Embed Size (px)
Citation preview

[CANCER RESEARCH 54, 1035-11141, February 15. 19941
Inhibitory Effect of Bombesin Receptor Antagonist RC-3095 on the Growth of
Human Pancreatic Cancer Cells in Vivo and in Vitro ~
Y u n f e n g Qin , T i b o r Ert l , R e n - Z h i Cai , G a b o r H a l m o s , a n d A n d r e w V. S e h a l l y 2
Endocrine, Polypeptide and Cancer Institute, Veterans AfJhirs Medical Center, and Section of Experimental Medicine, Department of Medicine, Tulane University School of Medicine, New Orleans, Louisiana 70146 [Y. Q., T. E., R-Z. C., G. H., A. V. S.]
A B S T R A C T
In this study, we investigated the effect of bombesin/GRP antagonist RC-3095 on the growth of CFPAC-1 human pancreatic cancer cells trans- planted to nude mice or cultured in vitro. Nude mice bearing xenografts of the CFPAC-1 cell line received s.c. injections of RC-3095 (10/tg twice a day) or the vehicle (control) for 25 days. Chronic administration of RC- 3095 inhibited the growth of CFPAC-1 tumors in nude mice as shown by a significant decrease in tumor volume throughout the period of treat- ment. Tumor volume doubling time was prolonged by RC-3095 treatment from 7.2 days to 10 days, and the tumor growth rate was decreased by 49%. In mice treated with RC-3095, the tumor growth delay time was 5.8 days. ~I~reatment with RC-3095 decreased the final tumor weight by 37% and reduced DNA and protein contents in tumor tissues by 44 and 39.9%, respectively, compared to the controls. In cultures of the CFPAC-I cell line, the addition of bombesin(1-14) (1 p~a-0.1 pM) to the medium induced a dose-dependent increase in cell number. RC-3095 at 1 nM concentration effectively inhibited the bombesin-stimulated growth of CFPAC-I cells in cultures. In the presence of 1 btM RC-3095 in the culture medium, the bombesin-induced growth of CFPAC-1 cells was totally suppressed. Bom- besin was also shown to stimulate the DNA synthesis in CFPAC-1 cells in vitro as based on [3H]thymidine incorporation assay. When the cells were cultured in the presence of 1-100 nM bombesin, the uptake of [3H]thymi- dine by the cells was increased by 89-131%. RC-3095 inhibited both the basal and bombesin-stimulated DNA synthesis of CFPAC-1 cells. Addition of RC-3095 (10-100 nM) alone to the cultures caused a 39-40% decrease in the [3H]thymidine incorporation by the cells. Concomitant addition of RC-3095 (1 p~) and bombesin (1-100 nM) to the cultures induced a sig- nificant reduction in the uptake of [aH]thymidine by the cells compared to the values obtained with bombesin alone. Receptor binding assays showed the presence of two classes of specific binding sites for bombesin on CFPAC-1 cells, one with high affinity (Ka = 4.25 _+ 0.77 n~a) and low capacity (B,,ox = 0.268 +_ 0.052 pmol/106 cells) and the other with low affinity (K,t -" 321.70 + 68.46 nM) and high capacity (B .... - 3.991 +_ 0.374 pmoi/106 cells). Antagonist RC-3095 inhibited the binding of ~2SI-Tyr 4- bombesin to CFPAC-1 cell membranes in a dose-dependent manner. These observations suggest that bombesin acts as a growth factor and stimulates proliferation of CFPAC-1 human pancreatic cancer through specific re- ceptors for bombesin/GRP present on the cells. RC-3095 appears to inhibit the growth of CFPAC-1 cells by blocking the interaction of bombesin with its receptors. Bombesin/GRP receptor antagonist RC-3095 could be con- sidered for the development of new approaches for treatment of human pancreatic cancers.
I N T R O D U C T I O N
Pancreatic cancer is one of the greatest challenges for oncologists (1-5). Carcinoma of the exocrine pancreas is the fifth leading cause of death from cancer in the United States (1). Most of pancreatic cancers are histologically ductal cell carcinomas, which constitute about 80- 90% of the cases (2). In the past two decades, great efforts have been made to improve the therapies for pancreatic cancers. However, the
overall prognosis of patients with pancreatic cancer is still very poor, and the 5-year survival rate is only 2-5% (2-4). Less than 15-20% of pancreatic tumors are resectable, mostly due to the difficulties in early diagnosis and the frequent occurrence of local or distal metastases, and fewer than 5% of the patients can survive for over 5 years postoperatively (3-5). Radiotherapy and chemotherapy are usually ineffective (2-5). Therefore, an urgent need exists to develop a new and effective therapy for treatment of patients with pancreatic cancers.
Recently, various investigations have demonstrated that gastroin- testinal hormones and growth factors may play important roles in the regulation of growth of normal and malignant exocrine pancreas (5- 10). In vitro studies have shown that gastrin, CCK, 3 and secretin stimulate the proliferation of pancreatic adenocarcinoma cells in tissue cultures (6, 7). Caerulein combined with secretin promotes the in vivo
growth of H-2-T pancreatic ductal adenocarcinoma cells in golden hamsters (8). Gastrointestinal hormones may also influence the phe- notypic transformation of pancreatic cells (6, 8, 9). These findings suggest that the growth of pancreatic cancers might be possibly con- trolled by hormonal manipulations (4-10), and various peptide ana- logues have been suggested for treatment (5-10).
Bombesin and its mammalian counterpart, GRP, are hormonal pep- tides which can exert diverse physiological or pharmacological ac- tions in various systems (5, 11, 12). In the gastrointestinal tract, bombesin and GRP stimulate gastric and pancreatic secretions, en- hance the release of several gastrointestinal hormones, and promote the growth of exocrine pancreas (5, 11, 12). Administration of bom- besin to rats produces hyperplasia and hypertrophy of pancreatic acinar cells (13). Bombesin and GRP appear to function as autocrine or paracrine growth factors and stimulate the growth of some normal or malignant cells including Swiss 3T3 fibroblast cells (14), human small cell lung cancer cells (15), human mammary and gastric cancer cells (16), 4 mouse and human colon cancer cells (17, 18), and CAPAN human pancreatic tumor cells (19). The trophic effect of bombesin/ GRP has also been demonstrated on the growth of azaserine-induced pancreatic acinar-cell adenocarcinomas in rats (20, 21) and on the cell proliferation of acinar tumor in primary cultures (22). The action of bombesin/GRP is thought to be mediated through its specific receptors present on the target cells (19, 22, 23). The discovery that bombesin appears to act as an autocrine growth factor in human small cell lung carcinoma (15, 23) and may also be involved in other cancers (16-22) has aroused major interest in the development of competitive bombesin/GRP receptor antagonists.
During the past few years, various bombesin/GRP antagonists in- cluding RC-3095 have been synthesized in our laboratory and evalu- ated for antitumor activity (24, 25). Receptor studies showed that these synthetic peptide analogues inhibit the binding of bombesin/ GRP to specific receptors on the membranes of Swiss 3T3 cells, SCLC cells, and human gastric cancer cells (24, 25). 4 Bombesin
Received 9/20/93; accepted 12/17/93. The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 This work was supported by NIH Grant CA 40077 and the Medical Research Service of the Veterans Affairs (A. V. S.).
2 To whom requests for reprints should be addressed, at Veterans Affairs Medical Center, 1601 Perdido Street, New Orleans, LA 70146.
3 The abbreviations used are: CCK, cholecystokinin; GRP, gastrin-releasing peptide; FCS, fetal calf serum; IMDM, lscove's modified Dulbecco's medium; PBS, phosphate- buffered saline; GHRH, growth hormone-releasing hormone; Tpi, 2,3,4,9-tetrahydro-lH- pyrido[3,4-b]indol-3-carboxylic acid; EGF, epidermal growth factor; [D-Trp6]LH-RH, o-tryptophan-6-1uteinizing hormone-releasing hormone.
4 y. Qin, G. Halmos, R-Z. Cai, B. Szoke, T. Ertl, and A. V. Schally. Inhibition of specific binding of bombesin and bombesin-stimulated growth of human gastric cancer cells by bombesin antagonists, submitted for publication.
