Embed Size (px)
Citation preview

Abstract. Juvenile chronic myelogenous leukemia(JCML) is a hematologic malignancy of monocyte-macrophage lineage in which leukemic progressionis mediated in an autocrine manner by tumor necro-sis factor (TNF-α), GM-CSF and possibly othergrowth factors. Cytogenetic data showing involve-ment of both erythroid and monocyte-macrophagelineages in the JCML leukemic clone, as well as anobserved episode of B-lineage lymphoid blast crisisin JCML, has strengthened the thesis for a lympho-hematopoietic pluripotent stem cell origin for thedisorder. To study this further, JCML CD34+ cellsfrom bone marrow (BM) or spleen from six newlydiagnosed patients were isolated and cultured inclonogenic assays with combinations of recombi-nant cytokines. Compared to control CD34+ cells,JCML cells from all patients showed an aberrantgrowth pattern restricted almost exclusively to themonocyte-macrophage lineage. Most of the clono-genic activity was seen in a subsorted population ofCD34+, HLA-Dr– cells. Additionally, an exaggeratedgrowth response to minute doses of GM-CSF thathad no effect on control cells was observed withJCML CD34+ cells. Recloning (“self-renewal”) ofJCML CD34+ cells was also strongly promoted byGM-CSF. JCML colonies also formed spontaneouslyin the absence of exogenous cytokines but were aug-mented by GM-CSF, interleukin 1 and TNF-α, thelatter feature not seen with control CD34+ cells fromnormal BM. The abnormal spontaneous growth pat-tern of CD34+ JCML cells could be suppresseddirectly in vitro by anti-TNF-α antibodies and anti-GM-CSF antibodies alone or in combination, andby soluble TNF-α receptors (sTNF-R:Fc), consistent
with the notion that JCML CD34+ cells are stimu-lated by both cytokines in an autocrine manner. Inmalignant CD34+ cells from one patient, the cyto-genetic marker monosomy 7 proved leukemicinvolvement of monocyte-macrophage, erythroidand B-lymphoid lineages. We conclude that CD34+
JCML cells of multilineage potential exhibit exces-sive and aberrant monocyte-macrophage colony for-mation, a property that was previously observed inJCML progenitors found in light density cell frac-tions. Thus, within the CD34+ cellular compartmentis a subpopulation of JCML “stem” cells thataccounts for the abnormal leukemic proliferativeactivity in this disease. Stem Cells 1996;14:690-701
Introduction
In previous studies of patients with juve-nile chronic myelogenous leukemia (JCML), weobserved two reproducible abnormalities in vitrothat have become hallmarks of the disease:impaired growth of normal bone marrow (BM)and peripheral blood (PB) hematopoietic pro-genitors, and excessive proliferation of malig-nant clonal monocyte-macrophage elements inthe absence of an exogenous growth factor [1-8].
Further studies showed that JCML cellsendogenously produce tumor necrosis factor(TNF-α ) and GM-CSF that act as autocrinegrowth factors [9, 10]. Indeed, JCML cells showa selective hypersensitivity to the growth-pro-moting effect of GM-CSF compared to controls[11]. Data from Bagby et al. [12] also suggestedthat the abnormal proliferative activity of JCMLcells was dependent, in part, on the endogenousproduction of interleukin 1 (IL-1) which, in aparacrine manner, stimulated the release ofcolony-stimulating activity by other cells.
Correspondence: Dr. Melvin H. Freedman,Division of Hematology/Oncology, Hospital for SickChildren, 555 University Avenue, Toronto, Ontario,M5G 1X8, Canada.
Rece ived March 28 , 1996; accepted forpublication July 30, 1996. ©AlphaMed Press1066-5099/96/$5.00/0
STEM CELLS 1996;14:690-701
Juvenile Chronic Myelogenous Leukemia Multilineage CD34+ Cells: Aberrant Growth and Differentiation Properties
Melvin H. Freedman, Johann K. Hitzler, Nancy Bunin, Tom Grunberger, Jeremy SquireDivision of Hematology/Onocology, Department of Pediatrics, and Department of Pediatric Laboratory Medicine,Division of Pathology, Hospital for Sick Children, University of Toronto, Toronto, Canada; Division of Oncology,Children’s Hospital of Philadelphia, Philadelphia, USA
Key Words. Juvenile chronic myelogenous leukemia • CD34+ cells • Monocyte-macrophage colonies

All of these vitro studies were performedon heterogenous populations of BM or PBmononuclear cells, usually the light density ele-ments obtained following a Percoll or Ficoll-Hypaque fractionation step. Within these mixedpopulations of cells, there must be a subpopu-lation of JCML progenitors that accounts forthe abnormal proliferative properties [1-12] ofthe monocyte-macrophage progeny. Our goalis to identify and characterize these JCML pro-genitors in order to ask fundamental questionsabout the biology of this aggressive form ofleukemia. Since it was proposed [13, 14] andrecently confirmed [15] that JCML involvesmultiple hematopoietic lineages in a mannersimilar to Ph+ CML, we reasoned that the puta-tive JCML “stem” cells should be within thepluripotent CD34+ cellular compartment.
In support of this hypothesis, this reportdescribes the aberrant growth and differentia-tion characteristics of multilineage JCML CD34+
cells obtained from six patients with the disease.
Materials and Methods
SubjectsThe six previously unpublished patients in
this report fit the classical clinical and laboratorydescription of JCML [13, 14] and showed thecharacteristic in vitro hallmarks alluded to inthe introduction. They were designated aspatients 21-26 to correspond with our ongoingseries. Details on the other patients in the seriescan be found as follows: patients 1-9, [1-7];patient 10, [8]; patients 11-16, [9]; patient 17,[16]; and patients 18-20, [17].
All six patients had varying combinationsof lymphadenopathy, skin manifestations, variable
splenomegaly, pallor and hemorrhagic signs. Themain hematological findings were anemia, throm-bocytopenia, leukocytosis with a left shift andnucleated RBCs in the PB (Table 1). BM aspi-rates were cellular with granulocytic hyperpla-sia, active erythropoiesis and reduced to absentmegakaryocytes. BM specimens from all sixyielded high numbers of monocyte-macrophagecolonies in a colony-forming unit (CFU)-GMassay in the absence of exogenous growth factor,typical of JCML. Routine cytogenetics studies ofprimary cellular specimens and colonies after 14days in tissue culture were normal in five patients,but cells from patient 26 showed monosomy 7.For control studies, BM was obtained from hema-tologically normal donors for BM transplanta-tion during the harvest under anesthesia, and fromBM from adults with Ph+ CML in chronic phase.These specimens were obtained with informedconsent and were approved by the Human EthicsCommittee of our institution.
Preparation of CD34+ CellsPatients 21, 23, 25, and 26 underwent
splenectomy prior to BM transplantation.Splenic tissue was freshly obtained, minced andthen filtered. Heparinized BM, PB or spleencells were layered over Percoll (density 1.077g/ml; Pharmacia Fine Chemicals; Piscataway,NJ) and centrifuged (400 g) for 10 min at 4°C toremove neutrophils and RBCs.
