Embed Size (px)
Citation preview
Induced Responses to Herbivory and Increased Plant Performance
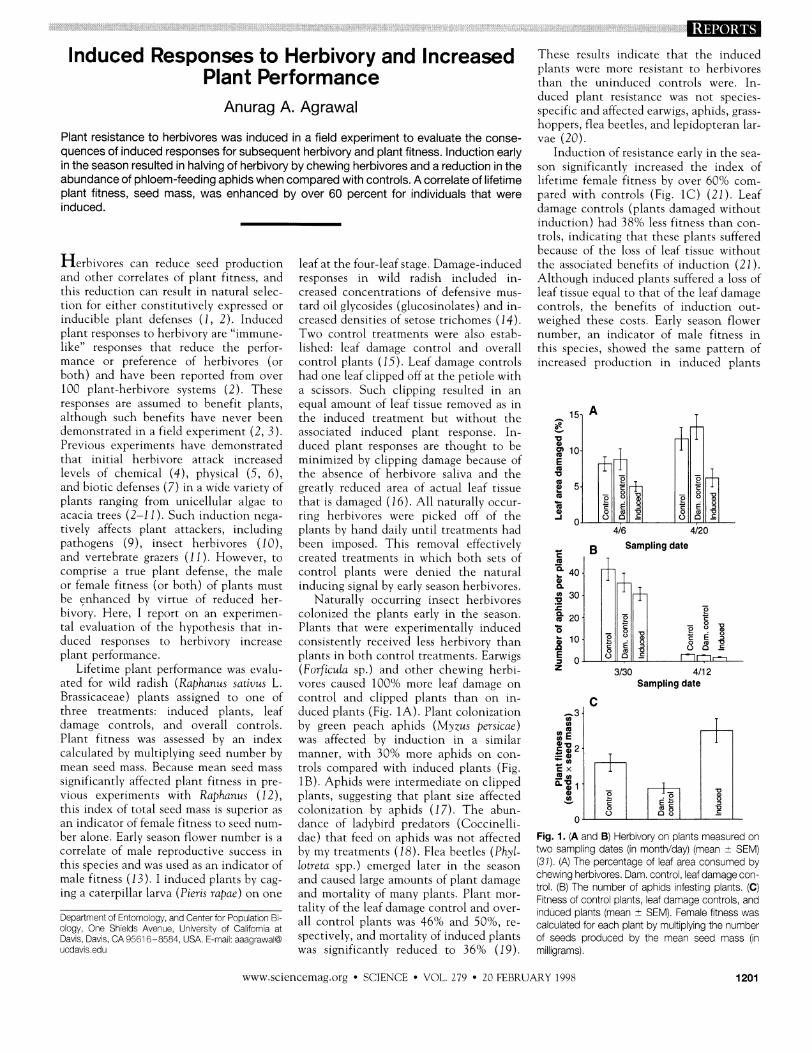
These results indicate that the induced plants were tnore resistant to herbivores than the uninduced controls were. In- duced plant resistance was not species- specific and affected earwigs, aphids, grass- hoppers, flea beetles, and lepidopteran lar- vae 120).
Anurag A. Agrawal
Plant resistance to herbivores was induced in a field experiment to evaluate the conse- quences of induced responses for subsequent herbivory and plant fitness. Induction early in the season resulted in halving of herbivory by chewing herbivores and a reduction in the abundance of phloem-feeding aphids when compared with controls. A correlate of lifetime plant fitness, seed mass, was enhanced by over 60 percent for individuals that were induced.
~,
Induction of resistance early in the sea- son significantly increased the index of lifetime female fitness by over 60% com- pared with controls (Fig. 1C) (21). Leaf datnage controls (plants datnaged without induction) had 38% less fitness than con- trols, indicating that these plants suffered because of the loss of leaf tissue without
Herbivores can reduce seed nroduction leaf at the four-leaf stage. Damage-induced responses in wild radish included in- creased concentrations of defensive mus- tard oil glycosides (glucosinolates) and in- creased densities of setose trichomes 114).
