die Tiere wogen im Mittel 50 g. Der Infektionsnachweis erfolgte über
einen ELISA-Test bei jedem Tier.
Kontrollexperimente wurden mit nicht infizierten Tieren durchgeführt. Dabei handelte es
sich um 34 weibliche NMR-1 Auszucht-Mäuse sowie 36 weibliche Gerbils.
Alle infizierten Tiere wurden unter L2-Bedingungen in einem Unterdruck-Laminar-flow
Schrank (Uniprotect 1555, Ehret, Emmendingen) untergebracht.

Methoden
14
2.2 Narkose 2.2.1 Narkosetyp und Kreislaufsystem
Bei der Narkose handelt es sich um eine halboffene Inhalationsnarkose unter
Spontanatmung. Vorteil dieses Narkosetyps für die hier beschriebene Anwendung ist, dass
keine künstliche Beatmung notwendig ist und der Zustand der Versuchstiere an ihrer Atmung
kontrolliert werden kann.
Die Abbildung 4 zeigt schematisch den Narkosekreislauf. Die Tiere werden mit einer
Kombination aus Lachgas (Stickoxydul, N2O) und Halothan narkotisiert, als Trägergase
fungieren Sauerstoff und Kohlendioxid.
Steuerung des Outflow
Flowmeter“in”
Flowmeter“out”
Optosensor
Zeit[ms]
Amplitude
Aufzeichnung der PulsfrequenzB
A
Abb. 4:
A. Darstellung des Narkosekreislaufs im halboffenen Minimal-flow System. Das Tier liegt mit dem Kopf im Narkosegasstrom. Eine Membranpumpe sorgt mit 5 l/min für die Zirkulation im Kreissystem, die Frischgaszufuhr wird über einen Gasmischer geregelt, der Ausstrom kann ebenfalls reguliert werden. Für die Narkoseeinleitung wird der Gasstrom auf eine Plexiglasröhre geschaltet. B. Die Herzfrequenz wird mit einem optischen Pulssensor über die Blutdruckschwankungen in der Schwanzarterie aufgezeichnet. Die Messung der Herzfrequenz wird auf einem PC visualisiert und entsprechend dokumentiert.

Methoden
15
Der Kopf des Tieres befindet sich in einer Inhalationsmaske, die in das Kreislaufsystem
integriert ist. Das Kreislaufsystem der Narkosegase ist halboffen, d.h. die Gase werden
definiert über einen Gasmischer eingespeist, überschüssiges Gas kann entweichen. Unter
Kontrollbedingungen wird das Narkoseträgergas gemischt aus 3 % CO2, 57 % N2O und
40 % O2 für die Maus und 5 % CO2, 55 % N2O und 40 % O2 für den Gerbil. Die Frischgaszufuhr beträgt 1 l/min, was deutlich über dem Atemzeitvolumen von Maus
und Gerbil liegt, so dass die Gasfraktionen im System konstant bleiben. Die
Halothankonzentration wird über einen Halothanverdampfer reguliert.
2.2.2 Messung der Atem- und Herzfrequenz
Die Atemfrequenz wird anfangs in einem Abstand von 5 min, nach beendeter Operation
von 10 min, gemessen und dokumentiert.
Die Herzfrequenz wird nicht invasiv in der Schwanzarterie gemessen. An den Schwanz
des narkotisierten Tieres wird ein optischer Pulssensor (Infrarot-Sensor) angebracht (siehe
Abb. 4). Die durch Blutdruckschwankungen verursachten Änderungen im Durchmesser
beider großen arteriellen Gefäße führen zu einer veränderten Lichtdurchlässigkeit, welche
vom Pulssensor registriert und in ein elektrisches Signal umgewandelt wird. Nach
elektronischer Verstärkung und Filterung steht dieses Signal zur Anzeige und Auswertung als
Pulskurve zur Verfügung. Die Herzfrequenz wird aus der Pulskurve durch Mittelwertbildung
errechnet.

Methoden
16
2.3 Operation 2.3.1 Eröffnung des Bauchraumes
Zur Stabilisierung der Körpertemperatur werden die Tiere nach der Narkoseeinleitung in
ein Heizbett überführt, welches auf 37°C erwärmt ist. Das Heizbett ist in einem Winkel von
10° nach caudal gekippt, damit die narkotisierten Tiere nicht aspirieren.
Zur Eröffnung des Bauchraumes wird ein Schnitt einige Millimeter unterhalb des
Brustbeines in sicherem Abstand zu Mediastinum und Zwerchfell angesetzt. Mit einem
Thermokauter wird die Bauchwand eröffnet. Bei der Schnittführung durchtrennte Gefäße, vor
allem die v. und a. epigastrica inferior und superior, werden durch Hitze direkt verödet, so
dass der Blutverlust minimal ist.
2.3.2 Fixierung des Magens
Während der Fixierung des Magens darf auf das Organ weder Druck noch Zug ausgeübt
werden, noch dürfen Beeinträchtigungen in der Gefäßversorgung oder Verletzungen der
umgebenden Strukturen entstehen. Nur ein völlig intakter und gut durchbluteter Magen
gewährleistet unverfälschte experimentelle Daten.
Der Magen wird durch den Einsatz eines Gewebehakens, der die Leber nach kranial
schiebt, lokalisiert und mit Gewebefasspinzetten vorsichtig aus dem Bauchraum
herausgehoben.
Gewebehaken
Spatel mit Pin
Gewebefass-pinzette
Unterbindungsnadel
Große Kurvatur mita. gastroepiploica
A B
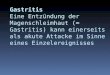
Abb. 5: Fotografische Darstellung der Operationsschritte. A: Lokalisierung des Magens im Bauchraum. Mit einem Haken wird die Leber nach kranial geschoben und der Magen mit einer Gewebefasspinzette vorsichtig aus dem Bauchraum herausgehoben. B: Fixierung des Magens. Durch das Omentum minus wird medial des Pylorus eine Unterbindungsnadel gestochen, die den Magen anhebt.
Methoden
17
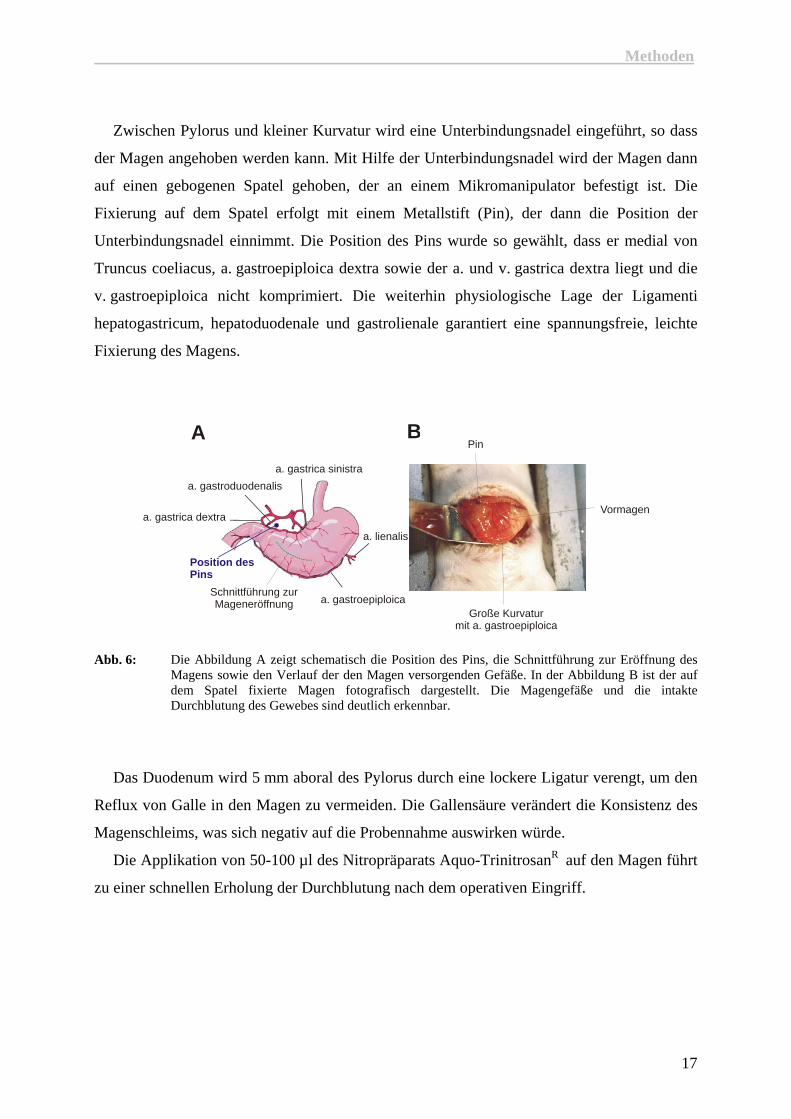
Zwischen Pylorus und kleiner Kurvatur wird eine Unterbindungsnadel eingeführt, so dass
der Magen angehoben werden kann. Mit Hilfe der Unterbindungsnadel wird der Magen dann
auf einen gebogenen Spatel gehoben, der an einem Mikromanipulator befestigt ist. Die
Fixierung auf dem Spatel erfolgt mit einem Metallstift (Pin), der dann die Position der
Unterbindungsnadel einnimmt. Die Position des Pins wurde so gewählt, dass er medial von
Truncus coeliacus, a. gastroepiploica dextra sowie der a. und v. gastrica dextra liegt und die
v. gastroepiploica nicht komprimiert. Die weiterhin physiologische Lage der Ligamenti
hepatogastricum, hepatoduodenale und gastrolienale garantiert eine spannungsfreie, leichte
Fixierung des Magens.
Position des Pins
Pin
Vormagen
Große Kurvaturmit a. gastroepiploica
Schnittführung zur Mageneröffnung a. gastroepiploica
a. lienalis
a. gastrica dextra
a. gastrica sinistraa. gastroduodenalis
A B
Abb. 6: Die Abbildung A zeigt schematisch die Position des Pins, die Schnittführung zur Eröffnung des Magens sowie den Verlauf der den Magen versorgenden Gefäße. In der Abbildung B ist der auf dem Spatel fixierte Magen fotografisch dargestellt. Die Magengefäße und die intakte Durchblutung des Gewebes sind deutlich erkennbar.
Das Duodenum wird 5 mm aboral des Pylorus durch eine lockere Ligatur verengt, um den
Reflux von Galle in den Magen zu vermeiden. Die Gallensäure verändert die Konsistenz des
Magenschleims, was sich negativ auf die Probennahme auswirken würde.
Die Applikation von 50-100 µl des Nitropräparats Aquo-TrinitrosanR auf den Magen führt
zu einer schnellen Erholung der Durchblutung nach dem operativen Eingriff.

Methoden
18
2.3.3 Peritoneale Spülung und Dialyse
Vor der Eröffnung des Magens werden die zu- und abführenden Schläuche für die
peritoneale Spülung angebracht, die als peritoneale Dialyse wirkt. Das Ziel der hier
eingesetzten Dialyse ist es, zum einen den Salz- und Flüssigkeitshaushalt der Tiere während
der Narkose stabil zu halten und damit die Durchblutung der Organe zu verbessern, und zum
anderen die Plasmakonzentrationen verschiedener Substanzen gezielt zu beeinflussen.
Damit eine peritoneale Spülung als Dialyse wirkt, muss sie das gastrointestinale
Durchblutungs-Minutenvolumen übersteigen. Mittels radioaktiver Microspheres haben
Matsumoto et al. gemessen, dass für den Mongolischen Gerbil das Minutenvolumen aller im
Peritonealraum gelegenen Organe, einschließlich der großen Verdauungsdrüsen, 5,6 ml/min
beträgt (Matsumoto, 1982). Da die Leber nur an ihrer caudalen Oberfläche und der
herausgehobene Magen praktisch nicht von der peritonealen Spülung erreicht wird, ist im
umspülten Durchblutungsgebiet mit einem Minutenvolumen von ca. 4,5 ml zu rechnen. Der
gewählte peritoneale Strom von 7 ml/min erwies sich für den Gerbil daher als effektive
Dialyse. Für eine 30 g Maus, ergibt sich daraus eine Dialysegeschwindigkeit von 3 ml/min.
Kleinmolekulare Substanzen ließen sich wenige Minuten nach Start der Dialyse im
Blutplasma nachweisen und erreichten nach 15 min im Plasma die Dialyse-Konzentration3.
2.3.4 Eröffnung und Reinigung des Magens
Zur Eröffnung des Magens wird mit einem unipolaren Hochfrequenzskalpell ein kleiner
Schnitt in die ventrale Magenwand gesetzt. Der Schnitt verläuft, wie aus der Abbildung 6
ersichtlich, parallel zur großen Kurvatur in Form eines Halbkreises, beginnend von der
Pylorusregion, lateral oralwärts, bis auf Höhe der größeren Äste der a. und v. gastrica dextra,
etwa zur Mitte des Magens.
Zur Reinigung des Magenlumens wird der Chymus vorsichtig entfernt, wobei wichtig ist,
dass die Schleimschicht nicht beschädigt wird. Anschließend wird die luminale Superfusion
angebracht.
3 Entsprechende Experimente wurden von der Firma Altana AG durchgeführt.

Methoden
19
2.3.5 Blutgasanalyse und Narkoseende
Nach Ende der Experimente wird der Säure-Basen Status im arteriellen Blut bestimmt.
Dazu wird die Aorta abdominalis dargestellt und sauber durchtrennt, was binnen Sekunden zu
Herz- und Atemstillstand der Tiere führt. Das arterielle Blut wird in heparinisierten
Glaskapillaren aufgefangen und zur Messung in den Säure-Basen-Analysator (ABL 510,
Firma Radiometer, Kopenhagen) eingebracht. Der Analysator misst den pH, den pCO2 und
den pO2 in der Blutprobe und berechnet daraus die Sauerstoffsättigung, die
Bicarbonatkonzentration sowie den Säure/Basen-Status des arteriellen Blutes. Zusätzlich kann
der Hämoglobingehalt des Blutes bestimmt werden.

Methoden
20
2.4 Beschreibung des Messfeldes Das Messfeld, aus welchem die Proben entnommen werden, ist der Antrumbereich der
dorsalen Magenwand. Orientierungspunkte des Antrums sind der Pyloruseingang sowie die
Plica salivarius, die nur bei der Maus und anderen Kleinnagern vorhanden ist. Bei der Plica
handelt es sich um eine natürliche Begrenzungslinie, die durch ihre weißliche Farbe gut zu
erkennen ist und an blütenblattähnliche Gewebeläppchen erinnert. Sie zieht sich etwa von der
Mitte der kleinen Kurvatur entlang schräg nach links caudal.
Duodenum
Ventrale Magenwand(aufgeklappt)
Feuchte LäppchenLuminale
Superfusion
Klemmen
Absaugung
Antrum(Messfeld)
Abb. 7: Dargestellt ist das von Nahrungsbrei gereinigte Messfeld in der dorsalen Antrumwand des
Magens. Die Magenwände werden von zwei Mikroklemmen fixiert, das Magenlumen wird von einer Lösung superfundiert. Der intakte Magenschleim (weißliche Schicht über dem Gewebe) sowie die gute Durchblutung (rosige Färbung des Gewebes) des Messfelds sind erkennbar.
Die Entnahme von Schleimproben erfolgt aus der rechtsseitigen Region um die Plica, aus
dem Bereich vor dem Pylorus und aus der zentralen Antrumregion. Voraussetzungen für eine
erfolgreiche Probennahme sind eine intakte Schleimschicht, das Fehlen von Erosionen, sowie
eine gleichmäßig gute Durchblutung des Messfeldes.

Methoden
21
2.5 Entnahme von Schleimproben aus der Magenmucosa Für die Entnahme von Schleimproben aus dem Magenschleim des Antrums von Maus und
Gerbil, wird ein von unserer Gruppe entwickeltes Nanoprobennahmesystem (Schreiber,
1998), das, neben präziser Nanoprobennahme aus dem Magenschleim, auch präzise
Hochdrucknanoinjektionen ausführen kann, verwendet.
Heizspirale
Temperatursensor
Nanoprobennahme Pipette
1000fach
15 µm
A
B
C
Abb. 8: Fotografie (A) sowie schematische Darstellung (B) der Nanoprobennahmeeinheit. Die
Nanoprobennahmeeinheit besteht aus einer Glaspipette, die in einen teflonummantelten Heizkopf eingespannt ist. Der Pipettenschaft ist von einer Heizspirale umgeben, über die die Temperatur exakt geregelt wird. Die Pipettenspitze und ca. 5 mm des Schaftes sind mit Pufferlösung befüllt, daran anschließend befindet sich eine 15 mm Silikonölstrecke. Durch einen in Epoxyklebstoff eingebetteten Glaskolben ist die Pipette druckfest verschlossen. Die Pipettespitze (C) ist auf einen Durchmesser von ca. 15 µm in einem Winkel von 20° geschliffen (Schreiber, 1998).
Das Nanoprobennahmesystem besteht aus einer druckfest verschlossenen Glaspipette,
deren Schaft mit Silikonöl gefüllt wurde und die in einen Heizkopf eingespannt wird. Die
Temperatur im Heizkopf kann über eine Steuereinheit definiert variiert werden, was dazu
führt, das Lösung aus der Pipettenspitze injiziert oder umgebendes Medium in die Pipette
aufgesaugt werden kann.