Ceprate LC Laboratory Cell SeparationSystems (CellPro, Inc.; Bothwell, WA) wereused to isolate CD34+cells in the following way:the post-Percoll mononuclear cells were incu-bated with biotinylated anti-CD34 monoclonalIgM and passed through a column of avidin-coated beads [18-20]. The nonadsorbed cellswere washed through with phosphate-buffered
Freedman, Hitzler, Bunin et al. 691
Table 1. Clinical and laboratory data on six patients with JCML at diagnosis
Patient
21 22 23 24 25 26
Sex M M M M M MAge (months) 4 3 30 2 5 42Hemoglobin (G/dl) 7.9 9.9 8.2 8.4 8.8 8WBC (× 109/l) 124.3 28.8 71.7 43.9 193.1 99.0Platelets (× 109/l) 63.0 52.0 17.0 349.0 146.0 6.0PB monocytes (× 109/l) 14.9 3.3 1.0 13.6 20.0 40.0PB blasts (× 109/l) 9.9 1.4 1.0 3.0 18.0 13.0BM M:E ratio 5:1 5:1 10:1 5:1 5:4 12:1BM monocytes (%) 5 2 3 14 0 3BM blasts (%) 15 6 5 3 27 9

saline (PBS); the adsorbed cells were detachedby squeezing the column several times followedby washing again with PBS. The enriched CD34+
cell population was analyzed by phenotype usingcytofluorometry to assess the degree of purity.
Clonogenic AssaysTo demonstrate the effect of specific cytokines
on CD34+ cells, a modification of the CFU-gran-ulocyte/erythroid/macrophage/megakaryocyte(GEMM) clonogenic assay was performed aspreviously described [2, 3]. A population of 3 × 103 CD34+ cells (BM, PB or spleen) was cul-tured in 0.8% methylcellulose with Iscove’smodified Dulbeco’s medium (Ontario CancerInstitute; Toronto, Canada), plasma obtainedfrom a pool of five normal donors (for studiesshown in Tables 2 and 3), and 5 × 10–5 M of 2-mercaptoethanol. For clonogenic assays otherthan those shown in Tables 2 and 3, 30% fetalcalf serum was substituted for pooled humanplasma. One ml of culture mixture was placedin 35 mm Petri dishes and incubated with 5%CO2 in air in a humidified atmosphere.
All cultures were evaluated after 14 daysfor the number of BFU-E colonies (defined asan aggregate of more than 500 hemoglobinizedcells, or three or more erythroid subcolonies),CFU-megakaryocyte (MK) colonies (CFU-GEMM) containing all lineages and CFU-GMcolonies defined as a cluster of more than 40cells consisting of granulocytes, monocyte-macrophages, or both. For secondary and ter-tiary in vitro recloning experiments, colonieswere counted after 14 days of incubation, resus-pended and then replated in fresh semisolidmedium. Cultures in the presence and absenceof exogenous growth factor were processed inparallel.
Microscopically, pure granulocyticcolonies could be easily distinguished frompure macrophage colonies because of smallercells and looser clustering in the former.Individual colonies were plucked from the cul-tures with a micropipette and spread on glassslides, and the cellular composition was deter-mined microscopically using Wright’s stainand nonspecific (a-naphthyl butyrate) esterasestaining with and without treatment with flu-oride [21]. Staining with benzidine was used toconfirm intracellular hemoglobin in coloniesscored as BFU-E.
Fluorescence in Situ Hybridization (FISH)After recording the coordinates, Wright-
stained smears of individual BFU-E colonieswere briefly destrained in ethanol prior to eitherfixation for FISH or for staining with benzi-dine. CD19+ lymphoid cell populations wereprepared by fluorescence-activated cell sorting(FACStar, Becton Dickinson; San Jose, CA)by labeling CD34+ JCML spleen cells withfluorescein isothiocyanate (FITC)-conjugatedanti-CD19 according to manufacturer’sinstructions (Coulter; Hialeah, FL). ResultantCD19+ cell populations (purity of 95% to 98%)were used to prepare cytospin slides for fixationand FISH.
To identify cells with monosomy 7, a biotiny-lated probe (D7ZI; Oncor, Inc; Gaithersburg, MD),hybridizing with the centromeric region of humanchromosome 7 was used. Normal human lym-phocytes were used as controls. FISH was basedon published procedures [22]. After preparing theslides and hybridization mixture, hybridiza-tion was carried out for 16-20 h at 37°C. The slideswere washed, soaked in 3% bovine serum albu-min (BSA), and then incubated FITC (Oncor,Inc.). The slides were then washed and counter-stained with 0.6 µg/ml–1 propidium iodide in PBS).After washing in PBS for 5 min, the slides weremounted in 20 mM Tris-HC1, pH 8.0, 90% glyc-erol containing 2.3% of the DAPCO antifade, 1.4diazabicyclo-(2,2,2) octane. Photomicrographswere obtained using a Nikon Microphot-FXA epi-fluorescence microscope equipped with dual bandFITC/Texas red filters (Omega Optical Inc.;Montreal, Canada). Kodak color Ektachromep800/1600 E-6p professional film was used with acamera setting of ASA 1600.
Recombinant Human Cytokines and TheirRespective Antibodies
Human TNF-α (lot #N9030AX; Genentech;San Francisco, CA) with the specific activityof 5.6 × 107 U/mg was used in a concentration of 1× 103 U/ml. Monoclonal anti-TNF-α neutralizingantibodies (mAb-Hu TNF-α , lot #5890-90;Genentech) were purified from ascites fluid to aconcentration of 2 mg/ml, and had a neutralizingtitre of more than 5 × 105 neutralizing U/ml. Thesame lot number of mAb-Hu TNF-α was used inall of the studies. For our assays, anti-TNF-α anti-bodies were added directly to cell cultures, inwhich an exogenous source of growth factor orcytokine was added or omitted. In quality control
692 JCML Multilineage CD34+ Cells: Aberrant Growth and Differentiation Properties

studies, 2,000 U/ml for anti-TNF-α antibodiesneutralized 1,000 U of TNF-α, and hence 2,000U/ml were used in all of our assays. The efficiencyof neutralization of TNF-α by anti-TNF-α anti-bodies was documented in a TNF-α cytoxicityassay using L929 mouse fibrosarcoma cells [23].Soluble TNF receptor (p80), linked to the Fc por-tion of human IgG1 (rHu TNF-R:FC, lot #3353-090; Immunex; Seattle, WA) was used in aconcentration of 5 mg/ml to neutralize the in vitroeffect on TNF-α [24].
