Embed Size (px)
Citation preview

et Biophysics &a ELSEVIER Biochimica et Biophysics Acta 1257 (1995) 293-296
Rapid Report
Incorporation of bovine adrenal 3 P-hydroxy+ene steroid dehydrogenase/A5-A4 isomerase into phospholipid vesicles
Takeshi Yamazaki, Shiro Kominami, Naofumi Nishida, Shigeki Takemori *
Faculty of Integrated Arts and Sciences, Hiroshima University, Higashi-hiroshima, 724, Japan
Received 20 March 1995; accepted 18 April 1995
Abstract
3PHydroxysteroid dehydrogenase/A5-A4 isomerase (3@HSD/I) and cytochrome P-450c2, were co-purified from bovine adreno- cortical microsomes by an improved method. The 3P-HSD/I was successfully incorporated into liposomal membranes in which the enzyme activity was greatly stabilized. Enzymatic activities and kinetic parameters of the 3P-HSD/I proteoliposomes were almost the same as those of the solubilized form.
Keywords: 3P-Hydroxy-5-ene steroid dehydrogenase/A’-A4-ene isomerase; F’roteoliposome; Steroidogenesis; (Bovine adrenal)
3P-Hydroxy-5-ene steroid dehydrogenase/A’-A4-ene isomerase (3P-HSD/I; E.C. 1.1.1.51 and E.C. 5.3.3.1) catalyzes both dehydrogenation and isomerization to form A4-3-ketosteroids from A5-3P-hydroxysteroids [l-3]. The 3 @-HSD/I functions in mammalian steroidogenic organs, namely adrenal cortex, testis, ovary and placenta, and also in several peripheral tissues [4]. The reactions thus play an essential role in the biosynthesis of all classes of steroid hormones [4,5]. The biosynthesis of steroid hormones from cholesterol in the adrenal cortex involves four distinct forms of cytochrome P-450 and 3@HSD/I. P-450,,, and P-450 * , a are located in the inner mitochondrial mem- branes whereas P-450,,, and P-450,,,,,,,,, are located in the membranes of the endoplasmic reticulum [5]. The 3/?-HSD/I is located in the membranes of both endoplas- mic reticulum and mitochondria [6-81. We have previously shown that adrenal cytochromes P-450 were incorporated with NADPH electron transfer components into liposome membranes and the cytochrome P-450 proteoliposomes were useful system for detailed investigations concerning molecular mechanism of adrenal steroidogenesis [9- 131.
Previous studies on purified 3/3-HSD/I have been car-
Abbreviations: 3P-HSD/I, 3/3-hydroxy-5-ene steroid dehydro- genase/A5-A4 isomerase; P-450,,, , cytochrome P-450XXIAl; P- 450 17aJyase. cytochrome P-450XVIIAl; P-450, ,b, cytochrome P- 450XIB 1; P-450,,, , cytochrome P-450XIA
* Corresponding author. Fax: + 81 824 24 0757.
0005-2760/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0005-2760(95)00108-5
ried out in the detergent-solubilized form [2,14-161, but the instability of the solubilized form has hindered the reconstitution of complete adrenal steroidogenic pathways consisting of cytochromes P-450 and 3PHSD/I.
In this study, the 3&HSD/I was purified from bovine adrenocortical microsomes by the improved method and successfully incorporated into liposome membranes. The 3@HSD/I embedded in the liposome membranes was found to be more stable than the detergent solubilized form.
3@-Hydroxysteroid dehydrogenase activity was esti- mated from the conversion rate of NAD+ to NADH [2]. The reaction was started by addition of 0.1 mM dehy- droepiandrosterone (Sigma, St. Louis, MO) into 1 ml of reaction mixture containing 100 mM potassium phosphate buffer (pH 7.5), 0.1 mM EDTA, 20% glycerol, 0.1 mM NAD+ and appropriate amount of the 3P-HSD/I at 25” C, and absorption change at 340 nm was measured with a Union giken SM-401 spectrophotometer. The 3 P-HSD/I reactions were assayed by the production of 3-keto-A4 steroid products from 3-hydroxy-A5 steroid substrates. The reactions were carried out under the same condition as the dehydrogenase assay described above. The steroid prod- ucts were extracted with chloroform and analyzed by HPLC as described previously [17].