Methoden
22
Die Herstellung der Probennahmepipetten sowie deren Handhabung bei der Entnahme von
Schleimproben kann in Schreiber, Stüben et al., 1998 (Schreiber, 1998) nachgelesen werden.
Vorteile dieses Systems sind, dass kleine Proben mit gleichbleibendem Volumen, die alle
Bereiche des Magenschleims inklusive des oberflächlichen Epithels enthalten, entnommen
werden können, ohne große Schäden in der Mucosa zu hinterlassen. Zusätzlich beträgt die
Zeitspanne von der Entnahme bis zur Abkühlung der Schleimprobe nur etwa 30 Sekunden,
wodurch der Erhalt der in vivo Bedingungen in der Probe gewährleistet bleibt.
Aus der Antrumregion von Maus und Gerbil werden während eines Experimentes bis zu 6
Nanoproben entnommen. Für die mikroskopische Auswertung werden die Proben auf einen
Diagnoseobjektträger in einen Flüssigkeitstropfen appliziert. Die Objektträger können auf 5°C
abgekühlt oder auf 37°C erwärmt werden. Eine Abkühlung erfolgt bei der Positions- und
Dichtebestimmung der Bakterien, um die Bedingungen in der Probe „einzufrieren“, die
Erwärmung wird bei der Messung der Motilität eingesetzt, um die in vivo Beweglichkeit der
Bakterien zu erhalten.

Methoden
23
2.6 Auswertung der Nanoschleimproben
Die aus dem Magenschleim des Antrums entnommene in vivo Probe besteht in der Regel
aus einem Gewebeanteil, aus epithelnahem fibrillären Schleims sowie einem großen Anteil
luminalem wolkigen Schleim.
Ziel der Auswertung ist es, die räumliche Lage der Bakterien zur Gewebeoberfläche
darzustellen und ihre Motilität zu bestimmen.
2.6.1 Ermittlung von Bakteriendichte und Abstand zur Gewebegrenze
Einzelne Bereiche, der auf 5°C abgekühlten Gesamtschleimprobe, werden digital
fotografiert und ihr Volumen berechnet. Die Anzahl der Bakterien in dem entsprechenden
Volumen wird ermittelt und die Dichte der Bakterien pro Nanoliter Schleim angegeben. In
den epithelnahen Bereichen wird zusätzlich der mittlere Abstand der Bakterien von der
Gewebegrenze ermittelt. Das genaue Verfahren wurde in Schreiber, Stüben et al. 1999
beschrieben (Schreiber, 1999).
2.6.2 Motilitätsmessung
Zur Bestimmung der mittleren Motilität der Bakterien in den Schleimproben, wird die
Temperatur innerhalb der Proben auf 37°C erhöht. Die motilen Bakterien im epithelnahen
Schleim werden mit 25 Einzelbildern pro Sekunde aufgezeichnet, woraus sich ein Zeitabstand
von 40 ms pro Bild ableitet. Die Strecke, die die Bakterien in eine Richtung zurücklegen, wird
vermessen und das dazugehörige Zeitintervall bestimmt. Um die curvilineare4
Geschwindigkeit der Bakterien im Magenschleim zu ermitteln, wird die zurückgelegte
Strecke den Größenverhältnissen der Probe angepasst und in Mikrometer pro Sekunde
angegeben.
4 Zerlegung einer komplexen räumlichen Bewegung in einzelne lineare Teilstücke, deren Länge pro Zeit gemessen werden kann

Methoden
24
Abb. 9: Digitale Vergrößerung einer epithelnahen Schleimregion mit H. felis. Die Bewegung der Bakterien im epithelnahen Schleim ist durch die Sternchen gekennzeichnet. Der Abstand zwischen zwei Sternchen entspricht einer Strecke von ca. 10 µm.
TissueMucus
10 µm
250 msec

Methoden
25
2.7 Versuchsbedingungen
2.7.1 Zeitliche Einteilung
Die einzelnen Experimente wurden in verschiedene Phasen unterteilt. Nach der Operation,
die ungefähr 40 min in Anspruch nahm, startete die Probennahmephase, die sich in eine
Kontrollphase und eine Phase veränderter Bedingungen (= Testphase) gliederte. In der
Abb. 10 ist die zeitliche Einteilung des Versuchsablaufes schematisiert dargestellt.
NarkosebeginnNarkoseendeCO -Applikation2
Operation35 - 40 min
Inkubationsphase
Kontrollphaseca. 15 min
90 min 120 min
Testphase(veränderte Bedingungen)
Proben-nahme
Proben-nahme
Proben-nahme
Proben-nahme
60 min
Abb. 10: Übersicht über den zeitlichen Ablauf eines Experimentes mit den einzelnen Versuchsphasen.
In der Kontrollphase wurden in schneller Abfolge ein bis drei Nanoproben entnommen, um
die Bakteriendichte unter Kontrollbedingungen zu ermitteln. Daran anschließend folgte eine
60 minütige Inkubation zur Einstellung der Testbedingungen am Magen. Nach der Inkubation
wurden im Abstand von 30 min drei weitere Nanoproben aus dem Magenschleim entnommen.

Methoden
26
2.7.2 Kontrollbedingungen
Unter Kontrollbedingungen bestand die luminale Superfusion aus einer 0,9 %
Natriumchloridlösung, die durch Begasung mit einem Gasgemisch aus 10 % CO2, 5 % O2 und
85 % N2 auf einen pH von 3,8 äquilibriert wurde. Der Fluss der luminalen Lösung betrug bei
allen Versuchen 30 ml/h bei der Maus, und 70 ml/h beim Gerbil.
Als Kontrolldialysat wurde eine peritoneale Lösung eingesetzt, die alle im Plasma
wichtigen Salze sowie Glucose, Harnstoff und bei der Maus zusätzlich 200 mg/l
Aminophyllin zur Steigerung des Atemantriebs enthielt. Das Aminophyllin gehört der Gruppe
der Methylxanthine an und ist ein Derivat des Theophyllins. Die Methylxanthine wirken im
Zentralnervensystem erregend auf die Herz-Kreislauffunktionen. Unter Narkose führte die
Gabe von Aminophyllin zu einer Steigerung der Atemfrequenz und damit zu einer dezenten
Hyperventilation. In den Experimenten mit verändertem Schleim-Gradient wurde
Aminophyllin eingesetzt, um den arteriellen pCO2 abzusenken.
Die leichte Hypotonie der Lösung, die durch eine dezente Absenkung der
Natriumkonzentration erzeugt wurde, führt zu einer effizienteren Dialyse, indem vermehrt
Flüssigkeit in die Gefäße einströmt und somit die zu dialysierenden Substanzen
mittransportiert werden (solvent drag).
Tabelle 1: Zusammensetzung der peritonealen Dialyse unter Kontrollbedingungen. Ausgehend von dieser
Lösung wurden alle weiteren in dieser Arbeit verwendeten Dialysen hergestellt.
Maus Gerbil
80 mM NaCl 4 mM KCl
35 mM NaHCO3 0,5 mM Mg2SO4 1 mM KH2PO4
20 mM Tris 2 mM CaCl2
pH 7,4 bei 37°C 11 mM Glucose 5 mM Harnstoff
200 mg/l Aminophyllin
80 mM NaCl 4 mM KCl
35 mM NaHCO3 0,5 mM Mg2SO4 1 mM KH2PO4
20 mM Tris 2 mM CaCl2
11 mM Glucose
5 mM Harnstoff
pH 7,4 bei 37°C

Methoden
27
2.7.3 Phase veränderter Bedingungen
Um Gradienten oder einzelne Substanzkonzentrationen im Magenschleim zu verändern,
wurden die luminale Superfusion, die peritoneale Dialyse sowie die inspiratorischen
Gasfraktionen je nach Bedarf variiert.
Inspiratorische Gasfraktion
Änderung von pCO2
Luminale SuperfusionÄnderung von
pH [HCO ] [CO ] [Harnstoff] [NH ]
3 2
4
-
+
Peritoneale DialyseÄnderung von
[HCO ] [Harnstoff] [NH ]
3
4
-
+
Abb. 11: Darstellung der Möglichkeiten, um am narkotisierten Tier die Bedingungen an der Mucosa zu verändern. Über die luminale Superfusion können der luminale pH, sowie die Konzentrationen von CO2, Bicarbonat, Harnstoff und Ammonium variiert werden. Die peritoneale Dialyse ermöglicht Änderungen der Plasmakonzentrationen von Bicarbonat, CO2, Harnstoff und Ammonium, über die inspiratorische Gasfraktionen kann der arterielle pCO2 verändert werden.
Die luminale Superfusion erlaubt die Veränderung der Bedingungen im Magenlumen.
Über die inspiratorische Gaszusammensetzung können die Partialdrücke von Sauerstoff und
Kohlendioxid im arteriellen Blut verändert werden. Die Dialyse ermöglicht die Einstellung
bestimmter Konzentrationen einzelner Substanzen im arteriellen Blut und somit an der
Gewebeseite des Magens.

Methoden
28
2.8 Statistische Auswertung der Daten
Für jeden einzelnen Versuch wurde ein Mittelwert der epithelnahen Bakteriendichte in
Bakterien pro Nanoliter Mucus für die Kontrollphase bzw. die Phase veränderter
Bedingungen ermittelt. Aus diesen beiden Zahlen wurde ein Quotient Testdichte, geteilt durch
Kontrolldichte, berechnet. Die Quotienten jeder Versuchsserie wurden gemittelt und die
Standardabweichungen angegeben. Lagen die Quotienten bei einem Wert um 1, fand keine
Umorientierung statt. Quotienten mit einem signifikant niedrigeren Wert wiesen auf eine
Umorientierung hin. Als Signifikanztest wurde der „students T-Test“ verwendet; als
signifikant galt ein p-Wert < 0,05.
Zur Bestimmung der Schleimprobenvolumina der ausgewerteten Regionen wurde das
mittlere, ausgewertete Schleimprobenvolumen unter Kontroll- und Testbedingungen für alle
Experimente mit der dazugehörigen Standardabweichung berechnet.
Die Bakterien befanden sich in einer 25 µm dicken Schleimschicht. Zur Darstellung der
Verteilung der Bakterien wurde dieser epithelnahe Mucus noch einmal in 5 µm dicke
Schichten unterteilt. Der Abstand der Bakterien von der Gewebeoberfläche wurde gemessen
und die Bakterien wurden jeweils einer der Schichten 0-5 µm, 5-10 µm, 10-15 µm, 15-20 µm,
20-25 µm zugeordnet. Auf diese Weise konnte nach Abschluss der Auswertung angegeben
werden, wie viel Prozent der Bakterien sich jeweils in welcher Schicht befanden. Aus allen
Abstandsmessungen wurde ein mittlerer Abstand ermittelt.
Die mittlere Motilität der Bakterien wurde unter Kontroll- und Testbedingungen mit der
dazugehörigen Standardabweichung berechnet.
Ergebnisse
29
3. ERGEBNISSE
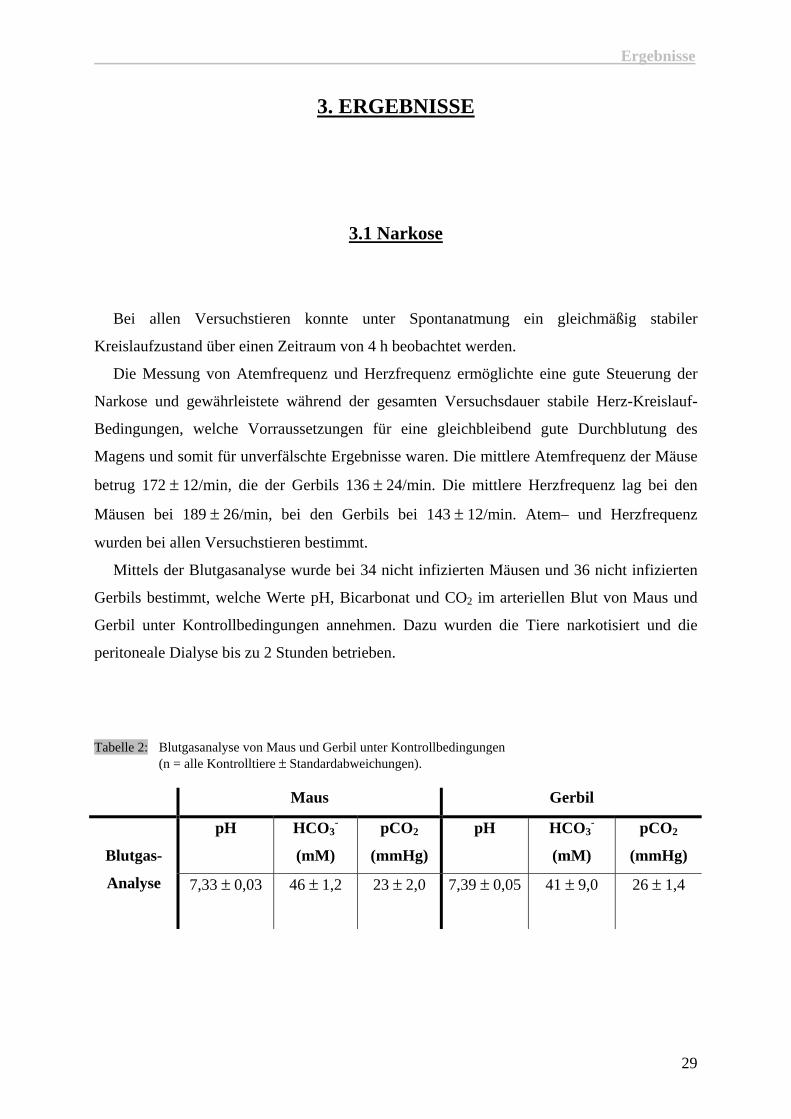
3.1 Narkose
Bei allen Versuchstieren konnte unter Spontanatmung ein gleichmäßig stabiler
Kreislaufzustand über einen Zeitraum von 4 h beobachtet werden.
Die Messung von Atemfrequenz und Herzfrequenz ermöglichte eine gute Steuerung der
Narkose und gewährleistete während der gesamten Versuchsdauer stabile Herz-Kreislauf-
Bedingungen, welche Vorraussetzungen für eine gleichbleibend gute Durchblutung des
Magens und somit für unverfälschte Ergebnisse waren. Die mittlere Atemfrequenz der Mäuse
betrug 172 ± 12/min, die der Gerbils 136 ± 24/min. Die mittlere Herzfrequenz lag bei den
Mäusen bei 189 ± 26/min, bei den Gerbils bei 143 ± 12/min. Atem– und Herzfrequenz
wurden bei allen Versuchstieren bestimmt.
Mittels der Blutgasanalyse wurde bei 34 nicht infizierten Mäusen und 36 nicht infizierten
Gerbils bestimmt, welche Werte pH, Bicarbonat und CO2 im arteriellen Blut von Maus und
Gerbil unter Kontrollbedingungen annehmen. Dazu wurden die Tiere narkotisiert und die
peritoneale Dialyse bis zu 2 Stunden betrieben.
Tabelle 2: Blutgasanalyse von Maus und Gerbil unter Kontrollbedingungen (n = alle Kontrolltiere ± Standardabweichungen).
Maus Gerbil
pH HCO3-
(mM)
pCO2
(mmHg)
pH HCO3-
(mM)
pCO2
(mmHg)
Blutgas-
Analyse 7,33 ± 0,03 46 ± 1,2 23 ± 2,0 7,39 ± 0,05 41 ± 9,0 26 ± 1,4

Ergebnisse
30
3.2 Operation
Die blutarme Operation belegt der stabile Hämoglobingehalt im arteriellen Blut der Tiere,
der bei 34 Mäusen und 36 Gerbils postoperativ bestimmt wurde. Bei den Mäusen lag der Hb
bei 160 ± 16 g/l und bei den Gerbils bei 140 ± 14 g/l.
Das Messfeld in der dorsalen Magenwand des eröffneten Magens zeichnete sich nach guter
Operation durch eine gleichmäßige rosige Färbung und eine intakte Schleimschicht aus.
3.3 Nanoprobennahme
Der Magenschleim der Maus und des Gerbils ist etwa 100 bzw. 150 µm dick und lässt sich
in mehrere Bereiche einteilen. Direkt dem Epithel aufgelagert findet man in einer Dicke von
25 µm den epithelnahen Schleim, das Habitat von Helicobacter. Dann folgt bis zum
Magenlumen der zentrale und luminale Schleim.
In der aus luminalem/zentralem und epithelnahem Schleim sowie Gewebeanteilen
bestehenden Nanoprobe können diese Regionen optisch unterschieden werden. Der
epithelnahe Schleim zeichnet sich durch eine durchsichtige, fibrilläre Struktur aus. Der
zentrale/luminale Schleim ist beim Gerbil ebenfalls durchsichtig, weist aber keine Fibrillen
auf. Bei der Maus ist der luminale Schleim hingegen undurchsichtig und wolkig.

