Embed Size (px)
Citation preview
18 4 P R E L I M I N A R Y N O T E S
PN 61o24 Isolement d'un complexe enzyme d'~change A D P - A T P - cytochrome c
r~dult
La r6action d'6change ADP-ATP est une 6tape terminale du processus de couplage en oxydation phosphorylantO& L'enzyme qui catalyse cette r6action a ~t6 purifi6 par GLAZE ET WADKINS 3. Une des caract6ristiques principales de cet enzyme est que ses propri&6s physio-chimiques sont modifi6es par l'addition de cytochrome c r6duit a; en particulier, l'inhibition de l'6change ADP-ATP caus6e par la pr6sence d'un exc+s d 'ADP dimintte si du cytochrome c r~duit est ajout~ au milieu d'incubation.
Nous avons r6cemment isol6, ~ partir de mitochondries de coeur de boeuf, une fraction prot6ique homog~ne qui correspond apparemment /t une combinaison de l 'enzyme d'6change ADP-ATP avec le cytochrome c r6duit. Cette note pr61iminaire porte sur l'isolement et les propri6t~s du complexe enzyme d'6change A D P - A T P - cytochrome c r6duit.
Les mitochondries de coeur de boeuf ont 6t6 pr~par6es selon CRANE et co113. Un extrait mitochondrial dont la pr6paration et la purification seront d6crites en d6tail ult6rieurement est chromatographi6 sur colonne de DEAL-cellulose ~quilibr6e avec un tampon Tris-HC1 0.025 M (pH 8.5). L'~lution des fractions prot~iques est r6alis6e par le passage successif sur la colonne d'une s6rie de tampons Tris dont le pH d6croit et dont la molarit6 crolt. Un absorptiom&tre Uvicord 47olA LKB permet l'enregistrement automatique du diagramme d'61ution. Le passage d'un tampon Tris o.I M (pH 7.5) entralne l'dlution d'une fraction de coloration rose (fraction
0.5
~¢mm
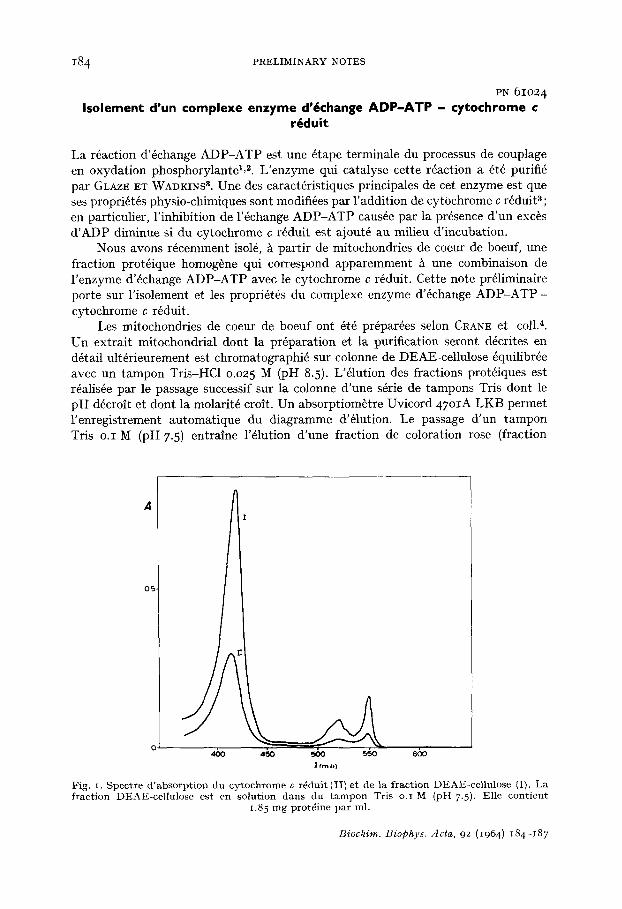
Fig. t. Spectre &absorption du cytochrome c r6duit (II) et de la fraction DEAL-cellulose (I). La fraction DEAE-ce]lulose est en solution dans du tampon Tris o.I M (pH 7.5). Elle contient
1.85 mg protdine par ml.