Adrenocortical microsomes were prepared from fresh adrenals of Holstein-Friesian cattle as previously described [18]. The microsomes were solubilized with 2% sodium cholate in 50 mM potassium phosphate buffer, pH 7.5,

294 T. Yamazaki et al. /Biochimicu et Biophvsica Acta 1257 (1995) 293-296
I II 1 I 2 o.4 - ---- f / t;L /
: 2 -1 g\ 0.2- &j
P Q
/ /
rx - /
0” 00’ c -
1 50 100
Fraction No.
Fig. 1. Elution profiles of 3P-HSD/I and P-450,, , on an ez-amino-octyl Sepharose 4B column chromatography. Enzymes were eluted from the column with a linear gradient of Emulgen 913 (O-0.4%). 3P-Hydroxy- steroid dehydrogenase activity (AA,,, nm /min) was measured as de- scribed in the text (0). P-450,,, content was measured by substrate-in- duced spectral change as described elsewhere (0) [18]. Emulgen concen- tration was shown with dotted line. The profile was reproducible in seven individual experiments.
containing 0.1 mM EDTA, 0.1 mM dithiothreitol (DTT) and 20% glycerol (buffer A), and centrifuged at 105 000 X
g for 1 h. Resulting supematant contained 88% of the 3/3_hydroxysteroid dehydrogenase activity of the original microsomes. The solution was applied to an *amino-n- octyl-Sepharose 4B column (2.5 X 10 cm) which was equi- librated with buffer A containing 0.4% sodium cholate. The elution was carried out with a linear gradient from 0 to 0.4% Emulgen 9 13 in the equilibration buffer. The 3@-HSD/I and P-450,, 1 were eluted at 0.15 and 0.4% Emulgen, respectively (Fig. 1). The recoveries for both the enzymes were higher than 90% at this step. The P-450,,, fractions were frozen at - 80” C for the further purification [ 181. Fractions containing 3P-HSD/I were concentrated by a Centrifugal Ultra free Filter Unit (Millipore) and dialyzed for 3 days 4 times against 1 1 of buffer A containing 0.2% Emulgen 913 to remove cholate. The dialyzed solution was applied to a hydroxylapatite column (1.5 X 10 cm) previously equilibrated with 0.2% Emulgen 913 in 10 mM potassium phosphate buffer, pH 7.5, con- taining 10 PM NAD+, 0.1 mM EDTA, 0.1 mM DTT and 20% glycerol (buffer B). After washing the column with the equilibration buffer, the 3P-HSD/I was eluted with a linear gradient from 10 to 150 mM potassium phosphate in buffer B. The purified enzyme fractions were combined, concentrated as above, and stored at - 80” C. The final recovery of the 3&HSD/I was 65%, which was the best recovery among previously reported methods [2,14,19]. In the previous purification methods for the 3P-HSD/I and P-450,, , , only one enzyme could be purified [2,14,18,19]. The new purification method described above could co- purify 3/3-HSD/I and P-450,,, from the same starting materials.
The purified 3/3-HSD/I gave a single protein band on SDS-PAGE (data not shown). The molecular weight of the protein was estimated to be 42 000 f 1000, which was in good agreement with the value, 42093, deduced from the
cDNA analysis [20]. The specific activity of the 3/?-hy- droxysteroid dehydrogenase against dehydroepiandros- terone as a substrate was 170 nmol NADH produced/min per mg protein.
The 3@HSD/I was incorporated into phospholipid vesicles by a dialysis method [lo]. The purified enzyme was applied again to a hydroxylapatite column (1.5 X 10 cm) previously equilibrated with 0.2% Emulgen 9 13 and 0.4% sodium cholate in buffer A. Since the 3@HSD/I cannot be integrated into liposome membranes in the presence of Emulgen 9 13, the enzyme on the column was washed to remove the detergent with 0.2% sodium cholate in buffer A, and subsequently eluted with 500 mM potas- sium phosphate buffer, pH 7.5, containing 0.1 mM EDTA, 0.1 mM DTT, 20% glycerol and 0.2% cholate, and concen- trated to 1 mg protein per ml. Phosphatidylcholine (Sigma, St. Louis, MO), phosphatidylethanolamine and phospha- tidylserine (both from Lipid Products, Surrey, UK) in organic solvent were mixed at the weight ratio of 5:3:1 which resembled lipid compositions of adrenal micro- somes [21 I. [ “C]Dipalmitoylphosphatidylcholine (5000 cpm/l mg phospholipids, New England Nuclear, Boston, MI) was added to the mixture for the estimation of the recovery of the lipids in proteoliposomes. After removal of the organic solvents under vigorous N, stream, 10 mg of the lipids mixture were dispersed with vigorous ultrasoni- cation (Heat system, W-225) in 0.2 ml of 500 mM potas- sium phosphate buffer, pH 7.2, containing 5% sodium cholate, 5% n-octyl-P-D-glucoside and 0.5 mM DTT. The Emulgen-free 3P-HSD/I was added to the dispersed lipids and diluted to 1 ml. The final solution contained 10 mg/ml phospholipids, 6 mM 3@-HSD/I, 1% cholate, 1% n-octyl-k-o-glucoside, 20% glycerol, 0.1 mM EDTA and 0.1 mM DTT in 50 mM potassium phosphate buffer, pH 7.2. The mixture was dialyzed for 12 h against 50 mM Tris-HCl buffer, pH 7.2, containing 200 mM NaCl, 0.1 mM EDTA and 0.1 mM DTT. The proteoliposomes con- taining 3P-HSD/I could be prepared during the removal of cholate. The proteoliposomes composed of 3P-HSD/I and phospholipids in a molar ratio of 1:2000.