Ergebnisse
31
3.4 Verteilung von Helicobacter im Magenschleim unter
Kontrollbedingungen
Wurden unter Kontrollbedingungen (siehe 2.7.2) Nanoproben aus dem Magen von Maus
und Gerbil entnommen und die Bakterienzahlen in epithelnahen und luminalen bzw. zentralen
Schleimregionen, sowie ihr mittlerer Abstand vom Epithel bestimmt, so ergaben sich
deutliche Unterschiede in der Verteilung von H. felis und H. pylori.
Die Dichte der Bakterien im epithelnahen Schleim war in beiden Spezies annähernd gleich
hoch (Maus: 3900 H.f./nl; Gerbil: 2900 H.p./nl), in luminalen und zentralen Schleimregionen
fanden sich bei Maus und Gerbil weniger als 50 Bakterien/nl Schleim.
Im Magen der Maus halten sich 90 % der H. felis vorwiegend in den Schleimanteilen
10-25 µm vom Epithel entfernt auf, während sich im Gerbilmagen 90 % der H. pylori in dem
Bereich 0-15 µm vom Epithel entfernt fanden. Einige wenige H. pylori fanden sich an
Epithelzellen adhäriert, während H. felis nicht adhärierte. Der mittlere Abstand von H. felis
zum Magenepithel der Maus betrug 16 ± 6 µm, H. pylori fand sich in einem mittleren
Abstand von 11 ± 5 µm zum Magenepithel des Gerbils. Die prozentuale Verteilung von
H. felis und H. pylori im epithelnahen Schleim ist in der Abbildung 12 durch die
unterschiedlichen Grauschattierungen schematisiert dargestellt.
Die von H. pylori besiedelte ökologische Nische befindet sich entsprechend etwas näher an
der Magenepithelgrenze, als die Region, die H. felis besiedelt.
Innerhalb von Drüsengängen lag die Dichte von H. felis und von H. pylori bei ca. 5 pro
100 µm Drüsenganglänge.

Ergebnisse
32
Abb. 12: Schematischer Querschnitt durch die Magenmucosa von Maus und Gerbil. Die prozentuale Verteilung von H. felis und H. pylori im epithelnahen Schleim unter Kontrollbedingungen ist prozentual farblich abgestuft dargestellt.
Magenlumen
Mucus
Gewebe
Maus
0% 0-10 µm
0 - 5 µm0%5 - 10 µm12%
10 - 15 µm36%15 - 20 µm28%20 - 25 µm24%
Gerbil
0 - 5 µm30%5 - 10 µm32%
10 - 15 µm26%15 - 20 µm8%20 - 25 µm4%
0% 0-10 µm
Die Motilität von H. pylori und H. felis unter Normalbedingungen wies keinen Unterschied
auf. H. felis bewegt sich mit einer mittlere Geschwindigkeit von 30 ± 20 µm/sec (n = 15) im
Mucus, H. pylori erreicht eine mittlere Geschwindigkeit von 30 ± 6 µm/sec (n = 7).

Ergebnisse
33
3.5 Orientierung von Helicobacter im Magenschleim unter veränderten
Bedingungen
3.5.1 Der luminale pH und die luminalen Bicarbonat/CO2-Konzentrationen
Zur Klärung der Hypothese, die Orientierung von Helicobacter basiere auf einer Repellent-
Wirkung des niedrigen luminalen pH wurde dieser bei den narkotisierten Tieren von 3 auf 6
erhöht. Dazu wurde als luminale Superfusion anstelle von 0,9 % NaCl ein 50 mM MES Puffer
eingesetzt. Die Dialyselösung blieb gegenüber den Kontrollbedingungen unverändert.
Weder bei der mit H. felis infizierten Maus noch dem H. pylori infizierten Gerbil
reagierten die Bakterien auf diese Veränderung.
Abb. 13: Schematischer Querschnitt durch die Mucosa von Maus und Gerbil. Dargestellt sind der Einfluss der Neutralisierung des luminalen pH auf die Verteilung der Bakterien im Magenschleim. Auch die Änderung der luminalen Bicarbonat/CO2-Konzentrationen (Maus) zeigte keine Wirkung.
Magenlumen
Mucus
Gewebe pH 7,4
pH 6,5
pH 3 6
[HCO ] [CO ]3 2-
1 pCO 0 mmHg
2 pCO = 40 mmHg
3 pCO = 80 mmHg
2
2
2
Neben der Anhebung des luminalen pH auf 6 wurde mittels Begasung mit CO2 zusätzlich
der pCO2 zwischen 0 und 80 mmHg, und damit entsprechend die Bicarbonatkonzentration der
luminalen Superfusion variiert, da bei einem luminalen pH von 6 Bicarbonat und CO2 im
Verhältnis 1:1 vorhanden sind (siehe Abbildung 13).
Ergebnisse
34
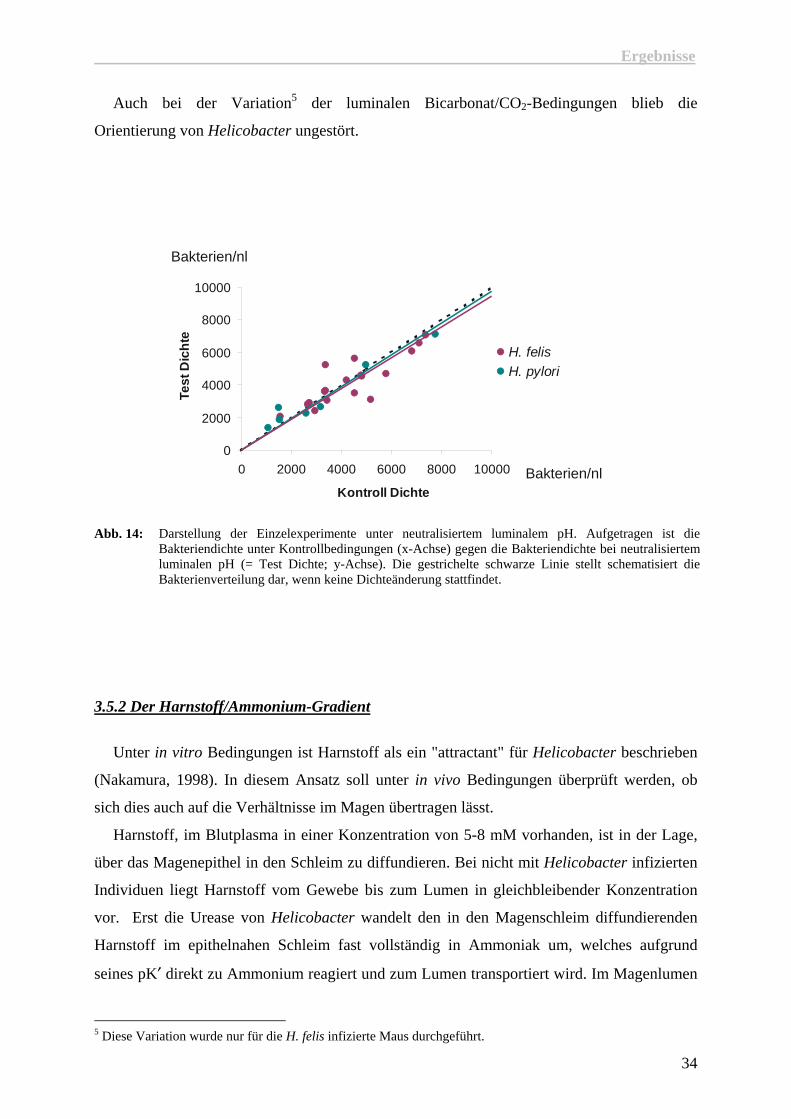
Auch bei der Variation5 der luminalen Bicarbonat/CO2-Bedingungen blieb die
Orientierung von Helicobacter ungestört.
0
2000
4000
6000
8000
10000
0 2000 4000 6000 8000 10000
Kontroll Dichte
Test
Dic
hte
H. felisH. pylori
Bakterien/nl
Bakterien/nl
Abb. 14: Darstellung der Einzelexperimente unter neutralisiertem luminalem pH. Aufgetragen ist die
Bakteriendichte unter Kontrollbedingungen (x-Achse) gegen die Bakteriendichte bei neutralisiertem luminalen pH (= Test Dichte; y-Achse). Die gestrichelte schwarze Linie stellt schematisiert die Bakterienverteilung dar, wenn keine Dichteänderung stattfindet.
3.5.2 Der Harnstoff/Ammonium-Gradient
Unter in vitro Bedingungen ist Harnstoff als ein "attractant" für Helicobacter beschrieben
(Nakamura, 1998). In diesem Ansatz soll unter in vivo Bedingungen überprüft werden, ob
sich dies auch auf die Verhältnisse im Magen übertragen lässt.
Harnstoff, im Blutplasma in einer Konzentration von 5-8 mM vorhanden, ist in der Lage,
über das Magenepithel in den Schleim zu diffundieren. Bei nicht mit Helicobacter infizierten
Individuen liegt Harnstoff vom Gewebe bis zum Lumen in gleichbleibender Konzentration
vor. Erst die Urease von Helicobacter wandelt den in den Magenschleim diffundierenden
Harnstoff im epithelnahen Schleim fast vollständig in Ammoniak um, welches aufgrund
seines pK′ direkt zu Ammonium reagiert und zum Lumen transportiert wird. Im Magenlumen
5 Diese Variation wurde nur für die H. felis infizierte Maus durchgeführt.

Ergebnisse
35
findet man bei mit Helicobacter infizierten Individuen anstelle von Harnstoff eine
Ammoniumkonzentration von 5-15 mM (Kim, 1990) .
(NH ) -CO2 2
(NH ) -CO2 2
(NH ) -CO2 2
Nicht infiziert
(NH ) -CO2 2urease
NH NH +H
4
3
+
+
NH4+
(NH ) -CO2 2
Infiziert
(NH ) -CO2 2
NH4+
NH4+
(NH ) -CO2 2
urease
Änderung des Gradienten
Abb. 15: Schematischer Querschnitt durch die Mucosa von Maus/Gerbil. Dargestellt sind die Änderungen des Harnstoff/Ammonium-Gradienten im Magenschleim.
Zur Änderung des Harnstoff/Ammonium-Gradienten im Magenschleim wurde über die
Dialyse die Blut Harnstoffkonzentration reduziert und die Ammonium-Konzentration erhöht.
Dazu wurde der Dialyselösung Ammoniumchlorid (5 mM) anstelle von Harnstoff zugesetzt.
Über die luminale Superfusion wurde die Harnstoffkonzentration im Magenlumen auf 20 mM
erhöht.
Unter dem so umgekehrten Gradienten waren keine Änderungen in der Verteilung von
H. felis und H. pylori im epithelnahen Magenschleim von Maus und Gerbil gegenüber den
Kontrollbedingungen zu finden. Die Blutgaswerte dieser Experimente entsprachen den
Kontrollwerten.
Ergebnisse
36
0
2000
4000
6000
8000
10000
0 2000 4000 6000 8000 10000
Kontroll Dichte
Test
Dic
hte
H. felis
H. pylori
Bakterien/nl
Bakterien/nl
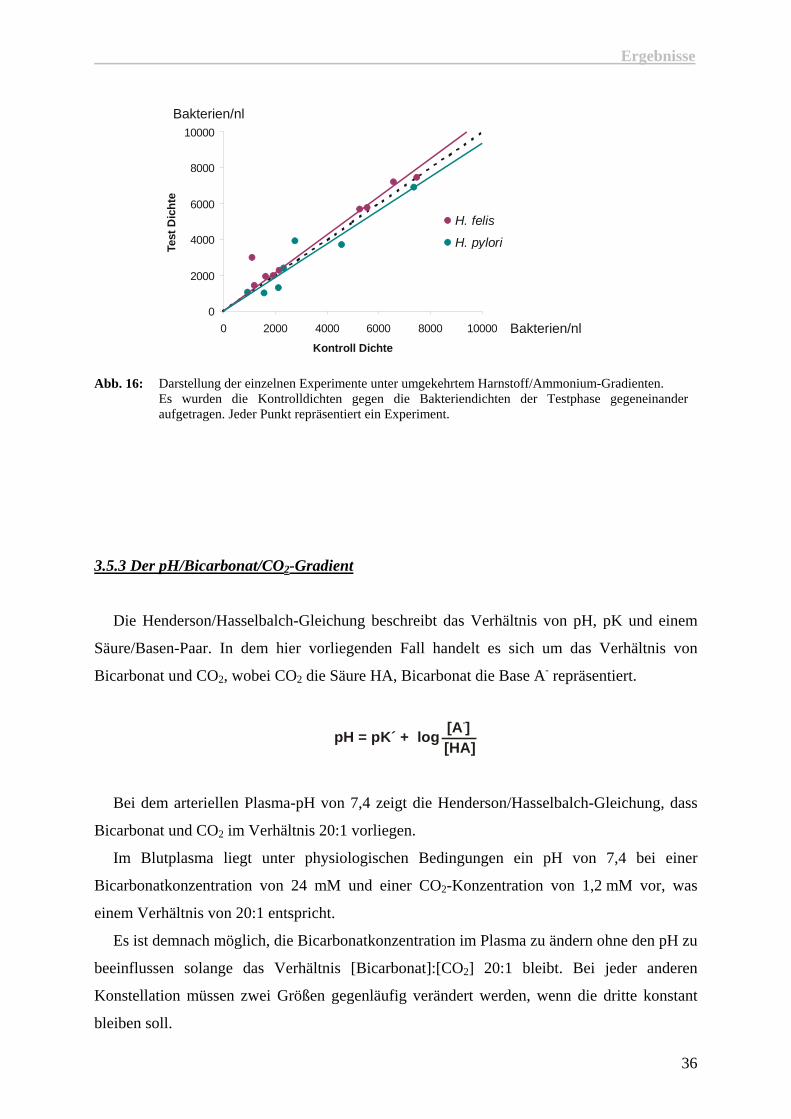
Abb. 16: Darstellung der einzelnen Experimente unter umgekehrtem Harnstoff/Ammonium-Gradienten.
Es wurden die Kontrolldichten gegen die Bakteriendichten der Testphase gegeneinander aufgetragen. Jeder Punkt repräsentiert ein Experiment.
3.5.3 Der pH/Bicarbonat/CO2-Gradient
Die Henderson/Hasselbalch-Gleichung beschreibt das Verhältnis von pH, pK und einem
Säure/Basen-Paar. In dem hier vorliegenden Fall handelt es sich um das Verhältnis von
Bicarbonat und CO2, wobei CO2 die Säure HA, Bicarbonat die Base A- repräsentiert.
pH = pK´ + log [A ]-
[HA]
Bei dem arteriellen Plasma-pH von 7,4 zeigt die Henderson/Hasselbalch-Gleichung, dass
Bicarbonat und CO2 im Verhältnis 20:1 vorliegen.
Im Blutplasma liegt unter physiologischen Bedingungen ein pH von 7,4 bei einer
Bicarbonatkonzentration von 24 mM und einer CO2-Konzentration von 1,2 mM vor, was
einem Verhältnis von 20:1 entspricht.
Es ist demnach möglich, die Bicarbonatkonzentration im Plasma zu ändern ohne den pH zu
beeinflussen solange das Verhältnis [Bicarbonat]:[CO2] 20:1 bleibt. Bei jeder anderen
Konstellation müssen zwei Größen gegenläufig verändert werden, wenn die dritte konstant
bleiben soll.

Ergebnisse
37
pH 7,4 = 6,1 + log art
24 mmol/l [HCO ]3-
1,2 mmol/l [CO ]2
Henderson/Hasselbalch
Epithelialer MucuspH 4,5 - 6
[HCO ] [CO ]3 2-
PlasmapH 7,4
[HCO ] 24 mmol/l[CO ] 12 mmol/l
3
2
-
LumenpH < 3
[CO ] > [HCO ]2 3-
Abb. 17: Schematischer Querschnitt durch die Mucosa von Maus und Gerbil. Dargestellt sind die pH-, Bicarbonat- und CO2-Verhältnisse an der Mucosa und im Schleim unter Normalbedingungen, über die Henderson/Hasselbalch-Gleichung sind diese drei Größen miteinander verknüpft.
Im epithelnahen Bereich des Magenschleims bei einem pH um 6 gleicht sich das
Verhältnis von Bicarbonat und CO2 auf äquimolare Konzentrationen an, im Magenlumen bei
saurem pH überwiegt die CO2-Konzentration. Es sind hier CO2-Konzentrationen von
≥ 60 mmHg beschrieben (Rune, 1969; Nagano, 1990). Bicarbonat wird von den Epithelzellen
der Mucosa sezerniert (Kauffman, Jr., 1980), wobei die Sekretionsrate der Epithelzellen von
einer ausreichend hohen Bicarbonatkonzentration im Plasma abhängig ist (Kivilaakso, 1981).
Im Folgenden wurde auf drei Arten in die pH- und Bicarbonat/CO2-Verhätnisse im
Magenschleim, bzw. im arteriellen Blut eingegriffen.

