Embed Size (px)
Citation preview

[CANCER RESEARCH 47, 2236-2242, May 1, 1987]
Induction of Differentiation of Human Myeloid Cell Lines by Tumor NecrosisFactor in Cooperation with la,25-Dihydroxy vitamin D3*
Giorgio Trinchieri,2 Mark Rosen, and Bice Perussia3
The Wistar Institute of Anatomy and Biology, 3601 Spruce Street, Philadelphia, Pennsylvania 19104
ABSTRACT
We analyzed the combined effect of tumor necrosis factor and la,25-dihydroxyvitamin 1)i on the differentiation of human myeloid cell linesIII -6(1,ML3, and U937. The two compounds synergize in inducing themorphological, phenotypic, enzymatic, and functional characteristics ofcells of the monocytic lineage. Immune 7-interferon synergizes with eachcompound to induce differentiation. However, recombinant tumor necrosis factor is much more effective than recombinant -y-interferon in potentiating the effect of la,2S-dihydroxyvitamin 1)( and, alone, is also moreeffective than recombinant 7-interferon in inducing expression of thehigh-affinity Fc receptor on ML3 cells. The possible physiological orpathological relevance of the synergistic effect of tumor necrosis factorand la,25-dihydroxyvitamin D3on monocytic differentiation is discussed.
INTRODUCTION
Leukemia-derived human myeloid cell lines, such as thepromyelocytic HL-60 (1) and ML3 (2) and the histiocytic U937(3), can be induced to differentiate in vitro by several syntheticand natural inducers and are often used as models to studyterminal differentiation of myelomonocytic cells. Dimethylsulf-oxide (4), retinoic acid (5), and other substances induce HL-60cells to differentiate along the myeloid pathway, whereas phor-bol diesters (6), 1,25(OH)2D3" (7), and leukocyte products contained in medium conditioned from PHA-CM (8) induce themto differentiate into cells with characteristics of monocyte/macrophages. HL-60 and U937 cells bear specific receptormolecules for 1,25(OH)2D3 (9,10); an HL-60 blast cell line (9),which does not differentiate in response to 1,25(OH)2D3, expresses only about 8% of the number of 1,25(OH)2D3 receptorsas the parental responsive cell line. HL-60 cells treated with1,25(OH)2D3 have been reported (7) to differentiate to cellswith the differentiated morphology of metamyelocytes andmonocytes, capable of phagocytosis and NBT reduction. Thesecells have been identified as monocyte/macrophages becausethey adhere to glass, contain nonspecific esterase, and expressmonocyte-specific enzymes (9, 11).
Partial monocytic differentiation of human myeloid cell lines(12-17) is induced by IFN-7, a potent inducer of the high-affinity FcR for monomeric IgG (13, 18) and of class II majorhistocompatibility antigens (19). IFN-7 is present in PHA-CMbut in concentrations insufficient to account for its differentiation-inducing activity, and other differentiation-inducing factors have been detected in IFN-7-depleted PHA-CM that syn-
Received 10/27/86; revised 1/30/87; accepted 2/2/87.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported, in pan, by USPHS Grants CAI08IS, CA20833,CA32898, CA37155, and CA40256.
3To whom requests for reprints should be addressed, at the Wistar Instituteof Anatomy and Biology, 3601 Spruce Street, Philadelphia, PA 19104.
J Scholar of the Leukemia Society of America.4The abbreviations used are: 1,25(OH)jD3,1 a,25-dihydroxyvitamin D3; PHA-
CM, medium conditioned by phytohemagglutinin-activated leukocytes; NBT,nitroblue tetrazolium; UN-,, immune interferon; rll N-->.recombinant immuneinterferoni FcR, Fc receptor, TNF, tumor necrosis factor, rTNF, recombinanttumor necrosis factor, II. lymphotoxin; Ab-CMC, antibody-dependent cell-mediated cytotoxicity; FBS, fetal bovine serum; ANAE, a-naphthyl acetate esterase; FITC, fluorescein isothiocyanate; [3H]dTdR, [3H]thymidine.
ergize with IFN-7 in inducing differentiation (16). IFN-7 wasshown to synergize with 1,25(OH)2D3 in inducing differentiation of human myeloid cell lines (20, 21). Also, a synergismbetween IFN-7 and 1,25(OH)2D3 was observed in the inductionof class II major histocompatibility antigens on murine WEHI-3 myelomonocytic cells (22). Interestingly, IFN-7 has beenshown to stimulate synthesis of 1,25(OH)2D3 by normal humanmacrophages (23). This interplay between IFN-7 and1,25(OH)2D3 may suggest a role for 1,25(OH)2D3 in the regulation of hematopoiesis by IFN-7-producing lymphocyte subsets, i.e., T and natural killer cells.
TNF and LT are two partially homologous factors originallydescribed on the basis of their cytostatic effects on tumor celllines, TNF as a tumor cytotoxic factor present in the serum ofanimals injected with endotoxin (24), and LT as a class ofcytotoxic/cytostatic substances released by lymphocytes uponantigenic or mitogenic stimulation (25, 26). The cytotoxic/cytostatic effects of both cytokines are potentiated by IFN-7(27, 28). Recently, purification and cloning of the genes forTNF and LT have allowed more detailed studies of the biologyof these factors (29-33). The two cytotoxins also appear tomediate regulatory effects on various cell types, e.g., they act asgrowth factors on fibroblasts (34,35) and regulate proliferation,differentiation, and functions of myelomonocytic cells at allstages of differentiation (36). In particular, we (36, 37) showedthat TNF and LT, at concentrations of the order of 10~" M,
induce monocytic differentiation of human myeloid cell lines.After 5 days of culture in the presence of TNF or LT, asignificant proportion of the HL-60 myeloid cells express monocyte differentiation antigens and nonspecific esterase activityand become able to reduce NBT and to mediate low levels ofAb-CMC against tumor target cells. These markers of differentiation, however, are maximally induced when IFN-7 is present simultaneously with the cytotoxin, and the two classes ofcytokines act synergistically to induce terminal differentiation.The appearance of monocytic antigens is accompanied by acquisition of the morphology and functional properties of maturemonocytic cells, such as chemiluminescence and phagocytosis,and by expression of FcR for monomeric IgG. A decrease incell proliferation accompanies induced differentiation.
The differentiation induced by TNF on myeloid cell linesclosely resembles that induced by 1,25(OH)2D3. We report herethat TNF and 1,25(OH)2D3 in combination induce differentiation of the HL-60, ML3, and U937 human cell lines. We showthat the interaction of the two differentiation-inducing compounds is synergistic, especially when the compounds are usedat suboptimal concentrations.
MATERIALS AND METHODS
Cell Lines. All cell lines were grown in RPMI 1640 medium (FlowLaboratories, Rockville, MD). The human promyelocytic cell line HL-60 was maintained in medium supplemented with 15% FBS (GIBCO,Grand Island, NY), the human promyelocytic ML3, the histiocyticU937, the B-lymphoblastoid RPMI 8866, the bladder carcinoma 5637,and the mouse mastocytoma line P815y were maintained in medium
2236
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from
INDUCTION OF DIFFERENTIATION BY TNF AND 1,25(OH)2D,
supplemented with 10% FBS. All cell lines used in this study weremycoplasma-free.
Monoclonal Antibodies. Monoclonal antibodies BS2.1, B33.1, andB147.2 were produced and characterized in our laboratory; OKM1, and5E9 were produced from cell lines obtained from the American TissueCulture Collection (Rockville, MD); 3G8 was kindly donated by Dr. J.Unkeless (Mount Sinai Hospital, New York) and KuFc79 was a gift ofDr. P. Gambel (Medical College of Virginia). Antibody OKM1 (IgG2b,38) reacts with an epitope on the C3bi receptor which is expressed onall differentiated neutrophilic granulocytes and monocytes and which,during differentiation, appears at the myelocytic and promyelocyticstages. Antibody BS2.1 (IgM, 16, 39) reacts with all peripheral bloodmonocytes and all ANAE-positive cells in the bone marrow, and cross-competes for binding to monocytes with antibody Mo2 (40, 41). Antibody B33.1 (IgG2a, 42) reacts with class II HLA antigens. Antibody5E9 (IgGl, 43) reacts with the transferrin receptor. Antibody 3G8(IgGl, 44) reacts with a surface molecule with a molecular weight of50,000-72,000 identified as the low-affinity receptor for aggregatedIgG on neutrophilic granulocytes, natural killer cells, and macrophages.B147.2 is a murine monoclonal IgG2a of unknown specificity thatbinds in monomi-rii- form to the II-Vy ¡nilueibli-FcR on monocytes
and myeloid cell lines. The binding of B147.2 to cells is Fc fragmentdependent and is blocked by monomeric human IgGl and murineIgG2a and IgG3. Sepharose-linked B147.2 precipitates from human
monocytes and myeloid cell lines a molecule with a molecular weightof 70,000, identified as the high-affinity FcR for monomeric IgG.Antibody KuFc79 (IgG2b, 45) reacts with an FcR species (M, 42,000)present on monocytes, granulocytes, and B-cells.