1035
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from

BOMBESIN ANTAGONIST ON GROWTH OF PANCREATIC CANCER
antagonist RC-3095 has been shown to effectively suppress the
growth of HT 29 human colon cancers (26), PC-82 human prostate
cancers (27), MKN45 human gastric cancers xenograf ted in nude
mice (28), M X T breast cancers in mice (29), and ni t rosamine-induced
pancreatic cancers in hamsters (30). The inhibitory effect of antagonist
RC-3095 on the cell proliferation in vitro has also been demonstrated
in the cultures of some of these cancer cell lines (16, 25, 28). 4
RC-3095 and other novel bombesin antagonists could be considered
for the development of hormonal therapy for pancreatic cancers. How-
ever, the presence of specific receptors for bombesin /GRP has been
reported so far only in CAPAN human pancreatic cancer cells (19),
and the effects of bombes in /GRP and their antagonists on the growth
o f human pancreat ic ductal cancer cells must be evaluated more
extensively.
In the present study, we investigated the effect of bombesin antago-
nist RC-3095 on the growth of CFPAC-1 human ductal pancreatic
cancer cells implanted in nude mice. Direct effects of bombesin and
RC-3095 on the proliferation of CFPAC-1 cells were evaluated in cell
cultures. Specific receptors for bombes in /GRP on CFPAC-I cells
were characterized, and the ability of RC-3095 to inhibit the binding
of bombesin to the receptors was also studied.
M A T E R I A L S A N D M E T H O D S
Peptides and Chemicals. Bombesin antagonist D-Tpi6,Leul3aII[CH2NH] - Leu14-bombesin(6--14) (RC-3095), originally synthesized by a solid phase method and characterized in this laboratory (24, 25), was provided by Asta Pharm Co. (Frankfurt/M, Germany) for this study. Tyrn-bombesin and somat- ostatin-14 were obtained from Bachem (Torrence, CA) and Wyeth-Ayerst (Philadelphia, PA), respectively. Bombesin(1-14), GHRH(1-29), cholecysto- kinin-8, and D-Trp6-LHRH were all synthesized by a solid phase method in this laboratory (25, 31). 125I-Na and [methyl-3H]thymidine were purchased from Amersham (Arlington Heights, IL). Enzymobead iodination reagent was ob- tained from Bio-Rad (South Richmond, CA). FCS, IMDM, PBS, and Hanks' balanced salt solution were purchased from Gibco (Grand Island, NY). Univer- sol scintillation cocktail was obtained from ICN (Costa Mesa, CA). Other chemicals were purchased from Sigma (St. Louis, MO).
Cancer Cell Line. CFPAC-1 human pancreatic cancer cell line, originally established from a well-differentiated ductal pancreatic adenocarcinoma of a 26-year-old white male with cystic fibrosis (32), was purchased from the American Type Culture Collection (Rockville, MD). The cancer cells were routinely maintained as a monolayer culture in Costar T75 culture flasks with IMDM medium containing 10% FCS, 0.5 g/liter L-glutamine, 25 mM HEPES, 3.7 g/liter NaHCO3, 100 units/ml penicillin, 100/xg/ml streptomycin, and 0.25 /xg/ml amphotericin-B under humidified 5% CO2 at 37~ The cells growing exponentially were harvested by an incubation with 0.25% Trypsin-EDTA in calcium- and magnesium-free Hanks' balanced salt solution for 5 min at 37~ For tumor implantation, a single cell suspension was prepared in serum-free IMDM by repeatedly passing the cells through a G-22 needle; then the cells was diluted to a concentration of 5 • 106 cells/ml.
Implantation of Tumors in Nude Mice. Male athymic BALB/c (nu/nu) 6-week-old mice were obtained from the National Cancer Institute (Bethesda, MD) and housed in a laminar airflow cabinet under pathogen-free conditions throughout the experiments. Implantation of CFPAC-1 tumors was performed according to a method described previously (33). Three nude mice received s.c. injections in the flanks with 0.2 ml of cell suspension (1 X 106 cells) and served as tumor donors. After 4 weeks, the implanted tumors grew to a size of about 5 mm in diameter and were removed from the mice. Tumor samples were dissected free of necrotic tissue and blood vessels and were cut into small fragments of about 8 mm 3. Under ether anesthesia, two pieces of tumor fragments were implanted s.c. by trocars on both sides of the flanks for each mouse. The mice bearing the implanted tumors were randomly divided into two groups with 10 mice in each group.
Treatment with Bombesin Antagonist RC-3095. The nude mice with implanted tumors started to receive injections of bombesin antagonist RC-3095 7 days after the tumor cell injection. RC-3095 solution was freshly prepared by dissolving the peptide in 200 lxl of 0.01 M acetic acid and was diluted with
0.9% NaCI containing 0.2% bovine serum albumin to a concentration of 125 p.g/ml. Each mouse in the treatment group was given s.c. injections of RC- 3095 at a dose of 10/.~g twice daily for 25 days. This dose of RC-3095 was chosen on the basis of a previous report of the inhibitory effect of RC-3095 on the growth of HT29 human colon cancers in nude mice (26). The animals in the control group were injected with the same volume of vehicle but without RC-3095.
Evaluation of Tumor Growth. During the treatment with RC-3095, the size of the implanted tumors was measured by calipers in each mouse at 3-4 day intervals for 25 days to construct the tumor growth curves in vivo. Tumor volume was calculated by the formula: Tumor volume = Length • width 2 • 0.5. Tumor growth parameters, Le., tumor volume doubling time, tumor growth delay time, and tumor growth rate, were calculated from the tumor growth curves as described previously (34). In this study, the tumor volume doubling time was defined as the time required for the tumors to grow from 50 mm 3 to 100 mm 3 for the control group and from 35 mm 3 to 70 mm 3 for the treatment group, respectively. The tumor growth delay time was estimated as the time difference for the treated tumors and the controls to reach a volume of 70 mm 3. At the end of the treatment, the animals were sacrificed with an overdose of ether. The tumors were removed from the animals, weighed, and immediately frozen in liquid nitrogen for measurements of DNA and protein contents in tumor tissues.
Determination of DNA and Protein in Tumor Tissues. DNA in tumor tissues was determined by a method of Labara and Paigen (35, 36), which is based on the enhancement of fluorescence reaction upon binding bisbenzimid- azole Hoechst 33258 (H33258) to DNA in cell nuclei in a high ionic strength solution. Tumors collected in each group were pooled and homogenized in 10 times their volumes of a buffer consisting of 0.05 M NaH2PO4, 2.0 M NaCl, and 2 mM EDTA (pH 7.4). H33258 was dissolved in the same buffer at a concen- tration of 1/xg/ml and filtered before use. An aliquot of tumor homogenate (0.4 ml) was suspended in 4 ml of H33258 solution, followed by an incubation in a dark room for 30 min. The reaction was measured by a fluorescence spec- trophotometer at excitation and emission wavelengths of 356 and 492 nm, respectively. Calf thymus DNA type I was used as a standard reference.
Protein in tumor tissues was measured by a modified method of Bradford (37) using bovine serum albumin fraction V as a standard.
Measurement of Cell Growth in Vitro. The effects of bombesin and bombesin antagonist RC-3095 on the growth of CFPAC-1 human pancreatic cancer cells in vitro was evaluated by direct cell counting and [3H]thymidine incorporation assay.
Direct Cell Counting. CFPAC-1 cells collected from 60-70% confluent cultures were used for this study and seeded to 24-well culture plates (1 • 104 cells/well). After the cells were cultured in IMDM containing 10% FCS for 48 h, the medium was replaced by IMDM supplemented with 2.5% FCS and various concentrations of bombesin(1-14), bombesin antagonist RC-3095, or a combination of both. The same volume of medium but without peptides was added to the control wells. Following another 24 h of incubation, the culture was terminated by aspiration of the medium from the wells and washing with PBS (0.5 ml/well). The cells were trypsinized by a 10-rain incubation with (0.5 ml/well) 0.25% Trypsin-EDTA. The detached cells were dispersed by repeated pipeting using a G-22 needle and syringe. The number of cells collected from each well was counted by an automated electronic cell counter (Coulter Counter Modei-ZF; Coulter Electronics, Inc., Hialeah, FL).
