Embed Size (px)
Citation preview

Eur. J. Biochem. 172, 325-332 (1988) 0 FEBS 1988
Inactivation of detergent-solubilized sarcoplasmic reticulum ATPase Gertrude SWOBODA and Wilhelm HASSELBACH Max-Planck-Institute for Medical Research, Department of Physiology, Heidelberg
(Received September l/November 16, 1987) - EJB 87 1004
Inactivation of sarcoplasmic ATPase in the solubilized state was studied in the absence and presence of Ca2 ', Mg2 ' and glycerol. The effects of the detergents octa(ethyleneglyco1) mono-n-dodecyl ether (C12E8), 1-0- tetradecylpropanediol-( 1,3)-3-phosphorylcholine and myristoylglycerophosphocholine were compared. All three detergents caused a rapid decline of the dinitrophenyl phosphatase activity of the unprotected enzyme. The stabilizing effect of Ca2' ions was kinetically analysed. It was found that the stability of the solubilized enzyme depends on the Ca2+ concentration in a manner which is best explained by assuming rapid inactivation of Ca2+- free enzyme accompanied by slow inactivation of a calcium-enzyme complex (EICa). The apparent affinity constants obtained are in the order of 106M-', suggesting that high-affinity Ca2+ binding must be involved. No indications of a contribution were found, either of low-affinity Ca2 +-binding sites of the conformational state E2 or of the high-affinity calcium complex E1Ca2. If CaZt was replaced by Mg2', which exerts a weaker protection, the apparent affinity constants for Mg2+ are in the range of 1 mM-'. The stoichiometry of the effect of Mg2+ depends on the detergent.
It is well established that practically all functions of the Ca2' ATPase of sarcoplasmic reticulum membranes can be retained during solubilization by certain detergents, such as poly(ethyleneglyco1) monoethers (e.g. C12E8 or Triton X-100) and monoacyl phospholipids (lysolecithins) (compare e.g. [l - 51). Although Ca2+ transport, requiring an intact vesicle structure, is, of course, not possible in solubilized prep- arations, all the partial reactions of the commonly used reac- tion cycle [6] can be demonstrated in both directions in the monomeric state [4, 5 , 71. These investigations are compli- cated, however, by the finding that the detergent-solubilized enzyme is unstable in the absence of CaZt ions (see [2 , 4, 5, 81 for C12Es-solubilized preparations). The protective effect of Ca2+ has already been observed earlier by Ikemoto et al. [9] and Hasselbach and Migala [lo] with Triton-solubilized sarcoplasmic membranes.
Our contribution to the problem of inactivation was de- signed to compare the effects of three different detergents on the stability of the enzyme in the absence and presence of
studied as a function of time and Ca2+ concentration. Ca2+ can be expected to interact with different conformational states of the reaction chain [6, 1 I]. Our kinetic analysis estab- lished that the apparent affinity constants of the enzyme for Ca2+ in detergent solution, derived from the dependence of the inactivation on the Ca" concentration, were in the micromolar range. The occupation of low-affinity Ca2+- binding sites could not be ascertained to contribute to enzyme stabilization by high Ca2 ' concentrations.
Ca2+ , M g 2 + and glycerol. The retention of activity was
Correspondence to W. Hasselbach, Abteilung Physiologie, Max- Planck-Institut fur Medizinische Forschung, JahnstraBe 29, D-6900 Heidelberg, Federal Republic of Germany
Abbreviation. C1 zEs, octa(ethyleneglyco1) mono-n-dodecyl ether. Enzyme. Ca2+ ATPase (EC 3.6.1.3).
MATERIALS AND METHODS
Sarcoplasmic vesicles from rabbit skeletal muscle were prepared according to Hasselbach and Makinose [12] as modi- fied by de Meis and Hasselbach [13]. Protein concentration was determined by the biuret method, standardized by Kjeldahl.
Octa(ethyleneglyco1) mono-n-dodecyl ether (C12E8) was obtained from Nikko Chemicals (Tokyo, Japan). L-CI- Myristoylglycerophosphocholine was from Serva (Heidel- berg, FRG) and Ca2 + ionophore A-231 87 from Calbiochem (GieSen/Lahn, FRG).
1 - 0 - Tetradecylpropanediol - (1,3) - 3 - phosphorylcholine was synthesized in our laboratories according to 1141. A 10% aqueous solution was freshly prepared every 3 - 4 days and kept frozen. Phenylmethylsulfonyl fluoride (from Serva, Heidelberg, FRG) was dissolved in isopropanol (0.1 M). 2,4- Dinitrophenyl phosphate was synthesized according to [15] and stored as the lutidinium salt at -20°C.
Inactivation of solubilized sarcoplasmic vesicles
a) In the presence of EGTA. The standard incubation me- dium consisted of 50 mM Mops, pH 7, 0.1 M KCl, usually 10 mM MgC12, 1 mM EGTA and detergent (CI2E8 : 5 mg/ml, tetradecylpropanediol phosphorylcholine or myristoyl- glycerophosphocholine: 0.8 mglml). The inactivation was started by the addition of the vesicle protein (final concentra- tion 0.5 mg/ml, room temperature). At the desired time inter- vals the reaction was stopped by pipetting aliquots (0.5 ml) into cuvettes containing 2.2 mM CaCI, (corresponding to 2 mM excess Ca2+, based on a final volume of 2.5 ml) and the other components of the dinitrophenyl-phosphate-splitting assay (see below). Protein and detergent were thus diluted fivefold for the assay, their ratio remaining constant. For the determination of the basal activities aliquots of the standard
326
incubation mixture (1 mM EGTA) were diluted into the split- ting assay, omitting Ca2+.
In some series of experiments Mg2+ was omitted or varied between 1 mM and 20 mM. Glycerol, if included, was usually 20% (by vol.), and also varied up to 30%. In the latter case the splitting assay contained 6% glycerol, which had no effect on the rate of dinitrophenol liberation, however. Other addi- tives checked were sucrose (1 M), dithioerythritol (5 mM), Na2S205 (10 mM) and phenylmethylsulfonyl fluoride (1 mM, with or without glycerol). In some other control experiments the detergent or the protein concentration was varied.
For comparison native vesicles were incubated with 1 mM EGTA in the absence of detergent (MgZ+ concentration 0 or lOmM, other conditions were the same as in the presence of detergents, described above). Ca2+ ionophore A-23187 (20 pM) was then included in the dinitrophenyl-phosphate- splitting assay only.
h ) In the presence of Ca2+ ions. Preincubation of the vesicles was performed under the same conditions as described with EGTA inactivation, except that EGTA was replaced by different Ca2+ concentrations (0.03 - 10 mM CaC12 in the medium). 10 mM MgC12 were employed in these series; omit- ting Mg2+ did not change the results in the presence of Ca2'. Incubation times were up to 50 h, depending on the stability of the solubilized preparations.
20% glycerol (by vol.) were included in several series con- taining0.03 mM or 0.1 mM CaCI2. Phenylmethylsulfonyl flu- oride (1 mM), which was added to some preparations to avoid inactivation due to the action of proteases, did not increase stability. On the other hand, other effects were observed that led to accelerated inactivation under certain conditions (at low Ca2+). For monitoring the residual activities 0.5-ml aliquots were pipetted into cuvettes containing the amount of Ca2+ missing to yield finally 2 mM (volume 2.5 ml) besides the other components of the dinitrophenyl-phosphate-splitting assay (see below).
