Embed Size (px)
Citation preview

LACTOSE METABOLISM
I. CARBOHYDRATE METABOLISM OF LACTOBACILLUS BULGARI CUS STRAIN GERE A*
BY W. J. RUTTER AND R. G. HANSEN
(From the Laboratory of Biochemistry, Department of Dairy Science, University of Illinois, Urbana, Illinois)
(Received for publication, November 10, 1952)
Snell et al. (1) have shown that LactobaciZZus bulguricus strain Gere A grows in a medium containing lactose or other p-galactosides as the carbon source, but does not grow initially in a medium containing galactose or glucose or mixtures of the two. Only after prolonged incubation with these monosaccharides, and presumably through adaptation, can this mi- croorganism utilize galactose or glucose for growth. The carbohydrate metabolism of microorganisms giving evidence of utilizing lactose by some other means than through hydrolysis is of interest in connection with the mechanisms of “direct” utilization of disaccharides (2, 3) and may perhaps lend information concerning the general biochemical problem of lactose metabolism. Accordingly, the purpose of this report is to describe certain aspects of the metabolism of L. bulgaricus Gere A with particular reference to lactose and its constituent monosaccharides, glucose and galactose.
Methods
Bacteriological-Cultures of L. bulgaricus Gere A were obtained through the kindness of Dr. E. E. Snell and Dr. W. B. Sarles. Cells were grown by transfer of inocula to suitable quantities of the medium of Snell et al. (1). When the bacteria were consistently derived from media containing lactose, they were termed unadapted cells. In certain experiments, galac- tose-adapted or glucose-adapted cells were used. These were grown in the same medium except that the appropriate sugar was substituted for lactose. The cells were harvested at the end of the logarithmic growth phase (about 24 hours) by means of a Sharples centrifuge. Resting cell suspensions were prepared by washing the harvested cells twice in saline and suspend- ing them in the desired volume. Cell-free extracts were prepared by 1 hour sonic oscillation (Raytheon 200 watt sonic oscillator with a 10 kilo- cycle driver) of resting cells suspended in 0.15 M potassium chloride.
For the experiments involving the growth of this microorganism in de- fined nutrients, a double strength basal medium was prepared with the
* Supported in part by a grant from the United States Atomic Energy Commission (contract No. AT(ll-l)-67 project No. 10).
311
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from

312 LACTOSE METABOLISM OF L. BULGARICUS
composition shown in Table I and autoclaved in 5 ml. lots for 10 minutes at 120’. The carbohydrate source and other factors to be tested for growth-promoting activity were sterilized by passage through a bacteri- ological filter and added, with sterile H%O, to a final volume of 10 ml. per tube. 1 drop of inoculum of unadapted cells washed three times wit,h sterile saline was added to each tube. Incubation at 37” followed.
Chemical-Acid production was measured in the Warburg instrument by release of CO, from a bicarbonate buffer in an atmosphere of 95 per
TABLE I Composition of Basal Medium*
nL-Alanine 1 gm. DL-Asparagine 1 “ L-Glutamic acid 200 mg. L-Arginine hydrochloride 200 (‘ L-Lysine hydrochloride H20 100 “ L-Cysteine 100 lL L-Cystine 100 “ L-Histidine 100 I‘ L-Isoleucine 100 I‘ L-Leucine 100 ‘I L-Methionine 100 (‘ L-Phenylalanine 100 “ L-Proline 100 “ L-Threonine 100 (‘ L-Tyrosine 100 ‘r L-Valine 100 “ L-Tryptophan 100 <‘ L-Serine 100 ‘I Glycine 100 “
* For 500 ml. double strength medium.
Biotin Pteroylglutamic acid p-Aminobenzoic acid Pyridoxal Thiamine hydrochloride Calcium pantothenate Riboflavin Nicotinic acid Salts A
“ B Sodium acetate Tween 80 Adenine sulfate Guanine hydrochloride Uracil Lactose Water
10 gm. 100 “ 200 “ 200 ‘(
1 mg. 1 ‘C 1 “
1 “ 5 ml. 5 ‘C
6 gm. 1 I‘
10 mg. 10 ‘[ 10 “ 10 gm.