Ergebnisse
38
Epithelnaher Epithelnaher
pH 7,2[HCO3
-] 15 mMpCO2 40 mmHg
pH 7,2[HCO3
-] 15 mMpCO2 40 mmHg
pH = 6
[HCO3-] 15 mM
pCO2 40 mmHg
Erniedrigung des arteriellen pH und der
arteriellen Bicarbonatkonzentration
C
pH 7,4[HCO3
-] 15 mMpCO2 25 mmHg
pH 7,4[HCO3
-] 15 mMpCO2 25 mmHg
pH = 6
pH 7,4[HCO3
-] 15 mMpCO2 25 mmHg
pH 7,2[HCO3
-] 24 mMpCO2 60 mmHg
pH 7,2[HCO3
-] 24 mMpCO2 60 mmHg
Erniedrigung des arteriellen pH
pH = 6
pH 7,2[HCO3
-] 24 mMpCO2 60 mmHg
Erniedrigung der arteriellen
Bicarbonatkonzentration
BA
Mucus
Lumen
Gewebe
-
3 2[HCO] pCO- ↓ ↑ pH 3,0≈
pH 5,5[HCO] [CO]3 2
- ≈
pH 7,4[HCO] 24 mMpCO 40 mmHg
3
2
-
Mucus
Lumen
Gewebe
-
pH 3,0≈
pH 5,5
pH 7,4[HCO ] 24 mMpCO 40 mmHg
3
2
-
Normalbedingungen
Abb. 18: Schematischer Querschnitt durch die Mucosa von Maus/Gerbil. Dargestellt sind die Änderungen zur Beeinflussung der pH, Bicarbonat- und CO2 Verhältnisse an der Mucosa. A. Erniedrigter arterieller pH; B. Erniedrigte arterielle Bicarbonatkonzentration; C. Erniedrigter arterieller pH und erniedrigte arterielle Bicarbonatkonzentration.
Die Erniedrigung des arteriellen pH (Abb. 18 A) wird mittels einer Hyperkapnie
erzeugt, welche durch eine Kombination aus einer Erhöhung der inspiratorischen CO2-
Fraktion und einer durch die Dialyse normalisierten Bicarbonatkonzentration hervorgerufen
wird. Daraus resultiert ein niedriger, arterieller pH bei normaler Bicarbonatkonzentration und
erhöhtem arteriellem pCO2. Da man annimmt, dass der pH des frisch sezernierten Schleims
dem intrazellulären pH der mucusproduzierenden Zellen entspricht (Schreiber, 2000) und man
ferner weiß, dass die Epithelzellen sensitiv auf eine Ansäuerung des arteriellen Blutes
reagieren (Ashley, 1987), gingen wir davon aus, so den pH im epithelnahen Schleim
abzusenken. Die daraus tatsächlich resultierenden Blutgaswerte sind in der Tabelle 4
aufgeführt.
Unter diesen Bedingungen ändert sich die Verteilung von H. felis und H. pylori im
epithelnahen Magenschleim von Maus und Gerbil nicht. Auch der mittlere Abstand der
Bakterien vom Magenepithel sowie die Motilität bleiben unverändert.
Ergebnisse
39
0
2000
4000
6000
8000
10000
0 2000 4000 6000 8000 10000
Kontroll Dichte
Test
Dic
hte
H. felisH. pylori
Bakterien/nl
Bakterien/nl
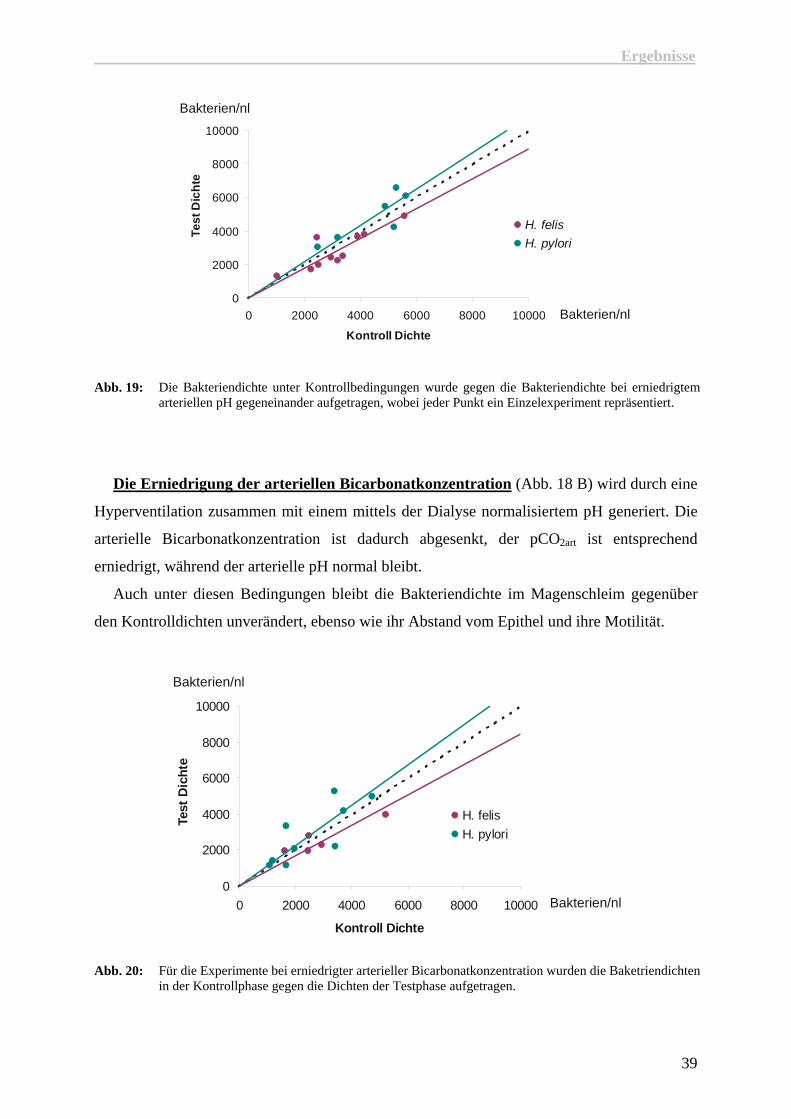
Abb. 19: Die Bakteriendichte unter Kontrollbedingungen wurde gegen die Bakteriendichte bei erniedrigtem arteriellen pH gegeneinander aufgetragen, wobei jeder Punkt ein Einzelexperiment repräsentiert.
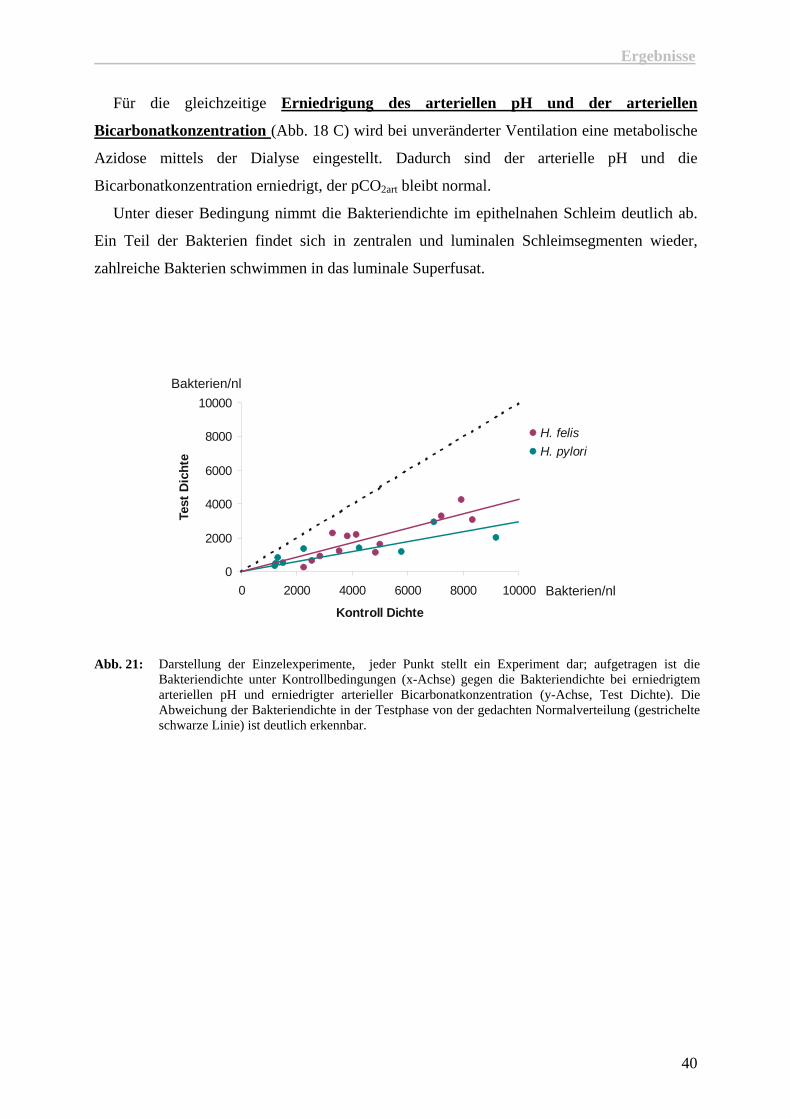
Die Erniedrigung der arteriellen Bicarbonatkonzentration (Abb. 18 B) wird durch eine
Hyperventilation zusammen mit einem mittels der Dialyse normalisiertem pH generiert. Die
arterielle Bicarbonatkonzentration ist dadurch abgesenkt, der pCO2art ist entsprechend
erniedrigt, während der arterielle pH normal bleibt.
Auch unter diesen Bedingungen bleibt die Bakteriendichte im Magenschleim gegenüber
den Kontrolldichten unverändert, ebenso wie ihr Abstand vom Epithel und ihre Motilität.
0
2000
4000
6000
8000
10000
0 2000 4000 6000 8000 10000
Kontroll Dichte
Test
Dic
hte
H. felisH. pylori
Bakterien/nl
Bakterien/nl
Abb. 20: Für die Experimente bei erniedrigter arterieller Bicarbonatkonzentration wurden die Baketriendichten in der Kontrollphase gegen die Dichten der Testphase aufgetragen.
Ergebnisse
40
Für die gleichzeitige Erniedrigung des arteriellen pH und der arteriellen
Bicarbonatkonzentration (Abb. 18 C) wird bei unveränderter Ventilation eine metabolische
Azidose mittels der Dialyse eingestellt. Dadurch sind der arterielle pH und die
Bicarbonatkonzentration erniedrigt, der pCO2art bleibt normal.
Unter dieser Bedingung nimmt die Bakteriendichte im epithelnahen Schleim deutlich ab.
Ein Teil der Bakterien findet sich in zentralen und luminalen Schleimsegmenten wieder,
zahlreiche Bakterien schwimmen in das luminale Superfusat.
0
2000
4000
6000
8000
10000
0 2000 4000 6000 8000 10000
Kontroll Dichte
Test
Dic
hte
H. felisH. pylori
Bakterien/nl
Bakterien/nl
Abb. 21: Darstellung der Einzelexperimente, jeder Punkt stellt ein Experiment dar; aufgetragen ist die Bakteriendichte unter Kontrollbedingungen (x-Achse) gegen die Bakteriendichte bei erniedrigtem arteriellen pH und erniedrigter arterieller Bicarbonatkonzentration (y-Achse, Test Dichte). Die Abweichung der Bakteriendichte in der Testphase von der gedachten Normalverteilung (gestrichelte schwarze Linie) ist deutlich erkennbar.

Ergebnisse
41
3.6 Messung des epithelnahen pH in vitro
Um zu überprüfen, ob die Änderung der pH-/Bicarbonat-
/CO2-Verhältnisse im arteriellen Blut eine Änderung der
Bedingungen im Magenschleim nach sich zieht, die die
Orientierung der Bakterien im epithelnahen Schleim
beeinflussen kann, wurden pH-Messungen an der isolierten
Meerschweinmucosa durchgeführt.
Dazu wurde eine ionensensitive Mikroelektrode im
epithelnahen Schleim einer explanierten Meerschwein-Antrum-
Mucosa positioniert. Das genaue Verfahren ist in Schreiber et
al., 1997 (Schreiber, 1997) nachzulesen.
Die gewebeseitige Lösung sowie die luminale Superfusion,
wurden nach Positionierung der Elektrode im epithelnahen
Schleim den veränderten pH/Bicarbonat/CO2-Bedingungen
angepasst. Über einen Zeitraum von 60 min wurde der pH registriert, nach 10 min unter
normalen Bedingungen wurden die Plasmalösungen geändert.
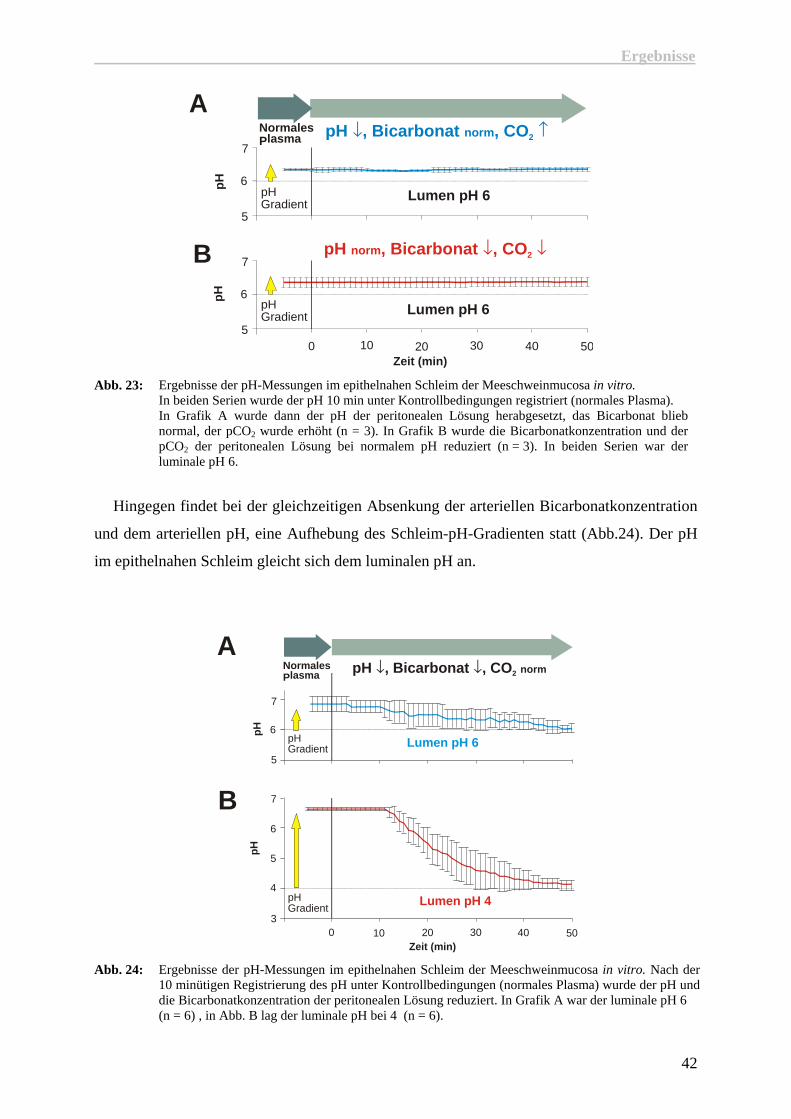
Wie in der Abbildung 23 zu sehen ist, veränderte sich der epithelnahe pH nicht, wenn nur
der arterielle pH (A) oder nur die arterielle Bicarbonatkonzentration (B) reduziert wurden.
PlasmaKonzentrationen
Epithelialer pH
Mucus
Lumen Ionenselektive Mikroelektrode
Abb. 22: Schematische Darstellung der pH-Messung im epithelnahen Schleim.
Ergebnisse
42
Zeit (min)
Lumen pH 6
7
6
5pH
Lumen pH 6
7
6
50 10 20 30 40
pH
50
NormalesPlasma pH , Bicarbonat , CO ↓ ↑norm 2
pH , Bicarbonat , CO norm ↓ ↓2
pHGradient
pHGradient
A
B
Abb. 23: Ergebnisse der pH-Messungen im epithelnahen Schleim der Meeschweinmucosa in vitro.
In beiden Serien wurde der pH 10 min unter Kontrollbedingungen registriert (normales Plasma). In Grafik A wurde dann der pH der peritonealen Lösung herabgesetzt, das Bicarbonat blieb normal, der pCO2 wurde erhöht (n = 3). In Grafik B wurde die Bicarbonatkonzentration und der pCO2 der peritonealen Lösung bei normalem pH reduziert (n = 3). In beiden Serien war der luminale pH 6.
Hingegen findet bei der gleichzeitigen Absenkung der arteriellen Bicarbonatkonzentration
und dem arteriellen pH, eine Aufhebung des Schleim-pH-Gradienten statt (Abb.24). Der pH
im epithelnahen Schleim gleicht sich dem luminalen pH an.
Lumen pH 6
7
6
5
Zeit (min)
pH
0 10 20 30 40 50
7
6
5
4
3
pH
Lumen pH 4
NormalesPlasma pH , Bicarbonat , CO ↓ ↓ 2 norm
pHGradient
pHGradient
A
B
Abb. 24: Ergebnisse der pH-Messungen im epithelnahen Schleim der Meeschweinmucosa in vitro. Nach der
10 minütigen Registrierung des pH unter Kontrollbedingungen (normales Plasma) wurde der pH und die Bicarbonatkonzentration der peritonealen Lösung reduziert. In Grafik A war der luminale pH 6 (n = 6) , in Abb. B lag der luminale pH bei 4 (n = 6).