the associated benefits of induction (21). Although induced plants suffered a loss of and other correlates of plant f~tness, and
this reduction can result in natural selec- "
leaf tissue equal to that of the leaf damage controls, the benefits of induction out- tion for either constitutively expressed or
inducible plant defenses (1. 2). Induced weighed these costs. Early season flower nutnber, an indicator of tnale fitness in plant responses to herbivory are "immune-
like" resmmses that reduce the nerfor-
\ . , Two control treatments were also estab- lished: leaf damage control and overall this specles, showed the same pattern of
increased production In induced plants mance or preference of herbivores (or both) and have been reported frotn over 100 plant-herbivore systetns (2). These responses are assumed to benefit plants, although such benefits have never been detnonstrated in a field experitnent (2, 3) . Previous experiments have demonstrated
- control plants (15). Leaf damage controls had one leaf clipped off at the petiole with a scissors. Such clipping resulted in an euual atnount of leaf tissue removed as in the induced treatment but without the associated induced plant resnonse. In- duced plant responses are thought to be minimized by clipping damage because of the absence of herbivore saliva and the greatly reduced area of actual leaf tissue that is damaged (16). All naturally occur- ring herbivores were picked off of the plants by hand daily until treatments had been imposed. This removal effectively created treatments in which both sets of control plants were denied the natural inducing signal by early season herbivores.
Naturally occurring insect herbivores colonized the plants early in the season. Plants that were experimentally induced consistentlv received less herbivorv than
that initial herbivore attack increased levels of chemical (4), physical (5, 6) , and biotic defenses (7) in a wide variety of plants ranging from unicellular algae to acacia trees (2-1 1 ). Such induction nega- tively affects plant attackers, including pathogens (9), insect herbivores ( l o ) , and vertebrate grazers 11 1 ). However. to
date " , ,
cotnprise a true plant defense, the male or female fitness (or both) of plants must , L
be enhanced by virtue of reduced her- bivory. Here, I report on an experitnen- tal evaluation of the hypothesis that in- duced responses to herbivory increase plant performance.
Lifetime plant performance was evalu- ated for wild radish (Raphanus sativus L. Brassicaceae) plants assigned to one of three treatments: induced plants, leaf damage controls, and overall controls. Plant fitness was assessed by an index calculated by multiplying seed number by mean seed mass. Because mean seed mass significantly affected plant fitness in pre- vious experiments with Raphanus (1 2), this index of total seed mass is superior as an indicator of female fitness to seed num-
plants in both control treatments. ~ a r w i ~ s (Forficula sp.) and other chewing herbi- vores caused 100% tnore leaf damage on control and clipped plants than on in- duced plants (Fig. 1A). Plant colonization by green peach aphids (bf>zus persicae) was affected by induction in a similar manner, with 30% more anhids on con-
Sampling date
trols compared with induced plants (Fig. 16) . Aphids were intermediate on clipped plants, suggesting that plant size affected colonization by aphids (17). The abun- dance of ladybird predators (Coccinelli- dae) that feed on aphids was not affected by my treatments (18). Flea beetles (Phyi- lotreta spp.) emerged later in the season and caused large atnounts of plant damage and mortality of many plants. Plant mor- tality of the leaf damage control and over- all control plants was 46% and 50%, re- spectively, and mortality of induced plants was significantly reduced to 36% (19).
ber alone. Early season flower nutnber is a correlate of male re~roductive success in
Fig. 1. (A and B) Herbvory on plants measured on two sampl~ng dates (In month/day) (mean i- SEM) (37). (A) The percentage of leaf area consumed by chew~ng herbivores. Dam. control, leaf damage con- trol. (B) The number of aphids infesting plants. (C) Fitness of control plants, leaf damage controls, a ~ d induced plants (mean 2 SEMI. Female f~tness was calculated for each plant by mult~plyng the number of seeds produced by the mean seed mass (In m~ll~grams).
this species and was used as an indicator of male fitness (13). I induced plants by cag- ing a caterpillar larva (Pieris rapae) on one
Department of Eritomoogy, and Ceriter for Popuaton B - ology, One Shields Avenue, Univers~ty of Caforri~a at Dav~s. Dav~s. CA 9551 6-8584, USA. E-mat aaagrawam ucdavs edu
wwu,.Sclencemag.org SCIENCE VOL. 279 2L1 FEBRU,

(22) . Th i s increase in the fitness of in duced plants was found even though wild radish can tolerate large alllounts of her- hivory without reductions in fitness (23, 24). Only two previous experiments have attempted to detect fitness benefits of in- duced responses to herbivory; these exper- iments failed t o detect fitness benefits, probably because they involved long-lived perennial plants (25) .