[3H]Thymidine Incorporation Assay. DNA synthesis in tumor cells was measured by [3H]thymidine incorporation assay as reported previously (38). Single cell suspension was prepared in IMDM with 10% FCS and was seeded to 24-well culture plates (1 • 104 cells/well). After 48 h of culture, the medium was changed to IMDM (0.5 ml/well) containing 2.5% FCS and various con- centrations of bombesin(1-14), bombesin antagonist RC-3095, or a combina- tion of both. The control wells received the same medium without bombesin or RC-3095. After 24 h of culture, [methyl-3H]thymidine (radioactivity 25 Ci/ml) was added to each well (1 /zCi/well) to pulse the cells. After a 4-h incubation, the medium was removed from the wells, and the cells were fixed by Camoy's solution (1 ml/well; methanol:glacial acetic acid, 3:1 v/v) for 20 min. After washing three times with PBS, the cells in each well were dissolved with 0.5 ml of 0.3 N NaOH for 15 min at room temperature. The cell lysate was collected from each well and mixed with 3 ml of Universol scintillation cocktail. The radioactivity was measured for 1 min by a liquid scintillation beta counter (Mark III; Searle Analytic, Inc., Des Plaines, IL).
1036
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from
BOMBESIN ANTAGONIST ON GROWTH OF PAN('RkAT[(" CANCER
Receptor Binding Assay. Receptor binding assay was performed using intact CFPAC-I cells in monolayer cultures. Tyr4-bombesin was radiolabeled with '25I-Na using a Bio-Rad enzymobead iodination kit. Mono-J25I-Tyr 4- bombesin was purified by high performance liquid chromatography as de- scribed (39). The specific activity of '25I-Tyr4-bombesin was about 2,000 Ci/mmol. CFPAC-1 cells were seeded to 24-well culture plates (1 • 10 4
cells/well) and cultured with IMDM containing 10% FCS for 48 h. The cells in subconfluent culture were washed once with serum-free IMDM supple- mented with 25 mM 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid, 10 mM MgCI2, 1 mM EGTA, 10 mM monothioglycerol, 0.25 mM phenylmethyl- sulfonyl fluoride, aprotinin 10,000 kallikrein inactivator units/liter, and 0.1% bovine serum albumin (pH 7.5), followed by an incubation for 2 h at 22~ with the same medium containing (0.5 riM) '25I-Tyr4-bombesin in the presence or absence of various concentrations of unlabeled bombesin(1-14), bombesin antagonist RC-3095, or structurally unrelated peptides somatostatin-14, cho- lecystokinin-8, GHRH(1-29), and D-Trp6-LHRH. The binding reaction was terminated by adding 0.5 ml of ice-cold medium to each well. After washing four times with ice-cold PBS (pH 7.4), the cells in each well were dissolved with 0.5 ml of 0.3 N NaOH. The resultant cellular lysate was collected from each well for measurement of radioactivity by a gamma counter (APEX; Microdemic System, Inc., Huntsville, AL).
Statistical Analysis. All data are expressed as the mean _+ SEM of dupli- cate or triplicate observations from at least 2-3 repeated experiments. Mean values between the RC-3095 treatment group and the control group were analyzed by the Student t test, Mann-Whitney U test, or one-way analysis of variance. The difference was statistically considered to be significant when a two-tailed value of P was less than 0.05.
Data from receptor binding assays were analyzed by a ligand-PC comput- erized curve-fitting program created by Munson and Rodbard as modified by McPherson (40) to determine the types of binding sites, the dissociation constants (Kd), and the maximal binding capacity of receptors (B,,,,=).
RESULTS
Effect of RC-3095 on Growth of CFPAC-1 Tumors in Nude Mice. Nude mice with implanted CFPAC-1 human pancreatic cancers started to receive injections of bombesin antagonist RC-3095 7 days after the tumor inoculation when all animals had palpable tumors in the flanks. Injections of RC-3095 at a dose of 10/~g twice a day for 25 days did not show any obvious side effects on the growth of the treated animals. There was no significant difference in body weights between the RC-3095-treated and the control groups (P > 0.05).
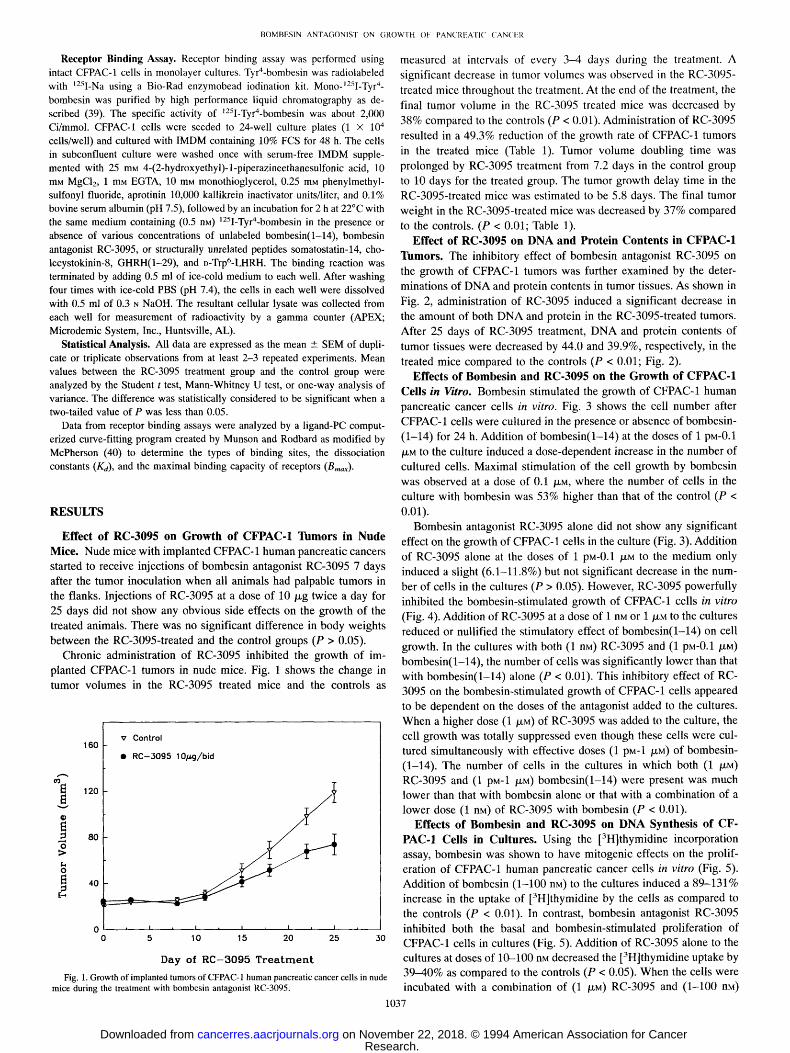
Chronic administration of RC-3095 inhibited the growth of im- planted CFPAC-1 tumors in nude mice. Fig. 1 shows the change in tumor volumes in the RC-3095 treated mice and the controls as
v C o n t r o l 1 6 0
�9 R C - 3 0 9 5 l O / ~ : j / b i d
d-"
1 2 0
80 O ~>
O
40 [.-,
0 i
0 3 0 I , , I , I J l , I
5 10 15 20 25
Day of RC-3095 T r e a t m e n t
Fig. 1. Growth of implanted tumors of CFPAC-1 human pancreatic cancer cells in nude mice during the treatment with bombesin antagonist RC-3095.
measured at intervals of every 3--4 days during the treatment. A significant decrease in tumor volumes was observed in the RC-3095- treated mice throughout the treatment. At the end of the treatment, the final tumor volume in the RC-3095 treated mice was decreased by 38% compared to the controls (P < 0.01). Administration of RC-3095 resulted in a 49.3% reduction of the growth rate of CFPAC-1 tumors in the treated mice (Table 1). Tumor volume doubling time was prolonged by RC-3095 treatment from 7.2 days in the control group to 10 days for the treated group. The tumor growth delay time in the RC-3095-treated mice was estimated to be 5.8 days. The final tumor weight in the RC-3095-treated mice was decreased by 37% compared to the controls. (P < 0.01; Table 1).
Effect of RC-3095 on DNA and Protein Contents in CFPAC-1 Tumors. The inhibitory effect of bombesin antagonist RC-3095 on the growth of CFPAC-1 tumors was further examined by the deter- minations of DNA and protein contents in tumor tissues. As shown in Fig. 2, administration of RC-3095 induced a significant decrease in the amount of both DNA and protein in the RC-3095-treated tumors. After 25 days of RC-3095 treatment, DNA and protein contents of tumor tissues were decreased by 44.0 and 39.9%, respectively, in the treated mice compared to the controls (P < 0.01; Fig. 2).