Dinitrophenyl phosphatase activity Dinitrophenyl phosphate splitting was measured spec-
trophotometrically at 420 nm in a medium containing 50 mM Mops, pH 7,O.l M KCI, 10 mM MgCI2 and 2 mM CaCI2 (or 0.1 mM CaC12 for comparison in some experiments). The detergent: protein ratio was kept constant, after fivefold di- lution of the incubation mixtures yielding the following final concentrations: vesicular protein 0.1 mg/ml, C12E8 1 mg/ml, tetradecylpropanediol phosphorylcholine or myristoylgly- cerophosphocholine 0.1 6 mgiml). The reaction was started with 1 mM dinitrophenyl phosphate (from a 25 mM aqueous stock solution which was stable for several days at -20°C) and monitored at 22 - 25 "C (thermostat) over at least 2 min (1st min evaluated). For the calculation of activity a E~~~ value of 6800 M-' cm-' was used, determined with 2,4- dinitrophenol under the same conditions.
The basal activities, measured in the presence of 0.2 mM EGTA instead of Ca2+, although quite low, were subtracted from the total values to yield the Ca2+-dependent dini- trophenyl phosphatase activities.
In some experiments 20 mM histidine, pH 7, was used instead of the Mops buffer. The activities of C12E8-solubilized enzyme were then somewhat lower.
RESULTS Dinitrophenyl phosphatase activities in detergent solution
Dinitrophenyl phosphate splitting was chosen for moni- toring the activity of sarcoplasmic vesicles, solubilized under
10 2 4 6 8 1 0 15 20
Incubation time (min 1
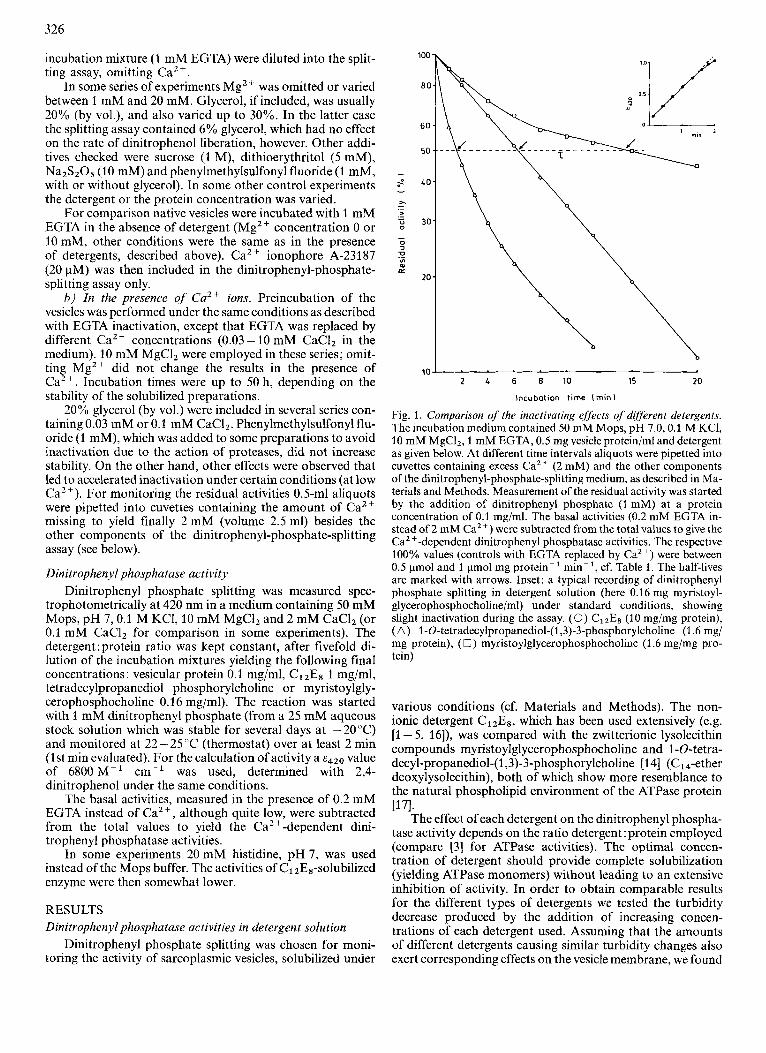
Fig. 1. Comparison of the inactivating effects of dqferent detergents. The incubation medium contained 50 mM Mops, pH 7.0,O.l M KCI, 10 mM MgCI2, 1 mM EGTA, 0.5 mg vesicle protein/ml and detergent as given below. At different time intervals aliquots were pipetted into cuvettes containing excess Caz+ (2 mM) and the other components of the dinitrophenyl-phosphate-splitting medium, as described in Ma- terials and Methods. Measurement of the residual activity was started by the addition of dinitrophenyl phosphate (1 mM) at a protein concentration of 0.1 mg/ml. The basal activities (0.2 mM EGTA in- stead of 2 mM CaZ +) were subtracted from the total values to give the CaZ +-dependent dinitrophenyl phosphatase activities. The respective 100% values (controls with EGTA replaced by Ca2+) were between 0.5 pmol and 1 pmol mg protein-' min-', cf. Table 1 . The half-lives are marked with arrows. Inset: a typical recording of dinitrophenyl phosphate splitting in detergent solution (here 0.16 mg myristoyl- glycerophosphocholine/ml) under standard conditions, showing slight inactivation during the assay. (0) ClzEs (10 mg/mg protein), (A) I-O-tetradecylpropanediol-(1,3)-3-phosphorylcholine (1.6 mg/ mg protein), (0) myristoylglycerophosphocholine (1.6 mg/mg pro- tein)
various conditions (cf. Materials and Methods). The non- ionic detergent Cl2E8, which has been used extensively (e.g. [l - 5, 16]), was compared with the zwitterionic lysolecithin compounds myristoylglycerophosphocholine and 1 -0-tetra- decyl-propanediol-(l,3)-3-phosphorylcholine [I41 (CI4-ether deoxylysolecithin), both of which show more resemblance to the natural phospholipid environment of the ATPase protein
The effect of each detergent on the dinitrophenyl phospha- tase activity depends on the ratio detergent: protein employed (compare (31 for ATPase activities). The optimal concen- tration of detergent should provide complete solubilization (yielding ATPase monomers) without leading to an extensive inhibition of activity. In order to obtain comparable results for the different types of detergents we tested the turbidity decrease produced by the addition of increasing concen- trations of each detergent used. Assuming that the amounts of different detergents causing similar turbidity changes also exert corresponding effects on the vesicle membrane, we found
1171.