Ergebnisse
43
Um dieses Ergebnis zu verdeutlichen, wurde, wie in Abbildung 24 dargestellt, neben dem
luminalen pH 6 zusätzlich noch der luminale pH 4 untersucht. Es ist deutlich zu erkennen,
dass in beiden Fällen eine Absenkung des epithelnahen pH auf den luminalen pH erfolgt und
damit eine Absenkung des pH-Gradienten im Mucus.
Ergebnisse
44
3.7 Zusammenfassung der Ergebnisse
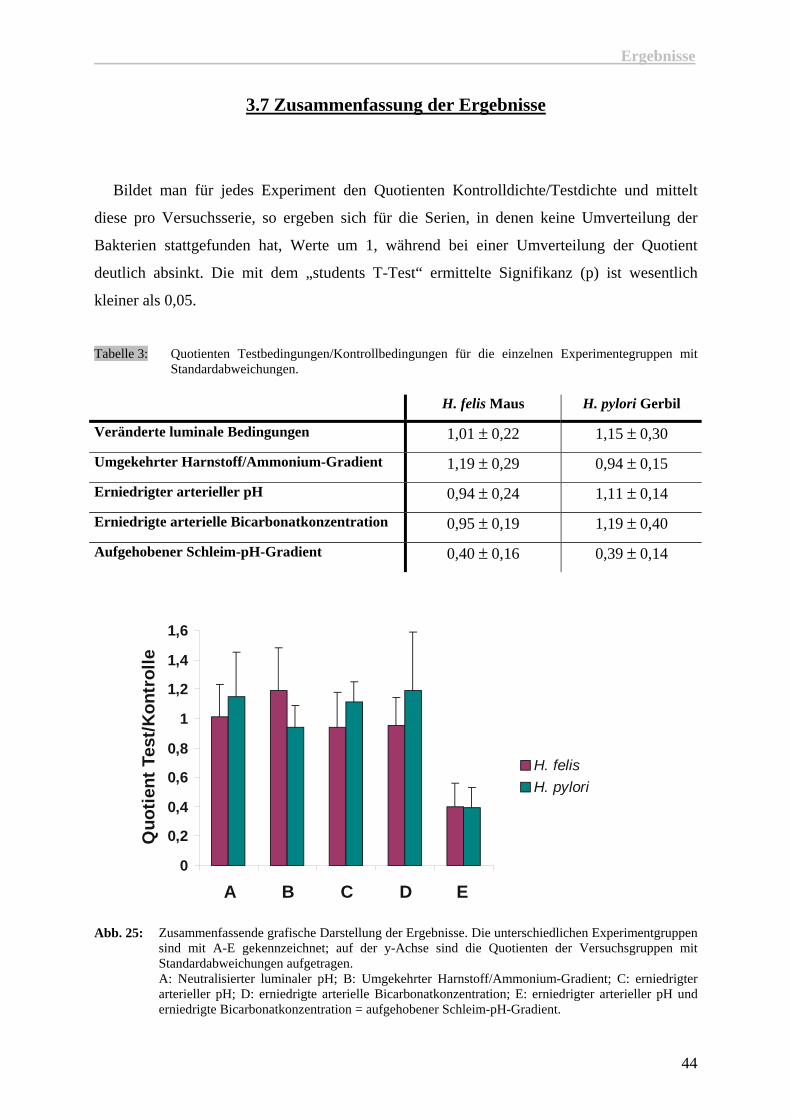
Bildet man für jedes Experiment den Quotienten Kontrolldichte/Testdichte und mittelt
diese pro Versuchsserie, so ergeben sich für die Serien, in denen keine Umverteilung der
Bakterien stattgefunden hat, Werte um 1, während bei einer Umverteilung der Quotient
deutlich absinkt. Die mit dem „students T-Test“ ermittelte Signifikanz (p) ist wesentlich
kleiner als 0,05.
Tabelle 3: Quotienten Testbedingungen/Kontrollbedingungen für die einzelnen Experimentegruppen mit
Standardabweichungen.
H. felis Maus H. pylori Gerbil
Veränderte luminale Bedingungen 1,01 ± 0,22 1,15 ± 0,30
Umgekehrter Harnstoff/Ammonium-Gradient 1,19 ± 0,29 0,94 ± 0,15
Erniedrigter arterieller pH 0,94 ± 0,24 1,11 ± 0,14
Erniedrigte arterielle Bicarbonatkonzentration 0,95 ± 0,19 1,19 ± 0,40
Aufgehobener Schleim-pH-Gradient 0,40 ± 0,16 0,39 ± 0,14
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
A B C D E
Quo
tient
Test
/Kon
trol
le
H. felisH. pylori
Abb. 25: Zusammenfassende grafische Darstellung der Ergebnisse. Die unterschiedlichen Experimentgruppen
sind mit A-E gekennzeichnet; auf der y-Achse sind die Quotienten der Versuchsgruppen mit Standardabweichungen aufgetragen. A: Neutralisierter luminaler pH; B: Umgekehrter Harnstoff/Ammonium-Gradient; C: erniedrigter arterieller pH; D: erniedrigte arterielle Bicarbonatkonzentration; E: erniedrigter arterieller pH und erniedrigte Bicarbonatkonzentration = aufgehobener Schleim-pH-Gradient.

Ergebnisse
45
Die Änderungen des luminalen pH- und der luminalen Bicarbonat/CO2-Konzentrationen,
des Harnstoff/Ammonium-Gradienten, sowie die Absenkung des arteriellen pH bei normaler
Bicarbonatkonzentration, als auch die Absenkung der arteriellen Bicarbonatkonzentration bei
normalem pH, beeinflussen die Orientierung von Helicobacter nicht.
Erst die gleichzeitige Absenkung des arteriellen pH und der Bicarbonatkonzentration, die
zu einer Aufhebung des Schleim-pH-Gradienten führen, erzeugen eine deutliche
Dichteabnahme von Helicobacter im epithelnahen Schleim. Die Bakterien verlieren ihre
Orientierung und verteilen sich über die gesamte Schleimschicht, sowie in das Magenlumen.
Der Schleim-pH-Gradient ist unter physiologischen Bedingungen immer vorhanden und
passt sich an die herrschenden Sekretionsbedingungen an, wobei er sich in dem von
Helicobacter besiedelten Schleimbereich nur minimal ändert. Er eignet sich also besonders
gut als Orientierungsgeber für Helicobacter.
Ergebnisse
46
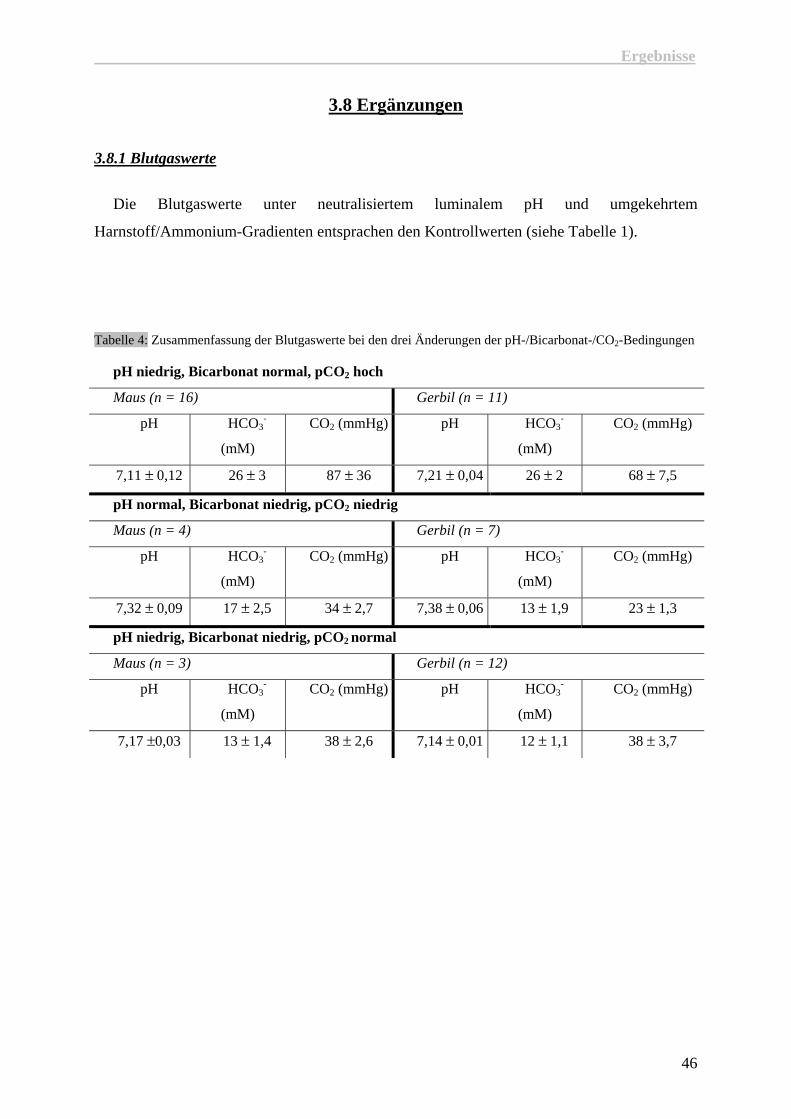
3.8 Ergänzungen
3.8.1 Blutgaswerte
Die Blutgaswerte unter neutralisiertem luminalem pH und umgekehrtem
Harnstoff/Ammonium-Gradienten entsprachen den Kontrollwerten (siehe Tabelle 1).
Tabelle 4: Zusammenfassung der Blutgaswerte bei den drei Änderungen der pH-/Bicarbonat-/CO2-Bedingungen
pH niedrig, Bicarbonat normal, pCO2 hoch
Maus (n = 16) Gerbil (n = 11)
pH HCO3-
(mM)
CO2 (mmHg) pH HCO3-
(mM)
CO2 (mmHg)
7,11 ± 0,12 26 ± 3 87 ± 36 7,21 ± 0,04 26 ± 2 68 ± 7,5
pH normal, Bicarbonat niedrig, pCO2 niedrig
Maus (n = 4) Gerbil (n = 7)
pH HCO3-
(mM)
CO2 (mmHg) pH HCO3-
(mM)
CO2 (mmHg)
7,32 ± 0,09 17 ± 2,5 34 ± 2,7 7,38 ± 0,06 13 ± 1,9 23 ± 1,3
pH niedrig, Bicarbonat niedrig, pCO2 normal
Maus (n = 3) Gerbil (n = 12)
pH HCO3-
(mM)
CO2 (mmHg) pH HCO3-
(mM)
CO2 (mmHg)
7,17 ±0,03 13 ± 1,4 38 ± 2,6 7,14 ± 0,01 12 ± 1,1 38 ± 3,7

Diskussion
47
4. DISKUSSION
Die Möglichkeiten pathogene Bakterien in ihrem Habitat in vivo zu beobachten, sind sehr
begrenzt. In dieser Arbeit wurde eine Nanoprobennahme-Technologie verwendet (Schreiber,
1998), die es ermöglicht, das Verhalten des humanpathogenen Keims H. pylori, sowie der eng
verwandten Spezies H. felis in ihrem Habitat, der Magen-Mucosa in vivo zu studieren. Um
das chemotaktische Verhalten von Helicobacter im Magenschleim in vivo zu studieren,
wurden als Tiermodelle die H. felis infizierte Maus und der H. pylori infizierte Gerbil
eingesetzt.
Die Experimente wurden zuerst an der H. felis infizierten Maus durchgeführt. Dieses
Tiermodell ist im Labor leicht zu handhaben, da keine Übertragbarkeit von H. felis auf den
Menschen beschrieben ist. Zudem liegen in unserem Labor Erfahrung mit diesem Modell vor,
siehe (Schreiber, 1999).
Um eine höhere Übertragbarkeit der Ergebnisse auf die humane Helicobacter Infektion zu
erlangen, wurden die Experimente zusätzlich an dem mit H. pylori infizierten Gerbil
durchgeführt. Der Gerbilmagen ähnelt in seiner Anatomie und Physiologie dem menschlichen
Magen mehr als der der Maus (Hirayama, 1996). Beispielsweise findet man im Gerbilmagen
wie beim Menschen basal pH Werte bis unter 1 während im Magenlumen der Maus der
Basal-pH um 3 liegt.
Die Ergebnisse zeigen, dass sich das chemotaktische Verhalten von H. felis in der Maus
und H. pylori im Gerbil nicht unterscheidet, in beiden Modellen verloren die Bakterien ihre
Orientierung nur bei einem aufgehobenen Schleim-pH-Gradienten.
Diese Ähnlichkeit in der Chemotaxis der beiden Spezies H. felis und H. pylori stützt die
Annahme, dass die hier erhobenen Daten auf die humane H. pylori Infektion übertragen
werden können.
Um das chemotaktische Verhalten von Helicobacter im Magenschleim in vivo zu
untersuchen, wurden die Bedingungen im Magenlumen und im arteriellen Blut bei den
narkotisierten Tieren, gezielt durch die luminale Superfusion, die inspiratorischen
Gasfraktionen und die Peritonealdialyse verändert (siehe Abb. 11, Kap. 2.7.3).