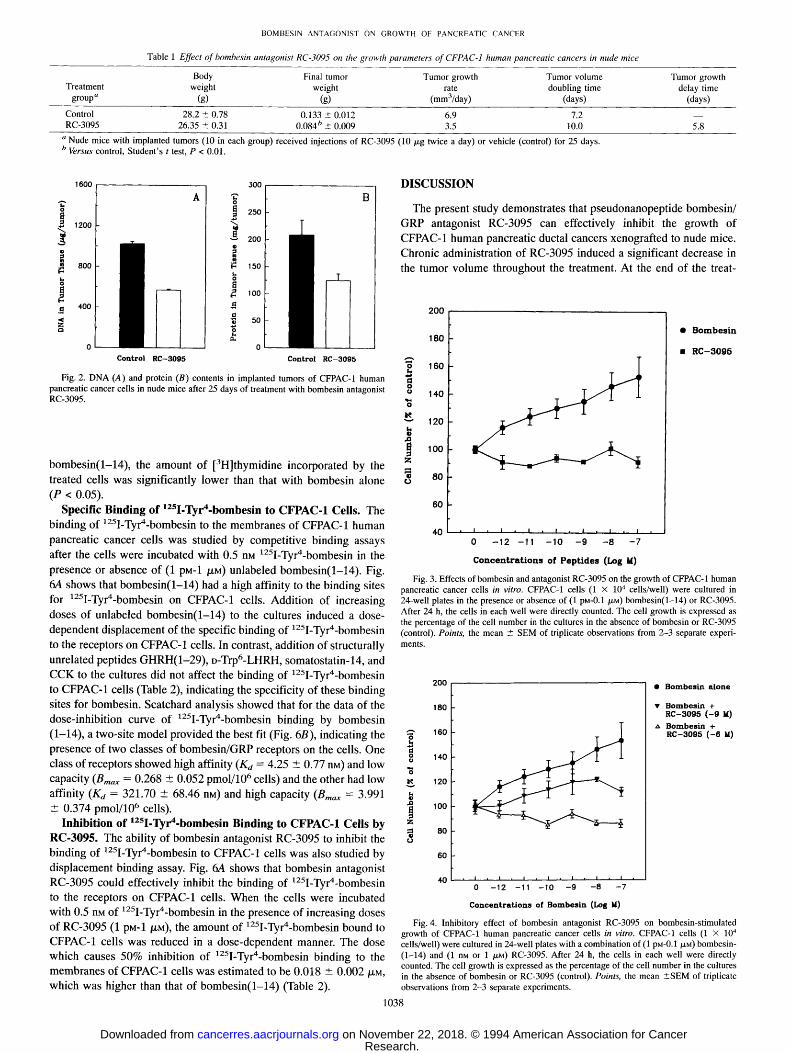
Effects of Bombesin and RC-3095 on the Growth of CFPAC-1 Cells in Vitro. Bombesin stimulated the growth of CFPAC-1 human pancreatic cancer cells in vitro. Fig. 3 shows the cell number after CFPAC- 1 cells were cultured in the presence or absence of bombesin- (1-14) for 24 h. Addition of bombesin(1-14) at the doses of i pM-0.1 /xM to the culture induced a dose-dependent increase in the number of cultured cells. Maximal stimulation of the cell growth by bombesin was observed at a close of 0.1 /~M, where the number of cells in the culture with bombesin was 53% higher than that of the control (P < 0.01).
Bombesin antagonist RC-3095 alone did not show any significant effect on the growth of CFPAC-1 cells in the culture (Fig. 3). Addition of RC-3095 alone at the doses of 1 pM-0.1 /XM to the medium only induced a slight (6.1-11.8%) but not significant decrease in the num- ber of cells in the cultures (P > 0.05). However, RC-3095 powerfully inhibited the bombesin-stimulated growth of CFPAC-1 cells in vitro
(Fig. 4). Addition of RC-3095 at a dose of 1 nM or 1 /XM to the cultures reduced or nullified the stimulatory effect of bombesin(1-14) on cell growth. In the cultures with both (1 riM) RC-3095 and (1 pM-0.1 p~M) bombesin(1-14), the number of cells was significantly lower than that with bombesin(1-14) alone (P < 0.01). This inhibitory effect of RC- 3095 on the bombesin-stimulated growth of CFPAC-1 cells appeared to be dependent on the doses of the antagonist added to the cultures. When a higher dose (1 /XM) of RC-3095 was added to the culture, the cell growth was totally suppressed even though these cells were cul- tured simultaneously with effective doses (1 pM-1 /.ZM) of bombesin- (1-14). The number of cells in the cultures in which both (1 /~M) RC-3095 and (1 pM-1 /XM) bombesin(1-14) were present was much lower than that with bombesin alone or that with a combination of a lower dose (1 riM) of RC-3095 with bombesin (P < 0.01).
Effects of Bombesin and RC-3095 on DNA Synthesis of CF- PAC-1 Cells in Cultures. Using the [3H]thymidine incorporation assay, bombesin was shown to have mitogenic effects on the prolif- eration of CFPAC-1 human pancreatic cancer cells in vitro (Fig. 5). Addition of bombesin (1-100 nM) to the cultures induced a 89-131% increase in the uptake of [3H]thymidine by the cells as compared to the controls (P < 0.01). In contrast, bombesin antagonist RC-3095 inhibited both the basal and bombesin-stimulated proliferation of CFPAC-1 cells in cultures (Fig. 5). Addition of RC-3095 alone to the cultures at doses of 10-100 nM decreased the [3H]thymidine uptake by 39-40% as compared to the controls (P < 0.05). When the cells were incubated with a combination of (1 /XM) RC-3095 and (1-100 riM)
1037
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from
BOMBESIN ANTAGONIST ON GROWTtt OF PANCREATIC CANCER
Table 1 Effect of bombesin antagonist RC-3095 on the growth parameters of CFPAC-1 human pancreatic cancers in nude mice
Body Final tumor Tumor growth Tumor volume Tumor growth Treatment weight weight rate doubling time delay time
group" (g) (g) (mm3/day) (days) (days)
Control 28.2 • 0.78 0.133 • 0.012 6.9 7.2 - - RC-3095 26.35 • 0.31 0.084 t' • 0.009 3.5 10.0 5.8
a Nude mice with implanted tumors (10 in each group) received injections of RC-3095 (10 ~g twice a day) or vehicle (control) for 25 days. b Versus control, Student's t test, P < 0.01.
1 6 0 0 D I S C U S S I O N
3 1 2 0 0
== 8oo
.~ 400
3 0 0
250
~ 2 0 0
= ~, 15o
I O0
50
0
Contro l R C - 3 0 9 5 Contro l R C - 3 0 9 5
Fig. 2. DNA (A) and protein (B) contents in implanted tumors of CFPAC-1 human pancreatic cancer cells in nude mice after 25 days of treatment with bombesin antagonist RC-3095.