327
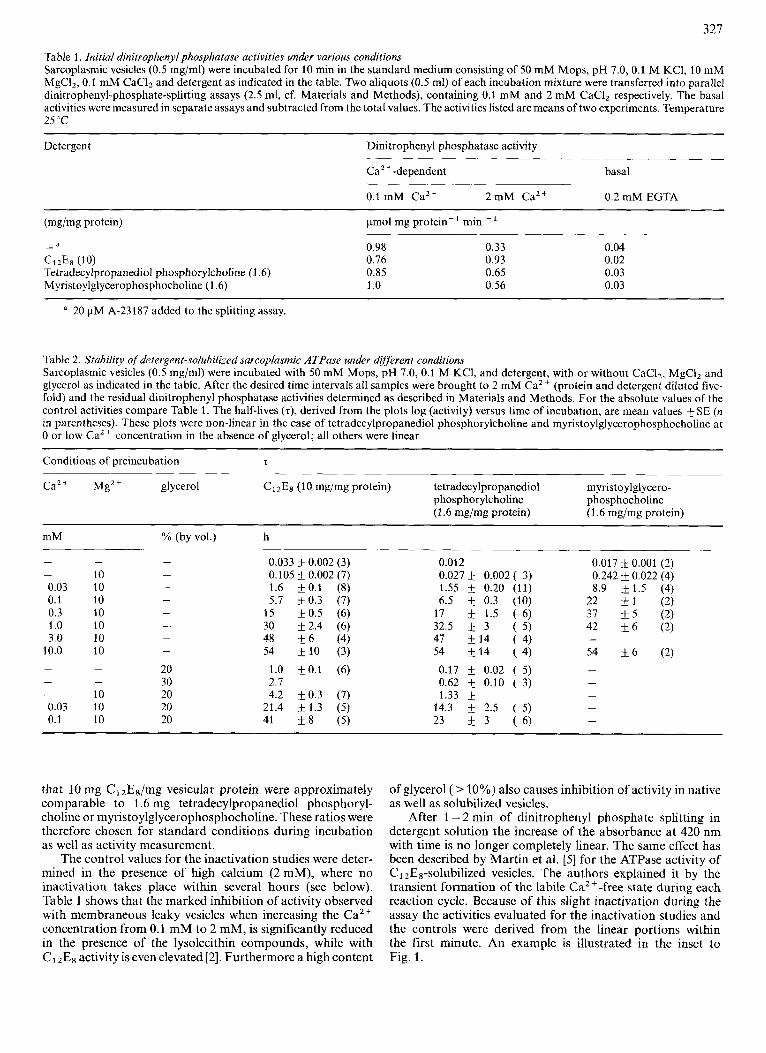
Table 1. Initial dinitroplienyl phosphatase activities under vurious conditions Sarcoplasmic vesicles (0.5 mg/ml) were incubated for 10 min in the standard medium consisting of 50 mM Mops, pH 7.0,O.l M KCI, 10 mM MgCI2, 0.1 mM CaC12 and detergent as indicated in the table. Two aliquots (0.5 ml) of each incubation mixture were transferred into parallel dinitrophenyl-phosphate-splitting assays (2.5 ml, cf. Materials and Methods), containing 0.1 mM and 2 mM CaCI, respectively. The basal activities were measured in separate assays and subtracted from the total values. The activities listed are means of two experiments. Temperature 25 "C
Detergent Dinitrophenyl phosphatase activity
CaZ +-dependent basal
0.1 mM Ca2+ 2 m M Ca2+ 0.2 mM EGTA
(mg/mg protein) pmol mg protein - min - ' a -
CllE8 (lo) Tetradecylpropanediol phosphorylcholine (1 -6) Myristoylglycerophosphocholine (1.6)
0.98 0.76 0.85 1 .o
0.33 0.93 0.65 0.56
0.04 0.02 0.03 0.03
a 20 pM A-23187 added to the splitting assay.
Table 2. Stability of detergelit-soluhilized sarcoplusmic ATPase under different conditions Sarcoplasmic vesicles (0.5 mg/ml) were incubated with 50 mM Mops, pH 7.0,O.l M KCI, and detergent, with or without CaCI2, MgCI2 and glycerol as indicated in the table. After the desired time intervals all samples were brought to 2 mM CaZ+ (protein and detergent diluted five- fold) and the residual dinitrophenyl phosphatase activities determined as described in Materials and Methods. For the absolute values of the control activities compare Table 1. The half-lives (z), derived from the plots log (activity) versus time of incubation, are mean values 5 SE (n in parentheses). These plots were non-linear in the case of tetradecylpropanediol phosphorylcholine and myristoylglycerophosphocholine at 0 or low Ca2+ concentration in the absence of glycerol; all others were linear
Conditions of preincubation z
Ca2+ Mg2+ glycerol ClzEs (10 mg/mg protein) tetradecylpropanediol myristoylglycero- phosphor ylcholine phosphocholine (1.6 mg/mg protein) (1.6 mg/mg protein)
mM % (by vol.) h
-
0.03 0.1 0.3 1 .o 3.0
10.0
- 0.03 0.1
-
10 10 10 10 10 10 10
-
10 10 10
20 30 20 20 20
0.033 k 0.002 (3) 0.105 f 0.002 (7) 1.6 kO.1 (8) 5.7 k O . 3 (7)
15 k0.5 (6) 30 2 2.4 (6) 48 + 6 (4)
1.0 kO.1 (6) 2.7 4.2 f 0 . 3 (7)
21.4 1.3 (5) 41 ,8 ( 5 )
54 ; 10 (3)
0.012 0.027 f 0.002 ( 3) 1.55 t- 0.20 (11) 6.5 f 0.3 (10)
17 t- 1.5 ( 6) 32.5 t 3 ( 5 ) 47 t-34 ( 4) 54 k 1 4 ( 4) 0.17 f 0.02 ( 5) 0.62 5 0.10 ( 3) 1.33 f
14.3 i: 2.5 ( 5 ) 23 f 3 ( 6)
-
0.017 & 0.001 (2) 0.242 -t 0.022 (4) 8.9 & 1.5 (4)
22 i 1 (2) 37 * 5 (2) 42 ,6 (2)
54 f 6 (2) -
that 10 mg ClzE8/mg vesicular protein were approximately comparable to 1.6 mg tetradecylpropanediol phosphoryl- choline or myristoylglycerophosphocholine. These ratios were therefore chosen for standard conditions during incubation as well as activity measurement.
The control values for the inactivation studies were deter- mined in the presence of high calcium (2 mM), where no inactivation takes place within several hours (see below). Table 1 shows that the marked inhibition of activity observed with membraneous leaky vesicles when increasing the Ca2+ concentration from 0.1 mM to 2 mM, is significantly reduced in the presence of the lysolecithin compounds, while with ClzEs activity is even elevated [2]. Furthermore a high content
of glycerol (> 10%) also causes inhibition of activity in native as well as solubilized vesicles.
After 1 - 2 min of dinitrophenyl phosphate splitting in detergent solution the increase of the absorbance at 420 nm with time is no longer completely linear. The same effect has been described by Martin et al. [5] for the ATPase activity of C1 zEs-solubilized vesicles. The authors explained it by the transient formation of the labile Ca2+-free state during each reaction cycle. Because of this slight inactivation during the assay the activities evaluated for the inactivation studies and the controls were derived from the linear portions within the first minute. An example is illustrated in the inset to Fig. 1.