GM-CSF (Genetics Institute; Cambridge,MA) with a specific activity in a CML clono-genic assay of 10.6 × 106 U/mg was used in aconcentration of 5 U/ml. Anti-GM-CSF neu-tralizing antibodies (Genetics Institute) wereused in a concentration of 10 mg/ml. In qualitycontrol bioassay neutralization testing, 1 mg/mlblocked the activity of 5 U/ml of GM-CSF. Inour clonogenic assays, the anti-GM-CSF anti-bodies were added directly to the cell cultures.
IL-1α (Hoffmann-LaRoche; Nutley, NJ)with specific activity of 3 × 108 U/mg was usedin concentration of 100 U/ml. Rabbit poly-clonal antihuman IL-1 neutralizing antibodies(product LP-712; Genzyme; Boston, MA) wereused directly in cultures in a concentration of10 U/ml, which was effective in reducing pro-liferation of hematopoietic colonies from Ph+ CMLpatients. Erythropoietin (Eprex; Ortho BiologicalInc.; Raritan, NJ) with activity of 1 × 104 U/mlwas used in a concentration of 2 U/ml. Il-3 (lot#15168-138 B, Genetics Institute) with a spe-cific activity of 1.58 × 106 U/ml was used in aconcentration of 40 U/ml. MGF (Steel factor, c-kit ligand, lot #3913-035; Immunex) with aspecific activity of 1 × 105 ng/ml was used in aconcentration of 50 ng/ml.
Statistical AnalysisThe probability of a significant difference
between colony numbers was determined byStudent’s t-test.
Results
Enrichment of Marrow and Spleen Specimensfor CD34+ Cells
The Ceprate LC Laboratory Cell Separationcolumns were highly effective in fractionatingCD34+ cells from monouclear elements in post-Percoll mononuclear preparations. With normal
BM specimens (n = 7), the average yield of CD34+
cells was 1.6% of the total (range 0.5%-2.5%);the purity of the fractionation by flow cytometryvaried from 85% to 99%. With the JCML mar-row or spleen specimens (n = 6), the average yieldof CD34+ cells was 2.7% (range 1.5%-5%) with the same degree of purity. With Ph+ CML mar-row (n = 2), the average yield was 0.36% (range0.3%-0.4%).
To test the magnitude of enrichment ofclonogenic CD34+ cells by the column frac-tionation technique, normal marrow was cul-tured in a CFU-GEMM assay before and afterthe fractionation step. With the postcolumnCD34+ cell preparation, 3 × 103 cells platedyielded comparable colony numbers 2 × 105
precolumn, post-Precoll mononuclear cells(Table 2). Thus, there was approximately a two-log enrichment of clonogenic cells using thepostcolumn CD34+ cell preparation.
Abnormal Colony Formation by JCML CD34+ Cells
Table 3 shows the colony growth pattern ofJCML CD34+ cells in response to a cytokine“cocktail” of IL-3, GM-CSF, Steel factor and ery-thropoietin. Compared to CD34+ cells fraction-ated from the BM of healthy controls and a newlydiagnosed patient with Ph+ CML in chronic phase,the JCML colony numbers and cellular composi-tion of individual colonies were markedly andconsistently different for all six patients. JCMLBFU-E-derived colonies were reduced in numberand size, mixed colonies were absent, and
Freedman, Hitzler, Bunin et al. 693
Table 2. Comparison of colony numbers from nor-mal marrow before and after column fractionationfor CD34+ cells
Colonies/Culture
Cell preparation BFU-E CFU-GM Mixed
Post-Percoll(2 × 105 cells plated) 111 163 9
Postcolumn CD34+ cells(3 × 103 cells plated) 82 165 2
A CFU-GEMM assay utilizing pooled humanplasma was used in the comparative study. All val-ues shown are the mean of duplicate cultures. Thefollowing cytokines were added to all cultures: GM-CSF, 5 U/ml; Steel factor, 50 ng/ml; IL-3, 40 U/ml;erythropoietin, 2 U/ml.
CFU-GM colony numbers were consistentlyhigher compared to the control study and the Ph+
CML results. Individual JCML CFU-“GM”colonies at harvest showed all stages of the mono-cytic lineage including blast forms, promonocytes,monocytes and histiocytic macrophages, whereasin the control and Ph+ CML cultures, at least 50%of the colonies were granulocytic. These findingswere confirmed by morphological and cyto-chemical analysis of individual colonies pluckedfrom the cultures and by examination of the cel-lular composition of entire harvested culture plates.It is noteworthy that the CD34+ cells from patients21, 25 and 26 in these experiments were fraction-ated from the spleen after its surgical removal;the abnormal growth pattern of the cells was iden-tical to that seen with the marrow specimensfrom the other three patients.
In a separate experiment, CD34+ cells frompatients 24 were subsorted by flow cytometryinto HLA-Dr+ and HLA-Dr– populations, eachof 99% purity. Each population was cultured (3 × 103 cells plated) with IL-3, GM-CSF, Steelfactor and erythropoietin. The HLA-Dr+ pop-ulation yielded six BFU-E colonies, zero mixedcolonies and 175 monocyte-macrophagecolonies; the HLA-Dr– population yielded 18BFU-E colonies, zero mixed colonies, and > 1,000monocyte-macrophage colonies.
Removal of Adherent Cell Does Not ImpairJCML CD34+ Plating Efficiency
PB cells from patient 26 were fractionatedinto CD34+ cells then divided into two sam-ples. The first sample was not processed fur-ther; the other sample (106 cells/ml) wasincubated in 100 mm plastic Petri dishes withculture medium and 10% fetal calf serum for 6h. The nonadherent cells were then harvestedand subjected to a second 12 h incubation inplastic Petri dishes in order to achieve a thor-ough depletion of adherent cells. The resultantcellular preparation (reduced in number by 60%)was analyzed by flow cytometry using mono-cyte-associated surface markers; no monocyticelements were detected.
The two preparations of JCML CD34+ cellswere then compared for clonogenic activity in thepresence or absence of added GM-CSF (Table 4).Using the sample prior to adherent cell removal,spontaneous monocyte-macrophage colony growthwas observed in the absence of added GM-CSF;colony number was dependent on the cell num-ber plated. Added GM-CSF markedly enhancedthe clonogenic activity and induced higher mono-cyte-macrophage colony numbers in all the studies compared to cultures lacking GM-CSF.
Using the sample with adherent cellsremoved, spontaneous growth was stillobserved without added GM-CSF, and the num-bers were higher compared to the sample priorto adherent cell removal. Added GM-CSF
694 JCML Multilineage CD34+ Cells: Aberrant Growth and Differentiation Properties
Table 3. Comparison of growth response of JCMLand control CD34+ cells to IL-3, GM-CSF, Steelfactor and erythropoietin
Colonies/Culture
BFU-E CFU-GM Mixed
Control 170 99 6Ph+ CML 100 35 4JCML: PT 21 40 429 0
PT 22 8 455 0PT 23 7 761 0PT 24 7 625 0PT 25 3 501 0PT 26 20 434 0
A CFU-GEMM assay utilizing pooled human plasmawas used in these studies. The values shown are themean of duplicate cultures. The CD34+ cell preparationfrom JCML patients 21, 25 and 26 was derived fromsplenic tissue obtained at surgical removal. The otherJCML specimens were from BM. The followingcytokines were added: IL-3, 40 U/ml; GM-CSF, 5U/ml; Steel factor, 50 ng/ml; erythropoietin, 2 U/ml.