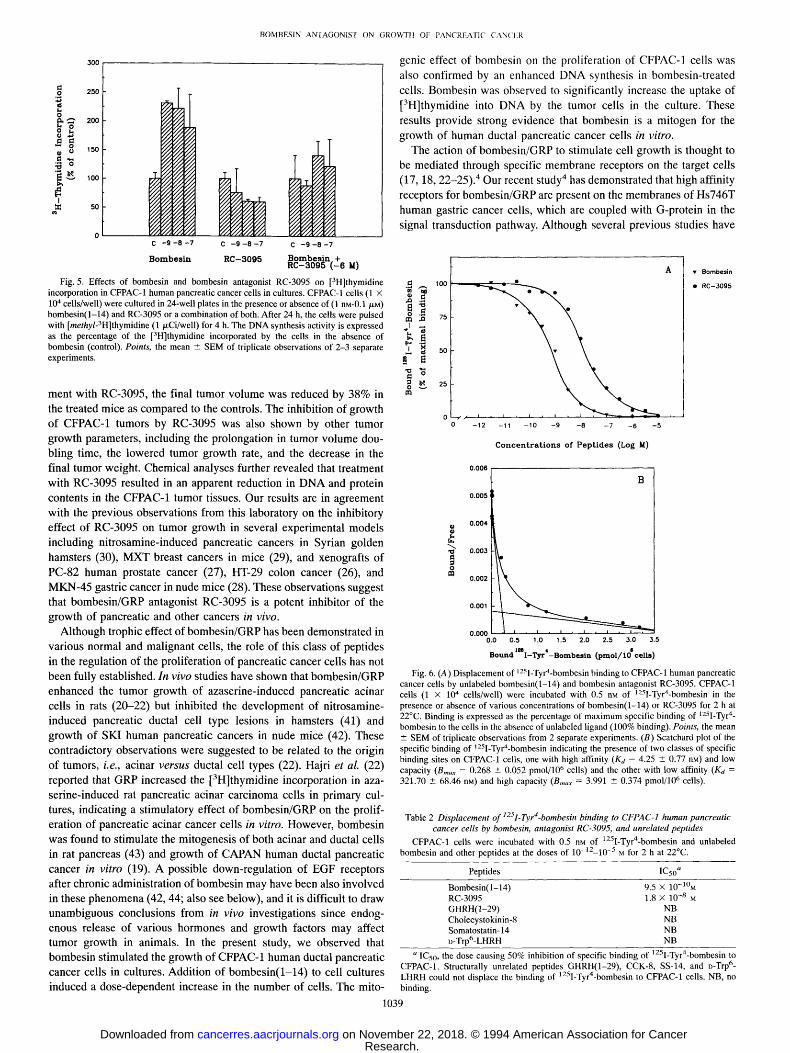
bombesin(1-14), the amount of [3H]thymidine incorporated by the treated cells was significantly lower than that with bombesin alone (e < 0.05).
Specific Binding of lZSI-Tyr4-bombesin to CFPAC-1 Cells. The binding of 125I-Tyr4-bombesin to the membranes o f CFPAC-1 human
pancreatic cancer cells was studied by compet i t ive binding assays
after the cells were incubated with 0.5 nM 125I-Tyr4-bombesin in the
presence or absence o f (1 pM-1 /xM) unlabeled bombes in (1 -14) . Fig.
6,4 shows that bombes in (1 -14 ) had a high affinity to the binding sites
for 12SI-Tyr4-bombesin on CFPAC-1 cells. Addit ion of increasing
doses of unlabeled bombes in (1 -14 ) to the cultures induced a dose-
dependent displacement of the specific binding o f 12SI-Tyr4-bombesin
to the receptors on CFPAC-1 cells. In contrast, addition of structurally
unrelated pept ides G H R H ( 1 - 2 9 ) , D-Trp6-LHRH, somatostat in-14, and
C C K to the cultures did not affect the binding of 125I-Tyr4-bombesin
to CFPAC-1 cells (Table 2), indicating the specifici ty of these binding
sites for bombesin . Scatchard analysis showed that for the data of the
dose-inhibi t ion curve of 125I-Tyr4-bombesin binding by bombes in
(1-14) , a two-si te model provided the best fit (Fig. 6B), indicating the
presence o f two classes of b o m b e s i n / G R P receptors on the cells. One
class of receptors showed high affinity (Ka = 4.25 _ 0.77 nM) and low
capaci ty (Bmax : 0.268 ___ 0.052 pmol/106 cells) and the other had low
affinity (Ka = 321.70 ___ 68.46 riM) and high capaci ty (Bmax --- 3.991
- 0.374 pmol/106 cells).
Inh ib i t ion o f 12SI-Tyr4-bombesin B ind ing to C F P A C - I Cells by
RC-3095 . The ability of bombes in antagonist RC-3095 to inhibit the
binding of 125I-Tyr4-bombesin to CFPAC-1 cells was also studied by
displacement binding assay. Fig. 6A shows that bombes in antagonist 1 2 5 4 RC-3095 could effectively inhibit the binding of I-Tyr -bombesin
to the receptors on CFPAC-1 cells. When the cells were incubated 1 2 5 4 with 0.5 nM of I-Tyr -bombesin in the presence of increasing doses
1 2 5 4 of RC-3095 (1 pr~-I/xM), the amount of I-Tyr -bombesin bound to CFPAC-1 cells was reduced in a dose-dependent manner. The dose
1 2 5 4 which causes 50% inhibition of I-Tyr -bombesin binding to the membranes of CFPAC-1 cells was est imated to be 0.018 + 0.002 p,M,
which was higher than that o f bombes in (1 -14 ) (Table 2).
The present study demonstrates that pseudonanopeptide bombesin/ GRP antagonist RC-3095 can effectively inhibit the growth of CFPAC-1 human pancreatic ductal cancers xenografted to nude mice. Chronic administration of RC-3095 induced a significant decrease in the tumor volume throughout the treatment. At the end of the treat-
2 0 0
180
. 16o
o 14.0
" " 120
,ID
I::I 1 0 0 :::I Z
d 8o
60
�9 Bombesin
�9 RC-3095
4 0 i i . i , I , i . i , i .
0 - 1 2 - 1 1 - 1 0 - 9 - 8 - 7
Concentrations of Peptides ( log M)
Fig. 3. Effects of bombesin and antagonist RC-3095 on the growth of CFPAC-1 human pancreatic cancer cells in vitro. CFPAC-1 cells (1 • 104 cells/well) were cultured in 24-well plates in the presence or absence of (1 pM-0.1 ~M) bombesin(1-14) or RC-3095. After 24 h, the cells in each well were directly counted. The cell growth is expressed as the percentage of the cell number in the cultures in the absence of bombesin or RC-3095 (control). Points, the mean - SEM of triplicate observations from 2-3 separate experi- ments.
2 0 0 �9 B o m b e s i n a l o n e
1 8 0 �9 B o m b e s i n § ~c-ao95 ( - 9 u)
A Bombes in + 1 6 0 R C - 3 0 9 5 ( - 6 M) o
"~ 140
o 1 2 0 ~ , ~
"~ 100
z RO
60
4.0 , i , i , i , i , i , i , i ,
0 - 1 2 - 1 1 - 1 0 - 9 - 8 - 7
C o n c e n t r a t i o n s o f B o m b e s i n ( L o g M)
Fig. 4. Inhibitory effect of bombesin antagonist RC-3095 on bombesin-stimulated growth of CFPAC-1 human pancreatic cancer cells in vitro. CFPAC-1 cells (1 • 104
cells/well) were cultured in 24-well plates with a combination of (1 pM-0.1 tXM) bombesin- (1-14) and (1 nM or 1 /xr~) RC-3095. After 24 h, the cells in each well were directly counted. The cell growth is expressed as the percentage of the cell number in the cultures in the absence of bombesin or RC-3095 (control). Points, the mean • of triplicate observations from 2-3 separate experiments.
1038
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from
BOMBESIN ANTAGONIST ON GROWTH OF PANCREATIC C A N ( E R
300
o 250
r
IZ, ~ 200
o 150
-B o ~ loo
.=
I 50
C - 9 - 8 C - 9 - 8 - 7 C - 9 - 8 - 7
genic effect of bombesin on the proliferation of CFPAC-I cells was also confirmed by an enhanced DNA synthesis in bombesin-treated cells. Bombesin was observed to significantly increase the uptake of [3H]thymidine into DNA by the tumor cells in the culture. These results provide strong evidence that bombesin is a mitogen for the growth of human ductal pancreatic cancer cells in vitro.
The action of bombesin/GRP to stimulate cell growth is thought to be mediated through specific membrane receptors on the target cells (17, 18, 22-25). 4 Our recent study 4 has demonstrated that high affinity receptors for bombesin/GRP are present on the membranes of Hs746T human gastric cancer cells, which are coupled with G-protein in the signal transduction pathway. Although several previous studies have
Bombesin RC-3095 Bombesin + RC-3095 (-6 M)
Fig. 5. Effects of bombesin and bombesin antagonist RC-3095 on [3H]thymidine incorporation in CFPAC-1 human pancreatic cancer cells in cultures. CFPAC-1 cells (1 x 104 cells/well) were cultured in 24-well plates in the presence or absence of (1 nM-0.1 #.M) bombesin(1-14) and RC-3095 or a combination of both. After 24 h, the cells were pulsed with [methyl-3H]thymidine (1 ~Ci/well) for 4 h. The DNA synthesis activity is expressed as the percentage of the [3H]thymidine incorporated by the ceils in the absence of bombesin (control). Points, the mean +- SEM of triplicate observations of 2-3 separate experiments.
ment with RC-3095, the final tumor volume was reduced by 38% in the treated mice as compared to the controls. The inhibition of growth of CFPAC-1 tumors by RC-3095 was also shown by other tumor growth parameters, including the prolongation in tumor volume dou- bling time, the lowered tumor growth rate, and the decrease in the final tumor weight. Chemical analyses further revealed that treatment with RC-3095 resulted in an apparent reduction in DNA and protein contents in the CFPAC-1 tumor tissues. Our results are in agreement with the previous observations from this laboratory on the inhibitory effect of RC-3095 on tumor growth in several experimental models including nitrosamine-induced pancreatic cancers in Syrian golden hamsters (30), MXT breast cancers in mice (29), and xenografts of PC-82 human prostate cancer (27), HT-29 colon cancer (26), and MKN-45 gastric cancer in nude mice (28). These observations suggest that bombesin/GRP antagonist RC-3095 is a potent inhibitor of the growth of pancreatic and other cancers in vivo.