To 500 ml.
cent Nz and 5 per cent COZ. p-Galactose-l-phosphate and p-glucose-l- phosphate were synthesized essentially according to the method of Reithel (4). a-Galactose-l-phosphate was synthesized by the procedure described by Kosterlitz (5). a-Glucose-l-phosphate was obtained through the kind- ness of Dr. H. A. Lardy. Galactose-6-phosphate was synthesized by the method of Levene and Raymond (6). The specific rotations of the various intermediates and the final phosphate esters were in agreement with the values reported in the literature for these compounds. Triphosphopyri- dine nucleotide (TPN) was procured from the Sigma Chemical Company. Uridine-diphosphoglucose (UDPG) was isolated and purified according to the directions of Caputto et al. (7). This concentrate has been shown (8) to contain, in addition to UDPG, small amounts of another unidenti- fied uridine-diphosphate derivative and uridylic acid. L. bulgur&s factor
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from

W. J. RU’FTER AND R. G. HANSEN 313
(pantethine) was furnished by Parke, Davis and Company. Pyridoxal phosphate was obtained through the kindness of Dr. W. A. Wood. Crys- talline vitamin BIz was obtained from Merck and Company, Inc. Uracil, erotic acid, uridine, and uridylic acid were procured from Nutritional Bio- chemicals Incorporated. Phosphoglucomutase was isolated from rabbit muscle by the method of Najjar (9). A crude Zwischenferment was pre- pared by modification of the method of Kornberg (10) to include a heat treatment at pH 9.0 and 40’. Phosphorus was determined by the method of Fiske and Subbarow (11). Lactase activity was followed by the use of the modified Steinhof reagent described by Caputto et al. (12). Reducing power was determined by the Somogyi-Nelson method (13, 14). Total carbohydrate was measured with the anthrone reagent modified for use by Trevelyan and Harrison (15). Paper chromatography of sugars was ef- fected with the solvents ethyl acetate-pyridine-water in the proportions 2 : 1: 2, as described by Jermyn and Isherwood (16). The aniline oxalate reagent of Partridge (17) was satisfactory for the detection of sugar spots.
Results
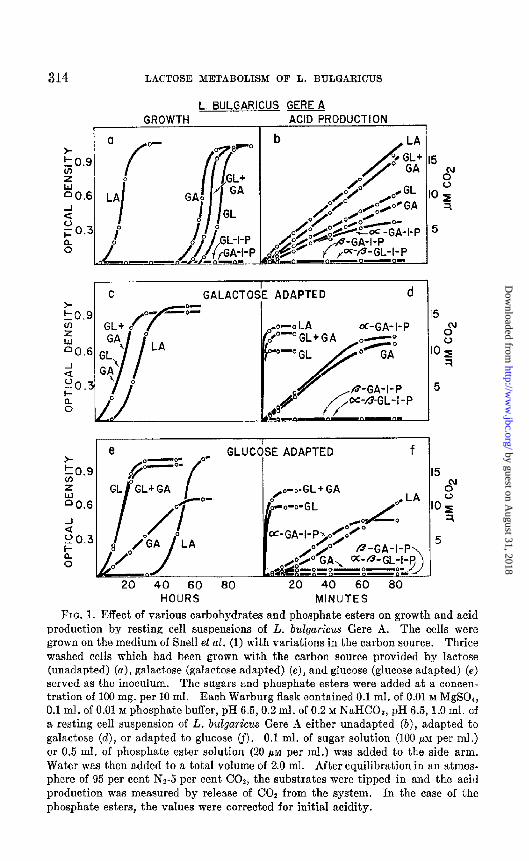
Comparison of Growth with Acid Production by Resting Cell Suspensions in Presence of Various Substrates-The growth characteristics of unadapted L. buZgaricus Gere A are shown in Fig. 1, a; immediate growth was obtained only with lactose as the carbon source. The organism was modified to grow on glucose or gaIactose by incubating it for a period of 60 to 70 hours in a medium containing these sugars. When any of the (II- or P-glucose or galactose-l-phosphates were used as the carbon source, no growth of this organism occurred. It is shown in Fig. 1, b that resting cell suspensions which would grow only in a medium containing lactose or other ,&galacto- sides nevertheless readily produced acid from free glucose or galactose or a mixture of the two. When 10 PM of glucose and 10 PM of galactose were added simultaneously to resting cell suspensions, the rate of acid production was similar to that produced from 10 PM of lactose. Other experiments have shown that under these conditions the rate of acid production from either 20 PM of glucose or 20 ,LLM of galactose is similar to that from 10 PM
of lactose. Addition of cr-galactose-l-phosphate caused acid production at a similar rate to that produced from galactose. None of the other phos- phate esters tested gave this response.