Diskussion
48
Weder die Neutralisierung des luminalen pH von sauren Werten auf 6 noch die
Umkehrung des Harnstoff/Ammonium-Gradienten im Magenschleim, beeinflussen die
Verteilung von H. felis in der Maus oder H. pylori im Gerbil, verglichen mit der
Bakterienverteilung unter Kontroll-Bedingungen.
Die Absenkung des arteriellen pH oder der arteriellen Bicarbonat/CO2-Konzentration
ändern ebenfalls nicht das Verhalten von H. felis und H. pylori im Magenschleim von Maus
und Gerbil.
Erst die gleichzeitige Reduktion des arteriellen pH und der arteriellen
Bicarbonatkonzentration und die daraus resultierende Eliminierung des Mucus-pH-Gradienten
führen zur Desorientierung von H. felis und H. pylori. Sie befinden sich nur noch in
reduzierter Dichte in ihrer ökologischen Nische, dem epithelnahen Schleim.
Der Zusammenhang von Bicarbonat, CO2 und pH
Bicarbonat und CO2 sind das höchstkonzentrierte Pufferpaar im Blut und den Organen. Als
Säure/Basenpaar sind sie über die Henderson/Hasselbalch Gleichung mit dem pH verknüpft.
Es wurden drei Konstellationen der drei Größen untersucht:
1. Eine Erniedrigung des pH durch einen hohen inspiratorischen pCO2, bei durch Dialyse
normaler Plasmakonzentration von Bicarbonat.
2. Eine Erniedrigung der Bicarbonatkonzentration durch Hyperventilation mit
entsprechender Erniedrigung des arteriellen pCO2, bei durch Dialyse normalem pH.
3. Eine Erniedrigung der Bicarbonatkonzentration und des arteriellen pH durch Dialyse bei
konstanter Ventilation und damit normalem pCO2.
In den in vitro Untersuchungen am Meerschwein konnten wir zeigen, dass ausschließlich
die dritte Bedingung zu einer Aufhebung des Schleim-pH-Gradienten führt. Genau diese
dritte Bedingung war es auch, die allein zu einer Störung der Helicobacter Orientierung
geführt hat. Die in den Bedingungen 1 und 2 auftretenden ausgeprägten Änderungen des
pCO2 und der Bicarbonatkonzentration haben weder zu einer Reaktion an der Mucosa, noch
zu einer Orientierungsstörung von Helicobacter geführt. Wir glauben daher, bewiesen zu

Diskussion
49
haben, dass es der gewebenahe pH-Gradient ist, der die Orientierung von Helicobacter
ermöglicht.
Wie registriert Helicobacter den pH?
Helicobacter muss über Rezeptoren verfügen, welche es ihm erlauben, den Umgebungs-
pH direkt zu registrieren. Die Erkennung von pH- oder Protonen-Gradienten ist bei anderen
Bakterien als Teil der „Energy taxis“ beschrieben. Unter der „Energy taxis“ werden die
Aerotaxis, die Phototaxis, die Redoxtaxis sowie die Chemotaxis von alternativen
Energielieferanten und Kohlenstoffquellen zusammengefasst.
Der Mechanismus der Energiestoff-Erkennung und der daraus resultierenden gerichteten
Bewegung (Taxis), ist bis heute noch nicht vollständig aufgeklärt, aber es ist bekannt, dass
chemotaktische Signalkaskaden eine wichtige Rolle spielen. Die dabei benutzten Proteine
können sehr unterschiedlich aufgebaut sein.
Einige Proteine enthalten als Gemeinsamkeit die sogenannte PAS-Domäne6 , einen
Genabschnitt, der erstmals bei drei Proteinen von Drosophila gefunden wurde (Taylor, 1999).
Die auf die Aktivierung eines Chemorezeptors folgenden Signalkaskaden können
wahrscheinlich unterschiedlich ausfallen. Es können unter anderem Häm-enthaltende Sensor-
Proteine, sogenannte Transducer, zwischengeschaltet sein (Hou, 2001).
Helicobacter besitzt drei klassische membrangebundene Chemorezeptoren sowie einen
mutmaßlichen cytoplasmatischen Chemorezeptor (HP599) (Tomb, 1997), welcher begrenzte
Homologien zu den Häm-ähnlichen Sensor Proteinen, nicht aber zu den PAS-Sensor
Proteinen aufweist.
Das Genom von Helicobacter weist außerdem Homologien zu den, in die chemotaktischen
Signalkaskaden von Salmonella enterica und Escherichia coli involvierten, Proteinen CheA,
CheW und CheY auf (Pittman, 2001; Beier, 1997; Alm, 1999).
Die Registrierung des extrazellulären pH bei Helicobacter könnte so ablaufen, dass die
Protonenkonzentration von einem Chemorezeptor erkannt wird. Daraufhin kommt es zur
Aktivierung der Autokinase CheA, welche rezeptorgekoppelt ist. Weiter wird über eine
intrazelluläre Signalkaskade, an der wahrscheinlich auch CheW beteiligt ist, die Information
6 PAS steht für drei Proteine von Drosophila, bei denen dieser Genabschnitt erstmals gefunden wurde, nämlich PER, AINT und SIM

Diskussion
50
an den Flagellenmotor weitergeleitet. CheY interagiert direkt mit dem Flagellenmotor und es
folgt die gerichtete Bewegung.
Da Helicobacter mehrere Chemorezeptoren besitzt, ist es vorstellbar, dass es neben dem
pH weitere Substanzen im Magenschleim gibt, die unabhängig von der räumlichen
Orientierung das chemotaktisches Verhalten beeinflussen.
Bakteriendichte in vivo
Trotz der Übereinstimmung im chemotaktischen Verhalten von H. felis und H. pylori gibt es
einige Unterschiede zwischen den beiden Spezies.
Die hier erzielten Ergebnisse zeigen, dass H. pylori und H. felis sich in ihrem
Kolonisationsverhalten unterscheiden. H. pylori ist in der Lage, an die Magenepithelzellen zu
adhärieren und besiedelt eine näher an der epithelialen Oberfläche liegende Schleimschicht
als H. felis. Diese Tatsache könnte dazu beitragen, den Austausch zwischen adhärierenden
und schwimmenden H. pylori-Populationen zu erleichtern. Zudem könnte der Besiedlungsort
im Magenschleim mit entscheidend für die Virulenz des Bakteriums sein. H. felis adhäriert
nicht an die Magenepithelzellen (Enno, 1995), hält sich etwas weiter vom Magenepithel
entfernt auf und erzeugt in der Maus keine Geschwüre, Lymphome oder Karzinome (Enno,
1995).
Die unerwartet hohen Bakteriendichten, die sich durch die sehr schmale Schleimschicht
ergeben, welche von den Bakterien besiedelt wird, könnten den genetischen Austausch
zwischen verschiedenen Helicobacter Stämmen fördern und die große genetische Variabilität
von Helicobacter erklären.

Diskussion
51
Triple-Therapie und Orientierung von Helicobacter
Die hier vorgestellten Ergebnisse könnten dabei helfen, die Rolle der
Protonenpumpenblocker in der Therapie der humanen H. pylori-Infektion besser zu verstehen.
Momentan werden mit H. pylori infizierte Patienten mit einer Kombination aus einem
Protonenpumpenblocker (PPI) sowie zwei hochdosierten Antibiotika behandelt (Axon, 1996).
Die Protonenpumpenblocker wirken auf die Parietalzellen im Corpus des Magens. Sie
blockieren die Protonenpumpen in den Canaliculi.
Aufgrund der hier vorgestellten Ergebnisse erscheint es unwahrscheinlich, dass die
dadurch ausgelöste luminale Neutralisierung einen direkten Einfluss auf die Bakterien im
epithelnahen Schleim des Antrums haben könnte. Bei neutralisiertem luminalem pH hat sich
die Orientierung von H. felis und H. pylori nicht verändert (Kapitel 3.5.1). Dieses Ergebnis
wird noch dadurch unterstützt, dass die alleinige Verabreichung eines PPI nicht zur
Helicobacter-Eliminierung führt (Hunt, 1993).
Aus in vitro Experimenten weiß man, dass die PPI zusätzlich die Motilität von
Helicobacter inhibieren (Tsutsui, 2000), jedoch ist dieser Effekt sehr gering und unterscheidet
sich in seiner Wirksamkeit unter den einzelnen auf dem Markt verfügbaren Produkten.
Beispielsweise ist er für das sehr wirksame Omeprazol besonders gering. Somit ist die
Erklärung, dass die PPI durch die verminderte Bakterienmotilität wirken können, ebenfalls
nicht zufriedenstellend.
Eine mögliche Erklärung für den synergistischen Effekt von PPI und Antibiotika ist, dass
die gehemmte Säuresekretion im Corpus die Bicarbonat- und pH-Regulation im Antrum stört.
Dies könnte zur Störung der Antrum-pH-Gradienten, und damit zum Verlust der Orientierung
von Helicobacter führen.
Ein präzises Verständnis der ökologischen Nische von Helicobacter ist Vorraussetzung,
um Hypothesen zur PPI-Wirkung zu überprüfen und neue, effektivere Wege zur Therapie der
Helicobacter-Infektion entwickeln zu können.

Literaturverzeichnis
52
LITERATURLISTE 1. Alm, R. A., Ling, L. S., Moir, D. T., King, B. L., Brown, E. D., Doig, P. C., Smith, D.
R., Noonan, B., Guild, B. C., deJonge, B. L., Carmel, G., Tummino, P. J., Caruso, A.,
Uria-Nickelsen, M., Mills, D. M., Ives, C., Gibson, R., Merberg, D., Mills, S. D.,
Jiang, Q., Taylor, D. E., Vovis, G. F., and Trust, T. J. (1999). Genomic-sequence
comparison of two unrelated isolates of the human gastric pathogen Helicobacter
pylori. Nature 397, 176-180.
2. Ashley, S. W., Soybel, D. I., Moore, C. D., and Cheung, L. Y. (1987). Intracellular pH
(pHi) in gastric surface epithelium is more susceptible to serosal than mucosal
acidification. Surgery 102, 371-379.
3. Atherton, J. C., Cao, P., Peek, R. M. J., Tummuru, M. K., Blaser, M. J., and Cover, T.
L. (1995). Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori.
Association of specific vacA types with cytotoxin production and peptic ulceration. J
Biol Chem 270, 17771.
4. Axon, A. T. and Moayyedi, P. (1996). Eradication of Helicobacter pylori: omeprazole
in combination with antibiotics. Scand J Gastroenterol Suppl 215, 82-89.
5. Bebelman, J. P., Evers, M. P., Zelle, B., Bank, R., Pronk, J. C., Meuwissen, S. G.,
Mager, W. H., Planta, R. J., Eriksson, A. W., and Frants, R. R. (1989). Family and
population studies on the human pepsinogen A multigene family. Hum Genet 82, 142-
146.
6. Beier, D., Spohn, G., Rappuoli, R., and Scarlato, V. (1997). Identification and
characterization of an operon of Helicobacter pylori that is involved in motility and
stress adaptation. J Bacteriol 179, 4676-4683.
7. Blaser, M. J. (1996). Role of vacA and the cagA locus of Helicobacter pylori in human
disease. Aliment Pharmacol Ther 10 (suppl 1), 73-77.
8. Borén, T., Falk, P., Roth, K. A., Larson, G., and Normark, S. (1993). Attachment of
Helicobacter pylori to human gastric epithelium mediated by blood group antigens
[see comments]. Science 262, 1892-1895.

Literaturverzeichnis
53
9. Bren, A. and Eisenbach, M. (2000). How signals are heard during bacterial
chemotaxis: protein-protein interactions in sensory signal propagation. J Bacteriol
182, 6865-6873.
10. Bronsdon, M. A., Goodwin, C. S., Sly, L. I., Chilvers, T., and Schoenknecht, F. D.
(1991). Helicobacter nemestrinae sp. nov., a spiral bacterium found in the stomach of
a pigtailed macaque (Macaca nemestrina) [see comments]. Int J Syst Bacteriol 41,
148-153.
11. Censini, S., Lange, C., Xiang, Z., Crabtree, J. E., Ghiara, P., Borodovsky, M.,
Rappuoli, R., and Covacci, A. (1996). Cag, a pathogenicity island of Helicobacter
pylori, encodes type I-specific and disease-associated virulence factors. Proc Natl
Acad Sci U S A 93, 14648-14653.
12. Cover, T. L., Hanson, P. I., and Heuser, J. E. (1997). Acid-induced dissociation of
VacA, the Helicobacter pylori vacuolating cytotoxin, reveals its pattern of assembly. J
Cell Biol 138, 759-769.
13. Debets-Ossenkopp Y.J., Sparrius M., Kusters J.G., Kolkman J.J., and Vandenbroucke-
Grauls C.M.J.E. (1996). Mechanism of clarithromycin resistance in clinical isolates of
Helicobacter pylori. FEMS Microbiology letters 142, 37-42.
14. Dewhirst, F. E., Seymour, C., Fraser, G. J., Paster, B. J., and Fox, J. G. (1994).
Phylogeny of Helicobacter isolates from bird and swine feces and description of
Helicobacter pametensis sp. nov.. Int J Syst Bacteriol 44, 553-560.
15. Dixon M.F. (1990). Helicobacter pylori and peptic ulceration: Histopathological
aspects. Jornal of Gastroenterology & Hepatology 6, 125-130.
16. Dixon M.F. and Ectors N.L. (1995). Gastric cancer. Current opinion in
gastroenterology 11 (suppl 1), 38-41.
17. Eaton, K. A., Dewhirst, F. E., Radin, M. J., Fox, J. G., Paster, B. J., Krakowka, S., and
Morgan, D. R. (1993). Helicobacter acinonyx sp. nov., isolated from cheetahs with
gastritis. Int J Syst Bacteriol 43, 99-106.

Literaturverzeichnis
54
18. Eaton, K. A., Suerbaum, S., Josenhans, C., and Krakowka, S. (1996). Colonization of
gnotobiotic piglets by Helicobacter pylori deficient in two flagellin genes. Infect
Immun 64, 2445-2448.
19. Enno, A., O'Rourke, J. L., Howlett, C. R., Jack, A., Dixon, M. F., and Lee, A. (1995).
MALToma-like lesions in the murine gastric mucosa after long-term infection with
Helicobacter felis. A mouse model of Helicobacter pylori-induced gastric lymphoma.
Am J Pathol 147, 217-222.
20. Ernst, P. B. and Gold, B. D. (1999). Helicobacter pylori in childhood: new insights
into the immunopathogenesis of gastric disease and implications for managing
infection in children. J Pediatr Gastroenterol Nutr 28, 462-473.
21. Evers, M. P., Zelle, B., Bebelman, J. P., van, B., V, Kraakman, L., Hoffer, M. J.,
Pronk, J. C., Mager, W. H., Planta, R. J., Eriksson, A. W., and . (1989). Nucleotide
sequence comparison of five human pepsinogen A (PGA) genes: evolution of the PGA
multigene family. Genomics 4, 232-239.
22. Ferrero, R. L. and Lee, A. (1991). The importance of urease in acid production for the
gastric-colonising bacteria Helicobacter pylori and Heliocbacter felis sp. nov..
Microbial ecology in health and disease 4, 121-134.
23. Fiocca R., Villani L., De Giacomo C., Perego M, Trespi E, and Solcia E. (1989).
Morphological evidence of campylobacter pylori pathogenicity in chronic gastritis and
peptic ulcer. Acta Gastro-Enterologica Beligica 52, 324-335.
24. Fiocca, R., Villani, L., Luinetti, O., Gianatti, A., Perego, M., Alvisi, C., Turpini, F.,
and Solcia, E. (1992). Helicobacter colonization and histopathological profile of
chronic gastritis in patients with or without dyspepsia, mucosal erosion and peptic
ulcer: a morphological approach to the study of ulcerogenesis in man. Virchows Arch
A Pathol Anat Histopathol 420, 489-498.
25. Forman, D., Newell, D. G., Fullerton, F., Yarnell, J. W., Stacey, A. R., Wald, N., and
Sitas, F. (1991). Association between infection with Helicobacter pylori and risk of
gastric cancer: evidence from a prospective investigation [see comments]. BMJ 302,
1302-1305.

Literaturverzeichnis
55
26. Fox, J. G., Correa, P., Taylor, N. S., Lee, A., Otto, G., Murphy, J. C., and Rose, R.
(1990). Helicobacter mustelae-associated gastritis in ferrets. An animal model of
Helicobacter pylori gastritis in humans. Gastroenterology 99, 352-361.
27. Foynes, S., Dorrell, N., Ward, S. J., Stabler, R. A., McColm, A. A., Rycroft, A. N.,
and Wren, B. W. (2000). Helicobacter pylori possesses two CheY response regulators
and a histidine kinase sensor, CheA, which are essential for chemotaxis and
colonization of the gastric mucosa. Infect Immun 68, 2016-2023.
28. Geis, G., Suerbaum, S., Forsthoff, B., Leying, H., and Opferkuch, W. (1993).
Ultrastructure and biochemical studies of the flagellar sheath of Helicobacter pylori. J
Med Microbiol 38, 371-377.
29. Ghiara, P., Marchetti, M., Blaser, M. J., Tummuru, M. K., Cover, T. L., Segal, E. D.,
Tompkins, L. S., and Rappuoli, R. (1995). Role of the Helicobacter pylori virulence
factors vacuolating cytotoxin, CagA, and urease in a mouse model of disease. Infect
Immun 63, 4154-4160.
30. Goodman, K. J. and Correa, P. (1995). The transmission of Helicobacter pylori. A
critical review of the evidence. Int J Epidemiol 24, 875-887.
31. Goodwin, C. S., Armstrong J.A., Chilvers T., Peters M., Collins M.D., Sly L.,
Mcdonnell W., and Harper W.E.S. (1989). Transfer of campylobacter pylori and
campylobacter mustelae to helicobacter gen. nov. as helicobacter comb. nov. and
helicobacter mustelae comb. nov., respectively. Int J Systematic Bac 397-405.
32. Graham, D. Y., Adam E., Reddy G.T., Agarwal P.J., Agarwal R., Evans jr.D.E.,
Malaty H.M., and Evans, D. G. (1991). Seroepidemiology of helicobacter pylori
infection in India - Comparison of developing and developed countries. Digestive
diseases and sciences 36, 1084-1088.
33. Graham, D. Y., Klein, P. D., Evans, D. J. J., Evans, D. G., Alpert, L. C., Opekun, A.
R., and Boutton, T. W. (1987). Campylobacter pylori detected noninvasively by the
13C-urea breath test. Lancet 1, 1174-1177.