Table 4. Comparison of colony growth of PB JCMLCD34+ cell (patient 26) before and after removal ofadherent cells
Colonies/Culture
Cells plated Without GM-CSF With GM-CSF
Before Adherent Cell Depletion104/ml 28 1135 × 103/ml 14 43103/ml 1 11
After Adherent Cell Depletion104/ml 54 2495 × 103/ml 21 66103/ml 5 8
A CFU-GEMM assay utilizing 30% fetal calf serumwas used. Cell numbers were plated as shown with-out and with added GM-CSF, 5 U/ml. Values shownare the mean of duplicate cultures.

augmented colony numbers in all of the stud-ies compared to cultures lacking GM-CSF. Thecolony numbers were also higher when con-trasted to those seen from the sample prior toadherent cell removal.
A similar study was performed on spleenderived CD34+ cells after adherent cell depletionfrom patient 25 and spontaneous monocyte-macrophage colony growth in the absence ofadded growth factors was again observed (datanot shown).
TNF-α , IL-1 and GM-CSF Augment JCMLCD34+ Colony Numbers
Table 5 shows the growth response of JCMLCD34+ cells from four patients to TNF-α ,IL-1 and to GM-CSF when studied in CFU-GMclonogenic assays. Compared to CD34+ cells froma normal control marrow and from a Ph+ CMLmarrow specimen, the growth properties of theJCML cells were very different. Remarkable,without added cytokines, all four JCML speci-mens showed “autonomous” colony formation, afeature not seen with the controls. Once again,these “autonomous” CFU-“GM” colonies wererestricted exclusively to the monocyte-macrophagelineage. Daily observation of the culture platesshowed very early “autonomous” colony growthafter only 48 h of culture which, by day 7, becameexcessive numerically and very large and cellu-lar. The monocytic macrophage colony growthwas further augmented by adding IL-1, TNF-αand GM-CSF individually, and TNF-α and GM-CSF together. (Colony increments induced by allthree cytokines reached significant p values). TheJCML CD34+ cell fractions for these experimentswere obtained from spleen specimens from
patients 21 and 23, and from marrow from patients22 and 24; yet the unusual growth pattern andcytokine response were identical for all fourpatients. In the controls, CFU-GM colony forma-tion was only observed in cultures containingadded GM-CSF, and the colonies in these plateswere about 50% granulocytic in composition.
Hyper-Responsiveness of JCML CD34+ Cellsto GM-CSF
Table 6 shows the colony growth of JCMLBM CD34+ cells from patient 21 in response tograded doses of GM-CSF. Without addedGM-CSF, exuberant “autonomous” colonygrowth was again observed, and the colonieswere exclusively monocyte-macrophage in com-position. With increasing concentrations of addedGM-CSF, there was a dose-responsive increase inmonocyte-macrophage colonies, a characteris-tic not seen in the Ph+ CML or the normal controlarms of the study in which no growth of any sortwas seen. Even with minute concentrations ofGM-CSF, for example with 0.3 and 0.6 U/ml,enhanced monocyte-macrophage JCML colonygrowth was observed. Some CFU-GM granulo-cytic colony formation was noted in the Ph+ CMLcultures, but only at the highest concentration ofGM-CSF that was used (5.0 U/ml). The numberof JCML colonies in response to 5.0 U/ml ofGM-CSF was 8.7 fold higher than the Ph+ CMLCFU-GM colony numbers, however (p < 0.001).
The exaggerated response of JCML CD34+
cells to GM-CSF was confirmed in a similarbut modified study of BM from patient 24.Without added cytokine, a mean of 136 “autono-mous” colonies/culture were seen; in responseto two doses of GM-CSF, 0.6 U/ml and to 5.0
Freedman, Hitzler, Bunin et al. 695
Table 5. Response of JCML CD34+ cells to IL-1α , TNF-α and GM-CSF
CFU-GM Colonies/Culture
JCML JCML JCML JCML Ph+ NormalCytokines PT 21 PT 22 PT 23 PT 24 CML control
None 100 127 589 108 4 3IL-1 130 180 688 143 0 0TNF * 202 1,022 148 0 0GM-CSF 224 265 1,495 188 89 3TNF+ GM-CSF 243 503 1,136 503 15 0
A CFU-GEMM assay utilizing 30% fetal calf serum was used but the following cytokines were substituted for the“standard” growth factor “cocktail” shown in Table 3: IL-1α, 100 U/ml; TNF-α, 103 U/ml; GM-CSF, 5 U/ml. Valuesshown are the mean of duplicate cultures. Specimens from JCML patients 21 and 23 were derived from splenictissue obtained at surgical removal. The other JCML, Ph+ CML and control specimens were from marrow.*Not performed in this experiment, but growth augmentation was confirmed with TNF-α in subsequent studies.
U/ml, monocyte-macrophage colony growthincreased to 179 (p < 0.1) and 240 (p < 0.01)colonies/culture, respectively.
Interplay of TNF-α , GM-CSF and TheirBlockers on JCML CD34+ Cell Growth
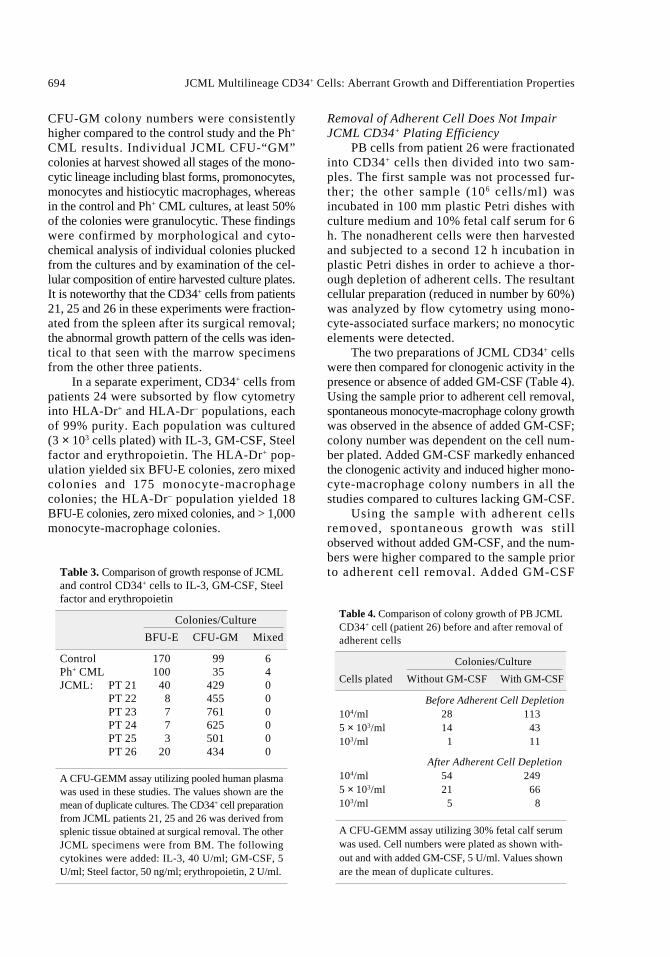
Regulation of JCML CD34+ cell growth byTNF-α, GM-CSF and their respective antagonistswas studied using fractionated marrow cells frompatient 24 (Figs. 1 and 2). Data in Figure 1 confirmthat in the absence of added cytokines,“autonomous” JCML colony formation ensued,and that GM-CSF and TNF-α individuallyaugmented the colony formation. When the respec-tive antibodies were added to cultures containingGM-CSF or TNF-α, the growth augmentation wasreversed and JCML CD34+ cell growth was inhib-ited. The suppressive effect by the respective anti-bodies could be reversed by adding excess doses ofcytokine (data not shown). Remarkably, anti-TNF-α antibodies reversed the growth-promoting effectof GM-CSF, and anti-GM-CSF antibodiesreversed the growth-promoting effect of TNF-α.