328
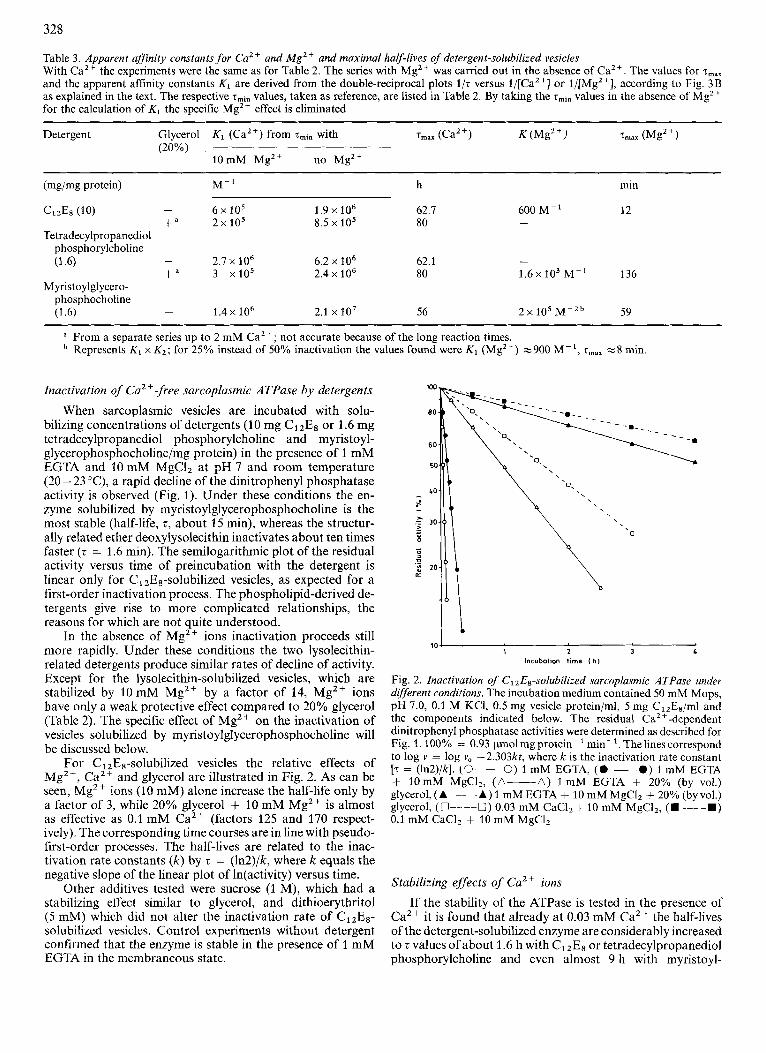
Table 3. Apparent affinity constants for Ca2+ and MgZt and maximal half-lives of detergent-solubilized vesicles With CaZt the experiments were the same as for Table 2. The series with Mgz+ was carried out in the absence of CaZt. The values for zmax and the apparent affinity constants Ki are derived from the double-reciprocal plots l /z versus l/[Ca'+] or l/[MgZ+], according to Fig. 3B as explained in the text. The respective rmin values, taken as reference, are listed in Table 2. By taking the zmin values in the absence of Mg2+ for the calculation of K1 the specific Mgz+ effect is eliminated
Detergent Glycerol K 1 (Ca") from zmin with z,,, (CaZ+) K (Mg2 +) 7max (Mgz+) (20%)
10mM Mgzi no Mg2+
(mg/mg protein) M- ' h min
- 6 x lo5 1.9 x lo6 62.1 600 M-' 12 +" 2x105 8.5 x 105 80
- 2.1 x 106 6.2 x lo6 62.1 -
- CizEs (10)
Tetradecylpropanediol phosphorylcholine (1.6) +" 3 xi05 2.4 x lo6 80 1 . 6 ~ lo3 M-' 136
Myristoylglycero- phosphocholine (1.6) - 1.4 x lo6 2.1 x 107 56 2 x 105 M - Z ~ 59
a From a separate series up to 2 mM Ca2+; not accurate because of the long reaction times. Represents K , x KZ; for 25% instead of 50% inactivation the values found were K , (Mg") w900 M-', zmaX w8 min.
Inactivation of Ca2 ' -jiree sarcoplasmic ATPase by detergents When sarcoplasmic vesicles are incubated with solu-
bilizing concentrations of detergents (10 mg Cl& or 1.6 mg tetradecylpropanediol phosphorylcholine and myristoyl- glycerophosphocholine/mg protein) in the presence of 1 mM EGTA and 10 mM MgC12 at pH 7 and room temperature (20 - 23 "C), a rapid decline of the dinitrophenyl phosphatase activity is observed (Fig. 1). Under these conditions the en- zyme solubilized by myristoylglycerophosphocholine is the most stable (half-life, z, about 15 min), whereas the structur- ally related ether deoxylysolecithin inactivates about ten times faster (z = 1.6 min). The semilogarithmic plot of the residual activity versus time of preincubation with the detergent is linear only for C12E8-solubilized vesicles, as expected for a first-order inactivation process. The phospholipid-derived de- tergents give rise to more complicated relationships, the reasons for which are not quite understood.
In the absence of Mg2+ ions inactivation proceeds still more rapidly. Under these conditions the two lysolecithin- related detergents produce similar rates of decline of activity. Except for the lysolecithin-solubilized vesicles, which are stabilized by 10 mM Mg2' by a factor of 14, Mg2+ ions have only a weak protective effect compared to 20% glycerol (Table 2). The specific effect of Mg2+ on the inactivation of vesicles solubilized by myristoylglycerophosphocholine will be discussed below.
For C1 2E8-sohbilized vesicles the relative effects of Mg", Ca2+ and glycerol are illustrated in Fig. 2. As can be seen, Mg2+ ions (10 mM) alone increase the half-life only by a factor of 3, while 20% glycerol + 10 mM Mg2+ is almost as effective as 0.1 mM Ca2+ (factors 125 and 170 respect- ively). The corresponding time courses are in line with pseudo- first-order processes. The half-lives are related to the inac- tivation rate constants ( k ) by t = (ln2)/k, where k equals the negative slope of the linear plot of ln(activity) versus time.
Other additives tested were sucrose (1 M), which had a stabilizing effect similar to glycerol, and dithioerythritol ( 5 mM) which did not alter the inactivation rate of C12E8-
solubilized vesicles. Control experiments without detergent confirmed that the enzyme is stable in the presence of 1 mM EGTA in the membraneous state.
10 1 2 3 1.