Biochim. Biophys. Acta, 92 (1964) I84-I87
PRELIMINARY NOTES I 8 5
DEAE-cellulose). L'~lectrophor~se de cette fraction en g61ose ~ pH 8.2 montre l 'examen direct et apr~s coloration par une solution de vert Lissamine qu'il s'agit d'une prot6ine migrant d'une fa~on homog~ne et poss~dant une mobilit6 cathodique distincte du cytochrome c r6duit (sa mobilit6 est interm6diaire entre celle des flA- globulines et celle des ~,-globulines du s6rum humain normal).
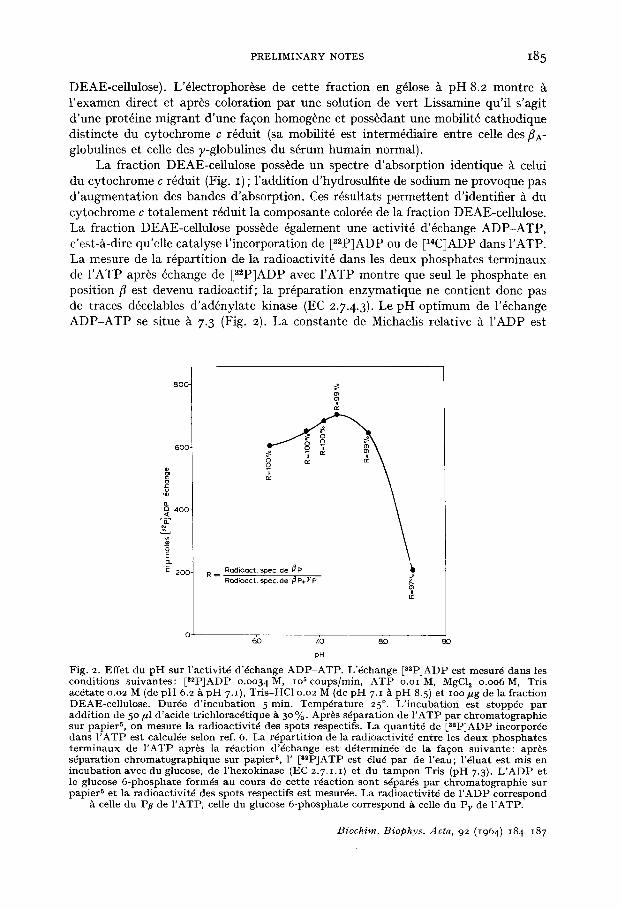
La fraction DEAE-cellulose poss~de un spectre d'absorption identique ~t celui du cytochrome c r6duit (Fig. I) ; l 'addition d'hydrosulfite de sodium ne provoque pas d'augmentation des bandes d'absorption. Ces r6sultats permettent d'identifier h du cytochrome c totalement r6duit la composante color6e de la fraction DEAE-cellulose. La fraction DEAE-cellulose poss~de 6galement une activit6 d'6change ADP-ATP, c'est-/~-dire qu'elle catalyse l'incorporation de [a~PJADP ou de ~CJADP dans I'ATP. La mesure de la r6partition de la radioactivit6 dans les deux phosphates terminaux de I 'ATP apr~s 6change de [s*p]ADP avec I'ATP montre que seul le phosphate en position fl est devenu radioactif; la pr6paration enzymatique ne contient donc pas de traces d6celables d'ad6nylate kinase (EC 2.7.4.3). Le pH optimum de l'6change ADP-ATP se situe ~ 7.3 (Fig. 2). La constante de Michaelis relative h I'ADP est
8 0 0
600
g 5 ,o
400- <
E 200- R ~ Radioact. spec. de ~ P Radioclct. spec. de /5 P+ ? P
6.0 ~0 8O 9O
pH
Fig. 2. Effe t du p H sur l ' ac t iv i t6 d '~change A D P - A T P . L 'dchange [3~P]ADP est mesur~ dans les condi t ions su ivan t e s : [s*P]ADP o .oo34M, ioScoups /min , A T P O.o lM, MgCI~ o .oo6M, Tris ac6 ta te o.o2 M (de p H 6.2 £ p H 7.I), Tr is -HC1 0.02 M (de p H 7.1 A p H 8.5) et ioo ~ug de la f rac t ion DEAE-cel lu lose . Durde d ' i n c u b a t i o n 5 min . T e m p d r a t u r e 25 °. L ' i n c u b a t i o n es t s toppde pa r add i t ion de 5 ° /z l d ' ac ide t r ich loracdt ique A 3 ° %. Apr~s sdpara t ion de I ' A T P pa r c h r o m a t o g r a p h