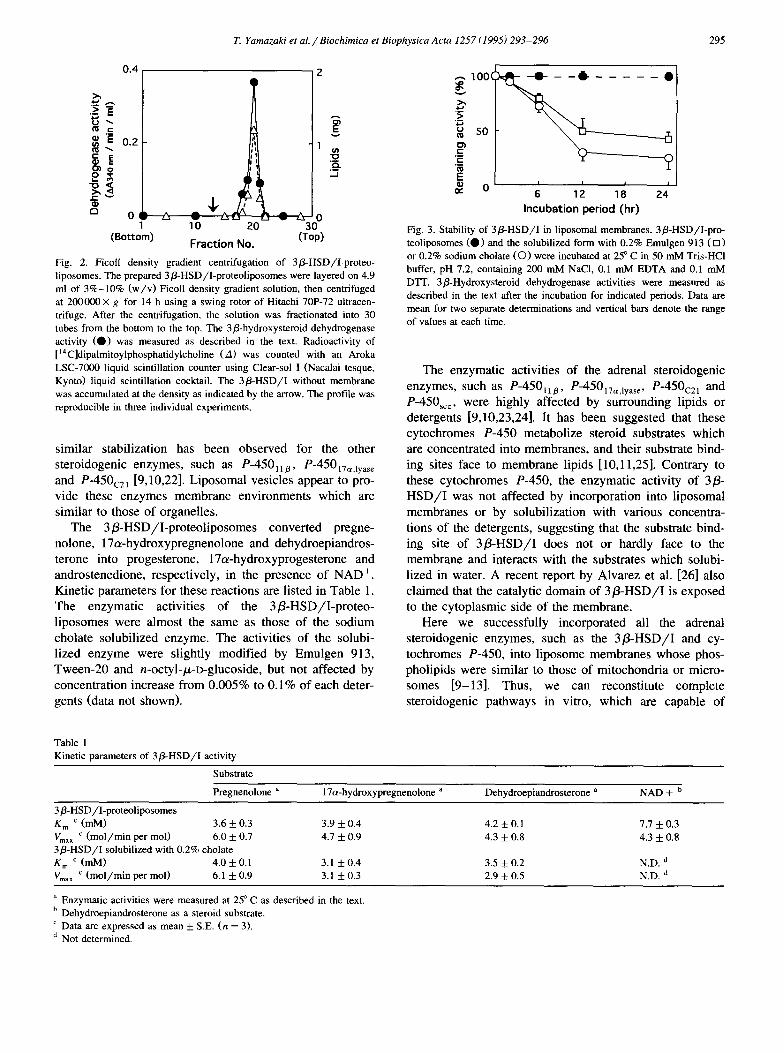
The incorporation of 3P-HSD/I into the liposomal membranes was ascertained by the Ficoll density gradient centrifugation [9]. As shown in Fig. 2, the activity of 3P-HSD/I was recovered at the same position in the density gradient as the liposomes. The phospholipid free 3P-HSD/I showed a peak at higher density (indicated as an arrow in Fig. 2) and empty liposomes at a little lower density. This result indicates the successful incorporation of the 3@-HSD/I into the phospholipid membranes.
The 3P-HSD/I was greatly stabilized by the incorpora- tion into liposome membranes. The 3@-HSD/I-proteo- liposomes sustained full activity after 24 h incubation at 25” C even in the absence of 20% glycerol (Fig. 3). On the contrary, when the proteoliposomes were solubilized with Emulgen 913 or sodium cholate, the enzyme lost the activity more than 60% by the same incubation. The
T. Yamazaki et al. /Biochimica et Biophysics Acta 1257 (1995) 293-296 295
(Bottom) Fraction No.