Incubation time I h j
Fig. 2. Inactivation of C12E8-solubilized sarcoplasmic ATPase under dqferent conditions. The incubation medium contained 50 mM Mops, pH 7.0, 0.1 M KCl, 0.5 mg vesicle protein/ml, 5 mg C12E8/ml and the components indicated below. The residual Ca2+-dependent dinitrophenyl phosphatase activities were determined as described for Fig. 1.100% = 0.93 pmolmg protein- min- I . The lines correspond to log v = log v, -2.303kt, where k is the inactivation rate constant [z = (ln2)/k]. (0-0) 1 mM EGTA, (0-0) 1 mM EGTA + 10 mM MgC12, (A-A) 1 mM EGTA + 20% (by vol.) glycerol, (A--A) 1 mM EGTA + 10 mM MgClz + 20% (by vol.) glycerol, (O----O) 0.03 mM CaClz + 10 mM MgC12, (m----H) 0.1 mM CaC12 + 10 mM MgC12
Stabilizing eJjCects of Ca2+ ions If the stability of the ATPase is tested in the presence of
Ca2 ' it is found that already at 0.03 mM Ca2+ the half-lives of the detergent-solubilized enzyme are considerably increased to z values of about 1.6 h with C12Es or tetradecylpropanediol phosphorylcholine and even almost 9 h with myristoyl-
329
A
1 5 10
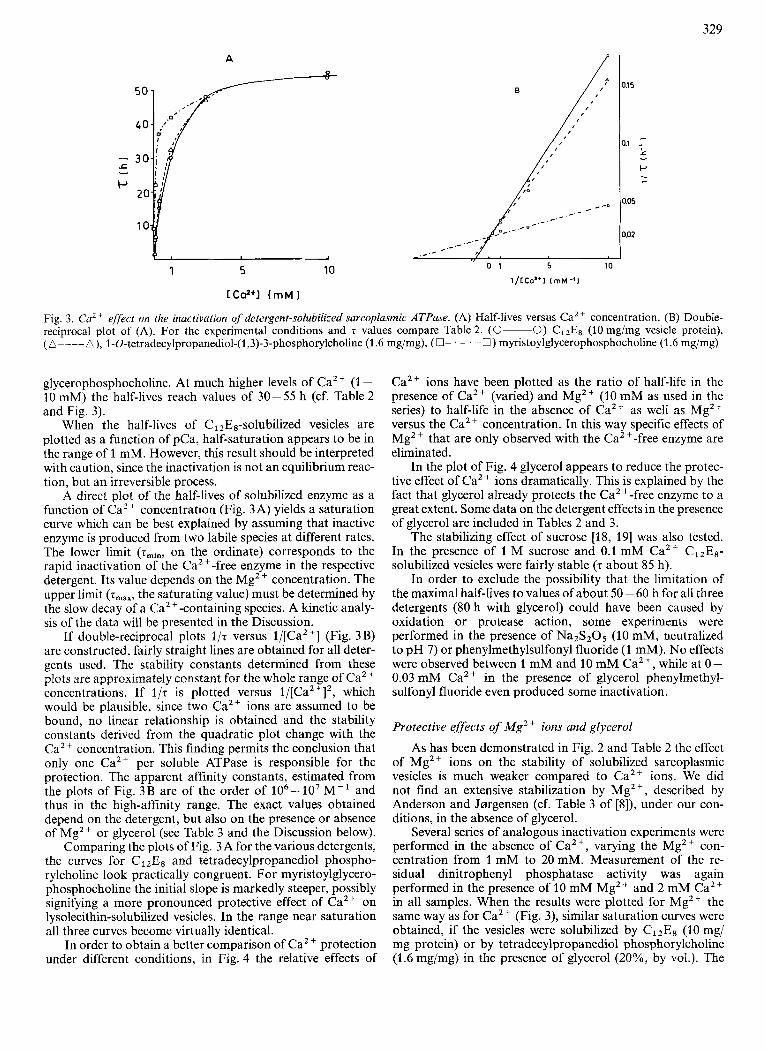
Fig. 3. Ca2 + effect on the irtactivation uf detergent-solubilized sarcoplasmic ATPase. (A) Half-lives versus Ca2+ concentration. (8) Double- reciprocal plot of (A). For the experimental conditions and 7 values compare Table 2. (0-0) ClzEs (10 mg/mg vesicle protein), (A----A), l-O-tetradecylpropanediol-(1,3)-3-phosphorylcholine (1.6 mgimg), (0- . - . -0) myristoylglycerophosphocholine (1.6 mg/mg)
glycerophosphocholine. At much higher levels of Ca2+ (1 - 10 mM) the half-lives reach values of 30 - 55 h (cf. Table 2 and Fig. 3).
When the half-lives of Cl2E8-solubilized vesicles are plotted as a function of pCa, half-saturation appears to be in the range of 1 mM. However, this result should be interpreted with caution, since the inactivation is not an equilibrium reac- tion, but an irreversible process.
A direct plot of the half-lives of solubilized enzyme as a function of CaZ + concentration (Fig. 3A) yields a saturation curve which can be best explained by assuming that inactive enzyme is produced from two labile species at different rates. The lower limit (zminr on the ordinate) corresponds to the rapid inactivation of the Ca2+-free enzyme in the respective detergent. Its value depends on the Mg2+ concentration. The upper limit (zmax, the saturating value) must be determined by the slow decay of a Ca'+-containing species. A kinetic analy- sis of the data will be presented in the Discussion.
If double-reciprocal plots 1/z versus l/[Ca2+] (Fig. 3B) are constructed, fairly straight lines are obtained for all deter- gents used. The stability constants determined from these plots are approximately constant for the whole range of Ca2+ concentrations. If l / z is plotted versus 1/[CaZ+]2, which would be plausible, since two Ca2+ ions are assumed to be bound, no linear relationship is obtained and the stability constants derived from the quadratic plot change with the Ca2+ concentration. This finding permits the conclusion that only one CaZt per soluble ATPase is responsible for the protection. The apparent affinity constants, estimated from the plots of Fig. 3B are of the order of lo6- lo7 M - l and thus in the high-affinity range. The exact values obtained depend on the detergent, but also on the presence or absence of Mg2+ or glycerol (see Table 3 and the Discussion below).
Comparing the plots of Fig. 3A for the various detergents, the curves for C12E8 and tetradecylpropanediol phospho- rylcholine look practically congruent. For myristoylglycero- phosphocholine the initial slope is markedly steeper, possibly signifying a more pronounced protective effect of Ca2+ on lysolecithin-solubilized vesicles. In the range near saturation all three curves become virtually identical.
In order to obtain a better comparison of CaZ+ protection under different conditions, in Fig. 4 the relative effects of
Ca2+ ions have been plotted as the ratio of half-life in the presence of Ca2+ (varied) and Mg2+ (10 mM as used in the series) to half-life in the absence of Ca2+ as well as Mg2+ versus the Ca2+ concentration. In this way specific effects of Mgz+ that are only observed with the Ca2+-free enzyme are eliminated.
In the plot of Fig. 4 glycerol appears to reduce the protec- tive effect of Ca2+ ions dramatically. This is explained by the fact that glycerol already protects the Ca2+-free enzyme to a great extent. Some data on the detergent effects in the presence of glycerol are included in Tables 2 and 3.
The stabilizing effect of sucrose [18, 191 was also tested. In the presence of 1 M sucrose and 0.1 mM Ca2+ C12Es- solubilized vesicles were fairly stable (z about 85 h).
In order to exclude the possibility that the limitation of the maximal half-lives to values of about 50 - 60 h for all three detergents (80 h with glycerol) could have been caused by oxidation or protease action, some experiments were performed in the presence of Na2S205 (10 mM, neutralized to pH 7) or phenylmethylsulfonyl fluoride (1 mM). No effects were observed between 1 mM and 10 mM Ca2+, while at 0- 0.03 mM Ca2+ in the presence of glycerol phenylmethyl- sulfonyl fluoride even produced some inactivation.