Fig. 1, c, indicates that cells adapted to grow on galactose also acquired the ability to grow on glucose and retained their ability to grow on lactose. Resting cell preparations of a culture adapted to grow on galactose (Fig. 1, d) produced acid more readily from glucose or lactose than from gaIac- tose or cu-galactose-l-phosphate. None of the other hexose phosphate es- ters elicited acid production.
As illustrated in Fig. 1, e, cells adapted to grow on glucose grew slowly
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from
314 LACTOSE METABOLISM OF L. BULGARICUS
L. BULGARICUS GERE A GROWTH ACID PRODUCTION
I C GALACTOSE ADAPTED d ]
.6
GLUCdSE ADAPTED f
15 0”
10; q
5
BO HOURS MINUTES
FIG. 1. Effect of various carbohydrates and phosphate esters on growth and acid production by resting cell suspensions of L. bdgaricus Gere A. The cells were grown on the medium of Snell et al. (1) with variations in the carbon source. Thrice washed cells which had been grown with the carbon source provided by lactose (unadapted) (a), galactose (galactose adapted) (c), and glucose (glucose adapted) (e) served as the inoculum. The sugars and phosphate esters were added at a concen- tration of 100 mg. per 10 ml. Each Warburg flask contained 0.1 ml. of 0.01 M MgS04, 0.1 ml. of 0.01 M phosphate buffer, pH 6.5,0.2 ml. of 0.2 M NaHCOa, pH 6.5,1.0 ml. of a resting cell suspension of L. bulgur&s Gere ,4 either unadapted (b), adapted to galactose (d), or adapted to glucose cf). 0.1 ml. of sugar solution (100 PM per ml.) or 0.5 ml. of phosphate ester solution (20 pi per ml.) was added to the side arm. Water was then added to a total volume of 2.0 ml. After equilibration in an atmos- phere of 95 per cent NQ-5 per cent COe, the substrates were tipped in and the acid production was measured by release of COz from the system. In the case of the phosphate esters, the values were corrected for initial acidity.
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from

W. J. RUTTER AND R. G. HANSEN 315
on galactose but would not initially grow on lactose. After a 40 to 50 hour incubation in the presence of lactose, however, growth resulted. Resting cells harvested from a cuhure adapted to grow on glucose produced acid readily only from glucose (Fig. 1, f); lactose and a-galactose-l-phosphate were utilized much less effectively, while galactose caused little if any pro- duction of acid. Other phosphate esters were again without effect.
Enzyme Activity of Cell-Free Extracts-It was observed that cell-free ex- tracts prepared from unadapted cells contained lactose activity. Thus, an extract from approximately 10 mg. of dried cells could hydrolyze 10 mg. of lactose in 1 hour. These extracts also contained galactokinase activity and hexokinase activity.