Cytokine Preparations and 1,25(011);.I),. Purified human recombinant IFN (rIFN-7) from Escherichia coli was kindly supplied by Dr. H.M. Shepard (Genentech Inc., South San Francisco, CA) and has anantiviral activity titer of 7 x IO7 U/mg on He I a cells. Human rTNF(90% pure, IO7 U/mg on L-929, a subline, cells) was kindly supplied
by Dr. J. S. Price (Cetus Corp., Emeryville, NJ). 1,25(OH)2D3 was agenerous gift from Dr. M. Uskokovic (Hoffmann-LaRoche, Nutley,NJ). In order to evaluate whether a given effect of a combination oftwo cytokines is additive, antagonistic, or synergistic, isobologramanalysis, as previously described (37), was used.
Induction of Myeloid Cell Lines. 111-60 cells were seeded at 1.5 x10s cells/ml in RPMI 1640 supplemented with 15% FBS. ML3 andU937 were seeded at 1 x 10s and 0.75 x 10s cells/ml, respectively, in
medium supplemented with 10% FBS. The various inducers of differentiation were present throughout the culture period, as indicated.
Cell Morphology and Cytochemistry. Cell morphology was examinedon May-Grunwald-Giemsa stained cytocentrifuge preparations (Cyto-spin centrifuge, Shandon Southern Instruments, Inc., Sewickley, PA).Staining for ANAE was according to Platt (46). Ability of the cells toreduce nitroblue tetrazolium (Sigma Co.) was determined by mixing ona microscope slide 1 volume of cells in medium (5 x IO6cells/ml) and1 volume of 1 mg/ml NBT in 0.15 M NaCl. After a 30-min incubationat 37°Cin a humidified atmosphere, slides were dried, stained with
Wright-Giemsa, and the percentage of cells containing blue-black for-mazan deposits in at least 200 cells was scored.
Antibody-dependent Cell-mediated Cytotoxicity. A 3-h s'Cr-release
assay was carried out by incubating various numbers of effector cells inround-bottomed microtiter plates with IO4 "Cr-labeled P815y cells
sensitized with a 1:200 dilution of a rabbit anti-PS 15y antiserum.Supernatants were collected and percentage of specific cytotoxicitycalculated as previously reported (47).
Indirect and Two-color Direct Immunofluorescence. For indirect immunofluorescence, cells were sequentially incubated (30 min at ()"(')
with appropriate dilutions of the different monoclonal antibodies (always in the presence of 5% human serum, except for antibody BI47.2)and with FITC-labeled goat F(ab')z anti-mouse Ig. The antibodies used
for two-color immunofluorescence were labeled with FITC and biotin.Biotin-labeled antibodies were detected using phycoerythrin-labeledavidin. The proportion of cells reacting with the different antibodieswas scored on a Cytofluorograf 50H (Ortho Diagnostics, Westwood,MA) connected to a Data General MP/200 microprocessor. To determine the proportion of positive cells, the threshold fluorescence inten
sity used was that at which 99% of the cell population treated withcontrol supernatant from the parental myeloma cells and ITI( untimouse Ig antibodies were negative. Mean fluorescence intensity wasreported on a linear or, when indicated, exponential scale in arbitraryunits, from 1 to 200, which corresponds to the mean fluorescencechannel of the positive cells. Fluorescence intensity allows comparisonof results only within each experiment; in order to allow determinationson a sufficient number of cells, mean fluorescence intensity is reportedonly when the percentage of positive cells is more than 5%.
|'H|d IdK Uptake. Cells were cultured in 96-well flat-bottomed microtiter plates (10* cells/ml, 200 ¿¿I/well)for the indicated periods oftime and pulsed for the last 6 h of culture with 1 /¿Ci/wellof | '11]dl dK
(New England Nuclear); cells were collected on glass-fiber filters withan automated cell harvester (Skatron, Sterling, VA), and the cell-associated radioactivity was assayed by liquid scintillography.
RESULTS
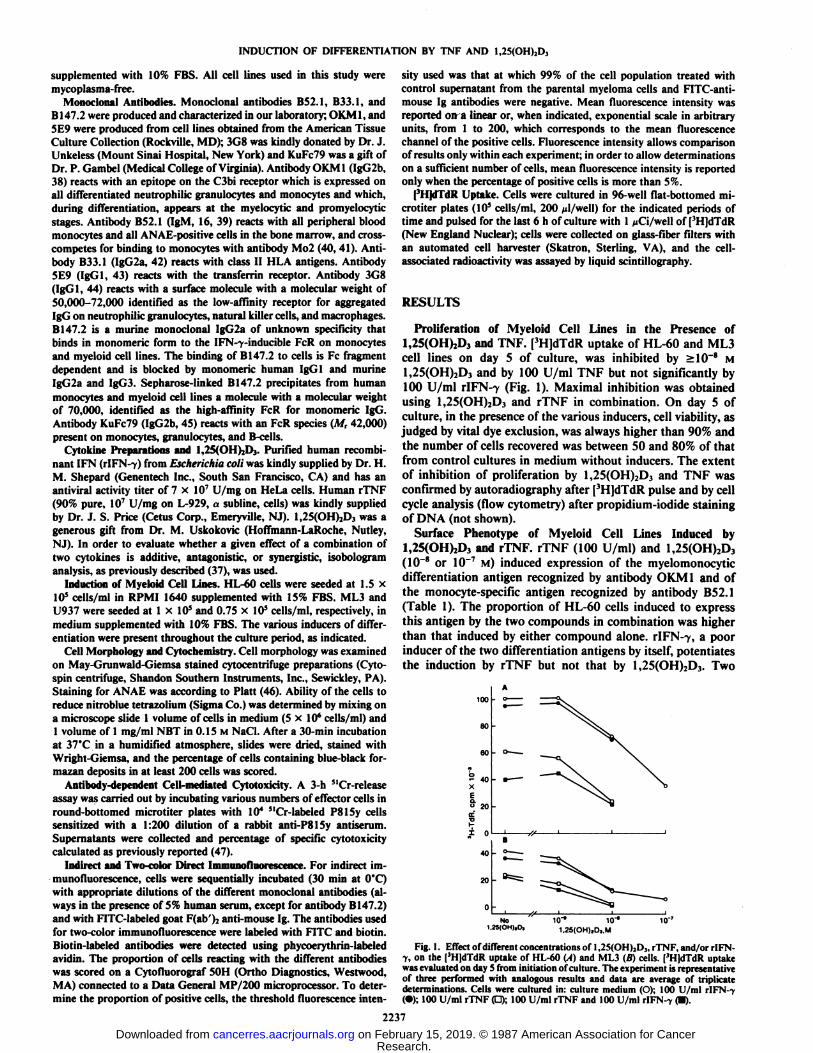
Proliferation of Myeloid Cell Lines in the Presence of1,25(OH)2D3 and TNF. [3H]dTdR uptake of HL-60 and ML3cell lines on day 5 of culture, was inhibited by >10~8 M
1,25(OH)2D3 and by 100 U/ml TNF but not significantly by100 U/ml r!FN--y (Fig. 1). Maximal inhibition was obtained
using 1,25(OH)2D3 and rTNF in combination. On day 5 ofculture, in the presence of the various inducers, cell viability, asjudged by vital dye exclusion, was always higher than 90% andthe number of cells recovered was between 50 and 80% of thatfrom control cultures in medium without inducers. The extentof inhibition of proliferation by 1,25(OH)2D3 and TNF wasconfirmed by autoradiography after [3H]dTdR pulse and by cellcycle analysis (flow cytometry) after propidium-iodide stainingof DNA (not shown).