Protective effects of Mg2+ ions and glycerol
As has been demonstrated in Fig. 2 and Table 2 the effect of Mg2+ ions on the stability of solubilized sarcoplasmic vesicles is much weaker compared to Ca2+ ions. We did not find an extensive stabilization by Mg2+, described by Anderson and Jorgensen (cf. Table 3 of [S]), under our con- ditions, in the absence of glycerol.
Several series of analogous inactivation experiments were performed in the absence of CaZf , varying the Mg2+ con- centration from 1 mM to 20 mM. Measurement of the re- sidual dinitrophenyl phosphatase activity was again performed in the presence of 10 mM Mgz+ and 2 mM CaZ+ in all samples. When the results were plotted for Mg2+ the same way as for CaZ + (Fig. 3), similar saturation curves were obtained, if the vesicles were solubilized by CIZEB (10 mg/ mg protein) or by tetradecylpropanediol phosphorylcholine (1.6 mg/mg) in the presence of glycerol (20%, by vol.). The
330
0.1 1 2 3 CCo**l (mM)
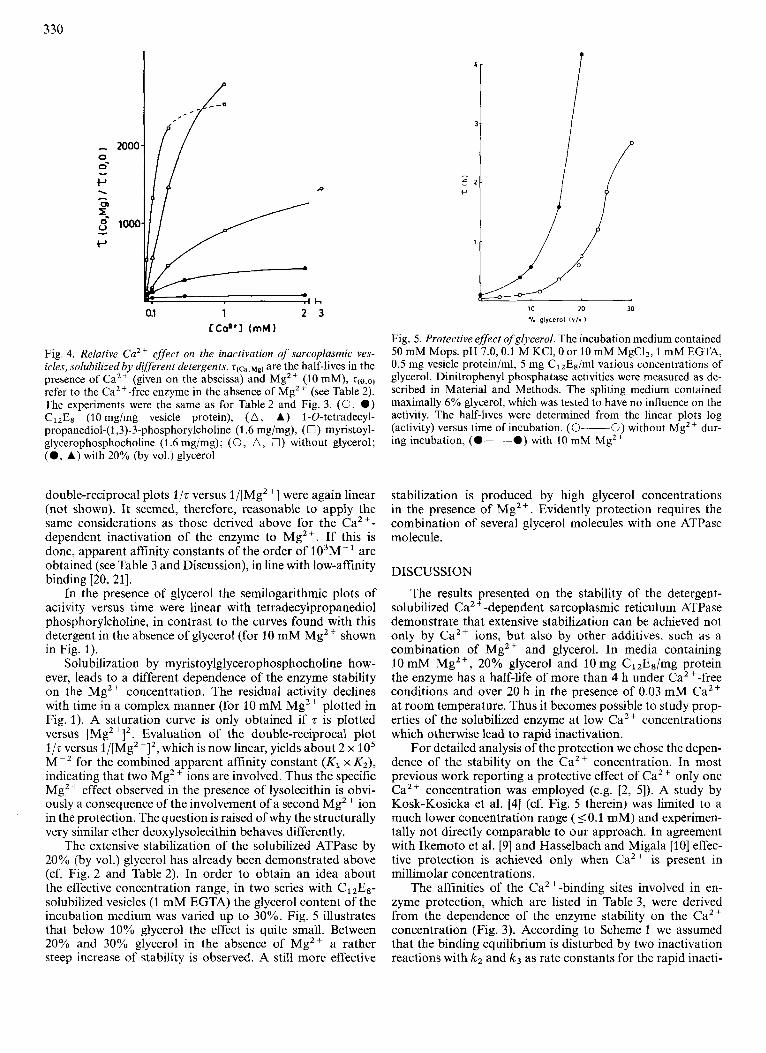
Fig. 4. Relative Caz+ effect on the inactivation of sarcoplasmic ves- icles, solubilized by different detergents. z ( ~ ~ , ~ ~ , are the half-lives in the presence of Ca2+ (given on the abscissa) and Mgzc (10 mM), q O , o )
refer to the Caz+-free enzyme in the absence of Mgz+ (see Table 2). The experiments were the same as for Table 2 and Fig. 3. (0, 0 ) C12Es (10 mg/mg vesicle protein), ( A , A) 1-0-tetradecyl- propanediol-(l,3)-3-phosphorylcholine (1.6 mgimg), (n) myristoyl- glycerophosphocholine (1.6 mgimg); (0, A, 0) without glycerol; (0 , A) with 20% (by vol.) glycerol
double-reciprocal plots l/t versus 1/[Mg2 '1 were again linear (not shown). It seemed, therefore, reasonable to apply the same considerations as those derived above for the Ca2 +-
dependent inactivation of the enzyme to Mg2+. If this is done, apparent affinity constants of the order of 103M-' are obtained (see Table 3 and Discussion), in line with low-affinity binding [20, 211.
In the presence of glycerol the semilogarithmic plots of activity versus time were linear with tetradecylpropanediol phosphorylcholine, in contrast to the curves found with this detergent in the absence of glycerol (for 10 mM Mg2+ shown in Fig. 1).
Solubilization by myristoylglycerophosphocholine how- ever, leads to a different dependence of the enzyme stability on the Mg2+ concentration. The residual activity declines with time in a complex manner (for 10 mM Mg2+ plotted in Fig. 1). A saturation curve is only obtained if z is plotted versus [Mg2 + I 2 . Evaluation of the double-reciprocal plot l / z versus 1/[Mg2+I2, which is now linear, yields about 2 x lo5 M-2 for the combined apparent affinity constant ( K , x Kz) , indicating that two Mg2+ ions are involved. Thus the specific Mg2+ effect observed in the presence of lysolecithin is obvi- ously a consequence of the involvement of a second Mg2+ ion in the protection. The question is raised ofwhy the structurally very similar ether deoxylysolecithin behaves differently.
The extensive stabilization of the solubilized ATPase by 20% (by vol.) glycerol has already been demonstrated above (cf. Fig. 2 and Table 2). In order to obtain an idea about the effective concentration range, in two series with C12E8- solubilized vesicles (1 mM EGTA) the glycerol content of the incubation medium was varied up to 30%. Fig. 5 illustrates that below 10% glycerol the effect is quite small. Between 20% and 30% glycerol in the absence of Mg2' a rather steep increase of stability is observed. A still more effective
10 20 30 % glycerol f v l v 1
Fig. 5. Protective effect ofglycerol. The incubation medium contained 50 mM Mops, pH 7.0,O.l M KCI, 0 or 10 mM MgCI2, 1 mM EGTA, 0.5 mg vesicle protein/ml, 5 mg CI2E8/ml various concentrations of glycerol. Dinitrophenyl phosphatase activities were measured as de- scribed in Material and Methods. The spliting medium contained maximally 6% glycerol, which was tested to have no influence on the activity. The half-lives were determined from the linear plots log (activity) versus time of incubation. (0-0) without Mg2+ dur- ing incubation, (0-0) with 10 mM Mgz+
stabilization is produced by high glycerol concentrations in the presence of Mg2+. Evidently protection requires the combination of several glycerol molecules with one ATPase molecule.