Although trophic effect of bombesin/GRP has been demonstrated in various normal and malignant cells, the role of this class of peptides in the regulation of the proliferation of pancreatic cancer cells has not been fully established. In vivo studies have shown that bombesin/GRP enhanced the tumor growth of azaserine-induced pancreatic acinar cells in rats (20-22) but inhibited the development of nitrosamine- induced pancreatic ductal cell type lesions in hamsters (41) and growth of SKI human pancreatic cancers in nude mice (42). These contradictory observations were suggested to be related to the origin of tumors, i.e., acinar versus ductal cell types (22). Hajri et al. (22) reported that GRP increased the [3H]thymidine incorporation in aza- serine-induced rat pancreatic acinar carcinoma cells in primary cul- tures, indicating a stimulatory effect of bombesin/GRP on the prolif- eration of pancreatic acinar cancer cells in vitro. However, bombesin was found to stimulate the mitogenesis of both acinar and ductal cells in rat pancreas (43) and growth of CAPAN human ductal pancreatic cancer in vitro (19). A possible down-regulation of EGF receptors after chronic administration of bombesin may have been also involved in these phenomena (42, 44; also see below), and it is difficult to draw unambiguous conclusions from in vivo investigations since endog- enous release of various hormones and growth factors may affect tumor growth in animals. In the present study, we observed that bombesin stimulated the growth of CFPAC-1 human ductal pancreatic cancer cells in cultures. Addition of bombesin(1-14) to cell cultures induced a dose-dependent increase in the number of cells. The mito-
~ 100
k ~" "~ 5o
0 ~ N 25 m
A
�9 - �9 �9 �9 �9
o - 1 2 -11 - 1 0 - 9 - 8 - 7 - 6 - 5
Concentrations of Peptides (Log M)
�9 Bombesln
�9 RC-3095
"r
o
0.006 .
0.005 f
0,004
0.003
0.002
0.001
0.000 i , , �9 , �9 , �9 , �9 0.0 0.5 1,0 1.5 2.0 2.5 3,0 3.5
118 4 l
Bound l-Tyr -Bombes in ( p m o l / l O c e l l s )
Fig. 6. (A) Displacement of 1251.Tyr4.bombesin binding to CFPAC-1 human pancreatic cancer cells by unlabeled bombesin(l-14) and bombesin antagonist RC-3095. CFPAC-I cells (1 x 104 cells/well) were incubated with 0.5 nM of ]25I-Tyr4-bombesin in the presence or absence of various concentrations of bombesin(1-14) or RC-3095 for 2 h at 22~ Binding is expressed as the percentage of maximum specific binding of 125I-Tyr 4- bombesin to the cells in the absence of unlabeled ligand (100% binding). Points, the mean -+ SEM of triplicate observations from 2 separate experiments. (B) Scatchard plot of the specific binding of 12-Sl-Tyr4-bombesin indicating the presence of two classes of specific binding sites on CFPAC-1 cells, one with high affinity (Ka = 4.25 +- 0.77 riM) and low capacity (Bmax = 0.268 -+ 0.052 pmol/106 cells) and the other with low affinity (Ka = 321.70 • 68.46 riM) and high capacity (Bmax = 3.991 _+ 0.374 pmol/106 cells).
Table 2 Displacement of 1251-Tyr4-bombesin binding to CFPAC-1 human pancreatic cancer cells by bombesin, antagonist RC-3095, and unrelated peptides
CFPAC-1 cells were incubated with 0.5 nM of 125I-Tyr4-bombesin and unlabeled bombesin and other peptides at the doses of 10-12-10 -5 M for 2 h at 22~
Peptides IC5o"
Bombesin(1-14) 9.5 • 10-10M RC-3095 1.8 • 10 -8 M GHRH(I-29) NB Cholecystokinin-8 NB Somatostatin- 14 NB D-Trp6-LHRH NB
'~ IC5o, the dose causing 50% inhibition of specific binding of 1251-Tyr4-bombesin to CFPAC-1. Structurally unrelated peptides GHRH(1-29), CCK-8, SS-14, and D-Trp 6- LHRH could not displace the binding of 125I-Tyr4-bombesin to CFPAC-1 cells. NB, no binding.
1039
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from

BOMBESIN ANTAGONIST ON GROWTtt OF I'AN('REATI(' CANCER
shown that bombesin/GRP receptors are widely distributed on the membranes of many benign and malignant tumor cells including rat pancreatic acinar cancer cells (17, 18, 22-26), 4 the presence of
bombesin/GRP receptors in human pancreatic cancer cells has been reported so far only by Avis et al. (19). In our previous study, antago- nist RC-3095 did not inhibit the growth of MIA PaCa-2 human pan- creatic cancer xenograft in nude mice (45). It is possible that our MIA PaCa-2 line has undergone some changes in morphology and receptor content. The absence of receptors may have been a contributing factor as specific binding sites for bombesin were not found in our cultures of MIA PaCa-2 cells and neither bombesin nor bombesin antagonist RC-3095 influenced cell growth in vitro (45). In the present study, we demonstrated that specific receptors for bombesin/GRP are present on the membranes of CFPAC-1 human ductal pancreatic cancer cells. In competitive receptor binding studies, we observed that bombesin (1-14) was able to effectively displace 125I-Tyra-bombesin from the binding sites on CFPAC-1 cells. In contrast, structurally unrelated peptides such as somatostatin-14, cholecystokinin-8, o-Trp6-LHRH, and GHRH(1-29) did not affect the binding of 125I-Tyr4-bombesin to the cell membranes. Scatchard analysis indicated that two types of bombesin/GRP receptors are present on the membranes of this cancer cell line, one class having a high affinity and low capacity and the other class having a low affinity and high capacity. Characteristics of bombesin receptors on CFPAC-1 cells are similar to those on MC-26 mouse colon cancer cells, which also show two classes of binding sites for bombesin/GRP (17). These receptors are apparently different from those on Swiss 3T3 cells (24) and many other tumor cell lines (18, 22, 23, 25) 4 in which only one class of binding sites is found. Because bombesin was observed to have direct stimulatory effects on the proliferation of CFPAC-1 cells in cultures, our receptor studies suggest that the specific binding sites for bombesin/GRP on CFPAC-1 cells may represent functional receptors which mediate the action of bombesin on the growth of these tumor cells.