Attempts to measure the presence of another enzyme capable of cata- lyzing the phosphorolysis of lactose were uniformly without success. No uptake of inorganic phosphate could be observed on incubation of the cell- free extracts with excess lactose. Various combinations of galactose-l- phosphate with glucose and glucose-l-phosphate were incubated with the extract in the presence of arsenate. If enzymes similar to sucrose phos- phorylase (18) or maltose phosphorylase (19) were present, an increase in reducing power could be expected; no increase was observed. In addition, short term incubations of sonic extracts with an excess of lactose together with added phosphoglucomutase and uridine-diphosphoglucose (the extract contained a galactowaldenase similar to that discovered by Caputto et al. (20) in yeasts) produced no accumulation of glucose-6-phosphate as meas- ured by the reduction of TPN in the presence of added Zwischenferment.
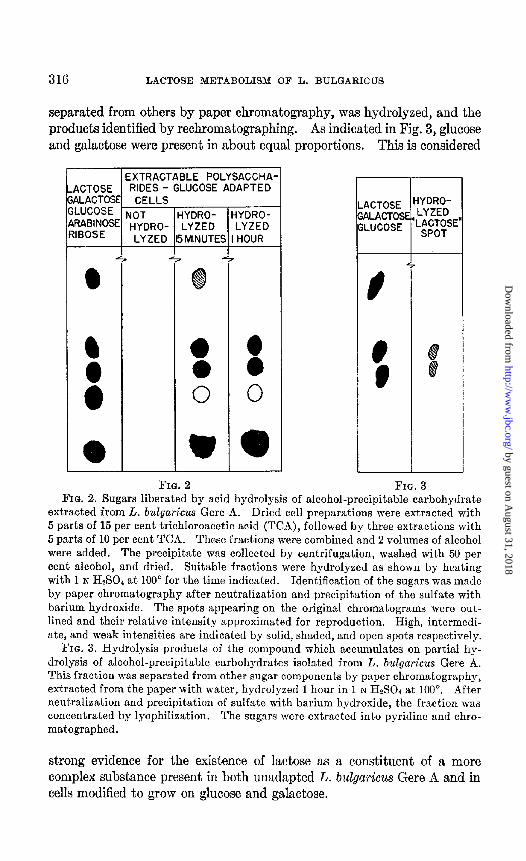
Trichloroacetic Acid-Extractable Carbohydrates from L. bulgaricus Gere A- It was found that repeated extractions of cells with trichloroacetic acid removed nearly all of the cellular carbohydrates as detected by the an- throne reagent. A qualitative investigation was made of the alcohol-pre- cipitable carbohydrates extracted from cells grown on lactose, eelIs grown on glucose, and cells grown on galactose. Fig. 2 pictures a typical chro- matogram of the hydrolysis products of the carbohydrate fraction of cells grown on glucose. Similar results were obtained when the carbohy- drates were extracted from cells grown on lactose or galactose; in every case, substances migrating similarly to ribose, arabinose, glucose, and ga- lactose were found in the hydrolysate. Moreover, the ratio of the result- ant sugars, as indicated by the intensity of the spots on the chromatogram, was similar and independent of the carbon source. In certain experiments, small amounts of a component migrating at the same rate as lactose ap- peared. It was shown that this component accumulated during the hydroIysis procedure. Thus in Fig, 2 this compound was present on the chromatogram of the fraction hydrolyzed for 15 minutes but was not detected in the unhydrolyzed sample or when that sample was hydrolyzed for 1 hour. To identify this sugar more positively, this substance was
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from
316 LACTOSE METABOLISM OF L. BULGARICUS
separated from others by paper chromatography, was hydrolyzed, and the products identified by rechromatographing. As indicated in Fig. 3, glucose and galactose were present in about equal proportions. This is considered
ACTOSE ALACT@ LUCOSE RABINOS ‘IBOSE
IXTRACTABLE POLYSAGCHA- RIDES - GLUCOSE ADAPTED
CELL:
JOT iYDRO- HYDRO, LYZED LYZEC HOUR
IYDRO- LYZED ) MINUTES
a
0
a
0
e
0
0
a
FIG. 2 FIG. 3 FIG. 2. Sugars liberated by acid hydrolysis of alcohol-precipitable carbohydrate
extracted from L. b&a&us Gere A. Dried cell preparations were extracted with 5 parts of 15 per cent trichloroacetic acid (TCA), followed by three extractions with 5 parts of 10 per cent TCA. These fractions were combined and 2 volumes of alcohol were added. The precipitate was collected by centrifugation, washed with 50 per cent alcohol, and dried. Suitable fractions were hydrolyzed as shown by heating with 1 N HzS04 at 100” for the time indicated. Identification of the sugars was made by paper chromatography after neutralization and precipitation of the sulfate with barium hydroxide. The spots appearing on the original chromatograms were out- lined and their relative intensity approximated for reproduction. High, intermedi- ate, and weak intensities are indicated by solid, shaded, and open spots respectively.