Surface Phenotype of Myeloid Cell Lines Induced by1,25(OH)2D3 and rTNF. rTNF (100 U/ml) and 1,25(OH)2D3(10~8 or 10~7 M) induced expression of the myelomonocytic
differentiation antigen recognized by antibody OKM1 and ofthe monocyte-specific antigen recognized by antibody B52.1(Table 1). The proportion of HL-60 cells induced to expressthis antigen by the two compounds in combination was higherthan that induced by either compound alone. rIFN-7, a poorinducer of the two differentiation antigens by itself, potentiatesthe induction by rTNF but not that by 1,25(OH)2D3. Two
80
°40
XEQ.o 20
I 0
40
20
Not.2S<OH),0.
io-- io-11.25(OH),Dj.M
Fig. 1. Effect of different concentrations of 1,2S(OH)2D3, rTNF, and/or rlFN-V, on the [3H]dTdR uptake of HL-60 (A) and ML3 (B) cells. [3H|dTdR uptake
was evaluated on day S from initiation of culture. The experiment is representativeof three performed with analogous results and data are average of triplicatedeterminations. Cells were cultured in: culture medium (O); 100 U/ml rIFN--y(•);100 U/ml rTNF (d); 100 U/ml rTNF and 100 U/ml rIFN-7 (•).
2237
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from
INDUCTION OF DIFFERENTIATION BY TNF AND 1,25(OH)2D3
Table I Effect ofl,25(OH)2D,, rTNF, and r¡FN-yon surface antigen expressionby HL-60 cells
HL-60 cells were incubated for S days with the indicated inducers and assayedin indirect immunofluorescence for the expression of the antigens recognized bythe three monoclonal antibodies.
Inducersl,25(OH)jD3(M)0000io-*10-'10-'io-*io-'io-«10-'io-'ir7rIFN-7
(U/ml)010001000100010001000too0rTNF(U/ml)001001000010010000too1000OKM132.5(84)*28.0
(92)60.9(131)69.4(133)27.8(113)31.4(116)75.1
(140)56.2(130)72.5
(103)87.3(107)73.3(171)68.6(151)64.2(158)Antibod/B52.18.6
(80)8.2(108)26.1(135)35.9(149)13.8(128)19.5(137)53.9(168)48.2
(163)43.9(159)44.5(166)66.0(178)57.9(173)79.6(171)5E995.7(187)92.6(175)73.1
(161)68.2(142)86.8(179)87.8
(173)65.1(148)73.3(146)81.1(174)87.3(162)53.8(143)53.9(137)58.2(158)
°Percentage of positive cells.* Numbers in parentheses, average intensity of fluorescence on the positive
cells (linear scale, arbitrary units from 1 to 200).
100 r* 100r
80
0 10-* 10"' 10" 0
1,25(OH),D3.M
10-' IO"1 10-
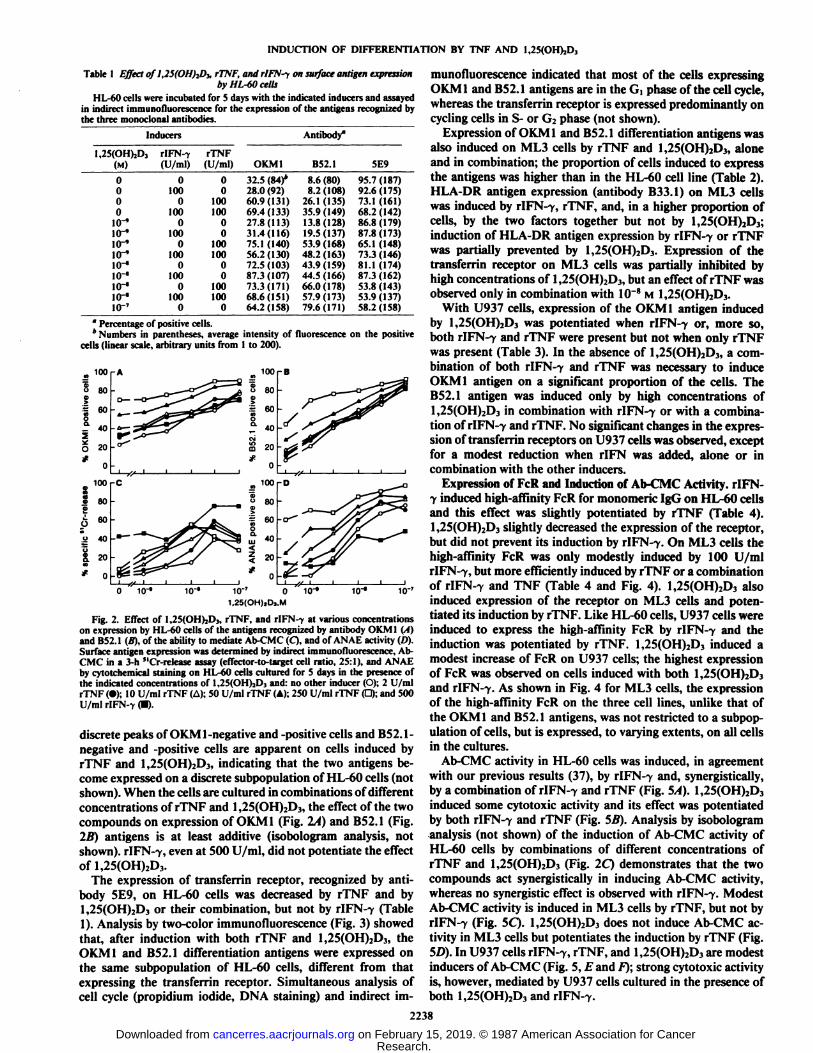
Fig. 2. Effect of l,25(OH)jD3, rTNF, and rlFN-f at various concentrationson expression by HL-60 cells of the antigens recognized by antibody OKM1 (A)and B52.1 (B), of the ability to mediate Ab-CMC (Q, and of ANAE activity (D).Surface antigen expression was determined by indirect immunofluorescence, Ab-CMC in a 3-h "Cr-release assay (effector-to-target cell ratio, 25:1), and ANAEby cytotchemical staining on HL-60 cells cultured for 5 days in the presence ofthe indicated concentrations of l,2?(()l I)..!), and: no other inducer (O); 2 U/mlrTNF (•);10 U/ml rTNF (A); 50 U/ml rTNF (A); 250 U/ml rTNF (D); and 500U/ml rIFN-7 (•).
discrete peaks of OKM1-negative and -positive cells and B52.1-negative and -positive cells are apparent on cells induced byrTNF and 1,25(OH)2D3, indicating that the two antigens become expressed on a discrete subpopulation of HL-60 cells (notshown). When the cells are cultured in combinations of differentconcentrations of rTNF and 1,25(OH)2D3, the effect of the twocompounds on expression of OKM 1 (Fig. 2A) and B52.1 (Fig.2li) antigens is at least additive (isobologram analysis, notshown). rIFN-7, even at 500 U/ml, did not potentiate the effectof 1,25(OH)2D3.