RC-3095 [D-Tpi6,Leu13~[CH2-NH]Leu14-bombesin(6-14)] is a potent bombesin antagonist, which was synthesized in our laboratory specially for antitumor therapy. Previous studies have shown that this synthetic peptide inhibits the binding of a25I-Tyr4-bombesin to the membranes of Swiss 3T3 cells (24) and H-345 human small cell lung cancer cells (25) and suppresses the basal and GRP-induced amylase release from rat pancreatic acini in a superfusion assay (24). RC-3095 was proven to be a potent tumor growth inhibitor in experimental models of nitrosamine-induced pancreatic cancers in hamsters (30), MXT breast cancers in mice (29), PC-82 human prostate cancers (27), MKN 45 gastric cancers (28), and HT-29 colon cancers in nude mice (26). The direct inhibitory effect of RC-3095 was observed on the cell proliferation of mammary (16) and gastric cancer cell lines (28). 4 The results of the present study further demonstrate that the addition of bombesin antagonist RC-3095 alone to the medium produced only a slight but not significant decrease in the number of cells in the culture. This suggests that RC-3095 itself does not affect the growth of CFPAC-1 cells in vitro, but it can effectively inhibit the bombesin- stimulated growth of CFPAC-1 cells in cultures. The presence of RC-3095 in culture medium reduced or nullified the stimulatory ac- tion of bombesin on the proliferation of CFPAC-1 cells as evidenced by a lower cell count in cultures treated with both RC-3095 and bombesin as compared to that with bombesin alone. Furthermore, RC-3095 inhibited DNA synthesis in CFPAC-1 cells during the cell proliferation. Addition of RC-3095 to the cultures reduced the bom- besin-enhanced DNA synthesis in the CFPAC-1 cells as measured by [3H]thymidine incorporation assay. The inhibitory effect of RC-3095 on bombesin-stimulated proliferation of CFPAC-1 cells was appar- ently dependent on the doses of RC-3095 added to the cultures. On the basis of both cell number and [3H]thymidine incorporation assay,
1040
addition of RC-3095 at a higher dose of 1 /xM completely suppressed the bombesin-induced growth of CFPAC-I cells in cultures, suggest- ing that the action of this antagonist is competitive. This view is supported by the observations of the inhibitory effect of RC-3095 on the binding of bombesin to its receptors on CFPAC-1 cells. In dis- placement binding assays, we noticed that bombesin antagonist RC- 3095 blocked, in a dose-dependent manner, the binding of 125I-Tyr4- bombesin to CFPAC-1 cells. Our findings indicate that the inhibitory effect of RC-3095 on bombesin-induced growth of CFPAC-1 cells is probably due to competitive occupation of the binding sites for bombesin/GRP on the cells.
However, the suppressive effect of RC-3095 on the growth of CFPAC-1 tumors in nude mice cannot be solely explained by the competitive action of this antagonist to displace the binding of bombesin/GRP to the receptors on the tumor cells. Several other regulatory mechanisms, such as a major down-regulation of EGF receptors, may also mediate the action of RC-3095 (26-30, 44). It has been found that in vivo treatment with RC-3095 induced a down- regulation of receptors for both bombesin/GRP and EGF on the mem- branes of MKN45 gastric cancers in nude mice (28) and greatly reduced concentration of EGF receptors in pancreatic, mammary, colorectal, and prostatic cancers (27, 29, 30, 44). RC-3095 may also affect EGF binding through an action on the protein kinase C system (46) and inhibition of up-regulation of EGF receptors produced by bombesin/GRP (47). Whether the effect of RC-3095 on the growth of CFPAC-1 tumors is related to the down-regulation of bombesin/GRP and EGF receptors on the membranes of CFPAC-1 cancer cell line or the interference with the function of EGF receptors (46, 47) needs to be elucidated. In addition, since bombesin/GRP can powerfully stimu- late the release of gastrointestinal hormones that may have trophic effects on the growth of pancreas and pancreatic cancers (5-12), the possible involvement of RC-3095 not only in the inhibition of action of bombesin/GRP but in the suppression of the release of these trophic hormones should be also considered. It is possible that the inhibitory action of RC-3095 on tumor growth of CFPAC-1 cells in vivo is mediated through several mechanisms.
In summary, our study shows that bombesin/GRP antagonist RC- 3095 can powerfully suppress the growth of CFPAC-1 human ductal pancreatic cancer cells xenografted into nude mice as well as cultured in vitro. The effect of RC-3095 appears to be mediated by the inter- ference with the binding of bombesin to its receptors on this cancer cell line. These findings suggest the merit of continued evaluation of bombesin/GRP antagonist RC-3095 for the possible development of new approaches for the treatment of pancreatic cancers.
A C K N O W L E D G M E N T S
We express our thanks to Drs. Balazs Szoke, Kate Groot, and Herta Reile, as well as Patti Armatis, for their help with iodination and cell cultures and to Harold Valerio and Nancy Hsi for excellent technical assistance.
REFERENCES
1. Boring, C. C., Squires, T. S., and Tong, T. Cancer Statistics, 1991. CA Cancer J. Clin., 41: 19-36, 1991.
2. Lin, R. S., Kessler, I. I. A multifactorial model for pancreatic cancer in man. Epide- miologic evidence. J. Am. Med. Assoc., 245: 147-152, 1981,
3. Williams, R. C. N. Pancreatic cancer: the greatest oncological challenge. Br. Med. J., 296: 445--446, ! 988.
4. Theve, N. O.. Pousctte, A., and Carlstrom, K. Adenocarcinoma of the pancreas--a hormone sensitive tumor? A preliminary report on Nolvadex treatment. Clin. Oncol., 9: 193-197, 1983.
5. Poston, G. J., Gillespie, J., and Guillou, P. J. The biology of pancreatic cancer (review). Gut, 32: 800--812, 1991.
6. Johnson, L. R. Effects of gastrointestinal hormones on pancreatic growth. Cancer (Phila.), 47: 1640-1645, 1981.
7. Edwards, B. E, Redding, T. W., and Schally, A. V. The effect of gastrointestinal hormones on the incorporation of tritiated thymidine in the pancreatic adenocarci-
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from

BOMBESIN ANTAGONIST ON GROWTH OF PANCREATIC CANCER
noma cell line (WD PaCa). Int. J. Pancreatol., 5: 191-201, 1989. 8. Townsend, C. M., Franklin, R. B., Watson, L. C., Glass, E., and Thompson, J.
Stimulation of pancreatic cancer growth by caerulein and secretin. Surg. Forum, 32: 228--229, 1981.
9. Schally, A. V. Oncological applications of somatostatin analogs. Cancer Res., 48: 6977-6985, 1988.
10. Liebow, C., and Du Sapin, K. Hormonal control of pancreatic cancer growth. Pan- creas, 1: 44-48, 1986.
11. Spindel, E. Mammalian bombesin-like peptides. Trends Neurosci., 9: 130-133, 1986. 12. Wood, S. M., Jung, R. T., Webster, J. D., Gasthei, M. A., Adrian, T. E., Yanaihara, N.,
Yanaihara, C., and Bloom, S. R. The effect of the mammalian neuropeptide, gastrin- releasing peptide (GRP) on gastrointestinal and pancreatic hormone secretion in man. Clin. Sci. (Lond.), 65: 365-371, 1983.
13. Lehy, T., Puccia, E, Chariot, J., and Labeille, D. Stimulating effect of bombesin on the growth of gastrointestinal tract and pancreas suckling rats. Gastroenterology, 90: 1942-1949, 1986.
14. Rozengurt, E., and Sinnet-Smith, J. Bombesin stimulation of DNA synthesis and cell division in cultures of Swiss 3T3 cells. Proc. Natl. Acad. Sci. USA, 80: 2936--2940, 1983.
15. Cuttitta, F., Carney, D. N., Mulshine, J., Moody, T. W., Fedorko, J., Fishier, A., and Minna, J. D. Bombesin-like peptides can function as autocrine growth factors in human small-cell lung cancer. Nature (Lond.), 316: 823-826, 1985.
16. Yano, T., Pinski, J., Groot, K., and Schally, A. V. Sitmulation by bombesin and inhibition by bombesin/gastrin-releasing peptide antagonist RC-3095 of growth of human breast cancer cell lines. Cancer Res., 52: 4545--4547, 1992.
17. Narayan, S., Guo, Y-S., Townsend, C. M., and Singh, P. Specific binding and growth effects of bombesin-related peptides on mouse colon cancer cells in vitro. Cancer Res., 50: 6772-6778, 1990.
18. Frucht, H., Gazdar, A. F., Park, J-A., Oie, H., and Jensen, R. T. Characterization of functional receptors for gastrointestinal hormones on human colon cancer cells. Cancer Res., 52: 1114-1122, 1992.
19. Avis, F. P., Maneckjee, R., Cuttitta, E, Nakanishi, Y., Mulshine, J., and Avis, I. The role of gastrin releasing peptide in a pancreatic tumor cell line (CAPAN). Proc. Am. Assoc. Cancer Res., 29: 54, 1988.
20. Lhoste, E. F., and Longnecker, D. S. Effects of bombesin and caerulein on early stages of carcinogenesis induced by azaserine in the rat pancreas. Cancer Res., 47: 3272- 3277, 1987.
21. Douglas, B. R., Woutersen, R. A., Jansen, J. B. M., deJong, A. J. L., Rovati, L. C., and Lamers, C. B. H. W. Influence of cholecystokinin antagonist on the effects of cho- lecystokinin and bombesin on azaserine induced lesions on rat pancreas. Gastroen- terology, 96: 462--469, 1989.