DISCUSSION
The results presented on the stability of the detergent- solubilized Ca2+-dependent sarcoplasmic reticulum ATPase demonstrate that extensive stabilization can be achieved not only by Ca2+ ions, but also by other additives, such as a combination of Mg2 + and glycerol. In media containing 10mM Mg2+, 20% glycerol and 10mg C12E8/mg protein the enzyme has a half-life of more than 4 h under Ca2+-free conditions and over 20 h in the presence of 0.03 mM Ca2+ at room temperature. Thus it becomes possible to study prop- erties of the solubilized enzyme at low Ca2+ concentrations which otherwise lead to rapid inactivation.
For detailed analysis of the protection we chose the depen- dence of the stability on the Ca2+ concentration. In most previous work reporting a protective effect of Ca2+ only one Ca2+ concentration was employed (e.g. [2 , 51). A study by Kosk-Kosicka et al. [4] (cf. Fig. 5 therein) was limited to a much lower concentration range (4 0.1 mM) and experimen- tally not directly comparable to our approach. In agreement with Ikemoto et al. [9] and Hasselbach and Migala [lo] effec- tive protection is achieved only when Ca2+ is present in millimolar concentrations.
The affinities of the Ca2+-binding sites involved in en- zyme protection, which are listed in Table 3, were derived from the dependence of the enzyme stability on the Ca2+ concentration (Fig. 3). According to Scheme 1 we assumed that the binding equilibrium is disturbed by two inactivation reactions with k2 and k3 as rate constants for the rapid inacti-

331
vation of the Ca2+-free enzyme and the slow inactivation of the Ca enzyme complex respectively.
[ECaI E + C a 2 s E C a ECa2 K1 = \ / [El [Ca2 +I
k\ Jk, Ei Scheme 1
From the scheme the following expression results for the accumulation of inactive enzyme.
-- IEiJ - k2 [El + k3 [ECa] = k ([Etntal] - [Ei] - [ECa2]), dt
where k is the inactivation rate constant derived from the plot log (residual activity) versus time at each Ca2 + concentration. With [Etotal] = [El + [ECa] + [ECa2J + [E;] and [ECaJ = K1 [El [Ca2 '1 the following relation was obtained:
k , + k3 K1 [Ca"] - ln2 k = -~ 1 + K 1 [Ca2+] T
As k3 in the equation is not directly experimentally accessible, graphical methods are employed for the estimation of the apparent affinity constants K1.
Fig. 3A corresponds to the plot according to Eqn ( l ) , rearranged for T . The double-reciprocal plot l / ~ versus l / [Ca2+] (Fig. 3B) is better suited to the graphical evaluation leading to an estimate of K1.
Eqn (2) contains the approximation K1 [Ca"] $1, which applies well to all concentrations studied. Even for the lowest Ca2+ concentration used (0.03 mM total Ca2+; about 0.026mM free Ca2+ at T ) the deviation due to this ap- proximation is less than the accuracy of experimentally deter- mining T .
The double-reciprocal plots, shown in Fig. 3B, are fairly linear for all detergents used. Linearity indicates that, apart from the free enzyme, only the ECa complex contributes to enzyme decay. If ECa, were also involved, quadratic components would be expected. ECa2, which is the prevailing species at high Ca2+ concentration, was considered to be stable. It is difficult to imagine that ECa2 could be more labile than ECa. K1 can be determined from Fig. 3B either from the slope (I/T,,,~,, . K , ) of from the negative intercept on the abscissa ( - T,~" . Kl/~max) . The T,,,,~ value equals the reciprocal ordinate intercept. The two different K 1 values listed for each detergent in Table 3 arise from the problem of assigning values for T,,,~" as reference, depending on the Mg2+ concentration employed in the absence of Ca2+. When Ca2+ is present, Mg2+ ions have no effect on the inactivation, which is con- sistent with a proposed competition between Ca2+ and Mg2+ for the two high-affinity binding sites of the El confor- mation [20, 221. In spite of this problem all apparent affinity constants obtained for Ca2+ were in the high-affinity range. The Ca2 + affinities in detergent solution, obtained by other methods at lower Ca2+ concentrations, have been found in the same range, provided that spontaneous inactivation could be kept low. They were shown to be only slightly or not at all different from the values determined in the vesicular state
Scheme 1 does not explicitly differentiate between the en- zyme conformations of high (El) and low (E,) affinity [6, 111, included in [El, as the equilibrium constant [E2]/[El] under
[2-4, 231.
the solubilized conditions is not known. Even in the vesicular state estimation is quite vague (according to [20] between 11 and lo4, probably around lo3). If, in the calculation, E is substituted by El + E2, k , in Eqn (1) will then become a mixed rate constant, relating the individual inactivation rate constants kil for El and ki2 for E2 in the following way:
,where KO = [E2]/[El]. ki1 + kiz Ko k2 = 1 + K, (3)
Since ki2 % kil (kil is probably very small or 0, if [El] were stable), for high K,, k2 would become approximately equal to ki2, representing the decay of the labile species E2.
Furthermore, if during the slow inactivation of ECa an intermediate species EiCa were involved, Eqn (1) would con- tain an additional term. However, assuming a low affinity of Ei for Ca2+, this could be neglected.
Thus the simplified scheme and the calculations derived from it can be assumed to give reasonable estimates of appar- ent affinity constants. A plausible pathway of the inactivation process in the presence of Ca2+ could include the following steps: Ca2+ could bind with high affinity to the proportion of El present in the equilibrium mixture El + E2, thus pro- moting a shift from E2 to El, which would counteract the equilibrium shift caused by the removal of E2 due to the irreversible formation of inactive enzyme. Our experiments did not yield an indication of the involvement of low-affinity binding sites (E2Ca) in the protective effect of Ca2+.
The weaker stabilizing effect of Mg2+ has been analysed in an analogous manner to the Ca2+ protection by substitut- ing Mg2+ for Ca2+ in Scheme 1 and Eqns (1) and (2), as the double-reciprocal plots l / ~ versus 1/[Mg2+] were again approximately linear, if C12E8 or tetradecylpropanediol phosphorylcholine and glycerol were employed. The relatively low T,,, values (Table 3) should correspond to the inactivation of EMg (probably E2Mg), which is obviously far more labile than ECa in the solubilized state. Hence the approximation K 1 [Mg2+] 9 1 i s not very well fulfilled even for the higher Mg2+ concentrations as K1 for Mg2+ is much lower than K1 for Ca2+ (cf. Table 3). Still the constants for Mg2+ estimated by this method are in the expected low-affinity range. For the binding of Mg2+ to E2 in the vesicular state an equilibrium constant of 128 M- ' was reported [20]. Ac- cording to the authors the El state of the enzyme has even three low-affinity binding sites for Mg2+, two of which are identical with the high-affinity Ca2 +-binding sites.
In the presence of myristoylglycerophosphocholine a lin- ear double-reciprocal relation was only obtained if, instead of the half-lives, the times corresponding to a low degree of inactivation (e.g. 25%) were plotted versus 1/[Mg2+]. Yet at 50% or 60% inactivation a quadratic relationship was found, i.e. I / T versus 1/[Mg2+I2 was linear. This can be explained by assuming that a second Mg2+ ion gets involved in the protection, as inactivation proceeds. Possibly Mg2+ binding to the El conformation could play a role, as according to [22] at high Mg2' concentrations El can be stabilized at the expense of E2.
