Embed Size (px)
Citation preview

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 226, 900–905 (1996)ARTICLE NO. 1447
Isolation of a New Mouse cDNA Clone: Hybrid Form of CytochromeP450 2b10 and NADPH-Cytochrome P450 Oxidoreductase
Marie Damon,*,† Alain Fautrel,* Nathalie Marc,* Andre Guillouzo,* and Laurent Corcos*,1
*Groupe Detoxication et Reparation Tissulaire, Faculte de Pharmacie, Universite de Rennes I, 2, Avenue du PrLeon Bernard, 35043 Rennes Cedex, France †INRA, Station de Recherche Porcine, 35590 St Gilles, France
Received August 9, 1996
We have isolated a novel cDNA sequence from a male mouse liver library. It consists of an open readingframe containing no stop codon of 1348 bp which share 98 % similarity with the cytochrome P450 2b10followed by 95 nucleotides sharing 88 % identity with the mouse NADPH cytochrome P450 oxidoreductase.Proteins are encoded by the opposite DNA strands. Genomic amplification excludes a library artefact or ade novo recombination event. RT-PCR analyses revealed expression of this clone in the liver of male andfemale mice as well as in kidney, lung and intestine. This cDNA was not, however, induced by phenobarbital.Preliminary results indicate that this cDNA could span more than 2.5 kb. This study is the first descriptionof a cytochrome P450 cDNA sequence encoding part of both a cytochrome P450 and the NADPH cyto-chrome P450 oxidoreductase in mammalian liver. q 1996 Academic Press, Inc.
Cytochrome P450 (Cyp) constitute a large superfamily containing at least 481 genes foundin bacteria, yeast, insects and mammals, which prove that this family is very ancient (1). Thesemonooxygenases play a critical role in the detoxication or sometimes activation of exogenouscompounds, and also in the metabolism of endogenous substances like steroids, fatty andbile acids (2). In most eukaryotic cells, Cyp-dependent monooxygenation requires anothermicrosomal protein, namely the NADPH cytochrome P450 oxidoreductase (P450 reductase).This protein is involved in transfer of electrons from NADPH to the heme of the Cyp and, incontrast to the multitude of Cyps, the P450 reductase is encoded by only one gene located onmouse chromosome 6 (3). Some data suggest that this reductase could be rate limiting in theoxidative process (4). Another system is found in the mitochondria of mammalian cells andin bacteria; it requires one more protein (redoxin) which transfers electrons from the reductaseto the Cyp. To date, there is only one known example of a one-protein system, which wasisolated from Bacillus megaterium (5).
The liver is the main organ involved in xenobiotic metabolism, but many Cyps are alsoexpressed in extrahepatic tissues such as lung, kidney and intestine (6). This tissue-specificexpression is regulated by some specific transcription factors (7). Moreover, Cyps have beenshown to be developmentally and hormonally regulated (2, 8, 9).
It is now well accepted that there are several distinct classes of Cyp inducers (polycyclicaromatic hydrocarbons, phenobarbital (PB), glucocorticoids, ethanol and clofibrate are theprototypes) that activate expression of one or more Cyp genes in distinct families and subfami-lies. Amongst these different regulatory processes, much remains to be uncovered about thePB mechanism of action (10). In the mouse, PB induces Cyps from family 2a, 2b, 2c and 3a(11). The Cyp 2b subfamily is one of the largest with at least 10 members but, until now,
1 Corresponding author. Fax number : 99. 33. 62. 42.Abbreviations used: Cyp, cytochrome P450 ; P450 reductase, NADPH cytochrome P450 oxidoreductase ; PB,
phenobarbital ; RT, reverse transcription ; PCR, polymerase chain reaction ; bp, base pair.
0006-291X/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
900
AID BBRC 5391 / 690a$$$261 09-06-96 13:47:51 bbrcas AP: BBRC

Vol. 226, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
only two genes, namely Cyp 2b9 and Cyp 2b10, have been cloned in the mouse and arelocated on chromosome 7 (12, 13).
In the present study, we have isolated a cDNA clone from a mouse liver cDNA library,which coding sequence is homologous to both Cyp 2b10 and P450 reductase, and studied itsexpression in hepatic and extrahepatic tissues as well as its regulation by PB in the liver.