22. Hajri, A., Balboni, G., Koening, M., Garaud, J-C., and Damge, C. Gastrin-releasing peptide: in vivo and in vitro growth effects on an acinar pancreatic carcinoma. Cancer Res., 52: 3726--3732, 1992.
23. Moody, T. W., Bertness, V., and Carney, D. N. Bombesin-like peptides and receptors in human tumor cell lines. Peptides, 4: 683--686, 1983.
24. Radulovic, S., Cai, R-Z., Serfozo, P., Groot, K., Redding, T. W., Pinski, J., and Schally, A. V. Biological effects and receptor binding affinities of new pseudonona- peptide bombesin/GRP receptor antagonist with N-terminal D-Trp or 9-Tpi. Int. J. Pept. Protein Res., 38: 591-600, 1991.
25. Cai, R-Z., Radulovic, S., Pinski, J., Nagy, A., Redding, T., Olsen, D., and Schally, A. V. Pseudononapeptide bombesin antagonists containing C-terminal Trp or Tpi. Pep- tides, 13: 267-271, 1992.
26. Radulovic, S., Miller, G., and Schally, A. V. Inhibition of growth of HT-29 human colon cancer xenografls in nude mice by treatment with bombesin/gastrin releasing peptide antagonist (RC-3095). Cancer Res., 51: 6006--6009, 1991.
27. Milovanovic, S. R., Radulovic, S., Groot, K., and Schally, A. V. Inhibition of growth of PC-82 human prostate cancer line xenografts in nude mice by bombesin antagonist RC-3095 or combination of agonist [n-Trp6]-luteinizing hormone-releasing hormone and somatostatin analog RC-160. Prostate, 20: 269-280, 1992
28. Pinski, J., Halmos, G., Yano, T., Szepeshazi, K., Qin, Y., Ertl, T., and Schally, A. V. Inhibition of growth of MKN45 human gastric adenocarcinoma exnografts in nude
mice by treatment with bombesin/gastrin releasing peptide antagonist (RC-3095) and somatostatin analogue RC-160. Int. J. Cancer, in press, 1994.
29. Szepeshazi, K., Schally, A. V., Halmos, G., Groot, K., and Radulovic, S. Growth inhibition of estrogen dependent and estrogen-independent MXT mammary cancers in mice by bombesin and gastrin-releasing peptide antagonist RC-3095. J. Natl. Cancer Inst., 84: 1915-1922, 1992.
30. Szepeshazi, K., Schally, A. V., Cai, R-Z., Radulovic, S., Milovanovic, S., and Szoke, B. Inhibitory effect of bombesin/GRP antagonist RC-3095 and high dose of somat- ostatin analogue RC-160 on nitrosamine-induced pancreatic cancers in hamsters. Cancer Res., 51: 5980-5986, 1991.
31. Redding, T. W., Schally, A. V., Tice, T. R., and Meyers, W. E. Long-acting delivery systems for peptides: inhibition of rat prostate tumors by controlled release of [t~-Trp6]luteinizing hormone-releasing hormone from injectable microcapsule. Proc. Natl. Acad. Sci. USA, 81: 5845-5848, 1984.
32. Schoumacher, R. A., Ram, J., lannuzzi, M. C., Bradbury, N. A., Wallace, R. W., Hon, C. T., Kelly, D. R., Schmid, S. M., Gelder, E B., Rado, T. A., and Frizzell, R. A. A cystic fibrosis pancreatic adenocarcinoma cell line. Proc. Natl. Acad. Sci. USA, 87: 4012--4016, 1990.
33. Qin, Y., Schally, A. V., and Wiilems, G. Somatostatin analogue RC-160 inhibits the growth of transplanted colon cancer in rats. Int. J. Cancer, 47: 765-770, 1991.
34. Ball, C. R., and Double, J. A. Transplantable colon tumors as chemotherapy screening models. Cancer (Phila.), 36: 2437-2440, 1975.
35. Labara, C., and Paigen, K. A simple, rapid and sensitive DNA assay procedure. Anal. Biochem., 102: 344-352, 1979.
36. Qin, Y., Schally, A. V., and Willems, G. Treatment of liver metastases of human colon cancers in nude mice with somatostatin analogue RC-160. Int. J. Cancer, 52: 791-796, 1992.
37. Bradford, M. A rapid and sensitive method for the quantitation of microgram quan- tities of protein using the principle of protein-dye binding. Anal. Biochem., 72: 248-254, 1976.
38. Qin, Y., van Cautern, M., Osteaux, M., Schally, A. V., and WiUems, G. Inhibitory effect of somatostatin analogue RC-160 on the growth of hepatic metastases of colon cancer in rats: a study with magnetic resonance imaging. Cancer Res., 52: 6025-6030, 1992.
39. Cai, R-Z., Reile, H., and Schally, A. V. Potent bombesin antagonists with a C-terminal Leu13a.I~(CH2N)Tac14-NH2. Int. J. Peptide Protein Res., in press, 1994.
40. McPherson, G. A. Analysis of radioligand binding experiments: a collection of com- puter programs for the IBM PC. J. Pharmacol. Methods, 14: 213-228, 1985.
41. Meijers, M., Graderen-Hoetmer, A. V., Lamers, C. B. H. W., Rovati, L. C., Jensen, J. B. M. J., and Voutersen, R. A. Effects of bombesin on the development of N-nitro- sobis(2-oxopropyl)amine-induced pancreatic lesions in hamsters. Cancer Lett., 59: 45-50, 1991.
42. Alexander, R. W., Upp, J. R., Jr., Poston, C. J., Townsend, C. M., Jr., Singh, E, and Thompson, J. C. Bombesin inhibits growth of human pancreatic adenocarcinoma in nude mice. Pancreas, 3: 297-302, 1988.
43. Lhoste, E., Aprahamian, M., Pousse, A., Hoeltzel, A., and Stock-Damge, C. Trophic effect of bombesin on the rat pancreas: is it mediated by the release of gastrin or cholecystokinin? Peptides, 6 (Suppl. 3): 89-95, 1985.
44. Szepeshazi, K., Schally, A. V., Groot, K., and Halmos, G. Effect of bombesin, gastrin-releasing peptide (GRP) (14-27) and bombesin/GRP receptor antagonist RC- 3095 on growth of nitrosamine-induced pancreatic cancers in hamsters. Int. J. Cancer, 54: 282-289, 1993.
45. Radulovic, S., Comaru-Schally, A. M., Milovanovic, S., and Schally, A. V. Somato- statin analogue RC-160 and LH-RH antagonist SB-75 inhibit growth of MIA PaCa-2 human pancreatic cancer xenografts in nude mice. Pancreas, 8: 88-97, 1993.
46. Zachary, I., Sinnett-Smith, J. W., and Rozengurt, E. Early events elicited by bombesin and structurally related peptides in quiescent Swiss 3T3 cells. I. Activation of protein kinase C and inhibition of epidermal growth factor binding. J. Cell Biol., 102: 221-222, 1986.
47. Liebow, C., Lee, M. T., Krebs, L. J., and Schally, A. V. Bombesin may stimulate growth through up-regulation of EGF receptors. Pancreas, 7: 746, 1992.
1041
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from

1994;54:1035-1041. Cancer Res Yunfeng Qin, Tibor Ertl, Ren-Zhi Cai, et al. in Vitro
and in Vivoon the Growth of Human Pancreatic Cancer Cells Inhibitory Effect of Bombesin Receptor Antagonist RC-3095
Updated version
http://cancerres.aacrjournals.org/content/54/4/1035
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/54/4/1035To request permission to re-use all or part of this article, use this link
Research. on November 22, 2018. © 1994 American Association for Cancercancerres.aacrjournals.org Downloaded from
![Humanlung small-cell carcinomacontains bombesin · methodofGrimelius (29) onBouin's fixed paraffin sections. Radioimmunoassays(RIAs). [Tyr8]Bombesin (donatedbyJ. E. Rivier, the Salk](https://img.pdfslide.net/doc/110x75/5f4a8f32d06af4400036e022/humanlung-small-cell-carcinomacontains-bombesin-methodofgrimelius-29-onbouins.jpg)




























