Embed Size (px)
Citation preview

Vol. 182, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS February 14, 1992 Pages 1277-l 281
Hideo Yemasaki, Shigeki Okayama*, Masatoshi Sbibata and Mitsuo Nishimura
Department of Biology, Faculty of Science, Kyushu University, BYigashi-ku, Fukuoka 812, Japan
*Biological Laboratory, College of General Education, Kyushu University, Ropponmatsu, Fukuoka 810, Japan
Received December 27, 1991
SUMMARY: The light-induced II+ release from thylakoids, which can be observed under completely uncoupled conditions, was inhibited by the SH reagent IIF ethylmaleimide (NEM) and its analogs, while the conventional H+ uptake and electron transfer were not affected. The half-inhibiting concentration of NEM for the H+ release was 10 mM and 4 mM in thylakoids in the presence of nigericin and in CFi-depleted thylakoids, respectively. The inhibitory effect increased with the increase in hydrophobicity of the NEM analogs: Z+ methylmaleimide < llFethylma.leimide < 1IFphenylmaleimide. It is suggested that SH groups in hydrophobic interior within the membrane are essential to the release of protons. 0 1992 Academic Press, Inc.
Illumination of thylakoid suspension causes H+ uptake from the
surrounding medium, a process corresponding to the vectorial H+ translocation
into the thylakoid vesicles. Removal of the catalytic moiety of the H+-ATPase
(CF1) from the membrane reverses the direction of the pH change under
illumination, namely, protons are released from thylakoids [l]. The hght-
induced H+ release is a unique phenomenon not observed in mitochondria and
bacteria [2]. The maximal amount of the H+ released is comparable to and
sometimes larger than that of the steady-state H+ uptake which can be
coupled with ATP synthesis [l]. Little is known concerning the mechanism by
which the light-response of pH change is reversed. Recently we found that
H+-conducting uncouplers and T&on-X100 also elicit the light-induced H'
release and concluded that the H+ release is a novel scalar reaction not due to
the splitting of water by PS II [2,3].
The H+ release is suppressed by inhibitors of the electron transfer [3].
However, there are no reports on specific inhibitors of the H+ release. We
report here that mthylmaleimide (NEM) and its analogs can inhibit the light-
induced H+ release from uncoupled thylakoid membranes without affecting other
photochemical activities.
OOD6-291X/92 $1.50
1277 Copyright 0 1992 by Academic Press, Inc.
All rights of reproduction in any form reserved.
Vol. 182, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0 1
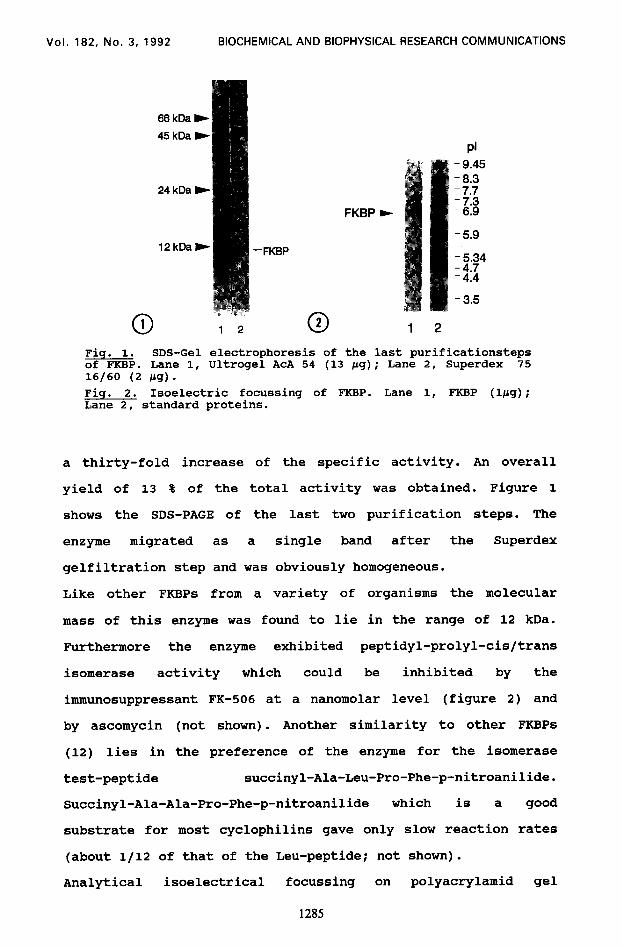
Fig. 1. SDS-Gel of FKBP. Lane 1, 16/60 (2 ccg) -
FKBP b
f3 w 12
PI - 9.45 - 8.3 -7.7 I g.8
electrophoresis of the last purificationsteps Ultrogel AcA 54 (13 pg); Lane 2, Superdex 75
Fig. 2. Isoelectric focussing of FKBP. Lane 1, FKBP (Qbg); Lane 2, standard proteins.
a thirty-fold increase of the specific activity. An overall
yield of 13 % of the total activity was obtained. Figure 1
shows the SDS-PAGE of the last two purification steps. The
enzyme migrated as a single band after the Superdex
gelfiltration step and was obviously homogeneous.