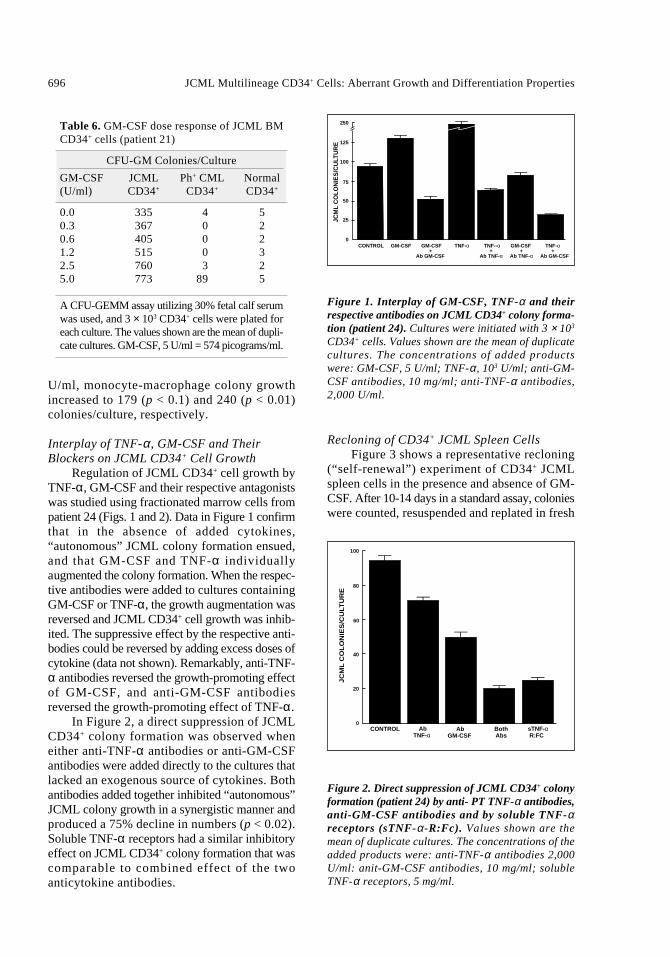
In Figure 2, a direct suppression of JCMLCD34+ colony formation was observed wheneither anti-TNF-α antibodies or anti-GM-CSFantibodies were added directly to the cultures thatlacked an exogenous source of cytokines. Bothantibodies added together inhibited “autonomous”JCML colony growth in a synergistic manner andproduced a 75% decline in numbers (p < 0.02).Soluble TNF-α receptors had a similar inhibitoryeffect on JCML CD34+ colony formation that wascomparable to combined effect of the two anticytokine antibodies.
Recloning of CD34+ JCML Spleen Cells Figure 3 shows a representative recloning
(“self-renewal”) experiment of CD34+ JCMLspleen cells in the presence and absence of GM-CSF. After 10-14 days in a standard assay, colonieswere counted, resuspended and replated in fresh
696 JCML Multilineage CD34+ Cells: Aberrant Growth and Differentiation Properties
Table 6. GM-CSF dose response of JCML BMCD34+ cells (patient 21)
CFU-GM Colonies/Culture
GM-CSF JCML Ph+ CML Normal(U/ml) CD34+ CD34+ CD34+
0.0 335 4 50.3 367 0 20.6 405 0 21.2 515 0 32.5 760 3 25.0 773 89 5
A CFU-GEMM assay utilizing 30% fetal calf serumwas used, and 3 × 103 CD34+ cells were plated foreach culture. The values shown are the mean of dupli-cate cultures. GM-CSF, 5 U/ml = 574 picograms/ml.
250
125
100
75
50
25
0CONTROL GM-CSF GM-CSF TNF-α TNF--α GM-CSF TNF-α
+ + + + Ab GM-CSF Ab TNF-α Ab TNF-α Ab GM-CSF
JCM
L C
OLO
NIE
S/C
ULT
UR
E
Figure 1. Interplay of GM-CSF, TNF-α and theirrespective antibodies on JCML CD34+ colony forma-tion (patient 24). Cultures were initiated with 3 × 103
CD34+ cells. Values shown are the mean of duplicatecultures. The concentrations of added productswere: GM-CSF, 5 U/ml; TNF-α, 103 U/ml; anti-GM-CSF antibodies, 10 mg/ml; anti-TNF-α antibodies,2,000 U/ml.
Figure 2. Direct suppression of JCML CD34+ colonyformation (patient 24) by anti- PT TNF-α antibodies,anti-GM-CSF antibodies and by soluble TNF-αreceptors (sTNF-α -R:Fc). Values shown are themean of duplicate cultures. The concentrations of theadded products were: anti-TNF-α antibodies 2,000U/ml: anit-GM-CSF antibodies, 10 mg/ml; solubleTNF-α receptors, 5 mg/ml.
JC
ML
CO
LO
NIE
S/C
ULT
UR
E
100
80
60
40
20
0Ab
TNF-α Ab
GM-CSF BothAbs
sTNF-αR:FC
CONTROL

semisolid medium with or without GM-CSF.Cultures exposed to GM-CSF during the primaryculture could be “self-renewed” for three passageswhether or not GM-CSF was added to further sub-cultures. JCML colonies, that were initially cul-tured in the absence of GM-CSF, however, hadmarkedly restricted recloning capability.