Induced responses to herbivory appear to be a n exalnple of a d a p t ~ v e plasticity in plants. Iniluced responses are nearly ubiq- uitous in the plant kingdom, yet it is still not u~lderstooil why plants have evolved facultative defenses instead of const~tut i \ ,e defenses. Energetic costs associated with plant defense have been suggested as the primary force favoring facultative defenses because of enertretic savines when the de- fenses are no t neeiled (26) . However, ev ide~ lce for such costs is weak 12. 27) Recent work has ilnplicated a broad array of o ther ecological and physiological trade-offs that nlay be inlportant con- straints o n plant defense favoring 11lduc- ible strategies (2 , 27). Al though the ben- efits of induction have received relatively little attention, inducible defenses may have many benefits for plants that consti- tutive ilefenses lack (28) . For example, induced responses may attract preilators and parasites of herbivores (29) . In addi- t ion, because some plant resistance traits attract specialist herbivores, i n i h ~ c ~ b l e de- fenses may make plants less obvious tar- gets for these adapted herbivores when the plants are not induced (30).
Deterlninlllg the ultimate fitness con- sequences of plant defense traits bears di- rectly o n the long-standing vlen. of plant parasites as selective agents anil plants as active participants in the evolutionary process ( I ). Induceil plant responses to h e r b i ~ ~ o r y provided high levels of plant defense that were reflected in increased plant performance. Leaf damage controls had lower fitness than overall controls, indicating that early season he rb~vory was costly. In this experiment, fitness benefits of induced defenses outweighed the costs associated with loss of leaf tissue and in- duction ~tsel f . T h e excess of benefits to costs makes induced defense a strategy that may be favored by natural selectloll in environments with herbivory.
REFERENCES AND NOTES
1. R. J. Marqu~s, Science 226. 537 (1984); E. L. S~mn-1s and M. D. Rausher, Evolution 43, 573 (1989): R. J. Marqus, In Plant Resistance to Herbi- vores and Pathogens. Ecology, Evolut!on and Ge- netics, R. S Fritz and E. L. Slmms, Eds. (Univ. of Chcago Press Chcago, L , 1992), pp. 392-425; J. Nunez-Farfan and R. Dlrzo. Evolution 48. 423 (1 994).
2. R. Karban arid I . T. Badwn , Induced Responses to Herbivory (Unv, of Chlcago Press, Chicago, L , 1997).
3 T R Green and C. A. Ryan, Science 175, 776 (1972); R. Karban and J. R. Carey, ibid. 225. 53 (1984); E Haukiola arid S. Neuvonen. Ecology 66. 1303 (1 995): I. T. Badwn , Oecologia 75. 367 (1 988); E. N Wold and R. J Marqu~s, Ecology 78, 1356 (1 997)
4 I T Baldwin, In Pl?ytochemical Induction by Herbi- vores, D W. Taalny and M. J. Raupp, Eds. (Wley. New York, 1991), pp. 47-69, M. J. Stout. K. V. Worklnan, S. S. Duffey. Entomol Exp. Appl 79, 255 (1996): J. L. BI, J. B. Murphy. G. W. Feton, J Chem. Ecol. 23, 97 (1 997)
5. T. P Young, Oecologia 71, 436 (1987); R. Baur, S. Blnder, G Benz. ibid. 87, 219 (1991), J. R Obeso. Plant Ecol. 129, 149 (1 997).
6 A A. Agrawal, in preparation. 7. L. A Mound. E~np . Cotton Grow. Rev 39. 254
(1962), S Koptur, n The E~/oIutiona!y Ecology of Plants. J. H. BockaridY B. L~riiiart. Eds. (Westvew, Boulder, 1989), pp. 323-339; L L. Smth, J. Lanza, G. C Smth. Ecology 71, 107 (1 990): A. A Agrawal, ibid., In press
8. L. A. Hansson, Proc. R Soc London Ser. B 263. 1241 (1996): G V. \A!olfe, M. Stellike. G. O Klrst. Nature 387. 894 (1 997)
9. J Kuc. Bioscience 32, 954 (1 982): r i Innovative Ap- pioacI?es to Plant Disease Control, I. Chet, Ed. (Wley, New York, 1997), pp 255-274; R Karban, R. Adamchak. \A! C Schnatiiorst, Science 235. 678 (1 987), E. L. Smms and T. J Vson, Oecologia 102, 494 (1995), M Schrieider, P Schweizer, P. Meuwy. J. P. Metraux, lnt Rev. Cytol. 168, 303 (1996).