METHODS
Animals. Male and female Balb/cByJ and male C57BL/6J mice were obtained from CERJ (Le Genest ST-ISLE,France). The animals were housed in plastic cages on wood chip bedding (U.A.R., Villemoisson-sur-Orge, France)and fed AO4 (U.A.R., Villemoisson-sur-Orge, France) and Wayne Breeder Box (Continental Grain Co., Chicago, IL)food ad libitum. At ages ranging between 8 and 12 weeks, mice were treated with one intraperitoneal injection of PB(GDH, Poole,UK) in 0.9 % NaCl, 18 hours before sacrifice. Control animals received vehicle alone. Animals weresacrificed by cervical dislocation and liver, kidney, lung and intestine samples were rapidly removed and frozen inliquid nitrogen. All experiments involving animals were done in compliance with French laws and regulations.
cDNA isolation. cDNA clones were isolated from a commercial liver cDNA library of a male Balb/cByJ mouseconstructed in the lgt11 vector (CLONTECH Laboratories, Palo Alto, CA). The screening was made by the plaquehybridization method as recommended by the supplier, using a 1250 bp rat cDNA probe corresponding to the 5*-endof the coding region of CYP 2B1. cDNAs were further subcloned in the PGem3zf(/)plasmid (Promega, Madison,WI) and analyzed by restriction enzyme digestion and hybridization with oligonucleotides specific for Cyp 2b9 andCyp 2b10 (14).
Southern blotting. About 2 mg of plasmid were digested with EcoRI (Eurogentec, Seraing, Belgium). The samplewas then subjected to electrophoresis in 1 % (w/v) agarose gel, denatured for 5 minin 0.25 N HCl, transferred in 0.4N NaOH onto Hybond-N/ nylon filters (Amersham, Arlington Heights, IL) and fixed with a UV crosslinker (Amersham,Arlington Heights, IL). Prehybridization, hybridization and washes were performed according to Church and Gilbert(15). cDNA probes were 32P-labeled by random priming using a T7 DNA polymerase random priming kit (Amersham,Arlington Heights, IL). The oligonucleotides were end-labeled using T4 polynucleotide kinase (Eurogentec, Seraing,Belgium). Filters were then exposed to autoradiography at 0807C. Probes used in this study were full length ratcDNAs for P450 reductase and CYP 2B1.
Sequencing. Double stranded cDNA was in part sequenced by the dideoxy method (16) with Sequenase Ver.2(Amersham, Arlington Heights, IL) and also carried out at Genome Express (Grenoble, France). The sequencingreaction was performed by PCR amplification in a final volume of 20 ml using 100 ng of PCR products, 5 pmoles ofprimer and 9.5 ml of dyeterminators premix according to Applied Biosystems protocol. After heating to 947C for 2min, the reaction was cycled as follows : 25 cycles of 30 s at 947C, 30 s at 557C and 4 min at 607C. Removal ofexcess of dyeterminators was performed using Quick spin columns (Boehringer, Manheim). The samples were driedin a vacuum centrifuge and dissolved in 4 ml of deionized formamide, EDTA pH 8. The samples were loaded ontoan Applied Biosystems 373A sequencer and run for 12 h on a 6% denaturing acrylamide gel. The nucleotide sequencewas then compared with sequences present in the Genbank database using BLAST network service at the NationalCenter for Biotechnology Information (NCBI).