Multilineage Potential of JCML CD34+ CellsThe cytogenetic marker monosomy 7 in
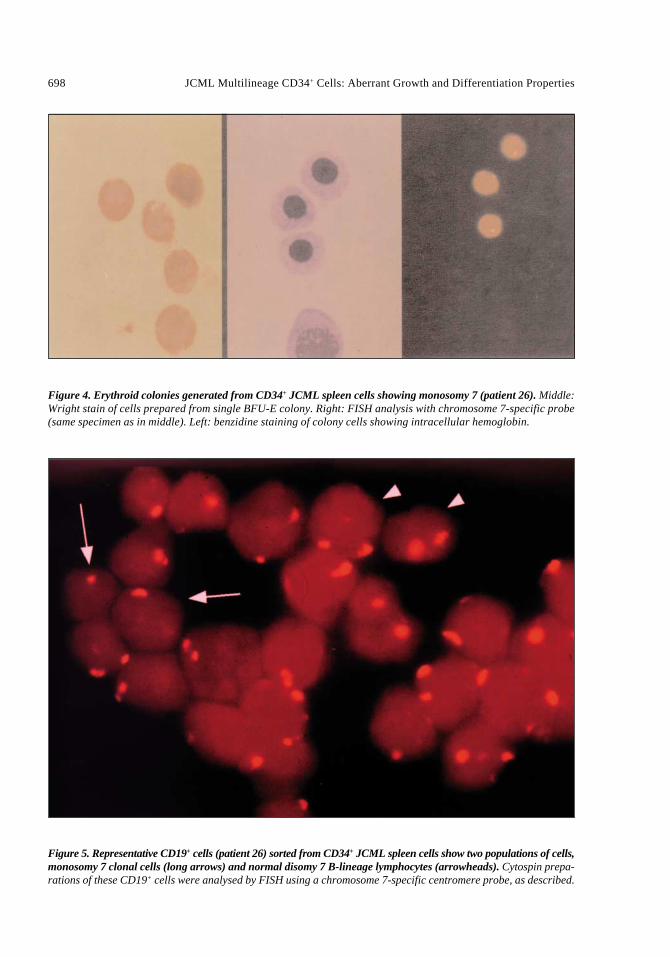
malignant spleen cells from patient 26 allowedus to investigate leukemic involvement in lin-eages other than monocyte-macrophage fromCD34+ cells and their progeny. Although thenumber of BFU-E colonies was very low com-pared to controls (Table 3), individual coloniescould be isolated for study. Monosomy 7 wasclearly demonstrated in benzidine-positive ery-throid cells generated from CD34+ spleen cells(Fig. 4). Similarly, CD19-enriched lymphoidpopulations from CD34+ spleen cells from thesame patient showed monosomy 7, as well asnormal disomy 7 B-lineage cells (Fig. 5).
Discussion
Although the hematologic phenotype is sim-ilar for all JCML patients, about 20% show aclonal cytogenetic change in myeloid cells, such astrisomy 8 and monosomy 7. The role of these cyto-genetic abnormalities in the pathogenesis of JCMLis unclear, but they serve as useful markers fortracking the cellular origin of the disease.
Thus, Inoue et al. [25] used cytogeneticdata of patients with JCML to demonstratethe presence of the same karyotypic abnor-mality (monosomy 7) in both initial mar-row specimen and pooled erythroid colonies,suggesting erythroid involvement in themalignant JCML cell clone in addition to themonocyte-macrophage lineage. Lau et al. [16]described a case of JCML terminating in B-lymphoid blast crisis. FISH demonstrated thatlymphoblasts isolated after blast transforma-tion, as well as myelomonocytic marrow cellsisolated during the “chronic” phase of the dis-ease, were positive for monosomy 7. Thus,the leukemic “event” in JCML clearly occursat an early stage of hematopoiesis judgingfrom these data showing multilineage involve-ment. This thesis is in complete agreementwith data by Busque et al. who demonstrated
in one female JCML patient, using a G6PDCT polymorphism clonality assay, that theleukemic clone involved multiple hematopoieticlineages [15].
To examine this directly, CD34+ spleen cellsfrom patient 26 were used to analyze lineagepotential of JCML cells. The CD34 antigen iden-tifies cell populations that are enriched forpluripotent and lineage-restricted hematopoieticprogenitor cells in vitro [26-36], some of whichare capable of marrow reconstitution in vivo[31-33]. The phenotype of the most primitivehuman hematopoietic cells isolated so far showsexpression of this membrane protein [34-36].Thus, we reasoned that CD34+ JCML cells were asuitably defined subset of cells for an appropriateanalysis of their multilineage potential.
Freedman, Hitzler, Bunin et al. 697
Control
GM-CSF
GM-CSF
GM-CSF
205
40306 244
0249 531
395
Figure 3. Recloning of JCML CD34+ spleen cells inthe presence or absence of GM-CSF (patient 26).CD34+ cells (3 × 103) were incubated with and without5 U/ml GM-CSF and serially recloned. Values shownare the mean of duplicate cultures of a representativeexperiment.
698 JCML Multilineage CD34+ Cells: Aberrant Growth and Differentiation Properties
Figure 4. Erythroid colonies generated from CD34+ JCML spleen cells showing monosomy 7 (patient 26). Middle:Wright stain of cells prepared from single BFU-E colony. Right: FISH analysis with chromosome 7-specific probe(same specimen as in middle). Left: benzidine staining of colony cells showing intracellular hemoglobin.
Figure 5. Representative CD19+ cells (patient 26) sorted from CD34+ JCML spleen cells show two populations of cells,monosomy 7 clonal cells (long arrows) and normal disomy 7 B-lineage lymphocytes (arrowheads). Cytospin prepa-rations of these CD19+ cells were analysed by FISH using a chromosome 7-specific centromere probe, as described.

The hemoglobin content of BFU-E coloniesderived from the CD34+ cells of patient 26 wasconfirmed by benzidine staining of cell smears.In addition, cells of B-lymphoid phenotype wereprepared from CD34+ JCML cells by fluores-cence-activated cell sorting using antibody toCD19. Both cell populations, erythroid and B-lymphoid cells, were shown by FISH to bepositive for monosomy 7, the marker present inthe diagnostic patient marrow and in monocyte-macrophage colonies derived from marrow andspleen cells. The malignant transformation inJCML therefore occurs in a multipotential cellwhich can generate monocyte-macrophage, ery-throid and B-lymphoid lineages. This in vitrofinding is in keeping with reported data [15] andsuggests a “stem” cell origin of JCML.
Excessive formation of monocyte-macro-phage colonies with impaired normal residualhematopoiesis is a characteristic pattern of JCMLcell proliferation in vitro [2, 3, 6]. Endogenousproduction of cytokines such as TNF-α, GM-CSFand IL-1 [9-12] by JCML cultures has been pro-posed as an explanation for this unique prolifera-tive behavior. Constitutive secretion of TNF-α byJCML cells inhibits normal hematopoietic prog-enitors in vitro [9] and, at the same time with GM-CSF, acts as an autocrine stimulator of excessivemalignant monocyte-macrophage colony forma-tion [9, 10]. Emmanuel et al. [11] demonstratedthe hypersensitivity of mononuclear JCML cells toexogenous GM-CSF as measured by monocyte-macrophage colony formation. IL-1 was found tostimulate the release of JCML growth-promotingcytokines by other BM or PB cells [12] and wasimplicated as a paracrine growth factor of JCMLcells. Importantly, in vitro proliferation of JCMLcells could be decreased by antibodies to TNF-αand GM-CSF [9], by IL-1 receptor antagonist [37],soluble IL-1 receptors, IL-4 (which inhibits IL-1and TNF-α production), and pentoxiphyllinewhich downregulates TNF-α mRNA [17, 26].