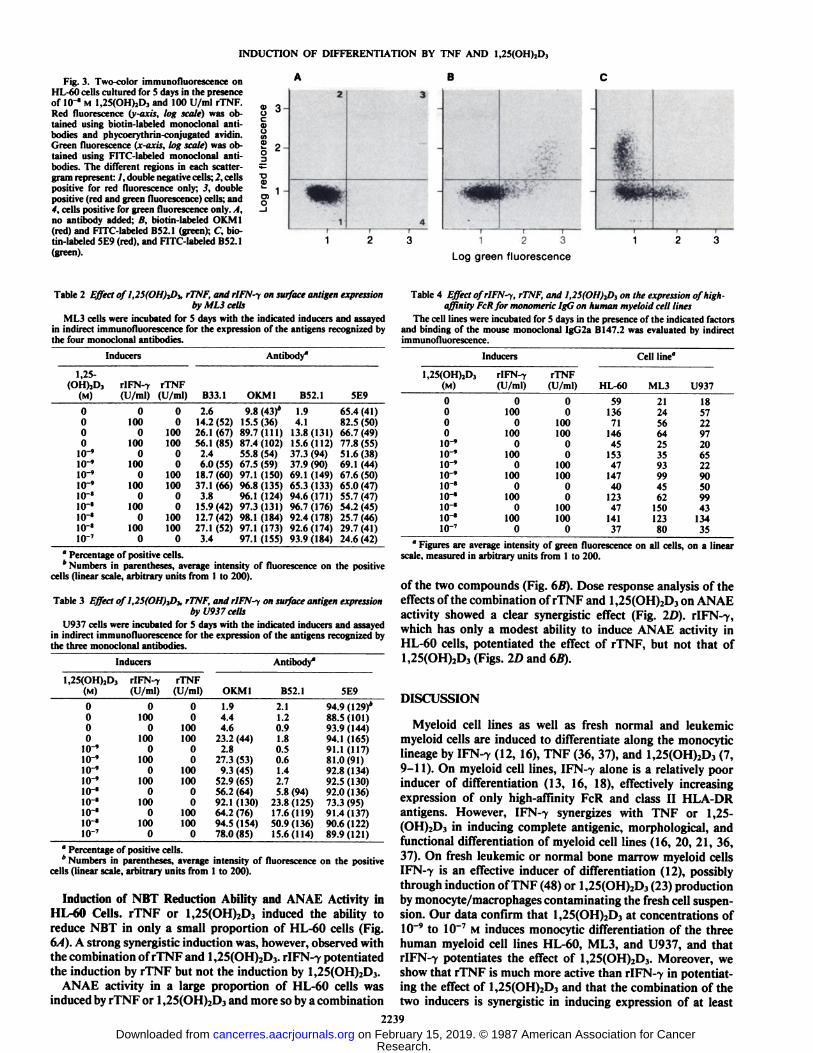
The expression of transferrin receptor, recognized by antibody 5E9, on HL-60 cells was decreased by rTNF and by1,25(OH)2D3 or their combination, but not by rIFN--y (Table1). Analysis by two-color immunofluorescence (Fig. 3) showedthat, after induction with both rTNF and 1,25(OH)2D3, theOKM1 and B52.1 differentiation antigens were expressed onthe same subpopulation of HL-60 cells, different from thatexpressing the transferrin receptor. Simultaneous analysis ofcell cycle (propidium iodide, DNA staining) and indirect im
munofluorescence indicated that most of the cells expressingOKM1 and B52.1 antigens are in the GÃŒphase of the cell cycle,whereas the transferrin receptor is expressed predominantly oncycling cells in S- or G2 phase (not shown).
Expression of OKM 1 and B52.1 differentiation antigens wasalso induced on ML3 cells by rTNF and 1,25(OH)2D3, aloneand in combination; the proportion of cells induced to expressthe antigens was higher than in the HL-60 cell line (Table 2).HLA-DR antigen expression (antibody B33.1) on ML3 cellswas induced by rIFN-7, rTNF, and, in a higher proportion ofcells, by the two factors together but not by 1,25(OH)2D3;induction of HLA-DR antigen expression by r!FN--y or rTNFwas partially prevented by 1,25(OH)2D3. Expression of thetransferrin receptor on ML3 cells was partially inhibited byhigh concentrations of 1,25(OH)2D3, but an effect of rTNF wasobserved only in combination with IO"8 M 1,25(OH)2D3.
With U937 cells, expression of the OKM1 antigen inducedby 1,25(OH)2D3 was potentiated when r!FN--y or, more so,both rIFN-7 and rTNF were present but not when only rTNFwas present (Table 3). In the absence of 1,25(OH)2D3, a combination of both rIFN-7 and rTNF was necessary to induceOKM1 antigen on a significant proportion of the cells. TheB52.1 antigen was induced only by high concentrations of1,25(OH)2D3 in combination with rIFN--y or with a combination of rIFN-7 and rTNF. No significant changes in the expression of transferrin receptors on U937 cells was observed, exceptfor a modest reduction when rIFN was added, alone or incombination with the other inducers.
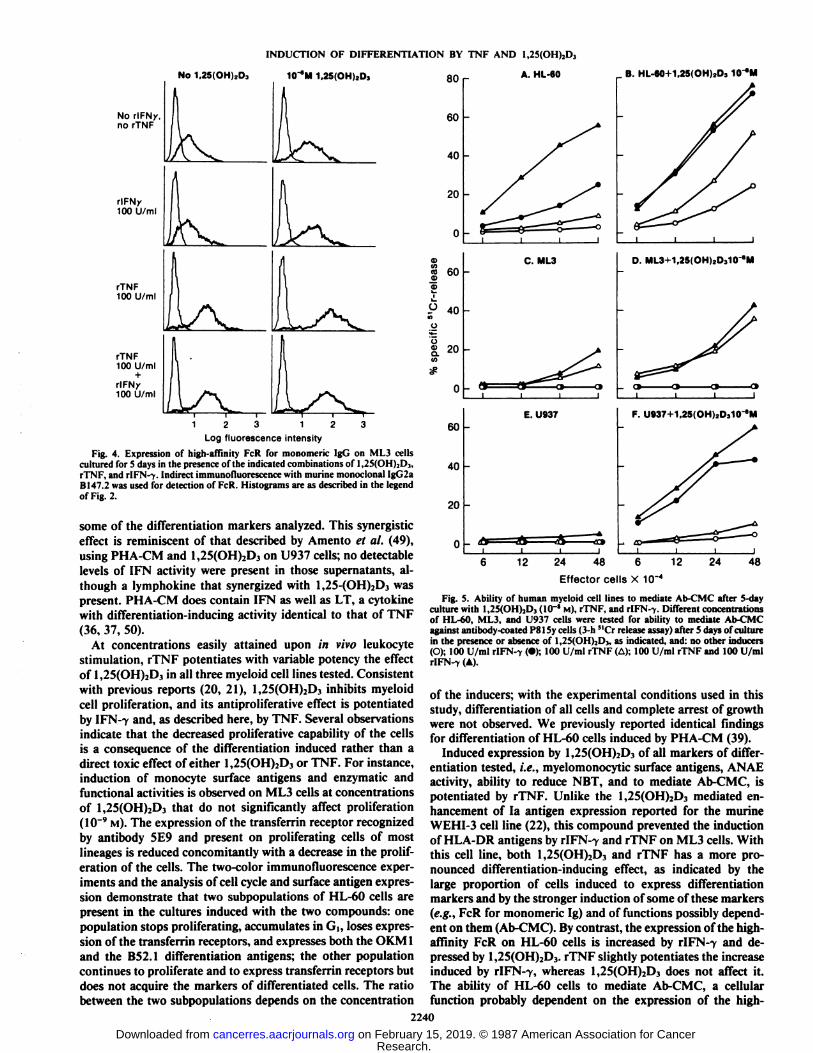
Expression of FcR and Induction of Ab-CMC Activity. rlFN-7 induced high-affinity FcR for monomeric IgG on HL-60 cellsand this effect was slightly potentiated by rTNF (Table 4).1,25(OH)2D3 slightly decreased the expression of the receptor,but did not prevent its induction by r!FN--y. On ML3 cells thehigh-affinity FcR was only modestly induced by 100 U/mlrIFN-7, but more efficiently induced by rTNF or a combinationof rIFN-7 and TNF (Table 4 and Fig. 4). 1,25(OH)2D3 alsoinduced expression of the receptor on ML3 cells and potentiated its induction by rTNF. Like HL-60 cells, U937 cells wereinduced to express the high-affinity FcR by rIFN-7 and theinduction was potentiated by rTNF. 1,25(OH)2D3 induced amodest increase of FcR on U937 cells; the highest expressionof FcR was observed on cells induced with both 1,25(OH)2D3and rIFN-7. As shown in Fig. 4 for ML3 cells, the expressionof the high-affinity FcR on the three cell lines, unlike that ofthe OKM1 and BS2.1 antigens, was not restricted to a subpop-ulation of cells, but is expressed, to varying extents, on all cellsin the cultures.
Ab-CMC activity in HL-60 cells was induced, in agreementwith our previous results (37), by rIFN-7 and, synergistically,by a combination of rIFN-7 and rTNF (Fig. 5A). 1,25(OH)2D3induced some cytotoxic activity and its effect was potentiatedby both rIFN-7 and rTNF (Fig. SB). Analysis by isobologramanalysis (not shown) of the induction of Ab-CMC activity ofHL-60 cells by combinations of different concentrations ofrTNF and 1,25(OH)2D3 (Fig. 2Q demonstrates that the twocompounds act synergistically in inducing Ab-CMC activity,whereas no synergistic effect is observed with rIFN-7. ModestAb-CMC activity is induced in ML3 cells by rTNF, but not byrIFN-7 (Fig. 5Q. 1,25(OH)2D3 does not induce Ab-CMC activity in ML3 cells but potentiates the induction by rTNF (Fig.5D). In U937 cells rIFN-7, rTNF, and 1,25(OH)2D3 are modestinducers of Ab-CMC (Fig. 5, fand F); strong cytotoxic activityis, however, mediated by U937 cells cultured in the presence ofboth 1,25(OH)2D3 and rIFN-7.