RT-PCR analyses. Total liver RNA was extracted by the one step guanidinium-thiocyanate-phenol-chloroformextraction method according to Chomezynski and Sacchi (17). Oligonucleotide primers (20 bp) derived from thecDNA sequences of mouse Cyp 2b10, P450 reductase and b-actin were synthesized. Base sequences of the primerswere chosen to encompass the junction between Cyp 2b10 and P450 reductase. In order to obtain specific annealingwith the mRNA, reverse transcription (RT) was performed using the P450 reductase oligonucleotide. Specific cDNAfor mouse mRNA was obtained from 1 to 2 mg of total RNA under incubation conditions recommended by thesupplier of the cDNA synthesis kit (Gibco BRL, Life Technologies, Cergy Pontoise, France). Subsequently, PCR wasperformed in 100 ml of PCR buffer : 20 mM Tris-HCl, 50 mM KCl, 300 nM of each primer, 1 mM MgCl2 , 125 mMdNTPs and 0.5 U Goldstar DNA polymerase (Eurogentec, Seraing, Belgium). Samples were subjected to 30 cyclesof amplification : 967C for 1 min, 55 7C for 50 sec and 727C for 1 min (for the new clone). Cyp 2b10 and b-actinwere amplified using the primers and methods described previously (18). Aliquots (20 ml) of the PCR reactions weresubjected to electrophoresis (1.5 % agarose gel) and products were visualized by ethidium bromide staining. Gelswere blotted onto Hybond-N/ filters (Amersham, Arlington Heights, IL) and subjected to hybridization with the newcDNA, rat b-actin or CYP 2B1 cDNA probes. After autoradiography, signals were analyzed by densitometry usingan imaging software (Microvision Instruments, Evry, France). A PCR product of the new clone was analyzed by NcoI(Eurogentec, Seraing, Belgium) digestion and led to 2 bands of 62 and 72 bp, as expected from the restriction mapof our clone. The hybrid PCR product was also sequenced and found to be 100 % identical with our cDNA sequence(data not shown).
901
AID BBRC 5391 / 690a$$$262 09-06-96 13:47:51 bbrcas AP: BBRC

902
AID BBRC 5391 / 690a$$5391 09-06-96 13:47:51 bbrcas AP: BBRC

Vol. 226, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
TABLE 1Homology of Nucleotide and Amino Acid Sequences between p24
and Cyp 2b10, Cyp 2b9 and P450 Reductase, in the Mouse
Cyp 2b10 Cyp2b9 RED
Nucleotide 98% 87% 88%Amino acid 96% 83% 88%Position in p24 sequence /1 to /1348 bp /1 to /1348 bp /1349 to /1443 bp
RESULTS AND DISCUSSION
Isolation and Structure of a New P450
In a previous report, 21 cDNA clones for mouse Cyp 2b were isolated from a cDNA libraryconstructed from Balb/cByJ female liver mRNA (14). This library screening was performedusing a rat CYP 2B2 cDNA probe. Full length nucleotide sequences were obtained for twotypes of cDNAs now known as Cyp 2b9 and Cyp 2b10 (1). In the present study, in order toget new insights into this Cyp 2b subfamily, which most probably contains more than 2members, we have screened a male liver cDNA library with a rat CYP 2B1 cDNA probe.Twenty two independent clones were obtained from approximately 106 plaques. Amongstthem, we identified Cyp 2b9 and Cyp 2b10 cDNAs, but also some interesting new ones. Oneclone, called p24, hybridized with two Cyp 2b10 oligonucleotides, one from the 5* end, andanother from the middle of this cDNA, but not at all with a more distal (3*-end) oligonucleotide.This observation was unexpected since, being 1.4 kb-long, this clone should encompass this3*-region. It was then sequenced using the strategy shown in Fig.1. Surprisingly, p24 wasshown to contain 1348 nucleotides which were 98 % homologous to Cyp 2b10 cDNA (14)and, just downstream, 95 bp which were 88 % homologous to mouse P450 reductase (19)(Fig. 1 and Table 1). This result was confirmed by hybridization with a rat P450 reductasecDNA probe (data not shown). The sequence of the P450 reductase found in the p24 clonebelongs to the flavin binding domain (20). Moreover, the reductase sequence of this clone wasout of phase and antisens of the Cyp sequence. This new nucleotide sequence also showedconsiderable similarity (87%) with Cyp 2b9 (Table 1).
These structural features were really unexpected since, in mammals, Cyp 2b and P450reductase mRNAs are transcribed from distinct genes, located on chomosomes 7 and 6, respec-tively. In bacteria, there is one exception found in Bacillus megaterium, in CYP 102, whichconsists of a soluble single polypeptide chain that combines both the electron transport and thesubstrate oxidation functions (5). Moreover, this protein is structurally much more analogous toliver microsomal Cyps than to any other bacterial Cyp characterized to date. The p24 clonecould be related to this bacterial Cyp, but it could also be a more recent form, produced bysome recombination event between Cyp 2b and P450 reductase genes. In fact, the P450reductase gene itself has been proposed to be the result of a fusion of two different genesencoding flavin-binding proteins (20).