Literaturverzeichnis
56
34. Han, S. R., Zschausch, H. C., Meyer, H. G., Schneider, T., Loos, M., Bhakdi, S., and
Maeurer, M. J. (2000). Helicobacter pylori: clonal population structure and restricted
transmission within families revealed by molecular typing. J Clin Microbiol 38, 3646-
3651.
35. Harper, C. G., Feng, Y., Xu, S., Taylor, N. S., Kinsel, M., Dewhirst, F. E., Paster, B.
J., Greenwell, M., Levine, G., Rogers, A., and Fox, J. G. (2002). Helicobacter cetorum
sp. nov., a urease-positive Helicobacter species isolated from dolphins and whales. J
Clin Microbiol 40, 4536-4543.
36. Hirayama, F., Takagi, S., Yokoyama, Y., Iwao, E., and Ikeda, Y. (1996).
Establishment of gastric Helicobacter pylori infection in Mongolian gerbils. J
Gastroenterol 31 Suppl 9, 24-28.
37. Honda, S., Fujioka, T., Tokieda, M., Gotoh, T., Nishizono, A., and Nasu, M. (1998).
Gastric ulcer, atrophic gastritis, and intestinal metaplasia caused by Helicobacter
pylori infection in Mongolian gerbils. Scand J Gastroenterol 33, 454-460.
38. Hou, S., Freitas, T., Larsen, R. W., Piatibratov, M., Sivozhelezov, V., Yamamoto, A.,
Meleshkevitch, E. A., Zimmer, M., Ordal, G. W., and Alam, M. (2001). Globin-
coupled sensors: a class of heme-containing sensors in Archaea and Bacteria. Proc
Natl Acad Sci U.S.A 98, 9353-9358.
39. Hunt, R. H. (1993). Hp and pH: implications for the eradication of Helicobacter
pylori. Scand J Gastroenterol Suppl 196, 12-16.
40. Hussell, T., Isaacson, P. G., Crabtree, J. E., and Spencer, J. (1993). The response of
cells from low-grade B-cell gastric lymphomas of mucosa-associated lymphoid tissue
to Helicobacter pylori [see comments]. Lancet 342, 571-574.
41. IARC (1999). Shistosomes, liver flukes and helicobacter pylori. WHO Publications
61, 177-240.
42. Ikeno, T., Ota, H., Sugiyama, A., Ishida, K., Katsuyama, T., Genta, R. M., and
Kawasaki, S. (1999). Helicobacter pylori-induced chronic active gastritis, intestinal
metaplasia, and gastric ulcer in Mongolian gerbils. Am J Pathol 154, 951-960.

Literaturverzeichnis
57
43. Ilver, D., Arnqvist, A., Ogren, J., Frick, I. M., Kersulyte, D., Incecik, E. T., Berg, D.
E., Covacci, A., Engstrand, L., and Boren, T. (1998). Helicobacter pylori adhesin
binding fucosylated histo-blood group antigens revealed by retagging. Science 279,
373-377.
44. Josenhans, C., Labigne, A., and Suerbaum, S. (1995). Comparative ultrastructural and
functional studies of Helicobacter pylori and Helicobacter mustelae flagellin mutants:
both flagellin subunits, FlaA and FlaB, are necessary for full motility in Helicobacter
species. J Bacteriol 177, 3010-3020.
45. Josenhans, C. and Suerbaum, S. (2002). The role of motility as a virulence factor in
bacteria. Int J Med Microbiol 291, 605-614.
46. Karim, Q. N., Logan, R. P., Puels, J., Karnholz, A., and Worku, M. L. (1998).
Measurement of motility of Helicobacter pylori, Campylobacter jejuni, and
Escherichia coli by real time computer tracking using the Hobson BacTracker. J Clin
Pathol 51, 623-628.
47. Kauffman, G. L., Jr. (1980). Gastric bicarbonate secretion: an update. Brain Res Bull 5
(suppl 1), 15-18.
48. Kavermann, H., Burns, B. P., Angermuller, K., Odenbreit, S., Fischer, W., Melchers,
K., and Haas, R. (2003). Identification and characterization of Helicobacter pylori
genes essential for gastric colonization. J Exp Med 197, 813-822.
49. Kihara, M. and Macnab, R. M. (1981). Cytoplasmic pH mediates pH taxis and weak-
acid repellent taxis of bacteria. J Bacteriol 145, 1209-1221.
50. Kim, H., Park, C., Jang, W. I., Lee, K. H., Kwon, S. O., Robey-Cafferty, S. S., Ro, J.
Y., and Lee, Y. B. (1990). The gastric juice urea and ammonia levels in patients with
Campylobacter pylori. Am J Clin Pathol 94, 187-191.
51. Kivilaakso, E. (1981). High plasma HCO3--protects gastric mucosa against acute
ulceration in the rat. Gastroenterology 81, 921-927.
52. Labenz J. and O'Morain C. (1995). Eradication. Current opinion in gastroenterology
11 (suppl 1), 47-51.

Literaturverzeichnis
58
53. Lee, A., Fox, J. G., Otto, G., and Murphy, J. (1990). A small animal model of human
Helicobacter pylori active chronic gastritis. Gastroenterology 99, 1315-1323.
54. Lee, A., Hazell, S. L., O'Rourke, J., and Kouprach, S. (1988). Isolation of a spiral-
shaped bacterium from the cat stomach. Infect Immun 56, 2843-2850.
55. Lee, A., O'Rourke, J., De Ungria, M. C., Robertson, B., Daskalopoulos, G., and
Dixon, M. F. (1997). A standardized mouse model of Helicobacter pylori infection:
introducing the Sydney strain [published erratum appears in Gastroenterology 1997
Aug;113(2):732]. Gastroenterology 112, 1386-1397.
56. Malaty, H. M. and Graham, D. Y. (1994). Importance of childhood socioeconomic
status on the current prevalence of Helicobacter pylori infection. Gut 35, 742-745.
57. Malaty, H. M., Graham, D. Y., Wattigney, W. A., Srinivasan, S. R., Osato, M., and
Berenson, G. S. (1999). Natural history of Helicobacter pylori infection in childhood:
12-year follow-up cohort study in a biracial community. Clin Infect Dis 28, 279-282.
58. Marshall B.J. (1986). Campylobacter pyloridis and gastritis. The Journal of infectios
disease 153, 650-669.
59. Matsumoto, M., Kimura, K., Fujisawa, A., Matsuyama, T., Asai, T., Uyama, O.,
Yoneda, S., and Abe, H. (1982). Regional blood flows measured in Mongolian gerbil
by a modified microsphere method. Am J Physiol 242, H990-H995.
60. Matsumoto, S., Washizuka, Y., Matsumoto, Y., Tawara, S., Ikeda, F., Yokota, Y., and
Karita, M. (1997). Induction of ulceration and severe gastritis in Mongolian gerbil by
Helicobacter pylori infection. J Med Microbiol 46, 391-397.
61. Megraud, F., Brassens-Rabbe, M. P., Denis, F., Belbouri, A., and Hoa, D. Q. (1989).
Seroepidemiology of Campylobacter pylori infection in various populations. J Clin
Microbiol 27, 1870-1873.
62. Mendall M.A., Goggin P.M., Molineaux N., Levy S., Toosy T., Strachan D., and
Northfield T.C. (1992). Childhood living conditions and helicobacter pylori
seropositivity in adult life. The Lancet 339, 896-897.

Literaturverzeichnis
59
63. Miehlke, S., Yu, J., Schuppler, M., Frings, C., Kirsch, C., Negraszus, N., Morgner, A.,
Stolte, M., Ehninger, G., and Bayerdorffer, E. (2001). Helicobacter pylori vacA, iceA,
and cagA status and pattern of gastritis in patients with malignant and benign
gastroduodenal disease. Am J Gastroenterol 96, 1008-1013.
64. Mitchell, H. M., Li, Y. Y., Hu, P. J., Liu, Q., Chen, M., Du, G. G., Wang, Z. J., Lee,
A., and Hazell, S. L. (1992). Epidemiology of Helicobacter pylori in southern China:
identification of early childhood as the critical period for acquisition. J Infect Dis 166,
149-153.
65. Mizote, T., Yoshiyama, H., and Nakazawa, T. (1997). Urease-independent
chemotactic responses of Helicobacter pylori to urea, urease inhibitors, and sodium
bicarbonate. Infect Immun 65, 1519-1521.
66. Mobley, H. L., Hu, L. T., and Foxal, P. A. (1991). Helicobacter pylori urease:
properties and role in pathogenesis. Scand J Gastroenterol Suppl 187, 39-46.
67. Mobley, H. L., Island, M. D., and Hausinger, R. P. (1995). Molecular biology of
microbial ureases. Microbiol Rev 59, 451-480.
68. Mohammadi, M., Redline, R., Nedrud, J., and Czinn, S. (1996). Role of the host in
pathogenesis of Helicobacter-associated gastritis: H. felis infection of inbred and
congenic mouse strains. Infect Immun 64, 238-245.
69. Nagano, F., Masaki, H., Fujimoto, M., Kotera, K., Yonehara, T., and Ohshiba, S.
(1990). Effects of H+ and HCO3- secretion on mucus gel pH in isolated antral mucosa
of bullfrog stomach. Bull Osaka Med Coll 36, 13-25.
70. Nakamura, H., Yoshiyama, H., Takeuchi, H., Mizote, T., Okita, K., and Nakazawa, T.
(1998). Urease plays an important role in the chemotactic motility of Helicobacter
pylori in a viscous environment. Infect Immun 66, 4832-4837.
71. Odenbreit, S., Faller, G., and Haas, R. (2002). Role of the alpAB proteins and
lipopolysaccharide in adhesion of Helicobacter pylori to human gastric tissue. Int J
Med Microbiol 292, 247-256.
72. Ottemann, K. M. and Lowenthal, A. C. (2002). Helicobacter pylori uses motility for
initial colonization and to attain robust infection. Infect Immun 70, 1984-1990.

Literaturverzeichnis
60
73. Parsonnet, J., Friedman, G. D., Orentreich, N., and Vogelman, H. (1997). Risk for
gastric cancer in people with CagA positive or CagA negative Helicobacter pylori
infection [see comments]. Gut 40, 297-301.
74. Parsonnet, J., Friedman, G. D., Vandersteen, D. P., Chang, Y., Vogelman, J. H.,
Orentreich, N., and Sibley, R. K. (1991). Helicobacter pylori infection and the risk of
gastric carcinoma [see comments]. N Engl J Med 325, 1127-1131.
75. Parsonnet, J., Shmuely, H., and Haggerty, T. (1999). Fecal and oral shedding of
Helicobacter pylori from healthy infected adults. JAMA 282, 2240-2245.
76. Paster, B. J., Lee, A., Fox, J. G., Dewhirst, F. E., Tordoff, L. A., Fraser, G. J.,
O'Rourke, J. L., Taylor, N. S., and Ferrero, R. (1991). Phylogeny of Helicobacter felis
sp. nov., Helicobacter mustelae, and related bacteria. Int J Syst Bacteriol 41, 31-38.
77. Pittman, M. S., Goodwin, M., and Kelly, D. J. (2001). Chemotaxis in the human
gastric pathogen Helicobacter pylori: different roles for CheW and the three CheV
paralogues, and evidence for CheV2 phosphorylation. Microbiology 147, 2493-2504.
78. Rees, W. D., Botham, D., and Turnberg, L. A. (1982). A demonstration of bicarbonate
production by the normal human stomach in vivo. Dig Dis Sci 27, 961-966.
79. Ross, I. N., Bahari, H. M., and Turnberg, L. A. (1981). The pH gradient across mucus
adherent to rat fundic mucosa in vivo and the effect of potential damaging agents.
Gastroenterology 81, 713-718.
80. Rothenbacher, D., Winkler, M., Gonser, T., Adler, G., and Brenner, H. (2002). Role of
infected parents in transmission of helicobacter pylori to their children. Pediatr Infect
Dis J 21, 674-679.
81. Rotter, J. I., Sones, J. Q., Samloff, I. M., Richardson, C. T., Gursky, J. M., Walsh, J.
H., and Rimoin, D. L. (1979). Duodenal-ulcer disease associated with elevated serum
pepsinogen I: an inherited autosomal dominant disorder. N Engl J Med 300, 63-66.
82. Ruggiero, P., Peppoloni, S., Berti, D., Rappuoli, R., and Giudice, G. D. (2002). New
strategies for the prevention and treatment of Helicobacter pylori infection. Expert
Opin Investig Drugs 11, 1127-1138.

Literaturverzeichnis
61
83. Rune, S. J. and Henriksen, F. W. (1969). Carbon dioxide tensions in the proximal part
of the canine gastrointestinal tract. Gastroenterology 56, 758-762.
84. Schreiber, S., Nguyen, T. H., Stüben, M., and Scheid, P. (2000). Demonstration of a
pH gradient in the gastric gland of the acid-secreting guinea pig mucosa. Am J Physiol
279, G597-G604.
85. Schreiber, S. and Scheid, P. (1997). Gastric mucus of the guinea pig: proton carrier
and diffusion barrier. Am J Physiol 272, G63-G70.
86. Schreiber, S., Stüben, M., Josenhans, C., Scheid, P., and Suerbaum, S. (1999). In vivo
distribution of Helicobacter felis in the gastric mucus of the mouse: experimental
method and results. Infect Immun 67, 5151-5156.
87. Schreiber, S., Stüben, M., Kröhan, P., and Scheid, P. (1998). Micropipette system for
sampling and injecting small volumes. J Neurosci Methods 85, 27-32.
88. Scott, D. R., Marcus, E. A., Weeks, D. L., and Sachs, G. (2002). Mechanisms of acid
resistance due to the urease system of Helicobacter pylori. Gastroenterology 123, 187-
195.
89. Sommi, P., Ricci, V., Fiocca, R., Romano, M., Ivey, K. J., Cova, E., Solcia, E., and
Ventura, U. (1996). Significance of ammonia in the genesis of gastric epithelial
lesions induced by Helicobacter pylori: an in vitro study with different bacterial strains
and urea concentrations. Digestion 57, 299-304.
90. Suerbaum, S. (1995). The complex flagella of gastric Helicobacter species. Trends
Microbiol 3, 168-161.
91. Suerbaum, S. and Michetti, P. (2002). Helicobacter pylori infection. N Engl J Med
347, 1175-1186.
92. Taggart, R. T., Karn, R. C., Merritt, A. D., Yu, P. L., and Conneally, P. M. (1979).
Urinary pepsinogen isozymes: a highly polymorphic locus in man. Hum Genet 52,
227-238.
93. Talley, N. J., Ormand, J. E., Frie, C. A., and Zinsmeister, A. R. (1992). Stability of pH
gradients in vivo across the stomach in Helicobacter pylori gastritis, dyspepsia, and
health. Am J Gastroenterol 87, 590-594.

Literaturverzeichnis
62
94. Taylor, B. L., Zhulin, I. B., and Johnson, M. S. (1999). Aerotaxis and other energy-
sensing behavior in bacteria. Annu Rev Microbiol 53, 103-128.
95. The Eurogast study group (1993a). An international association between helicobacter
pylori infection and gastric cancer. The Lancet 341, 1359-1362.
96. The Eurogast study group (1993b). Epidemiology of, and risk factors for, Helicobacter
pylori infection among 3194 asymptomatic subjects in 17 populations. The
EUROGAST Study Group [see comments]. Gut 34, 1672-1676.
97. Tomb, J. F., White, O., Kerlavage, A. R., Clayton, R. A., Sutton, G. G., Fleischmann,
R. D., Ketchum, K. A., Klenk, H. P., Gill, S., Dougherty, B. A., Nelson, K.,
Quackenbush, J., Zhou, L., Kirkness, E. F., Peterson, S., Loftus, B., Richardson, D.,
Dodson, R., Khalak, H. G., Glodek, A., McKenney, K., Fitzegerald, L. M., Lee, N.,
Adams, M. D., and Venter, J. C. (1997). The complete genome sequence of the gastric
pathogen Helicobacter pylori [see comments] [published erratum appears in Nature
1997 Sep 25;389(6649):412]. Nature 388, 539-547.
98. Tsujii, M., Kawano, S., Tsuji, S., Fusamoto, H., Kamada, T., and Sato, N. (1992).
Mechanism of gastric mucosal damage induced by ammonia. Gastroenterology 102,
1881-1888.
99. Tsutsui, N., Taneike, I., Ohara, T., Goshi, S., Kojio, S., Iwakura, N., Matsumaru, H.,
Wakisaka, S., Zhang, H. M., and Yamamoto, T. (2000). A novel action of the proton
pump inhibitor rabeprazole and its thioether derivative against the motility of
Helicobacter pylori. Antimicrob Agents Chemother 44, 3069-3073.
100. Warren J.R. (1983). Unidentified curved bacilli on gastric epithelium in active chronic
gastritis [letter]. Lancet 1, 1273-1275.
101. Watanabe, T., Tada, M., Nagai, H., Sasaki, S., and Nakao, M. (1998). Helicobacter
pylori infection induces gastric cancer in mongolian gerbils. Gastroenterology 115,
642-648.
102. Wirth, H. P., Beins, M. H., Yang, M., Tham, K. T., and Blaser, M. J. (1998).
Experimental infection of Mongolian gerbils with wild-type and mutant Helicobacter
pylori strains. Infect Immun 66, 4856-4866.