2238
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from
INDUCTION OF DIFFERENTIATION BY TNF AND 1,25(OH)2D3
Fig. 3. Two-color immunofluorescence onHL-60 cells cultured for 5 days in the presenceof IO'" M l,25(OH)2Dj and 100 U/ml rTNF.
Red fluorescence (y axis, log scale) was obtained using biotin-labeled monoclonal antibodies and phycoerythrin-conjugated avidin.Green fluorescence (x-axis, log scale) was obtained using FITC-labeled monoclonal antibodies. The different regions in each scatter -gram represent: /, double negative cells; 2, cellspositive for red fluorescence only; 3, doublepositive (red and green fluorescence) cells; and4, cells positive for green fluorescence only.. I,no antibody added; B, biotin-labeled OK Ml(red) and FITC-labeled B52.1 (green); C, biotin-labeled 5E9 (red), and FITC-labeled BS2.1(green).
g3-C5otfìI2'•oi••
1-0)o2dtÊ^i^•^^F134—
r— —r r
1 2 3
^^^^^^r
1 2 3
Log green fluorescence
Table 1 Effect ofl,25(OH)iD), rTNF, and rIFN-y on surface antigen expressionby MLÃŒcells
ML3 cells were incubated for 5 days with the indicated inducers and assayedin indirect immunofluorescence for the expression of the antigens recognized bythe four monoclonal antibodies.
Inducers1,25-(OH)2D,(M)000010-'10-'10-'10-'10-'io-'IO"'io-*io-7rlFN-y(U/ml)0100010001000100010001000rTNF(U/ml)0010010000100100001001000B33.12.614.2
(52)26.1(67)56.1
(85)2.46.0
(55)18.7(60)37.1(66)3.815.9(42)12.7(42)27.1
(52)3.4Antibody4OKMI9.8
(43)*15.5(36)89.7(111)87.4
(102)55.8(54)67.5(59)97.1(ISO)96.8(135)96.1
(124)97.3(131)98.1
(184)97.1(173)97.1
(155)B52.11.94.113.8(131)15.6(112)37.3
(94)37.9(90)69.1(149)65.3(133)94.6(171)96.7(176)92.4(178)92.6(174)93.9(184)5E965.4(41)82.5
(50)66.7(49)77.8(55)51.6(38)69.1
(44)67.6(50)65.0(47)55.7(47)54.2(45)25.7(46)29.7(41)24.6
(42)°Percentage of positive cells.* Numbers in parentheses, average intensity of fluorescence on the positive
cells (linear scale, arbitrary units from I to 200).
Table 3 Effect of 1,25(OH)iD), rTNF, and rIFN-y on surface antigen expressionby U937 cells
U937 cells were incubated for 5 days with the indicated inducers and assayedin indirect immunofluorescence for the expression of the antigens recognized bythe three monoclonal antibodies.
Inducersl,25(OH)2Dj(M)000010-'10-'10-'10-'10-'10-'10-'10-'io-7rIFN-y
(U/ml)0100010001000100010001000rTNF(U/ml)0010010000100100001001000OKMI1.94.44.623.2
(44)2.827.3
(53)9.3(45)52.9(65)56.2(64)92.1(130)64.2(76)94.5(154)78.0
(85)Antibody"B52.12.11.20.91.8O.S0.61.42.75.8
(94)23.8(125)17.6(119)50.9(136)15.6(114)5E994.9(129)*88.5(101)93.9
(144)94.1(165)91.1(117)81.0(91)92.8
(134)92.5(130)92.0(136)73.3
(95)91.4(137)90.6(122)89.9(121)
" Percentage of positive cells.* Numbers in parentheses, average intensity of fluorescence on the positive
cells (linear scale, arbitrary units from 1 to 200).
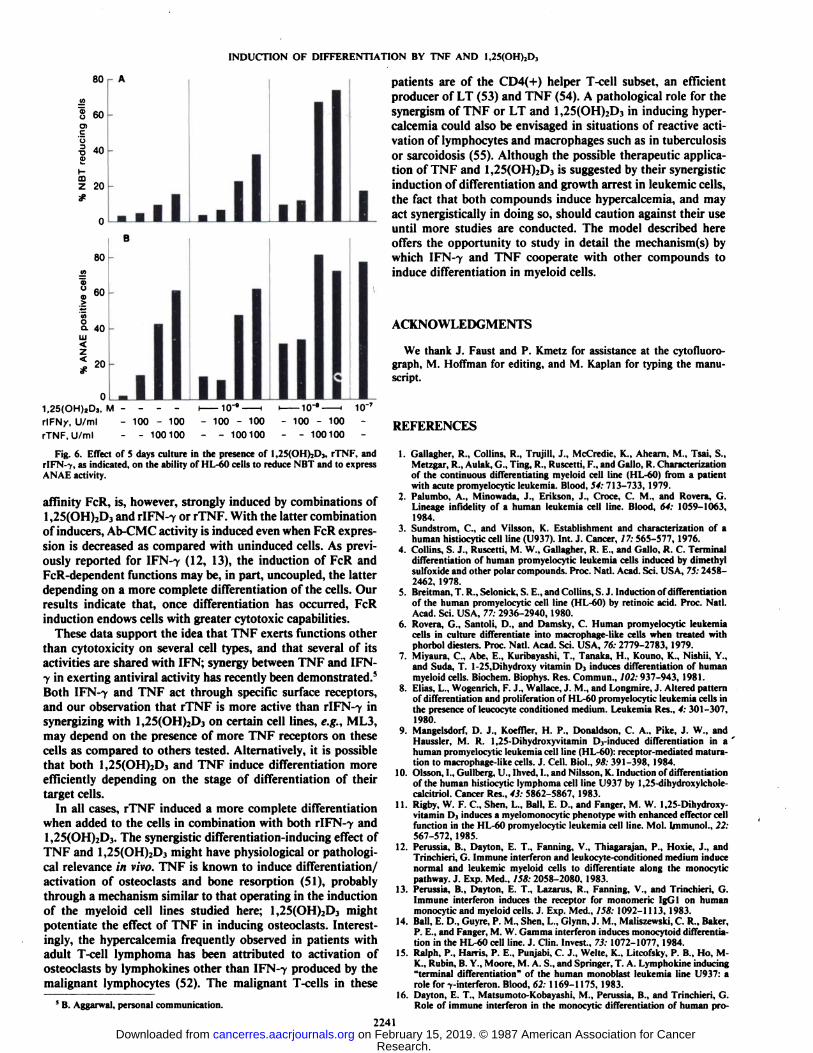
Induction of NBT Reduction Ability and ANAE Activity inHL-60 Cells. rTNF or 1,25(OH)2D3 induced the ability toreduce NBT in only a small proportion of HL-60 cells (Fig.6/Õ).A strong synergistic induction was, however, observed withthe combination of rTNF and 1,25(OH)2D3. rIFN-7 potentiatedthe induction by rTNF but not the induction by 1,25(OH)2D3.
ANAE activity in a large proportion of HL-60 cells wasinduced by rTNF or 1,25(OH)2D3 and more so by a combination
Table 4 Effect ofrlFN-y, rTNF, and 1,2S(OH)2D, on the expression of high-affinity FcR for monomeric IgG on human myeloid cell lines
The cell lines were incubated for 5 days in the presence of the indicated factorsand binding of the mouse monoclonal IgG2a B147.2 was evaluated by indirectimmunofluorescence.
Inducers1,25(OH),D,(M)000010-'10-'10-'10-'10-'io-'10-'10-'io-7rIFN-T
(U/ml)01000too01000100010001000rTNF(U/ml)0010010000100100001001000Cell
line"HL-6059136711464515347147401234714137ML321245664253593994562ISO12380U9371857229720652290SO994313435
' Figures are average intensity of green fluorescence on all cells, on a linear
scale, measured in arbitrary units from 1 to 200.
of the two compounds (Fig. 6B). Dose response analysis of theeffects of the combination of rTNF and 1,25(OH)2D3 on ANAEactivity showed a clear synergistic effect (Fig. 2D). r!FN--y,which has only a modest ability to induce ANAE activity inHL-60 cells, potentiated the effect of rTNF, but not that of1,25(OH)2D3 (Figs. 2D and 65).