L ACTOSE Gt ALACTOS G LUCOSE
YDRO- -YZED .ACTOSE SPOT
FIG. 3. Hydrolysis products of the compound which accumulates on partial hy- drolysis of alcohol-precipitabIe carbohydrates isolated from L. bulgaricus Gere A. This fraction was separated from other sugar components by paper chromatography, extracted from the paper with water, hydrolyzed 1 hour in 1 N HS04 at 100”. After neutralization and precipitation of sulfate with barium hydroxide, the fraction was concentrated by lyophilization. The sugars were extracted into pyridine and chro- matographed.
strong evidence for the existence of lactose as a constituent of a more complex substance present in both unadapted L. bulgaricus Gere A and in cells modified to grow on glucose and galactose.
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from
W. J. RUTTER AND R. G. HANSEN 317
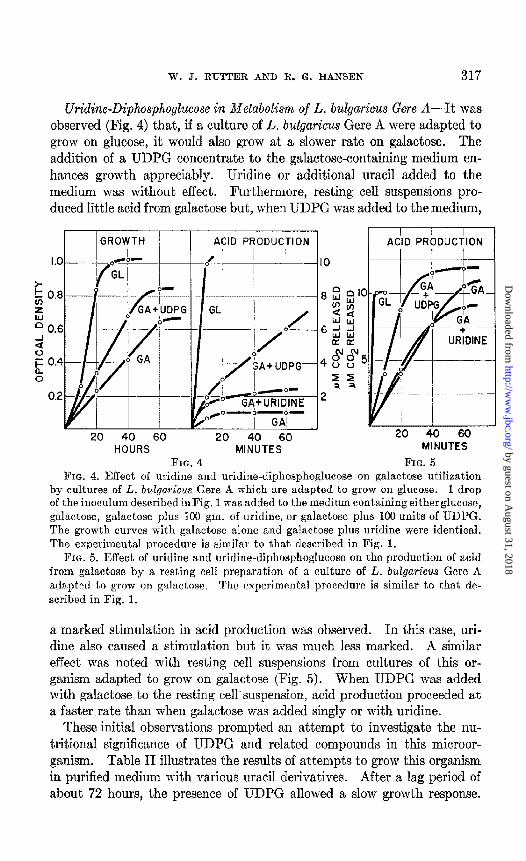
Uridine-Diphosphogkose in Metabolism of L. bulgaricus Gere A-It was observed (Fig. 4) that, if a culture of L. bulgaricus Gere A were adapted to grow on glucose, it would also grow at a slower rate on galactose. The addition of a UDPG concentrate to the galactose-containing medium en- hances growth appreciably. Uridine or additional uracil added to the medium was without effect. Furthermore, resting cell suspensions pro- duced little acid from galactose but, when UDPG was added to the medium,
GROWTH ~ ACID PRODUCTION ( A+lD PR~DUCjlON 1
20 40 60 20 40 60 HOURS MINUTES
11 URiDlNE 1
20 40 60 MINUTES
FIG. 4 FIG. 5 E’IG. 4. Effect of uridine and uridine-diphosphoglucose on galactose utilization
by cultures of L. 6ulgariczLs Gere A which are adapted to grow on glucose. 1 drop of the inoculum described inFig. 1 was added to the medium containing eitherglucose, galactosc, galactose plus 100 gm. of uridine, or galactose plus 100 units of UDPG. The growth cumes with galactose alone and galactose plus uridine were identical. The experimental procedure is similar to that described in Fig. 1.