According to Eqn (1) the stability of the solubilized en- zyme in the presence of Ca2+ or Mg2+ should be indepen- dent of the protein concentration. This was confirmed by a separate series of experiments in the range 0.25 - 2 mg protein/ml (not shown). In contrast, Gafni and Boyer [19] described a protein-dependent inactivation of Triton- solubilized sarcoplasmic vesicles and derived calculations on a monomer - dimer equilibrium from the results plotted in

332
Fig. 3 [19]. Yet in these experiments the protein was diluted by a Triton-containing medium, thereby varying the detergent/ protein ratio. In agreement with our findings Esmann [24] reported, for (Na’ + K’) ATPase at a CI2Es/protein ratio of 4, inactivation curves independent of the protein concen- tration between 0.25 mg/ml and 2 mg/ml. Apparently under the conditions generally employed for complete solubilization the detergent/protein ratio determines the behaviour of the enzyme rather than the detergent concentration per se, pro- vided that the critical micelle concentration is not approached during dilution. In our experiments the detergent concen- tration range used during incubation and activity measure- ment is far above the respective critical micelle concentration values [25], so that a change of the properties and effects of the detergents is not to be expected.
A problem which is not explained is the non-linear activity decline of vesicles solubilized by tetradecylpropanediol phosphorylcholine or myristoylglycerophosphocholine at 0 or low Ca2+ concentrations (cf. Fig. 1 and Table 2). If one species decays in a pseudo-first-order process according to v = v, e-k* the semilogarithmic plot of activity versus time should be linear. This was observed with C&s under all conditions employed, with the lysolecithin-related detergents, however, only in the presence of 20% glycerol or at high Ca2+ concentrations. For (Na’ + K+)ATPase Esmann [24] dem- onstrated a bi-exponential time course of inactivation by Cl2Es and explained it by assuming two species (a ‘protomer’ and a ‘diprotomer’) being inactivated at different rates with- out equilibrium exchange. If a mixture of monomeric and dimeric sarcoplasmic ATPase [26,27] were responsible for the curved plots of Fig. 1, equilibrium between the two species would have to be very slow. The use of detergentlprotein ratios much higher than 1.6, as employed in this study, would have led to a more rapid inactivation and extensive inhibition.
Finally we showed that in the absence of ligands (Ca2+ or MgZ+) and glycerol or sucrose none of the detergents used can prevent very rapid inactivation at room temperature. Stabilization by CaZ+ as well as Mg2+ is most effective in the presence of myristoylglycerophosphocholine. The order of the apparent affinity constants for Ca2+ (Table 3) in the different detergents, with zero Mg2+ taken as reference, cor- responds approximately to the relative Ca2+ effects, plotted in Fig. 4. This would mean that myristoylglycerophosphocho- line produces the most effective protection at a given Ca2 + concentration. This is also the detergent which is structurally most closely related to the native phospholipid environment of the ATPase. Thus, although an ether deoxylysolecithin is more resistant to degradation by cellular enzymes (25, 281 it is not the most suitable solubilizing agent for the sarcoplasmic ATPase.
REFERENCES
1. Dean, W. L. & Tanford, C. (1978) Biochemistry 17, 1683-
2. Msller, J. V., Lind, K. E. & Andersen, J. P. (1980) J . Biol. Chem.
3. Liidi, H., Rauch, B. & Hasselbach, W. (1982) Z . Naturforsch.
4. Kosk-Kosicka, D., Kurzmack, M. & Inesi, G. (1983) Biochemistry
5. Martin, D. W., Tanford, C. & Reynolds, J. A. (1984) Proc. Nut1
6. de Meis, L. & Vianna, A. L. (1979) Annu. Rev. Biochem. 48,275-
7. Tanford, C. (1984) Crit. Rev. Biochem. 17, 123-151. 8. Andersen, J. P. & Jsrgensen, P. L. (1985) J . Membr. Biol. 88,
9. Ikemoto, N., Bhatnagar, G. M. & Gergely, J. (1971) Biochem.
1690.
255,1912-1920.
3 7 ~ , 299 - 307.
22,2559-2567.
Acad. Sci. U S A 81,6623 - 6626.
292.
187- 198.
Biophys. Res. Commun. 44, 1510-1517. 10. Hasselbach, W. & Migala, A. (1972) FEBSLett. 26, 20-24. 11. de Meis, L. (1981) Thesarcoplasmic reticulum, Wiley, New York. 12. Hasselbach, W. & Makinose, M. (1963) Biochem. Z. 339, 94-
13. de Meis, L. & Hasselbach, W. (1971) J . Biol. Chem. 246, 4759-
14. Weltzien, H. U., Arnold, B. & Reuther, R. (1977) Biochim. Bio-
15. Ramirez, F. & Maracek, J. F. (1978) Synthesis 5, 601 -603. 16. Swoboda, G. & Hasselbach, W. (1985) Z. Nuturfursch. 40c, 863 -
17. Tanford, C. & Reynolds, J. A. (1976) Biochim. Biophys. Acta 457,
18. Murphy, A. J., Pepitone, M. & Highsmith, S. (1982) J . Bid.
19. Gafni, A. & Boyer, P. D. (1984) Biochemistry 23,4362-4367. 20. Tanford, C. & Martin, D. W. (1982) Z . Naturforsch. 37c, 522-
21. Punzengruber, C., Prager, R., Kolossa, N., Winkler, F. & Suko,
22. Loomis, C. R., Martin, D. W., McCaslin, D. R. & Tanford, C.
23. Andersen, J. P., Msller, J. V. & Jsrgensen, P. L. (1982) J . Biol.
24. Esmann, M. (1986) Biochim. Biophys. Acta 857, 38-47. 25. Helenius, A,, McCaslin, D. R., Fries, E. & Tanford, C. (1979)
26. Silva, J. I,. & Verjovski-Almeida, S. (1985) J. Biol. Chem. 260,
27. Nestruck-Goyke, A. C. & Hasselbach, W. (1981) Eur. J . Biochem.
28. Weltzien, H. U. (1975) Exp. Cell Res. 92, 11 1 - 121.
111.
4763.
phys. Acia 466,411 -421.
875.
133-170.
Chem. 257, 3551 -3554.
526.
J. (1978) Eur. J . Biochem. 92,349-359.
(1982) Biochemistry 21, 151 -156.
Chem. 257,8300-8307.
Methods in Enzymol. 56, pp. 734 - 749.
4764 - 4769.
114,339-347.







![Prevention of doxorubicin-induce renal function abnormalities ......ATPase, Mg2+-ATPase and Na+, K+-ATPase activities [15, 16]. Turmeric is a golden spice derived from the rhizome](https://img.pdfslide.net/doc/110x75/61385b7c0ad5d20676493447/prevention-of-doxorubicin-induce-renal-function-abnormalities-atpase-mg2-atpase.jpg)
![V-ATPase · From Wiki: Vacuolar-type H+ -ATPase (V-ATPase) is a highly conserved evolutionarily ancient enzyme with remarkably diverse functions in eukaryotic organisms.[1] membranes](https://img.pdfslide.net/doc/110x75/5fa3fb056ad5ca477269e2ce/v-atpase-from-wiki-vacuolar-type-h-atpase-v-atpase-is-a-highly-conserved-evolutionarily.jpg)