DISCUSSION
Myeloid cell lines as well as fresh normal and leukemicmyeloid cells are induced to differentiate along the monocyticlineage by IFN-7 (12, 16), TNF (36, 37), and 1,25(OH)2D3 (7,9-11). On myeloid cell lines, IFN-7 alone is a relatively poorinducer of differentiation (13, 16, 18), effectively increasingexpression of only high-affinity FcR and class II HLA-DRantigens. However, IFN-7 synergizes with TNF or 1,25-(OH)2D3 in inducing complete antigenic, morphological, andfunctional differentiation of myeloid cell lines (16, 20, 21, 36,37). On fresh leukemic or normal bone marrow myeloid cellsIFN-7 is an effective inducer of differentiation (12), possiblythrough induction of TNF (48) or 1,25(OH)2D3 (23) productionby monocyte/macrophages contaminating the fresh cell suspension. Our data confirm that 1,25(OH)2D3 at concentrations of10~9 to 10~7 M induces monocytic differentiation of the three
human myeloid cell lines HL-60, ML3, and U937, and thatrIFN-7 potentiates the effect of 1,25(OH)2D3. Moreover, weshow that rTNF is much more active than rIFN-7 in potentiating the effect of 1,25(OH)2D3 and that the combination of thetwo inducers is synergistic in inducing expression of at least
2239
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from
INDUCTION OF DIFFERENTIATION BY TNF AND 1,25(OH)2D3
No 1.25iOH,..D,
No rIFNy,no rTNF
rIFNy100 U/ml
rTNF100 U/ml
rTNF100 U/ml
+rIFNy100 U/ml
1<r"M 1.25(OH)2D3
Log fluorescence intensityFig. 4. Expression of high-affinity FcR for monomeric IgG on ML3 cells
cultured for 5 days in the presence of the indicated combinations of 1,25(OH)2Dj,rTNF, and rlFN-v Indirect immunofluorescence with murine monoclonal IgG2aBI47.2 was used for detection of FcR. Histograms are as described in the legendof Fig. 2.
some of the differentiation markers analyzed. This synergisticeffect is reminiscent of that described by Amento et al. (49),using PHA-CM and 1,25(OH)2D3 on U937 cells; no detectablelevels of IFN activity were present in those supernatants, although a lymphokine that synergized with 1,25-(OH)2D3 waspresent. PHA-CM does contain IFN as well as LT, a cytokinewith differentiation-inducing activity identical to that of TNF
(36, 37, 50).At concentrations easily attained upon /// vivo leukocyte
stimulation, rTNF potentiates with variable potency the effectof 1,25(OH)2D3 in all three myeloid cell lines tested. Consistentwith previous reports (20, 21), 1,25(OH)2D3 inhibits myeloidcell proliferation, and its antiproliferative effect is potentiatedby IFN-7 and, as described here, by TNF. Several observationsindicate that the decreased proliferative capability of the cellsis a consequence of the differentiation induced rather than adirect toxic effect of either 1,25(OH)2D3 or TNF. For instance,induction of monocyte surface antigens and enzymatic andfunctional activities is observed on ML3 cells at concentrationsof 1,25(OH)2D3 that do not significantly affect proliferation(10~9 M). The expression of the transferrin receptor recognized
by antibody 5E9 and present on proliferating cells of mostlineages is reduced concomitantly with a decrease in the proliferation of the cells. The two-color immunofluorescence experiments and the analysis of cell cycle and surface antigen expression demonstrate that two subpopulations of HL-60 cells arepresent in the cultures induced with the two compounds: onepopulation stops proliferating, accumulates in G,, loses expression of the transferrin receptors, and expresses both the OKM 1and the BS2.1 differentiation antigens; the other populationcontinues to proliferate and to express transferrin receptors butdoes not acquire the markers of differentiated cells. The ratiobetween the two subpopulations depends on the concentration
80 r
60
40
20
g 60
ü 40
üS. 2°
60
40
20
A. HL-60 ,_ B. HL-60+1,25(OH)2D3 KT'M
C. ML3
I I
E. U93T
D. ML3 1,25iOH),D ,10 "M
I I 1 1
F. U937+1,25(OH)2D310
12 24 48 6Effector cells X 10"
12 24 48
Fig. S. Ability of human myeloid cell lines to mediate Ab-CMC after 5-dayculture with 1,25(OH)2D3 (10"' M), rTNF, and rlFN--^. Different concentrationsof HL-60, ML3, and U937 cells were tested for ability to mediate Ab-CMCagainst antibody-coated PSISy cells (3-h "Cr release assay) after 5 days of culturein the presence or absence of 1,25(()HI..I),. as indicated, and: no other inducers(O); 100 U/ml rlFN-? (•);100 U/ml rTNF (A); 100 U/ml rTNF and 100 U/mlrIFN-> (A).
of the inducers; with the experimental conditions used in thisstudy, differentiation of all cells and complete arrest of growthwere not observed. We previously reported identical findingsfor differentiation of HL-60 cells induced by PHA-CM (39).
Induced expression by 1,25(OH)2D3 of all markers of differentiation tested, i.e., myelomonocytic surface antigens, ANAEactivity, ability to reduce NBT, and to mediate Ab-CMC, ispotentiated by rTNF. Unlike the 1,25(OH)2D3 mediated enhancement of la antigen expression reported for the murineWEHI-3 cell line (22), this compound prevented the inductionof HLA-DR antigens by rIFN-7 and rTNF on ML3 cells. Withthis cell line, both 1,25(OH)2D3 and rTNF has a more pronounced differentiation-inducing effect, as indicated by thelarge proportion of cells induced to express differentiationmarkers and by the stronger induction of some of these markers(e.g., FcR for monomeric Ig) and of functions possibly dependent on them (Ab-CMC). By contrast, the expression of the high-affinity FcR on HL-60 cells is increased by rIFN--y and de
pressed by 1,25(OH)2D3. rTNF slightly potentiates the increaseinduced by rIFN-7, whereas 1,25(OH)2D3 does not affect it.The ability of HL-60 cells to mediate Ab-CMC, a cellularfunction probably dependent on the expression of the high-
2240
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from
INDUCTION OF DIFFERENTIATION BY TNF AND 1,25(OH)2D3
80 r A
6001C
00Z 20
80
il I
60
Ut
z
40
20
.11,25(OH)îD3,M - - - - l IO'" 1 i IO'' 1 IO'7
rlFNy, U/ml - 100 - 100 - 100 - 100 - 100 - 100 -rTNF, U/ml - - 100100 - - 100100 - - 100100 -
Fig. 6. Effect of 5 days culture in the presence of 1,25(OH)2D3, rTNF, andrIFN--c, as indicated, on the ability of HL-60 cells to reduce NBT and to expressANAE activity.
affinity FcR, is, however, strongly induced by combinations of1,25(OH)2D3 and rIFN--c or rTNF. With the latter combinationof inducers, Ab-CMC activity is induced even when FcR expression is decreased as compared with uninduced cells. As previously reported for IFN--y (12, 13), the induction of FcR andFcR-dependent functions may be, in part, uncoupled, the latterdepending on a more complete differentiation of the cells. Ourresults indicate that, once differentiation has occurred, FcRinduction endows cells with greater cytotoxic capabilities.
These data support the idea that TNF exerts functions otherthan cytotoxicity on several cell types, and that several of itsactivities are shared with IFN; synergy between TNF and IFN-7 in exerting antiviral activity has recently been demonstrated.5Both IFN-7 and TNF act through specific surface receptors,and our observation that rTNF is more active than r!FN--y insynergizing with U5(()H):I)t on certain cell lines, e.g., ML3,may depend on the presence of more TNF receptors on thesecells as compared to others tested. Alternatively, it is possiblethat both 1,25(OH)2D3 and TNF induce differentiation moreefficiently depending on the stage of differentiation of theirtarget cells.