i e sur pap ie r 5, on mesu re la radioact iv i t6 des spo t s respectifs . La quan t i t 6 de [32P]ADP incorpor~e dans I ' A T P est calculde selon ref. 6. La rdpar t i t ion de la radioact iv i td en t re les deux p h o s p h a t e s t e r m i n a u x de I ' A T P apr~s la rdact ion d '6change es t d6terminde de la fa~on su ivan t e : apr6s s~para t ion c h r o m a t o g r a p h i q u e sur papier 5, 1' [s2P]ATP est 6lug pa r de r e a u ; l '~ luat es t mis en i ncuba t ion avec du glucose, de l ' hexok inase (EC 2.7.1.1 ) et du t a m p o n Tris (pH 7.3). L ' A D P et le glucose 6 - p h o s p h a t e formds au cours de cet te r6act ion son t s6par6s pa r c h r o m a t o g r a p h i e su r pap ie r 5 et la radioact iv i td des spo t s respect i fs es t mesurde. La radioact iv i t6 de I ' A D P cor respond
celle du Pt~ de I 'ATP, celle du glucose 6 -phospha t e cor respond A celle du Pv de I 'ATP.
Biochim. Biophys. Acta, 92 (1964) I 8 4 - I 8 7

PRELIMINARY NOTES 187
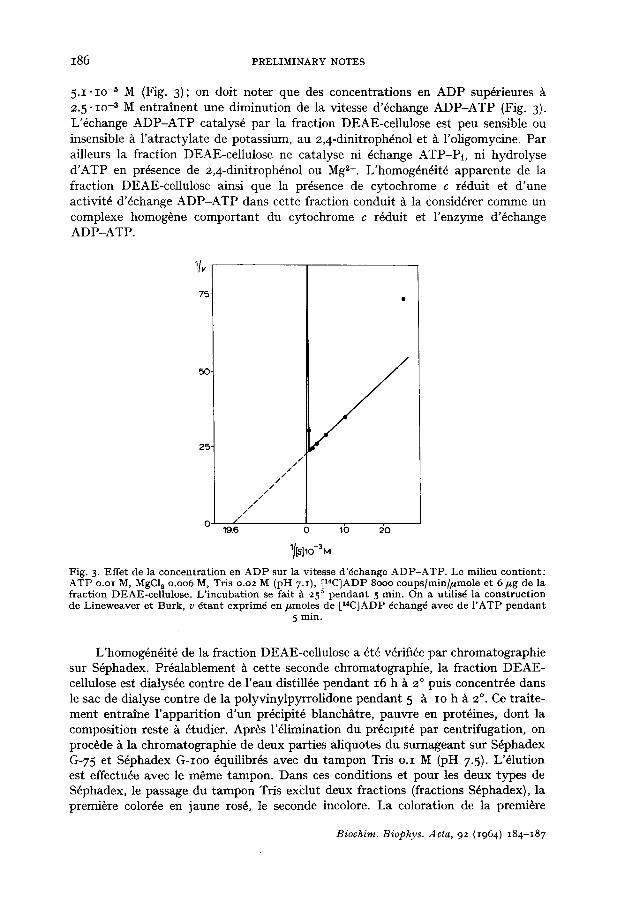
fraction est dfie A la pr6sence de cytochrome c r6duit, comme en t6moigne le spectre d'absorption. Les deux fractions S6phadex catalysent l'6change ADP-ATP, mais l 'activit6 sp6cifique de la fraction colorde est 3 ~t 4 fois plus grande que celle de la fraction incolord. D'autre part, les fractions color6es G-75 et G-Ioo migrent en 61ectrophor~se sur g61ose de fa~on homog~ne et ~t la m~me vitesse que la fraction DEAE-cellulose. Les fractions color6es (fraction DEAE-cellulose et fractions color6es S6phadex G-75 et G-Ioo) correspondent donc probablement A un m6me complexe homog&ne: cytochrome c r6duit - enzyme d'6change ADP-ATP. Cependant ce complexe semble dissociable puisqu'il lib~re apr~s dialyse l 'enzyme d'6change sous forme libre comme le montrent les chromatographies sur S6phadex et les mesures d'activit6 enzymatique. I1 est en fait possible que la stabilit6 de l'association entre le cytochrome c r6duit et l 'enzyme d'6change soit dfie ~t la participation d'un troisi&me constituant au complexe cytochrome c r6duit - enzyme d'6change; l'61imination de ce constituant (pr6cipit6 apparaissant au cours de la dialyse?) entrainerait une instabilit6 de l'association entre cytochrome c r6duit et enzyme d'6change.