The fractionation of CD34+ JCML cellsallowed experiments with a more clearly definedJCML cell population which did not appear “con-taminated” by a growth factor secreting adher-ent cells, as evidenced by enhanced clonogenicefficiency after exhaustive adherent cell removal(see below). The results were clear-cut, consistentin every patient studied and provided compellingevidence that JCML is a hematopoetic malig-nancy with in vitro growth characteristics entirelyreproducible by CD34+ JCML cells.
First, the colony growth pattern, in responseto a combination of IL-3, GM-CSF, Steel factorand erythropoietin, was strikingly different fromnormal BM controls as well as from Ph+ CMLcells. The JCML cultures always showed reducedto absent erythroid colony growth and increasedmonocyte-macrophage colony formation.
Second in contrast to normal BM cells,CD34+ JCML cells grew spontaneously in theabsence of exogenous growth factors. The addi-tion of GM-CSF, TNF-α and IL-1 resulted infurther augmentation of the already excessivemonocyte-macrophage growth. CD34+ JCMLcells demonstrated the hypersensitivity ofcolony formation in response to GM-CSF atminute concentrations that have no prolifera-tive effect on normal marrow cells. This fea-ture was previously described in mixed,nonadherent, T cell depleted populations oflight density JCML cells [11].
The spontaneous colony formation by JCMLCD34+ cells in the absence of added cytokinesprompts the question of whether growth factor-secreting “contaminating” monocytes are pre-sent in the samples. Although this is unlikelybecause of the high degree of purity of the CD34+
preparations proven by flow cytometry, detailedexperiments were performed to address this ques-tion directly. We observed that total removal ofadherent cells, proven by flow cytometry by anexhaustive double-depletion technique over 18h of incubation with plastic, did not impair ordecrease the plating efficiency of JCML CD34+
cells in the absence of added growth factors.Indeed, spontaneous formation of JCML mono-cyte-macrophage colonies were augmented, pos-sibly due to enrichment because of removal ofnonclonogenic CD34+ cells with surface adher-ence properties. Thus, the results suggest that ahighly purified population of JCML CD34+ can“self-start” and form colonies in keeping withan autocrine growth pattern. Our data differ fromprevious studies by others that showed impairedspontaneous JCML colony formation after adher-ent cell depletion [12, 38]. Their studies wereperformed on light density cells and ours onhighly purified CD34+ cells. Nonetheless, theexplanation for the contradictory results is notreadily evident. Taking an overview of their dataand ours, we would have to conclude that bothautocrine and paracrine growth loops are opera-tive in JCML cell proliferation and contribute tothe cytokine-driven leukemic process.
Freedman, Hitzler, Bunin et al. 699

Serial recloning experiments showed thatexposure of CD34+ JCML cells to GM-CSF dur-ing early culture passage was associated withsuccessful “self-renewal” for up to three pas-sages. Absence of GM-CSF during this phaseof proliferation could not be compensated bystimulation with GM-CSF during subsequent culture generations. In addition to hypersensitiv-ity of the overall JCML population to GM-CSF, atransient growth factor-sensitive period duringearly in vitro proliferation was demonstrated usingthe CD34+ JCML cells.
Finally, the abnormal growth of CD34+ JCMLcells could be suppressed by antagonists to TNF-αand GM-CSF. Working with light density cells,Emmanuel et al. also found that anti-GM-CSF, anti-bodies suppressed spontaneous JCML colonygrowth but anti-TNF antibodies had no significanteffect [38]. In our hands, however, mAbs to TNF-α and GM-CSF, alone or in combination, resultedin growth inhibition comparable to the effect ofsoluble TNF-α receptors. These data confirm ourprevious work [9] and suggest interference by theseagents on the effect of autocrine growth stimulationthat was previously identified in the light densitypopulation of cells [9]. Reversal of the effects ofGM-CSF by anti-TNF-α and TNF-α by anti-GM-CSF illustrates the complex interplay of cytokinesinvolved in JCML growth stimulation.
Thus, JCML is a disease that originates inpluripotent cells and can be identified in the CD34+
population. All of the in vitro features previouslyascribed to JCML “progenitors” in the mixed lightdensity cell population can be reproduced byCD34+ JCML cells. In addition, CD34+ JCML cellscan engraft in severe combined immunodeficiencymice and cause rapid infiltration of murine tissues,cachexia and hematopoetic failure similar to thehuman disease [39]. Therefore, JCML CD34+ cellsrepresent a close approximation of the cell of originof the disease and provide an entry point for fundamental investigations of this disorder.
Acknowledgements
We are grateful to Wilma Vanek for her cellculture expertise, and to Zong Mei Zheung andTeresa Scheidl for excellent technical assistancewith the FISH studies.
Supported by the National Cancer Institute ofCanada, the Histiocytosis Association of America,and the Medical Research Council of Canada.
References
1 Estrov Z. Zimmerman B, Grunberger T et al.Characterization of malignant peripheral bloodcells of juvenile chronic myelogenous leukemia.Cancer Res 1986;46:6456-6461.
2 Estrov Z, Grunberger T, Chan HSL et al. Juvenilechronic myelogenous leukemia: characterizationof the disease using cell cultures. Blood1986;67:1382-1387.
3 Estrov Z, Dube IA, Chan HSL et al. Residual juve-nile chronic myelogenous leukemia cells detectedin peripheral blood during clinical remission.Blood 1987;70:1466-1469.
4 Estrov Z, Lau AS, Williams BRG et al.Recombinant human interferon alpha-2 and juve-nile chronic myelogenous leukemia: cell receptorbinding, enzymatic induction, and growth sup-pression in vitro. Exp Hematol 1987;15:127-132.
5 Freedman MH, Estrov Z. Juvenile chronic myel-ogenous leukemia and histiocytosis syndromesin children. Lancet 1987;1:754.
6 Freedman MH, Estrov Z, Chan HSL. Juvenilechronic myelogenous leukemia. Am J PediatrHematol Oncol 1988;10:261-267.
7 Chan HSL, Estrov Z, Weitzman SS et al. Thevalue of intensive combination chemotherapy forjuvenile chronic myelogenous leukemia. J ClinOncol 1987;5:1960-1967.
8 Kirby MA, Weitzman S, Freedman MH. Juvenilechronic myelogenous leukemia: differentiationfrom infantile cytomegalovirus infection. Am J Pediatr Hematol Oncol 1990;12:292-296.