Like other FKBPs from a variety of organisms the molecular
mass of this enzyme was found to lie in the range of 12 kDa.
Furthermore the enzyme exhibited peptidyl-prolyl-cis/trans
isomerase activity which could be inhibited by the
immunosuppressant FK-506 at a nanomolar level (figure 2) and
by ascomycin (not shown). Another similarity to other FKBPs
(12) lies in the preference of the enzyme for the isomerase
test-peptide succinyl-Ala-Leu-Pro-Phe-p-nitroanilide.
Succinyl-Ala-Ala-Pro-Phe-p-nitroanilide which is a good
substrate for most cyclophilins gave only slow reaction rates
(about l/12 of that of the Leu-peptide; not shown).
Analytical isoelectrical focussing on polyacrylamid gel
1285
Vol. 182, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0 10 2OmM
NEM
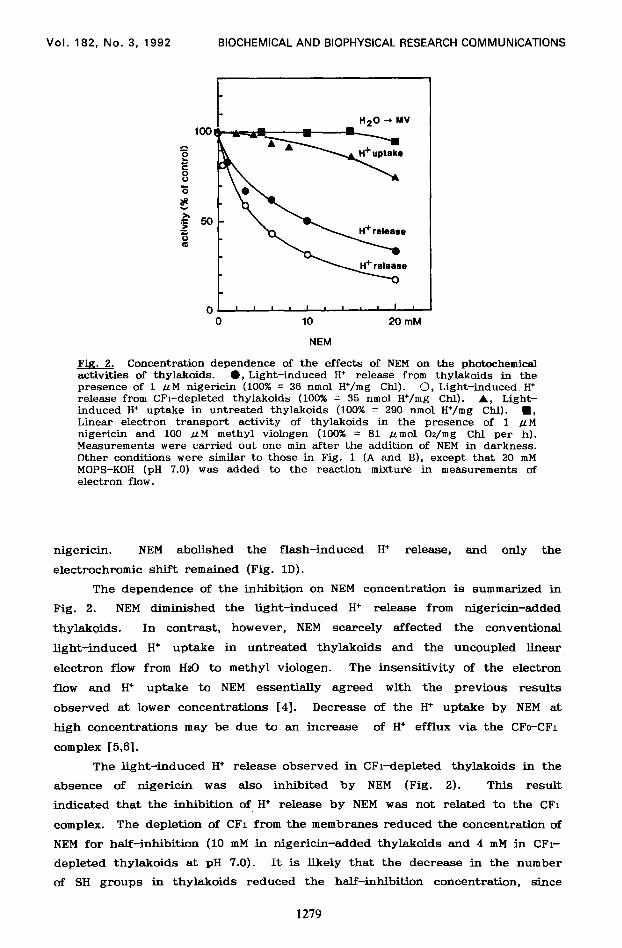
Fig. 2. Concentration dependence of the effects of NEM on the photochemical activities of thylakoids. l , Light-induced H* release from thylakoids in the presence of 1 ILM nigericin (100% = 36 nmol H+/mg Chl). 0, Light-induced H+ release from CFi-depleted thylakoids (100% = 35 nmol H*/mg Chl). A, Light- induced H+ uptake in untreated thylakoids (100% = 290 nmol H+/mg Chl). n , Linear electron transport activity of thylakoids in the presence of 1 .uM nigericin and 100 LLM methyl viologen (100% = 81 ~mol Oz/mg Chl per h). Measurements were carried out one min after the addition of NEM in darkness. Other conditions were similar to those in Fig. 1 (A and B), except that 20 mM MOPS-KOH (pH 7.0) was added to the reaction mixture in measurements of electron flow.
nigericin. NEM abolished the flash-induced H+ release, and only the
electrochromic shift remained (Fig. 1D).
The dependence of the inhibition on NEM concentration is summarized in
Fig. 2. NEM diminished the light-induced H+ release from nigericin-added
thylakoids. In contrast, however, NEM scarcely affected the conventional
light-induced H+ uptake in untreated thylakoids and the uncoupled linear
electron flow from HzO to methyl viologen. The insensitivity of the electron
flow and H+ uptake to NEM essentially agreed with the previous results
observed at lower concentrations [4]. Decrease of the H+ uptake by NEM at
high concentrations may be due to an increase of H+ efflux via the CFc-CF1
complex [5,6].