10. E. Haukoa and P Niemela, Ann. Zool. Fen17 14, 48 (1977); R. Karban, Ecology 74, 1 (1993); M. J. Stout arid S S Duffey, Entomol. Exp. Appl 79,273 (1 996)
11 J P Bryarit, Science 213. 999 (1991), A. V. Mewskl, T P Young, D Madden, Oecologia 86, 70 (1991); K A Ernest, Ecology 75, 1684 (1994); T. Seldal, K. J Andersen, G. Hogstedt. Oikos 70, 3 (1 994).
1 2. M L. Siantori, Ecology 65, 1 1 05 (1 994). 13. T.-L Ashman, F L. Galloway, M. L. Stanton, Oeco-
logia 96, 132 (1 993) 14. Chemlcal analyses ndlcated that leaf damage by
Pieris rapae caterpillars nduced hgh coricentrations of n d o e glucosinolates. whereas leaf dalnage by scssors alone d d not induce iridoe glucosinolates [F(1 ,30) = 6 490 P = 0 0161 (24). Setose trchomes were also found to be ~riduc~ble (ricrease In densty and total number) on newly formed leaves of prevl- ously dalnaged plarits [F(1,36) = 10.209. P = 3.3031 (6).
15. A total of 454 plants were dvlded n t o three treat- ments as follows: 151 induced plants, 150 leaf dam- age controls, and 153 overall (unmanipuated) con- trols. Half of the leaf damage control plants were clipped at the start of the treatments (that 1s when caterpillars were placed on induced treatment plants), and the other half were c p p e d when the caterpars on the Induced treatments had flnshed consumng the leaf (3 to 4 days later). The research po t was located In a plowed old f e d at the Universty of Caiforna Student Expermental Farm (Davs, CA). Seeds were collected from several wild popuatons of R, sativus in Davis and cotyledonary plants were transplanted from p ~ ~ g trays durng tlie t ypca perlod of juvene growth. The plot was not Irrigated.
16. I . T. Baldwin Trends Ecol. Evol. 5. 91 (1990); W. J. Mattson and S. R. Palmer, In Mechanisms of IUoody Plant Defenses Against Insects. Search for Pattern, W J Mattson, J. Levleux. C. Bernard-Dagan Eds. (Springer-Verag Berlin, 19881, pp 157-1 70, L. Mat- t acc . M. Dcke. M. A. Posthun-~~s. Proc. Natl. Acad. Sci, 0.S.A. 92, 2036 (1995). H. T. Alborn et a/., Scleiice 276, 945 (1 997).
17. Colonizing aphds choose larger plants over smaller plants. T . G . Whitham, Am. at. 155, 449 (1980): W. V. Zucker, Ecology 63.972 (1 982); A. A. Agrawal, ~ ~ n p u b s h e d data.