In all cases, rTNF induced a more complete differentiationwhen added to the cells in combination with both r!FN--y and1,25(OH)2D3. The synergistic differentiation-inducing effect ofTNF and 1,25(OH)2D3 might have physiological or pathological relevance in vivo. TNF is known to induce differentiation/activation of osteoclasts and bone résorption(51), probablythrough a mechanism similar to that operating in the inductionof the myeloid cell lines studied here; 1,25(OH)2D3 mightpotentiate the effect of TNF in inducing osteoclasts. Interestingly, the hypercalcemia frequently observed in patients withadult T cell lymphoma has been attributed to activation ofosteoclasts by lymphokines other than 1I-N--, produced by the
malignant lymphocytes (52). The malignant I cells in these
9 B. Aggarwal, personal communication.
patients are of the CD4(+) helper T-cell subset, an efficientproducer of LT (53) and TNF (54). A pathological role for thesynergism of TNF or LT and 1,25(OH)2D3 in inducing hypercalcemia could also be envisaged in situations of reactive activation of lymphocytes and macrophages such as in tuberculosisor sarcoidosis (55). Although the possible therapeutic application of TNF and 1,25(OH)2D3 is suggested by their synergisticinduction of differentiation and growth arrest in leukemic cells,the fact that both compounds induce hypercalcemia, and mayact synergistically in doing so, should caution against their useuntil more studies are conducted. The model described hereoffers the opportunity to study in detail the mechanism(s) bywhich IFN--y and TNF cooperate with other compounds to
induce differentiation in myeloid cells.
ACKNOWLEDGMENTS
We thank J. Faust and P. Kmetz for assistance at the cytofluoro-graph, M. Hoffman for editing, and M. Kaplan for typing the manuscript.
REFERENCES
1. Gallagher, R., Collins, R., Trujill, J., McCredie, K., Ahearn, M., Tsai, S.,Metzgar, R., Aulak, (... Ting, R., Ruscelli, F., and Gallo, R. Characterizationof the continuous differentiating myeloid cell line (HL-60) from a patientwith acute promyelocytic leukemia. Blood, 54:713-733, 1979.
2. Palumbo, A., Minowada, J., Erikson, J., Croce, C. M., and Rovera, G.Lineage infidelity of a human leukemia cell line. Blood, 64: 1059-1063,1984.
3. Sundstrom, C., and Vilsson, K. Establishment and characterization of ahuman histiocytic cell line (U937). Int. J. Cancer, 17: 565-577, 1976.
4. Collins, S. J., Ruscelli, M. W., Gallagher, R. E., and Gallo, R. C. Terminaldifferentiation of human promyelocytic leukemia cells induced by dimethylsulfoxide and other polar compounds. Proc. Nati. Acad. Sci. USA, 75:2458-2462, 1978.
5. Breitman, T. R., Selonick, S. E., and Collins, S. J. Induction of differentiationof the human promyelocytic cell line (HL-60) by retinole acid. Proc. Nati.Acad. Sci. USA, 77: 2936-2940, 1980.
6. Rovera, G., Santoli, D., and Damsky, C. Human promyelocytic leukemiacells in culture differentiate into macrophage-like cells when treated withphorbol diesters. Proc. Nati. Acad. Sci. USA, 76: 2779-2783, 1979.
7. Miyaura, C., Abe, E., Kuribayashi, T., Tanaka, H., Kouno, K., Nishii, Y.,and Suda, T. l-25,Dihydroxy vitamin l>, induces differentiation of humanmyeloid cells. Biochem. Biophys. Res. Commun., 702:937-943, 1981.
8. Elias, L., Wogenrich, F. J., Wallace, J. M., and Longmire, J. Altered patternof differentiation and proliferation of HL-60 promyelocytic leukemia cells inthe presence of leucocyte conditioned medium. Leukemia Res., 4: 301-307,1980.
9. Mangelsdorf, D. J., Koeffler, H. P., Donaldson, C. A., Pike, J. W., andHaussler, M. R. l,2S-Dihydroxyvitamin D3-induced differentiation in a '
human promyelocytic leukemia cell line (HL-60): receptor-mediated maturation to macrophage-like cells. J. Cell. Biol., 98: 391-398, 1984.
10. Olsson, I., Gullberg, I '.. lined. I., and Nilsson, K. Induction of differentiation
of the human histiocytic lymphoma cell line U937 by 1,25-dihydroxylchole-calcitriol. Cancer Res., 43: 5862-5867, 1983.
11. Rigby, W. F. C., Shen, L., Ball, E. p., and Fanger, M. W. 1,25-Dihydroxy-vitamin I>, induces a myelomonocytic phenotype with enhanced effector cellfunction in the HL-60 promyelocytic leukemia cell line. Mol. Immun«!..22:567-572, 1985.
12. Perussia, B., Dayton, E. T., Fanning, V., Thiagarajan, P., Hoxie, J., andTrinchieri, G. Immune Interferon and leukocyte-conditioned medium inducenormal and leukemic myeloid cells to differentiate along the monocyticpathway. J. Exp. Med., 158: 2058-2080. 1983.
13. Perussia, B., Dayton, E. T., Lazarus, R., Fanning, V., and Trinchieri, G.Immune interferon induces the receptor for monomeric IgGl on humanmonocytic and myeloid cells. J. Exp. Med., 158: 1092-1113, 1983.
14. Ball, E. I).. Guyre, P. M., Shen, L., Glynn, J. M., Maliszewski, C. R., Baker,P. E., and Fanger, M. W. Gamma interferon induces monocytoid differentiation in the HL-60 cell line. J. Clin. Invest., 73: 1072-1077,1984.
15. Ralph, P., Harris, P. E., Punjabi, C. J., Weite, K., Litcofsky, P. B., Ho, M-K., Rubin, B. Y., Moore, M. A. S., and Springer, T. A. Lymphokine inducing"terminal differentiation" of the human monoblast leukemia line U937: arole for -x-interferon. Blood, 62: 1169-1175, 1983.
16. Dayton, E. T., Matsumoto-Kobayashi, M., Perussia, B., and Trinchieri, G.Role of immune interferon in the monocytic differentiation of human pro-
2241
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from

INDUCTION OF DIFFERENTIATION BY TNF AND 1,25(OH)2D3
myelocytic cell lines induced by leukocyte conditioned medium. Blood, 66:583-594, 1985.
17. Harris, P. E., Ralph, P., Gabrilove, J., Weite, K., Kannali, R., and Moore,M. A. S. Distinct differentiation inducing activities of Y-interferon andcytokine factors acting on the human promyelocytic leukemia cell line HI60. Cancer Res., 45: 3090-3095, 1985.
18. Guyre, P. M . Morganelli, P. M., and Miller, R. Recombinant immuneinterferon increases immunoglobulin G Fc receptors on cultured humanmononuclear phagocytes. J. Clin. Invest., 72: 393-397, 1983.
19. Basham, T. Y., and Merigan, T. C. Recombinant interferone increasesHLA-DR synthesis and expression. J. Immunol., 130:1492-1497, 1983.
20. Gullberg, I . Nilsson, E., Einhorn, S., and Olsson, I. Combinations ofinicrlcron -) and retinole acid or la,25-dihydroxycholecalciferol induce differentiation of the human monoblast leukemia cell line U937. Exp. Hematol.,73:675-679,1985.
21. Weinberg, J. B., Misukonis, M. A., Hobbs, M. II.. and Borowitz, M. J.Cooperative effects of -Hnterferon and 1,25-dihydroxyvitamin 1><in inducingdifferentiation of human promyelocytic leukemia (HL-60) cells. Exp. Hematol., 14:138-142, 1986.
22. Morel, P. A., Manolagas, S. C., Prowedini, D. M., Wegmann, D. R., andChiller, J. M. Interferon-y-induced la expression in WEHI-3 cells is enhanced by the presence of 1,25-dihydroxyvitamin IX J. Immunol., 136:2181-2186, 1986.
23. Koefller, H. P., Reiche!, H., Bishop, J. E., and Norman, A. W. -y-Interferonstimulates production of 1,25-dihydroxyvitamin I) <by normal human macrophages. Biochem. Biophys. Res. Commun., ¡27:596-603, 1985.
24. Carswell, E. A., Old, L. J., Kassel, R. L., Green, S., Fiore, N., and Williamson, B. An endotoxin-induced serum factor that causes necrosis of tumors.Proc. Nati. Acad. Sci. USA, 72: 3666-3670, 1975.
25. Ruddle. N. H., and Waksman, B. W. Cytotoxicity mediated by soluble antigenand lymphocytes in delayed hypersensitivity. HI. Analysis of mechanism. J.Exp. Med., 128: 1267-1279, 1968.
26. Williams, T. W., and Granger, G. A. Lymphocyte in vitro cytotoxicity:lymphotoxin of several mammalian species. Nature (Lond.), 219: 1076-1077, 1968.
27. Williams, T. W., and Bellanti, J. A. In vitro synergism between interferonsand human lymphotoxin: enhancement of lymphotoxin-induced target cellkilling. J. Immunol., ¡30:518-520, 1983.