En r~sum6, les r~sultats rapport~s dans cette note sugg~rent que l 'enzyme d'6change A D P - A T P existe dans les mitochondries de coeur de boeuf au moins en partie sous une forme combin6e: enzyme d'6change - cytochrome c r6duit.
Facultd de Mddecine et de Pharrnacie et Centre d'Etudes Nuddaires,
Grenoble (France)
j . LATURAZE
P. V. VIGNAIS
x C. L. WADKINS ETA. L. LEHNINGER, J. Biol. Chem., ~38 (1963) 2355. C. L. WADKINS ETA. L. LEHNINGER, Federation Proc. Syrup., 22 (1963) lO92.
3 R. P. GLAZE ETC. L. WADKINS, Biochem. Biophys. Res. Commun., 15 (1964) 194. 4 F. L. CRANE, J. L. GLENN ET D. E. GREEN, Biochim. Biophys. Acta, 22 (1956) 475. • ~ H. A. KREBS ET R. HEMS, Biochim. Biophys. Acta, 12 (1953) 172. s p. D. BOYER, R. C. MILLS ET H. J. FROMM, Arch. Biochem. Biophys., 81 (1959) 249.
Re~u le 22 Juillet, 196 4 Biochim. Biophys. Acta, 92 (1964) I84-187
PN 6103O On the role of free radicals in tryptophan pyrrolase catalysis
Tryptophan pyrrolase is an oxygenase which catalyzes the insertion of molecular oxygen into the pyrrole ring of t ryptophan yielding formylkynurenine 1. Previous studies2, ~ which have recently been confirmed 4 indicate that a dissociating metal- loporphyrin coenzyme participates in this catalytic process. Further studies have suggested that this catalysis may be separated into distinct processes in which t ryptophan pyrrolase catalyzes the reduction by t ryptophan of hematin (ferriproto- porphyrin IX) to heme (ferroprotophorphyrin IX) and the reoxidation of the heine to hematin by oxygen 5,6. That electron transfer occurs in the t ryptophan pyrrolase reaction has been further documented by the recent demonstration of a substrate and enzyme-dependent anaerobic reduction of dichlorophenolindophenol, methylene blue, or cytochrome-c (ref. 7). Electron transfer from t ryptophan to oxygen may
Biochim. Biophys. Acta, 92 (1964) 187-19o





