FIG. 1. Nucleotide sequence of p24 compared with mouse Cyp 2b10, Cyp 2b9 and P450 reductase sequences. (A)Sequencing strategy. (B) The 1442 bp of the nucleotide sequence of p24 are shown. The different nucleotides fromp24 are underwritten for Cyp 2b10, Cyp 2b9 and P450 reductase. The initiation codon is boxed. The arrows indicatethe antisens P450 reductase coding sequence of p24. The broken line indicates deletions in these sequences. Theoligonucleotides used in this study for hybridizations and PCR analyses appear as bold faces and doubly underlined.
903
AID BBRC 5391 / 690a$$$262 09-06-96 13:47:51 bbrcas AP: BBRC
Vol. 226, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
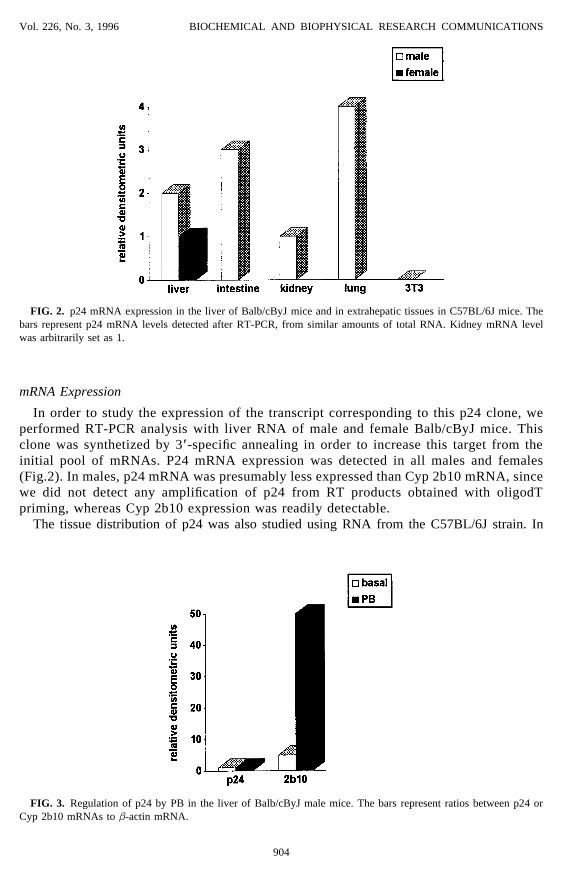
FIG. 2. p24 mRNA expression in the liver of Balb/cByJ mice and in extrahepatic tissues in C57BL/6J mice. Thebars represent p24 mRNA levels detected after RT-PCR, from similar amounts of total RNA. Kidney mRNA levelwas arbitrarily set as 1.
mRNA Expression
In order to study the expression of the transcript corresponding to this p24 clone, weperformed RT-PCR analysis with liver RNA of male and female Balb/cByJ mice. Thisclone was synthetized by 3*-specific annealing in order to increase this target from theinitial pool of mRNAs. P24 mRNA expression was detected in all males and females(Fig.2). In males, p24 mRNA was presumably less expressed than Cyp 2b10 mRNA, sincewe did not detect any amplification of p24 from RT products obtained with oligodTpriming, whereas Cyp 2b10 expression was readily detectable.
The tissue distribution of p24 was also studied using RNA from the C57BL/6J strain. In
FIG. 3. Regulation of p24 by PB in the liver of Balb/cByJ male mice. The bars represent ratios between p24 orCyp 2b10 mRNAs to b-actin mRNA.
904
AID BBRC 5391 / 690a$$$262 09-06-96 13:47:51 bbrcas AP: BBRC

Vol. 226, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
all the tissues examined, i.e liver (data not shown), lung, kidney and intestine, the p24 mRNAwas expressed (Fig.2). P24 was not, however, detected in a mouse fibroblastic cell line (3T3).