FIG. 5. Effect of uridine and uridine-diphosphoglucose on the production of acid from galactose by a resting cell preparation of a culture of L. bulgaricus Gere A a&v-ltxd tn I)*rnw on !gRlaCtOBP. Th- r--- -- 0--.. P f=xnw-imental nrncednre iq Gmi1fl.r to that de- -..=-.. .~......~ I~ .---.~~- -i ...-----.
scribed in Fig. 1.
a marked stimulation in acid production was observed. In this case, uri- dine also caused a stimulation but it was much less marked. A similar effect was noted with resting cell suspensions from cultures of this or- ganism adapted to grow on galactose (Fig. 5). When UDPG was added with galactose to the resting cell’suspension, acid production proceeded at a faster rate than when galactose was added singly or with uridine.
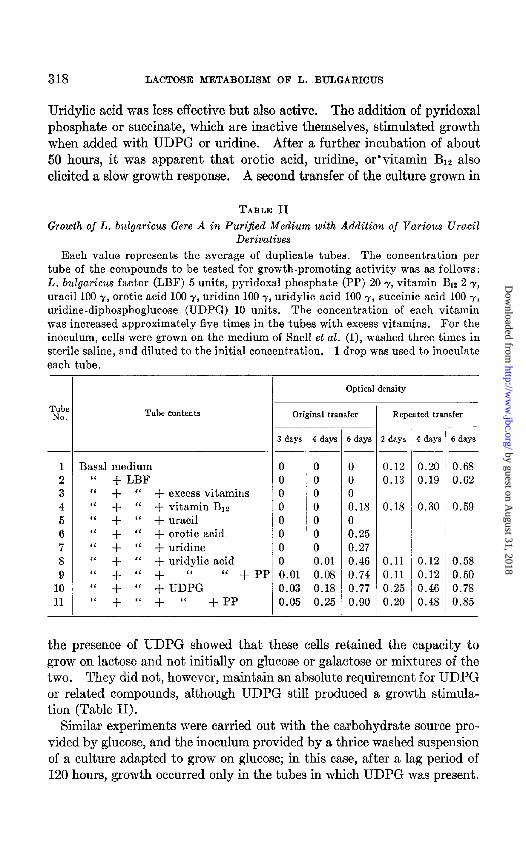
These initial observations prompted an attempt to investigate the nu- tritional significance of UDPG and related compounds in this microor- ganism . Table II illustrates the results of attempts to grow this organism in purified medium with various uraci1 derivatives. After a lag period of about 72 hours, the presence of UDPG allowed a slow growth response.
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from
318 LACTOSE METABOLISM OF L. BULGARICUS
Uridylic acid was less effective but also active. The addition of pyridoxal phosphate or succinate, which are inactive themselves, stimulated growth when added with UDPG or uridine. After a further incubation of about 50 hours, it was apparent that erotic acid, uridine, or’vitamin Bit also elicited a slow growth response. A second transfer of the culture grown in
TABLE II Growth of L. bulgaricus Gere A in Purified Medium with Addition of Various Uracil
Derivatives Each value represents the average of duplicate tubes. The concentration per
tube of the compounds to be tested for growth-promoting activity was as foIlows: L. bulgaricus factor (LBF) 5 units, pyridoxal phosphate (PP) 20 y, vitamin Brz 2 y, uracil 100 y, erotic acid 100 y, uridine 100 y, uridylic acid 100 y, succinic acid 100 7, uridine-diphosphoglucose (UDPG) 10 units. The concentration of each vitamin
was increased approximately five times in the tubes with excess vitamins. For the inoculum, cells were grown on the medium of Snell et aZ. (I), washed three times in sterile saline, and diluted to the initial concentration. 1 drop was used to inoculate each tube.