28. Stone-Wolff, D. S., Yip, Y. K., Kelker, H. C., Le, J., Henriksen-Destefano,D., Rubin, B. Y., Rinderknecht, E., Aggarwal, B. B., and Vilcek, J. Interrelationships of human interferon-gamma with lymphotoxin and monocytecytotoxin. J. Exp. Med., 759:828-843, 1984.
29. Aggarwal, B. B., Mollai. B., and Harkins, R. N. Human lymphotoxinproduction by a lymphoblastoid cell line; purification and initial characterization. J. Biol. Chem., 259: 686-691, 1984.
30. Aggarwal, B., Henzel, W., Mollai. B., Kohr, W., and Harkins, R. N. Primarystructure of human lymphotoxin derived from 1788 lymphoblastoid cell line.J. Biol. Chem., 260: 2334-2338, 1985.
31. Gray, P. W., Aggarwal, B. B., Benton, C. V., Bringman, T. S., Henzel, W.J., Jarre«, J. A., Leung, D. W., Moffat, B., Ng, P., Svedersky, L. P.,Palladino, M. A., and Nedwin, G. E. Cloning and expression of cDNA forhuman lymphotoxin, a lymphokine with tumor necrosis activity. Nature(Lond.), 312: 721-724, 1984.
32. Pennica, D., Nedwin, G. E., Hayflick, J. S., Seeburg, P. H., Derynck, R.,Palladino, M. A., Kohr, W. J., Aggarwal, B. B., and Goeddel, D. V. Humantumor necrosis factor: precursor structure, expression and homology tolymphotoxin. Nature (Lond.), 312: 724-729, 1984.
33. Wang, A. M., Creasy, A. A., Ladner, M. B., Lin, L. S., Strickler, J., VanArsdell, J. N., Yamamoto, R., and Mark, D. F. Molecular cloning of thecomplementary DNA for human tumor necrosis factor. Science (Wash.),22«:149-154, 1985.
34. Sugarman, B. J., Aggarwal, B. B., Hass, P. E., Figari, I. S.. Palladino, M.A., and Shepard, H. M. Recombinant human tumor necrosis factor-«:effectson proliferation of normal and transformed cells in vitro. Science (Wash.),250:943-945, 1985.
35. Vilcek, J., Palombella, V. J., Henriksen-Destefano, D., Swenson, C., Fein-man, R., Ilirai. M., and Tsujmoto, M. Fibroblast growth enhancing activityof tumor necrosis factor and its relationship to other polypeptide growthfactors. J. Exp. Med., 163: 632-643, 1986.
36. Trinchieri, G., Kobayashi, M., Murphy, M., and Perussia, B. Immune interferon and cytotoxins: regulatory effects on myeloid cells. Lymphokines, inpress, 1987.
37. Trinchieri, G., Kobayashi, M., Rosen, M., London, R., Murphy, M., andPerussia, B. Tumor necrosis factor and lymphotoxin induce differentiationof human myeloid cells in synergy with immune interferon. J. Exp. Med.,164: 1206-1225, 1986.
38. Breard, J., Reinherz, E. L., Kung, P. C, Goldstein, G., and Schlossman, S.F. A monoclonal antibody reactive with human peripheral blood monocytes.J. Immunol., 124:1943-1948, 1980.
39. Dayton, E. T., Perussia, B., and Trinchieri, G. Correlation between differentiation, expression of monocyte-specific antigens, and cytotoxic functionsin human promyelocytic cell lines treated with leukocyte-conditioned medium. J. Immunol., 130:1120-1128, 1983.
40. Todd, R. F., Ill, (,rillin. J. D., Ritz, J., Nadler, L. M., Abrams, T., andSchlossman, S. F. Expression of normal monocyte-macrophage differentiation antigens on HL-60 promyelocytes undergoing differentiation inducedby leukocyte conditioned medium or plu>rhol diester. Leukemia Res., S: 491-495, 1981.
41. Todd, R. F., Ill, Nadler, L. M., and Schlossman, S. F. Antigens on humanmonocytes identified by monoclonal antibodies. J. Immunol., 726: 1435-1442, 1981.
42. Perussia, B., Acuto, O., Terhorst, C., Faust, S., Lazarus, R., Fanning, V.,and Trinchieri, G. Human natural killer cells analyzed by B73.1, a monoclonal antibody blocking le-receptor functions. H. Studies of B73.1 antibody-antigen interaction on the lymphocyte membrane. J. Immunol., 730: 2142-2148, 1983.
43. Haynes, B. F., Hemler, M. E., Mann, D. L., Eisenbarth, G. S., Shelhamer,J., Mostowski, H. S., Thomas, C. A., Strominger, J. L., and Fauci, A. S.Characterization of a monoclonal antibody (4F2) that binds to humanmonocytes and to a subset of activated lymphocytes. J. Immunol., 726:1409-1414, 1981.
44. Fleit, H. B., Wright, S. D., and Unkeless, J. C. Human neutrophil Fc receptordistribution and structure. Proc. Nati. Acad. Sci. USA, 79:3275-3279,1982.
45. Vaughn, M., lavier. M., and Mohanakamur, T. Characterization of humanIgG Fc receptors. J. Immunol., 135:4059-4065, 1985.
46. Platt, W. R. Color Atlas and Textbook of Hematology, p. 518. Philadelphia:J. B. Lippincott Co., 1979.
47. Trinchieri, G., de Marchi, M., Mayr, W., Savi, M., and Ceppellini, R.Lymphocyte antibody lymphocytolytic interaction (LALI) with special emphasis on HLA. Transplant. Proc., 5: 1631-1646, 1973.
48. Nedwin, G. E., Svedersky, L. P., Bringman, T. S., Palladino, M. A., andGoeddel, D. V. Effect of interleukin 2, interferon and mitogens on theproduction of tumor necrosis factors a and .1.J. Immunol., 735: 2492-2497,1985.
49. Amento, E. P., Bhalia, A. K., Kurnick, J. T., Kadrin, R. L., Clemens, J. L.,Holick, S. A., Holick, M. F., and Krane, S. M. 1,25-Dihydroxyvitamin D,induces maturation of the human monocyte cell line U937, and, in associationwith a factor from human T lymphocytes, augments production of themonokine, mononuclear cell factor. J. Clin. Invest., 73: 731-739,1984.
50. Murphy, M., Loudon, R., Kobayashi, M., and Trinchieri, G. Gamma interferon and lymphotoxin, released by activated T cells, synergize to inhibitgranulocyte-monocyte colony formation. J. Exp. Med., 76* 263-279, 1986.
51. Bertolini, D. R., Nedwin, N. G.. Bringman, T., Smith, D., and Mundy, G.R. Stimulation of bone résorptionand inhibition of bone formation in vitroby human tumor necrosis factors. Nature (Lond.), 379: 516-518, 1986.
52. Dodd, R. C., Newman, S. L., Bunn, P. A., Winkler, C. F., Cohen, M. S., andGray, T. K. I ymphokinc induced monocytic differentiation as a possiblemechanism for hypercalcemia associated with I cell lymphoma. Cancer Res.,45: 2501-2506, 1985.
53. Conta, B. S., Powell, M. B., and Ruddle, N. H. Activation of Lyt-P and Lyt-2" T cell cloned lines: stimulation of proliferation, lymphokine production,and self destruction. J. Immunol., 134: 2185-2190, 1985.
54. Cuturi, M. C., Murphy, M., Costa-Giorni, M. P., Weinmann, R., Perussia,B., and Trindrieri, G. Independent regulation of tumor necrosis factor andlymphotoxin production by human peripheral blood lymphocytes. J. Exp.Med., in press, 1987.
55. Gkonos, P. I., Loudon, R., and Hendler, E. D. Hypercalcemia and elevated1.25-dili) droxy vitamin D levels in a patient with end stage renal disease andactive tuberculosis. N. Engl. J. Med., 377: 1683-1685, 1984.
2242
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from

1987;47:2236-2242. Cancer Res Giorgio Trinchieri, Mark Rosen and Bice Perussia 3,25-Dihydroxyvitamin D
αTumor Necrosis Factor in Cooperation with 1Induction of Differentiation of Human Myeloid Cell Lines by
Updated version
http://cancerres.aacrjournals.org/content/47/9/2236
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/47/9/2236To request permission to re-use all or part of this article, use this link
Research. on February 15, 2019. © 1987 American Association for Cancercancerres.aacrjournals.org Downloaded from





