Literaturverzeichnis
63
103. Wirth, H. P., Yang, M., Karita, M., and Blaser, M. J. (1996). Expression of the human
cell surface glycoconjugates Lewis x and Lewis y by Helicobacter pylori isolates is
related to cagA status. Infect Immun 64, 4598-4605.
104. Wotherspoon, A. C., Doglioni, C., Diss, T. C., Pan, L., Moschini, A., de Boni, M., and
Isaacson, P. G. (1993). Regression of primary low-grade B-cell gastric lymphoma of
mucosa-associated lymphoid tissue type after eradication of Helicobacter pylori.
Lancet 342, 575-577.
105. Yoshiyama, H., Nakamura, H., Kimoto, M., Okita, K., and Nakazawa, T. (1999).
Chemotaxis and motility of Helicobacter pylori in a viscous environment. J
Gastroenterol 34 (suppl 11), 18-23.

Anhang
64
ZUSAMMENFASSUNG
Helicobacter besiedelt den Magenschleim in unmittelbarer Nähe der epithelialen
Oberfläche. In einer vorangegangenen Untersuchung wurde in unserem Labor das
Verteilungsmuster von H. felis im Magen der Maus in vivo beschrieben.
H. felis siedelt sich in einem dünnen Schleimsegment, welches die ersten 25 µm oberhalb
des Epithels umfasst, an. Der Hauptteil der Bakterien findet sich in einem Bereich 10-20 µm
vom Epithel entfernt. Die übrige Schleimschicht, die in etwa 100 µm umfasst, ist frei von
Bakterien. Die Verteilung von H. pylori im Magenschleim des Gerbils unterscheidet sich in
einigen Punkten von der von H. felis, wie hier gezeigt werden konnte. Beide Spezies
besiedeln zwar ausschließlich die ersten 25 µm des Schleims, jedoch hält sich H. pylori näher
am Magenepithel auf. Der Hauptteil von H. pylori findet sich in den ersten 15 µm der
Schleimschicht, wobei sich ein Drittel der Bakterien in den ersten 5 µm, also in direktem
Kontakt zum Epithel, aufhält.
Es war wenig darüber bekannt, wie Helicobacter seine ökologische Nische im
Magenschleim erkennt. Aufgrund des kontinuierlichen Schleimflusses sind sowohl die hohe
Motilität als auch ihr Orientierungsvermögen entscheidend für die permanente Besiedlung des
Schleims. In dieser Arbeit wurde das chemotaktische Verhalten von H. felis und H. pylori in
vivo im Magenschleim von Maus und Gerbil untersucht, um die chemischen Gradienten zu
identifizieren, die Helicobacter für seine Orientierung benötigt.
Im Magenschleim sind drei permanent vorhandene Gradienten bekannt: der pH-Gradient,
der Bicarbonat/CO2-Gradient und der Harnstoff/Ammonium-Gradient, der durch die
bakterielle Urease bei infizierten Individuen hervorgerufen wird.
Sowohl Harnstoff als auch Bicarbonat sind als Lockstoffe von Helicobacter in vitro
beschrieben. Da es zusätzlich Spekulationen darüber gibt, ob die luminalen pH- und
Bicarbonat-Verhältnisse einen Einfluss auf die Orientierung von Helicobacter haben, wurden
auch diese Bedingungen in die hier vorgestellten Untersuchungen mit aufgenommen.

Anhang
65
In unserer Gruppe wurde vor einiger Zeit eine Methode entwickelt, die es erlaubt, das
Verteilungsmuster und die Kolonisationsdichte von Helicobacter im Magenschleim genau zu
beschreiben. Dabei werden aus dem eröffneten Magen der narkotisierten Tiere Nanoproben
entnommen. Der Magen der Tiere ist exponiert und wird von einer luminalen Superfusion
definierter Zusammensetzung durchströmt. Eine peritoneale Perfusion, die als Dialyse wirkt,
erlaubt die Modifikation von Harnstoff- und Ammoniumkonzentrationen im arteriellen Blut.
Die Dialyse, zusammen mit der inspiratorischen Gasfraktion, ermöglicht es außerdem,
Änderungen der arteriellen Bicarbonat- und CO2-Konzentrationen, sowie des arteriellen pH
durchzuführen.
Um die Gradienten zu finden, die bei der Orientierung von Helicobacter beteiligt sind,
modifizierten wir individuell Gradienten durch Austausch der luminalen Superfusion, der
Dialyse und der inspiratorischen Gasfraktion.
Weder die Neutralisierung des luminalen pH, noch die Umkehrung des
Harnstoff/Ammonium-Gradienten, beeinflussten die Verteilung von Helicobacter.
Zur Absenkung des epithelnahen Schleim-pH wurde der arterielle pH durch eine hohe
inspiratorische CO2-Fraktion, bei gleichbleibender arterieller Bicarbonatkonzentration
erniedrigt. Diese Änderung hatte keinen Effekt auf die bakterielle Orientierung.
Weiterhin wurde die Bicarbonatkonzentration im Magenschleim verändert. Dazu wurde
die Bicarbonatkonzentration und der pCO2 im arteriellen Blut erniedrigt, während der pH
unverändert blieb. Diese Änderung wurde durch eine Hyperventilation und eine Reduzierung
der Bicarbonatkonzentration mittels der Dialyse erzeugt. Es konnte kein Effekt auf die
Orientierung der Bakterien beobachtet werden.
Als dritte Möglichkeit wurden der arterielle pH, und die arterielle Bicarbonatkonzentration
durch die Dialyse bei konstanter Ventilation abgesenkt. Dies resultierte in einer Aufhebung
des Schleim-pH-Gradienten. Unter diesen Bedingungen nahm die Bakteriendichte im
epithelnahen Schleim signifikant ab. Etwa die Hälfte der Bakterien verblieb in den ersten 25
µm des Schleims, der Rest fand sich ungeordnet in der gesamten Schleimschicht und im
Magenlumen wieder.
Um zu bekräftigen, dass die Kombination aus einem erniedrigten arteriellen pH und einer
erniedrigten Bicarbonatkonzentration zur Aufhebung des Schleim-pH-Gradienten führt, haben

Anhang
66
wir eine zusätzliche Serie von Experimenten an der isolierten Antrum-Mucosa von
Meerschweinchen durchgeführt. Diese Methode benutzt ultrafeine, doppelläufige
ionensensitive Mikroelektroden, die den Schleim-pH direkt registrieren. Mit dieser Methode
haben wir gezeigt, dass nur unter der gleichzeitigen Absenkung des arteriellen pH und der
Bicarbonatkonzentration der epithelnahe Schleim-pH auf den luminalen pH abfällt, was den
Schleim-pH-Gradienten aufhebt.
Weiterhin konnten wir mit dieser Methode zeigen, dass eine alleinige Absenkung des pH
oder der Bicarbonatkonzentration den Schleim-pH-Gradienten nicht beeinflusst.
Zusammenfassend konnte gezeigt werden, dass der Schleim-pH-Gradient die Orientierung
von Helicobacter ermöglicht.

Anhang
67
SUMMARY
Helicobacter colonizes the gastric mucus layer close to the epithelial cell surface. Until
now, only the distribution pattern of H. felis in the murine stomach has been described in vivo.
H. felis colonizes the first 25 µm of the gastric mucus adjacent to the epithelial cell surface,
but the majority of the bacteria was found 10-20 µm from the epithelial cell surface. The
remaining part of the gastric mucus layer (approximately 100 µm) is virtually free of bacteria.
In the current study, we found that the distribution of H. pylori is somewhat different in the
gastric mucus of the Mongolian gerbil. Like H. felis, H. pylori is found only within the first
25 µm of the gastric mucus adjacent to the epithelial cells. However, unlike H. felis, the
majority of H. pylori was found within the first 15 µm, a third within the first 5 µm.
Up to now ist is not clear how Helicobacter is able to recognize its ecological niche in the
gastric mucus. Due to the rapid mucus turnover, both high motility and spatial orientation are
crucial for the persistence of the bacteria in this thin mucus layer. In this study, we have
examined the chemotactic behaviour of H. felis and H. pylori in the gastric mucus of mice and
gerbils in vivo to identify the chemical gradient that Helicobacter uses for spatial orientation.
Three chemical gradients are known to exist in the gastric mucus: the pH gradient, the
bicarbonate/CO2 gradient, and in Helicobacter-infected mucosa, a urea/ammonium gradient
created by the bacterial urease. Urea and bicarbonate have been described in vitro as
Helicobacter attractants, and there is also the supposition, that the low luminal pH may serve
as a chemorepellent and therefore enable bacterial orientation.
We previously developed a method to precisely determine the distribution and colonization
density of Helicobacter felis in the gastric mucus of mice in vivo. This method uses a
sampling device to take nanoliter samples from the mucus and mucosa of anesthetized
animals. The stomach of these animals was exposed and the lumen superfused by defined
solutions. A high-flow peritoneal perfusion worked as dialysis, which allowed the
modification of the urea and ammonium concentrations in the arterial plasma. The dialysis
together with changes in the inspired gas fractions made it also possible to change the arterial
concentrations of bicarbonate and CO2 as well as the arterial pH. In order to identify gradients

Anhang
68
involved in the orientation of Helicobacter spp, we inverted or modified individual gradients
by interchanging luminal superfusion, dialysis and inspiratory gas fractions.
Neither the neutralization of the luminal pH nor the inversion of the urea/ammonium
gradient affected the distribution of Helicobacter.
To examine the role of bicarbonate, CO2 and the pH in bacterial orientation we firstly
reduced the pH of the secreted mucus. This was achieved through the ventilation of a high
CO2 gas fraction, maintaining a constant bicarbonate concentration using the dialysis. The
resulting local acidosis did not change the distribution of the bacteria.
Next, we changed the bicarbonate concentration in the mucus. According to the
Henderson-Hasselbalch equation, it is possible to keep the arterial pH constant while reducing
the concentrations of bicarbonate and CO2 to the relevant extent. This change was achieved
by hyperventilation and reduction of the bicarbonate concentration through dialysis and had
no effect on the distribution of Helicobacter.
To eliminate the mucus pH gradient, we reduced both the arterial bicarbonate
concentration and the arterial pHthrough dialysis keeping the ventilation constant. Under
these conditions the density of bacteria in the epithelial mucus decreased significantly. About
a half of the bacteria left their 25 µm mucus zone, and spread over the remaining mucus layer
and lumen.
In order to confirm that the combined reduction of arterial pH and bicarbonate secretion
did lead to the predicted elimination of the mucus pH in vitro, we conducted additional series
of experiments in the explanted antrum mucosa of the guinea pig. This method uses ultrafine-
tipped microelectrodes to directly measure the mucus pH. Using this system we demonstrated
that only under the conditions of reduced arterial bicarbonate and pH, the pH of the
juxtamucosal mucus fell to the value of the lumen pH, thus eliminating the mucus pH
gradient. Under the above-described conditions, with low pH only and low bicarbonate
concentration only, the mucus pH gradient remained unaffected.
To summarise, we have shown that Helicobacter uses the gastric mucus pH gradient for
chemotactic orientation.

Anhang
69
DANKSAGUNGEN
Ich danke Herrn PD Dr. S. Schreiber für die hervorragende und zeitaufwendige praktische
und theoretische Anregung zu meiner Arbeit.
Für die Betreuung meiner Arbeit danke ich Herrn Prof. Dr. P Scheid und Herrn Prof. Dr. R.
Necker.
Allen Mitarbeitern des Lehrstuhls für Physiologie der Medizinischen Fakultät, insbesondere
Herrn Dr. K. Mückenhoff, Frau M. Duparc und Frau G. Reimus, danke ich für ihre tatkräftige
Unterstützung.
Allen Mitarbeitern unseres Teams danke ich für ihre Unterstützung, insbesondere Frau C.
Groll, ohne deren moralischen Beistand ich diese Arbeit nicht beendet hätte.
Der Werkstatt der Medizinischen Fakultät, vor allem Herrn E. Spangenberg danke ich für die
immer schnelle technische Problemlösung.
Herrn C. Ley (Bayer AG) und Herrn Dr. G. Hanauer (Altana AG) danke ich für die
Bereitstellung der infizierten Tiere.
Besonders danke ich meinem Mann K. Konradt, welcher mir immer wieder einen Freiraum
zum Arbeiten schaffte, indem er die Betreuung unserer Tochter Anna Lena übernahm und
überdies dieser Arbeit den letzten Schliff versah .
Ich danke meinen Eltern dafür, dass sie oftmals als Babysitter einsprangen und mich auch
ansonsten mit allen Mitteln unterstützten.

Anhang
70
LEBENSLAUF
Person Name:
Vorname:
Wohnort:
Geboren:
Staatsangehörigkeit :
Familienstand:
Kinder:
Konradt, geb. Stüben
Manuela
Posener Str. 34, 42283 Wuppertal
15.11.71 in Wuppertal
deutsch
verheiratet seit dem 20.12.02
eine Tochter (Anna Lena)
Eltern Vater:
Mutter:
Bernhard Stüben
Kaufmännischer Angestellter
geboren am 21.01.41 in Wuppertal
Rita Stüben, geb. Hansmann
Kunstglaserin
geboren am 24.01.44 in Braunschweig
Geschwister Schwester: Tanja Stüben
Sprach-Heil-Pädagogin
geboren am 09.05.75 in Wuppertal
Werdegang:
15.11.1971 Geboren in Wuppertal
1978 - 1982 Besuch der katholischen Grundschule Oberdörnen in Wuppertal
1982 - 1991 Besuch des katholischen St.-Anna Gymnasiums in Wuppertal
1991 Abschluss allgemeine Hochschulreife
1991-1998 Studium an der Ruhr-Universität Bochum, Studiengang Biologie
08.01.1998 Abschluss des Studiums mit Diplom
01.08.1995 - 30.09.1995 Anstellung als studentische Hilfskraft am Lehrstuhl für
Pflanzenphysiologie, Fakultät für Biologie, Ruhr-Universität
Bochum
02.10.1995 - 28.02.1998 Anstellung als studentische Hilfskraft am Institut für
Physiologie, Medizinische Fakultät, Ruhr-Universität Bochum
Im Rahmen dieser Stelle habe ich meine Diplomarbeit verfasst.

Anhang
71
01.03.1998 - 31.10.1998 Anstellung als wissenschaftliche Hilfskraft am Institut für
Physiologie, Medizinische Fakultät, Ruhr Universität Bochum
01.11.1998 - 31.10.2003 Arbeit als wissenschaftliche Mitarbeiterin am Institut für
Physiologie, Medizinische Fakultät, Ruhr-Universität Bochum
Im Rahmen dieser Stelle habe ich den experimentellen Teil dieser
Dissertation ausgeführt.
Im Rahmen meiner Arbeit an der Ruhr-Universität Bochum habe ich folgende
Lehrveranstaltungen mit betreut:
• Biologie für Mediziner, Praktikumsteil Osmose und kolloidosmotischer Druck
• Physiologiepraktikum für Mediziner, Praktikumsteil Atmungs- und
Leistungsphysiologie
• Tutorium zum Physiologiepraktikum für Mediziner
• POL (Problemorientiertes Lernen) für Mediziner
An den folgenden Kongressen und Tagungen habe ich während meiner Anstellung an der
Ruhr-Universität, zum Teil mit eigenem Beitrag, teilgenommen:
• Tagung der DPG (Deutschen Physiologischen Gesellschaft) in Hamburg 1998
• Tagung der DPG in Bonn 1999 (Posterpräsentation)
• Tagung der DPG in Ulm 2000 (Posterpräsentation)
• Tagung der DPG in Berlin 2001
• Tagung der DPG in Tübingen 2002
• 2. Helicobacter pylori-Workshop der DGHM (Deutsche Gesellschaft für Hygiene und
Mikrobiologie) 1997 in Bochum (Kurzvortrag)
• 3. Helicobacter pylori Workshop der DGHM 1999 in Herrsching
• 4. Helicobacter pylori Workshop der DGHM 2002 in Herrsching (Kurzvortrag)
• CHRO (Campylobacter, Helicobacter and related organisms) Workshop in Baltimore,
Maryland, USA 1999 (Posterpräsentation)
• CHRO Workshop in Freiburg, 2001 (Posterpräsentation)

Anhang
72
ERKLÄRUNG
Hiermit erkläre ich, dass ich die Arbeit selbstständig verfasst und bei keiner anderen
Fakultät eingereicht habe und dass es sich bei der heute von mir eingereichten Dissertation
um fünf in Wort und Bild völlig übereinstimmende Exemplare handelt. Weiterhin erkläre ich,
dass digitale Abbildungen nur die originalen Daten enthalten und in keinem Fall
inhaltsverändernde Bildbearbeitung vorgenommen wurde.