Fig. 2. Ficoll density gradient centrifugation of 3fi-HSD/I-proteo- liposomes. The prepared 36-HSD/I-proteoliposomes were layered on 4.9 ml of 3%-10% (w/v) Ficoll density gradient solution, then centrifuged at 200000X g for 14 h using a swing rotor of Hitachi 70F72 ultracen- trifuge. After the centrifugation, the solution was fractionated into 30 tubes from the bottom to the top. The 3phydroxysteroid dehydrogenase activity (0) was measured as described in the text. Radioactivity of [ “C]dipalmitoylphosphatidylcholine (A) was counted with an Aroka LSC-7000 liquid scintillation counter using Clear-sol I (Nacalai tesque, Kyoto) liquid scintillation cocktail. The 3@HSD/I without membrane was accumulated at the density as indicated by the arrow. The profile was reproducible in three individual experiments.
similar stabilization has been observed for the other steroidogenic enzymes, such as P-450,1p, P-450,,,,,,,,, and P-450,, , [9,10,22]. Liposomal vesicles appear to pro- vide these enzymes membrane environments which are similar to those of organelles.
The 3 P-HSD/I-proteoliposomes converted pregne- nolone, 17~hydroxypregnenolone and dehydroepiandros- terone into progesterone, 17a-hydroxyprogesterone and androstenedione, respectively, in the presence of NAD+. Kinetic parameters for these reactions are listed in Table 1. The enzymatic activities of the 3 @HSD/I-proteo- liposomes were almost the same as those of the sodium cholate solubilized enzyme. The activities of the solubi- lized enzyme were slightly modified by Emulgen 913, Tween-20 and n-octyl-p-D-glucoside, but not affected by concentration increase from 0.005% to 0.1% of each deter- gents (data not shown).
Table 1 Kinetic parameters of 3PHSD/I activity
‘E ._
E P d 0 6 12 18 24
Incubation period (hr)
Fig. 3. Stability of 3&HSD/I in liposomal membranes. 3S-HSD/I-pro teoliposomes (0 1 and the solubilized form with 0.2% Emulgen 913 (0 1 or 0.2% sodium cholate (0) were incubated at 25” C in 50 mM Tris-HCl buffer, pH 7.2, containing 200 mh4 NaCl, 0.1 mM EDTA and 0.1 mM D’IT. 3S-Hydroxysteroid dehydrogenase activities were measured as described in the text after the incubation for indicated periods. Data are mean for two separate determinations and vertical bars denote the range of values at each time.
The enzymatic activities of the adrenal steroidogenic enzymes, such as P-450, 1 B, P-450 17aJyase P-450,,, and P-450,, , were highly affected by surrounding lipids or detergents [9,10,23,24]. It has been suggested that these cytochromes P-450 metabolize steroid substrates which are concentrated into membranes, and their substrate bind- ing sites face to membrane lipids [ 10,11,25]. Contrary to these cytochromes P-450, the enzymatic activity of 3/3- HSD/I was not affected by incorporation into liposomal membranes or by solubilization with various concentra- tions of the detergents, suggesting that the substrate bind- ing site of 3P-HSD/I does not or hardly face to the membrane and interacts with the substrates which solubi- lized in water. A recent report by Alvarez et al. [26] also claimed that the catalytic domain of 3@HSD/I is exposed to the cytoplasmic side of the membrane.
Here we successfully incorporated all the adrenal steroidogenic enzymes, such as the 3@-HSD/I and cy- tochromes P-450, into liposome membranes whose phos- pholipids were similar to those of mitochondria or micro- somes [9-131. Thus, we can reconstitute complete steroidogenic pathways in vitro, which are capable of
Substrate
Pregnenolone a 17a-hydroxypregnenolone a Dehydroepiandrosterone a NAD+ b
3@HSD/Iproteoliposomes K, c (mM) 3.6 + 0.3 3.9 f 0.4 4.2 i- 0.1 7.7 * 0.3 V ’ max (mol/min per mol) 6.0 + 0.7 4.7 + 0.9 4.3 f 0.8 4.3 f 0.8 3P-HSD/I solubilized with 0.2% cholate K, c (mM) 4.0 + 0.1 3.1 + 0.4 3.5 f 0.2 N.D. d V ’ mar (mol/min per mol) 6.1 + 0.9 3.1 k 0.3 2.9 f 0.5 N.D. d
a Enzymatic activities were measured at 25” C as described in the text. b Dehydroepiandrosterone as a steroid substrate. ’ Data are expressed as mean + S.E. (n = 3). d Not determined.