9 Freedman MH, Cohen A, Grunberger T et al.Central role of tumour necrosis factor, GM-CSF,and interleukin 1 in the pathogenesis of juvenilechronic myelogenous leukaemia. Br J Haematol1992;80:40-48.
10 Gualtieri RJ, Emanuel PD, Zuckerman KS et al.Granulocyte-macrophage colony-stimulating fac-tor is an endogenous regulator of cell prolifera-tion in juvenile chronic myelogenous leukemia.Blood 1989;74:2360-2367.
11 Emanuel PD, Bates LJ, Castleberry RP et al.Selective hypersensitivity to granulocyte-macrophage colony-stimulating factor by juve-nile chronic myeloid leukemia hematopoieticprogenitors. Blood 1991;77:925-929.
12 Bagby GC, Dinarello CA, Neerhout RC et al.Interleukin-1-dependent paracrine granulopoiesisin chronic granuloycytic leukemia of the juve-nile type. J Clin Invest 1988;82:1430-1436.
13 Altman AJ. Schwartz AD. Malignant Diseasesof Infancy, Childhood and Adolescence.Philadelphia: W.B. Saunders, 1983:228-229.
700 JCML Multilineage CD34+ Cells: Aberrant Growth and Differentiation Properties

14 Nathan DG, Oski FA, Hematology of Infancy andChildhood, 4th Edition. Philadelphia: W.B.Saunders, 1993:1293-1295.
15 Busque L, Gilliland DG, Pryhcal JT et al.Clonality in juvenile chronic myelogenousleukemia. Blood 1995;85:21-30.
16 Lau RC, Squire J. Brisson L et al. Lymphoid blastcrisis of B-lineage phenotype with monosomy 7in a patient with juvenile chronic myelogenousleukemia (JCML). Leukemia 1994;8:903-908.
17 Estrov Z, O’Brien PM, Horvath A et al.Interruption of autocrine/paracrine growth factorstimulatory mechanisms: a potential approach forjuvenile chronic myelogenous leukemia therapy.Int J Pediatr Hematol Oncol 1994;1:253-258.
18 Berenson RJ, Bensinger WI, Kalamasz D. Positive selection of viable cell populations usingavidin-biotin immunoadsorption. J ImmunolMethods 1986;91:11-19.
19 Berenson RJ, Bensinger WI, Kalamasz D et al. Elim-ination of Daudi lymphoblasts from human bonemarrow using avidin-biotin immunoabsorption.Blood 1986;67:509-515.
20 Berenson RJ, Bensinger WI, Kalamasz D et al.Engraftment of dogs with Ia-positive marrow cellsisolated by avidin-biotin immunoadsorption. Blood 1987;69:1363-1367.
21 Tucker SB, Pierre RV, Jordan RE. Rapid identifi-cation of monocytes in mixed mononuclear cellpreparations. J Immunol Methods 1977;14:267-269.
22 Lichter P, Tang CJ, Call K et al. High resolutionmapping of human chromosome II by in situhybridization with cosmid clones. Science1990;247:64-69.
23 Lau AS, Livesey JF. Endotoxin induction oftumor necrosis factor is enhanced by acid-labileinterferon-alpha in acquired immunodeficiencysyndrome. J Clin Invest 1989;84:738-743.
24 Mohler KM, Torrance DS, Smith CA et al.Soluble tumor necrosis factor (TNF) receptorsare effective therapeutic agents in lethal endo-toxemia and function simultaneously as both TNFcarriers and TNF antagonists. J Immunol1993;151:1548-1561.
25 Inoue S, Shibata T, Ravindranath Y et al. Clonalorigin of erythroid cells in juvenile chronic myelogenous leukemia. Blood 1987;69:975-976.
26 Andrews RG, Singer JW, Bernstein ID. Monoclonalantibody 12-8 recognizes a 115-kd molecule pre-sent on both unipotent and multipotent hematopoi-etic colony-forming cells and their precursors.Blood 1986;67:842-845.
27 Brandt JE, Baird N, Lu L et al. Characterizationof a human hematopoietic progenitor cell capableof forming blast cells containing colonies in vitro.J Clin Invest 1988;82:1017-1027.
28 Andrews RG, Singer JW, Bernstein ID. Precursorsof colony-forming cells in humans can be distin-guished from colony forming cells by expressionof the CD33 and CD34 antigen and light scatterproperties. J Exp Med 1989;169:1721-1731.
29 Sutherland HJ, Eaves CJ, Eaves AC et al.Characterization and partial purification of humanmarrow cells capable of initiation long-termhematopoiesis in vitro. Blood 1989;74:1563-1570.
30 Sutherland HJ, Lansdorp PM, Henkelman DH etal. Functional characterization of individualhuman hematopoietic stem cells cultured at lim-iting dilution on supportive stromal layers. ProcNatl Acad Sci USA 1990;87:3584-3588.
31 Berenson RJ, Andrews RG, Bensinger WI et al.Antigen CD34+ marrow cells engraft lethally irra-diated baboons. J Clin Invest 1988;81:951-955.
32 Berenson RJ, Bensinger WI, Hill RS et al.Engraftment after infusion of CD34+ marrow cellsin patients with breast cancer or neuroblastoma.Blood 1991;77:1717-1722.
33 Peault B, Weissman IL, Baum C et al. Lymphoidreconstitution of the human fetal thymus in SCIDmice with CD34+ precursor cells. J Exp Med1991;174:1283-1286.
34 Baum CM, Weissman IL, Tsukamoto AS et al.Isolation of a candidate human hematopoieticstem-cell population. Proc Natl Acad Sci USA1992;89:2804-2808.
35 Craig W, Kay R, Cutler RL et al. Expression ofThy-1 on human hematopoietic progenitor cells.J Exp Med 1993;177:1331-1342.
36 Huang S, Terstappen WMM. Lymphoid andmyeloid differentiation of single human CD34+,HLA-Dr+, CD38– hematopoietic stem cells. Blood1994;83:1515-1526.
37 Schiro R, Longoni D, Rossi V et al. Suppressionof juvenile myelogenous leukemia colony growthby interleukin-1 receptor antagonist. Blood1994;83:460-465.
38 Emanuel PD, Bates LJ, Zhu S-W et al. The roleof monocyte-derived hematopoietic growth fac-tors in the regulation of myeloproliferation injuvenile chronic myelogenous leukemia. ExpHematol 1991;19:1017-1024.
39 Lapidot T, Grunberger T, Vermoor J et al.Identification of immature juvenile chronicmyelogenous leukemia cells capable of initiat-ing disease in primary and secondary SCIDmice. Blood 1996;88:2655-2664.
Freedman, Hitzler, Bunin et al. 701
















![[Ghiduri][Cancer]Chronic Myelogenous Leukemia](https://img.pdfslide.net/doc/110x75/577cc6ea1a28aba7119f80de/ghiduricancerchronic-myelogenous-leukemia.jpg)