These PCR products were specific for mRNA amplification, since we did not observe anyamplification from RNAse-treated samples. Moreover, this novel Cyp form corresponds to agene structure, since we also obtained a PCR product from genomic DNA (data not shown).
Taken together, these results lead to the conclusion that this clone really exists in the mouse.
Regulation of This Atypical mRNA by PB
Since Cyp 2b10 is induced by PB in the mouse (18, 21), we decided to examine the PBinducibility of p24 mRNA. In Balb/ cByJ male mice, we did not see any effect of PB treatmenton this mRNA, although Cyp 2b10 was induced (Fig.3). This observation confirmed that Cyp2b10 and p24 were two distinct forms of Cyps.
In conclusion, we have isolated a novel Cyp of atypical structure, expressed at the mRNAlevel in the liver and in several extrahepatic tissues. Preliminary results indicate that this clonecould span at least 2.5 kb. We are now adressing the challenging possibility that this clonecould indeed encode a functional protein by looking for the 3*-end of the mRNA as an initialstep for expressing the full length version in yeast cells.
ACKNOWLEDGMENTSWe thank V. Lecureur for providing 3T3 RNA samples and F. Morel for critical reading of the manuscript. This
work was supported by the INSERM and the Association pour la Recherche contre le Cancer. M.D. was supportedby the Institut National de la Recherche Agronomique.
REFERENCES1. Nelson, D. R., Koymans, L., Kamataki, T., Stegeman, J. J., Feyereisen, R., Waxman, M. R., Gotoh, O., Coon,
M. J., Estabrook, R. W., Gunsalus, I. C., and Nebert, D. W. (1996) Pharmacogenetics 6, 1–42.2. Gonzalez, F. J. (1989) Pharmacol. Rev. 40, 243–288.3. Simmons, D. L., Lalley, P. A., and Kasper, C. B. (1985) J. Biol. Chem. 260, 515–521.4. Dong, J., and Porter, T. D. (1996) Arch. Biochem. Biophys. 327, 254–259.5. Fulco, A. J. (1991) Annu. Rev. Pharmacol. Toxicol. 31, 177–203.6. Gonzalez, F. J., Liu, S. L., and Yano, M. (1993) Pharmacogenetics 3, 51–57.7. Yano, M., Falvey, E., and Gonzalez, F. J. (1992) Mol. Cell. Biol. 12, 2847–2854.8. Larsen, M. C., and Jefcoate, C. R. (1995) Arch. Biochem. Biophys. 321, 466–476.9. Lund, J., Zaphiropoulos, P. G., Mode, A., Warner, M., and Gustafsson, J. A. (1991) Adv. Pharmacol. 22, 325–
354.10. Waxman, D. J., and Azaroff, L. (1992) Biochem. J. 281, 577–592.11. Corcos, L. (1992) Drug. Metab. Dispos. 20, 797–801.12. Lakso, M., Masaki, R., Noshiro, M., and Negishi, M. (1991) Eur. J. Biochem. 195, 477–486.13. Honkakoski, P., Moore, R., Gynther, J., and Negishi, M. (1996) J. Biol. Chem. 271, 9746–9753.14. Noshiro, M., Lakso, M., Kawajiri, K., and Negishi, M. (1988) Biochemistry 27, 6434–6443.15. Church, G. M., and Gilbert, W. (1984) Proc. Natl. Acad. Sci. U.S.A. 81, 1991–1995.16. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl. Acad. Sci. USA 74, 5463–5467.17. Chomezynski, P., and Sacchi, N. (1987) Anal. Biochem. 162, 156–159.18. Damon, M., Fautrel, A., Guillouzo, A., and Corcos, L. (1996) Biochem. J. 317, 481–486.19. Ohgiya, S., Shinriki, N., Kamataki, T., and Ishizaki, K. (1994) Biochim. Biophys. Acta 1186, 137–141.20. Porter, T. D., and Kasper, C. B. (1985) Proc. Natl. Acad. Sci. USA 82, 973–977.21. Honkakoski, P., Kojo, A., and Lang, M. A. (1992) Biochem. J. 285, 979–983.
905
AID BBRC 5391 / 690a$$$262 09-06-96 13:47:51 bbrcas AP: BBRC





