The light-induced H+ release observed in CFl-depleted thylakoids in the
absence of nigericin was also inhibited by NEM (Fig. 2). This result
indicated that the inhibition of H+ release by NEM was not related to the CFi
complex. The depletion of CFi from the membranes reduced the concentration of
NEM for half-inhibition (10 mM in nigericin-added thylakoids and 4 mM in CFl-
depleted thylakoids at pH 7.0). It is likely that the decrease in the number
of SH groups in thylakoids reduced the half-inhibition concentration, since
1279

Vol. 182, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Table 1. Inhibitory effects of maleimides on the light-induced II+ release
Reagent H+ release Inhibition (nmol H+/mg Chl) (%I
No addition 43.1 f 0.7 -
N-MM 33.3 + 0.6 23 NEM 31.3 + 0.1 27 NPM 20.9 + 0.7 52
Each reagent was added to the reaction medium at 5 mM. The H+ release was measured in the presence of 1 .uM nigericin. Other conditions were similar to those in Fig. 1 (A and B). Abbreviations used: NEM, hkthylmaleimide; NMM, N- methylmaleimide; NPM, Wphenylmaleimide.
the treatment for depletion of CFi decreases the total amount of SH groups in
the membrane by removal of peripheral proteins including the CFi complex.
The inhibitory effect depended on the hydrophobicity of the reagents
(Table 1). NMM, NEM and NPM are N-substituted maleimides, whose side chains
are methyl, ethyl and phenyl group, respectively. The inhibition increased
with the hydrophobicity of the side chain, i.e., methyl < ethyl < phenyl. The
hydrophilic thiol reagent iodoacetic acid did not inhibit the H+ release (less
than 1 % inhibition at 5 mM, not shown). The dependence of the inhibition on
reagent hydrophobicity may be explained by the difference in their
solubilities in the thylakoid membranes. The requirement for somewhat high
NEM concentrations to inhibit the H+ release can be attributed to the location
of reactive thiol groups; those thiols may be located in the hydrophobic
interior within the thylakoid membrane.
As described above, NEM inhibits the light-induced H+ release without
affecting the linear electron flow and conventional H+ uptake. The difference
in sensitivities to NEM (Fig. 2) supports our earlier suggestion that the H+
release is not a reversed H+ translocation nor a part of reactions associated
with H+ uptake [21. The H+ releasing mechanism is still obscure. Data
presented here suggest the requirement of thiol groups within the membrane for
the H+ release.
Several enzymes involved in photosynthesis are regulated by the redox
state of the enzyme thiol groups; these include the CFo-CFi complex [7],
fructose-1,6-bisphosphatase [7] and glucose++phosphate dehydrogenase [S].
Those enzymes have a Cys-X-X-X-X-X-Cys motif [g-11]. Stromal thioredoxin
regulates such enzymic activities by reducing the disulfide in vivo [7]. But
the H+ release reaction does not involve such enzymes because it can be
observed in the absence of stroma and CFi (Fig. 2).
Thylakoid membranes have the xanthophyll cycle which is believed to
dissipate excess energy of light. Violaxanthin de-epoxidase, a component of
the xanthophyll cycle, is inhibited by the SH reagent dithiothreitol and is
thought to contain a disulfide group that is necessary for activity [12,13].
1280

Vol. 182, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
The de-epoxidase may be a candidate for NEM-sensitive enzymes involved in the
H* release. Further investigation are in progress to identify the H+ releasing
components.
ACKNOWLEDGMENTS
We thank Dr. K. Shimazaki of Kyushu University for helpful discussions. This work was partly supported by a Grant-in-Aid for the Promotion of Science for Japanese Junior Scientists (to H. Y.) from the Ministry of Education, Science and Culture of Japan.
REFERENCES
1. Kamienietzky, A. and Nelson, N. (1975) Plant Physiol. 55, 282-287. 2. Yamasaki, H., Furuya, S., Kawamura, A., Ito, A., Okayama, S. and
Nishimura, M. (1991) Plant Cell Physiol. 32, 925-934. 3. Yamasaki, H. and Nishimura, M. (1988) Plant Cell Physiol. 29, 1061-1064. 4. McCarty, R. E., Pittman, P. R. and Tsuchiya, Y. (1972) J. Biol. Chem.
247, 3048-3051. 5. Wagner, R. and Junge, W. (1977) Biochim. Biophys. Acta 462, 259-272. 6. Weiss, M. A. and McCarty, R. E. (1977) J. Biol. Chem. 252, 8007-8012. 7. Noctor, G. and MiBs, J. D. (1988) Bioch.im. Biophys. Acta 935, 53-60. 8. Brennan, T. and Anderson, L. E. (1980) Plant Physiol. 66, 815-817. 9. Inohara, N, Iwamoto, A., Moriyama, Y., Shimomura, S., Maeda, M. and
Futai, M. (1991) J. Mol. Chem. 266, 7333-7338. 10. Hartman, H., Syvanen, M. and Buchanan, B. B. (1990) Mol. Biol. Evol. 7,
247-254. 11. Miki, J., Maeda, M., Mukohata, Y. and Futai, M. (1988) FEBS Lett. 232,
221-226. 12. Yamamoto, H. Y. (1979) Pure Appl. Chem. 51, 639-648. 13. Pfundel, E. and Strasser, R. J. (1988) Photosynth. Res. 15, 67-73.
1281





