18. From the analysis of varance for the effect of treat- lnents on abundance of adybrd predators. F(2,451)
= 3.1 19, P = 0.889. From the l~kel~hood ratio x2 test for the effect of flea beetles on plant mo-taty, G = 7.532. degrees of freedom = 2, P = 0.023. In den tca experimerits peiormed or? a s b n g spe- ces (Raphanus raphanistrum) at a slte In the Serra Nevada foo th~s , I found s m a r results (G) , In these experiments, riduced responses to herblvory pro- tected parits from herbvory by grasshoppers and lepidopteran awae arid Increased flower producton arid seed set colnpared \vth control plants. Overall effects of treatments or? female plant f~triess are s~gnificant F(2.445) = 11.004. P i 3.3011: For the contrast of Induced parits colnpared \vth o\~erall controls, F( l ,445) = 6.567, P = 0.01 1, arid for the contrast of leaf dalnage coritrols compared w ~ t h o\~erall coritrols, F(1.445) = 4.954, P = 0.027. From the ariayss of vararice or? early season flower production, F;2,451) = 3 473, P = 0 032. R. Maurco, M. D Bowers. F. A. Bazzaz. Ecology 74, 2066 (1 993). A. A. Agrawal, S. Y. Strauss, M . J. Stout, In preparation. R. Karban, Entomol Exp. Appl 42.239 (1 986); Ecol- ogy 74, 9 (1 993). E. Haukiola and T Hakaa, Rep. Kevo Subarctic Res. Stat. 12, 1 (1975), D. F Rhoades, r i Herbivores. Their Interaction Yhth Secondar)) Plant Metabohtes, G. A Rosentha and D. H. Janzen Eds. (Academic Press, New York, 1979), pp. 3-54: C D. Ha lve, Q Rev. Biol 65, 323 (1990); A. R Zanger and F A. Bazzaz, In Plant Resistance to Herbivores and Patl?ogens. Ecology, El/olution, and Genetics, R S. Frltz and E, L. Smms, Eds. (Unv, of Chicago Press, Chcago, L , 1992), pp. 363-391; D. A. Herms arid W. J. Mattson, Q. Rev Biol. 67, 283 (1 992). E L. Simms, r i Plant Resistance to Herbivores and Pathogens. Ecology, E~/olution, and Genetics, R S Frltz and E L. Smms, Eds. (Unv, of Chicago Press, Chcago, L . 1992), pp. 363-391, A. A Agrawal and R. Karban, n Ecology and E~/olution oflnducible De- fenses, R. T o r a n and C. D. H a ~ ~ e l l , Eds. (Prncetori Unv. Press, Pririceton. NJ, in press). R. Karban, A A Agrawal, M. Marigel, Ecology 78, 1351 (1 997). T. C. J. Turngs J. H. Tuminsori, W J. Lewis, Sci- ence 250, 1251 (1993); B Drukker, P. Scutareanu, M. W. Sabes, Entomol. Exp. Appl. 77. 193 (1 995); J. Takabavash and M. Dicke. Trends Plant Sci 1 . 109 (1996); ?. Shmoda, J. ~akkbayash, W. ~shiha;a. A Takafull. J. Chem Ecol. 23, 2333 (1997). C. R. Carroll and C A. Hoffman, Science 209, 414 (1980); D. \A!. Taamy and E. S. McCoud. n Phyto- chemicallnd~ict~oii by Herbivores, D. W. Talamy and M. J. Raupp. Eds. (Wley, New York. '991), pp. 155-1 81 : A . Gamoustaris and R. Mithen.Ann Appl. 5/01, 126, 347 (1 995). From the anayss of variance for the effects of exper- mental nductlon on (I) chewlng herblvory and (li) plant colonizat~on by aphds measured on two sam- pling dates w ~ t h multivarlate analysis of variance, Wlks' lambda = 0.972, F(4 888) = 3.23 P = 0.012 for chewing herbvory: and Wlks lambda = 0.956, F(4,876) = 4.985, P = 0.001 for plant colonizat~on by aphids. Half-sib f a m e s also vared In their suscep- tibity to herbvoryand attack by aphds !Wks ' amb- da = 0.834 F(30 1694) = 2.625, P i 0.001]. I thank R. Karban, M. L. Stanton, B. L. Fisher, J L. Maron, J. A Rosenhen-I, T. G. Whtham, J S. Thaler, and K J. Baxter for helpful revews and discusson; S. Y. Strauss for introducing me to the w d radish system: M . Van Horn for og~st~ca l suppot at the Universlty of Caforna. Davis. Experimental Farm and K. J. Baxter, C. Wardaw, J. S. Thaler, C. E. Christan, S. M. Epply, Y. Sherman, and J. M . K n - skern for help w ~ t h the field work and seed countng. Supported by the Center for Popuaton Boogy at Unversity of Caforna. Davis, the Nothern Caforna Chapter of P h Beta Kappa, and NSF Doctoral Dls- sertatlon Improvement grant DEB-9701 109.
30 September 1997, accepted 23 December 1997
1202 SCIENCE * VOL. 279 20 FEBRUARY 1998 wna~.scienc





