296 T. Yamazaki et al. / Biochimica et Biophysics Acta 1257 (1995) 293-296
producing glucocorticoids, mineralocorticoids and andro- gen from cholesterol.
Acknowledgements
This work was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Sci- ence and Culture of Japan.
References
[l] Ford, H.C. and Engle, L.L. (1974) J. Biol. Chem. 249, 1363-1368. [2] Ishii-Ohba, H., Saiki, N., Inano, H. and Tamaoki, B.-I. (1986) J.
Steroid B&hem. 24, 753-760. [3] Lorence, M.C., Murry, B.A., Trant, J.M. and Mason, J.I. (1990)
Endocrinology 126, 2493-2498. [4] Labrie, F., Simard, J., Luu-The, V., Belanger, A. and Pelletier, G.
(1992) J. Steroids Biochem. Mol. Biol. 43, 805-826. [5] Takemori, S. and Kominami, S. (1984) Trends Biochem. Sci. 9,
393-396. 161 Chapman, J.C. and Sauer, L.A. (1979) J. Biol. Chem. 254, 6624-
6630. [7] Cherradi, N., Defaye, G. and Chambaz, E.M. (1993) J. Steroid
Biochem. Mol. Biol. 46, 773-779. [8] Cherradi, N., Defaye, G. and Chambaz, E.M. (1994) Endocrinology
134, 1358-1364. [9] Ikushiro, S., Kominami, S. and Takemori, S. (1989) Biochim.
Biophys. Acta 984, 50-56.
1101
1111
[121
1131
[141
iI51
1161
[171
iI81
[191
La
Dll WI
[231
L-J.41
1251
[261
Kominami, S., Higuchi, A. and Takemori, S. (1988) Biochim. Biophys. Acta 937, 177-183. Kominami, S., Itoh, Y. and Takemori, S. (1986) J. Biol. Chem. 261, 2077-2083. Kominami, S., moue, S., Higuchi, A. and Takemori, S. (1989) Biochim. Biophys. Acta 985, 293-299. Ikushiro, S., Kominami, S. and Takemori, S. (1992) J. Biol. Chem. 267, 1464-1469. Thomas, J.L., Berko, E.A., Faustino, A., Myers, R.P. and Strickler, R.C. (1988) J. Steroid B&hem. 31, 785-793. Thomas, J.L., Myers, R.P. and Strickler, R.C. (1989) J. Steroid Biochem. 33, 209-217. Ishii-Ohba, H., Saiki, N., Inano, H. and Tamaoki, B.-I. (1986) J. Steroid Biochem. 25, 555-560. Kominami, S., Ogawa, N., Morimune, R., De-Ying, H. and Take- mori, S. (1992) J. Steroid Biochem. Mol. Biol. 42, 57-64. Kominami, S., Ochi, H., kobayashi, Y. and Takemori, S. (1980) J. Biol. Chem. 255, 3386-3394. Rutherfurd, K.J., Chen, S. and Shively, J.E. (1991) Biochemistry 30, 8108-8116. Zhao, H.-F., Simard, J., Labrie, C., Breton, N., Rheaume, E., Luu-The, V. and Labrie, F. (1989) FEBS Lett. 259, 153-157. Narasimhulu, S. (19751 Adv. Exp. Med. Biol. 58, 271-286. Kominami, S., Tagashira, H., Ohta, Y., Yamada, M., Kawato, S. and Takemori, S. (1993) Biochemistry 32, 12935-12940. Kominami, S., Hara, H., Ogishima, T. and Takemori, S. (1984) J. Biol. Chem. 259, 2991-2999. Takikawa, O., Gomi, T., Suhara, Itagaki, E., Takemori, S. and Katagiri, M. (1978) Arch. Biochem. Biophys. 190, 300-306. Seybert, D.W., Lancaster, Jr., J.R., Lambeth, J.D. and Kamin, H. (1979) J. Biol. Chem. 254, 12088-12098. Alvarez, CL, Genti-Raimondi, S., Patrito, L.C. and Flury, A. (1994) Biochim. Biophys. Acta 1207, 102-108.





