1 Basal medium 2 “ + LBF 3 “ + “ + excess vitamins 4 “ + “ + vitamin Biz 5 ‘< + “ + uracil 6 ‘I + “ + erotic acid 7 “ + “ + uridine 8 “ + “ + uridylic acid 9 “ + ‘( + (‘ “ +PI
10 “ -I- ‘< + UDPG 11 “ + “ + “ + pp
Tube contents Original transfer
-
,
-
Optical density
3 days 4 days 6 days 2 days
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.01 0.01 0.08 0.03 0.18 0.05 0.25
- I
0 0 0 0.18 0 0.25 0.27 0.46 0.74 0.77 0.90
0.12 0.20 0.13 0.19
0.18 0.30
0.11 0.12 0.11 0.12 0.25 0.46 0.20 , 0.48 1 -
Repeated transfer
4 days 6 days
0.68 0.62
0.59
0.58 0.50 0.78 0.85
the presence of UDPG showed that these cells retained the capacity to grow on lactose and not initially on glucose or galactose or mixtures of the two. They did not, however, maintain an absolute requirement for UDPG or related compounds, although UDPG still produced a growth stimula- tion (Table II).
Similar experiments were carried out with the carbohydrate source pro- vided by glucose, and the inoculum provided by a thrice washed suspension of a culture adapted to grow on glucose; in this case, after a lag period of 120 hours, growth occurred only in the tubes in which UDPG was present.
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from

W. J. RUTTER AND R. G. HANSEN 319
DISCUSSION
The observations presented here indicate that with L. bdguricus Gere A the ability of a cell to metabolize a sugar readily does not necessarily mean that that cell can utilize this sugar for growth. Modification of the culture to permit growth on glucose or galactose could perhaps involve other sys- tems than those directly concerned with metabolism for energy purposes. The similarity of the complex carbohydrates extracted from unadapted cells and from eelIs adapted to grow upon glucose or galactose suggests, as one possibility, that the formation of a polysaccharide (perhaps a morpho- logical constituent of the cell) is also essential for growth. Thus, the un- adapted cell would produce this polysaccharide from lactose or certain P-galactosides by enzymes, the specificity of which would not allow the participation of glucose or galactose. Adaptation to growth on glucose or galactose would then primarily be the production or modification of en- zymes within the cell to utilize these compounds in the synthesis of the essential carbohydrate components.
Although the enzymes necessary to utilize lactose by way of its hy- drolysis products were found in cellular extracts of unadapted L. bdguricus Gere A, the existence of a phosphorolysis or another “direct” manner of lactose utilization and synthesis is not precluded by these observations. Since lactose was present, at least in a combined form, in the carbohydrates of cells adapted to glucose or galactose, its synthesis is indicated. The fact that resting cell suspension of this organism which utilizes p-galactosides produced acid from a-galactose-l-phosphate is interesting in this regard. Previously it has been suggested that /3-galactose-l-phosphate would be the logical precursor of lactose (21). A study of the stereochemical specific- ity of enzymes acting upon galactose in these extracts may lend informa- tion concerning this process.
The unexpected observation that lactose is present in a combined form in cells adapted to grow on glucose and galactose is interesting in regard to the findings of Caputto and Trucco (22), Malpress and Morrison (23), and Rutter and Hansen (24) relating to the isolation of complex substances containing lactose from the mammary gland. The hydrolysis procedures used in the experiments reported herein permitted only minimal recovery of lactose. Therefore, much of the glucose and galactose observed to ac- cumulate on hydrolysis of the cehular “polysaccharides” might have orig- inally been combined in lactose units. The significance of the existence of this disaccharide in cellular carbohydrates is not as yet completely under- stood.
Since UDPG stimulated acid production from galactose by cell suspen- sions, it is considered that this compound is involved in the utilization of galactose by this microorganism. This fact is understandable since UDPG
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from

320 LACTOSE METABOLISM OF L. BULGARICUS
is the coenzyme of galactowaldenase which facilitates the conversion of galactose-l-phosphate to glucose-l-phosphate (20). With regard to the experiments on the growth of this microorganism in a purified medium, during the long lag period before growth started undoubtedly modification or selection of the culture occurred. Moreover, for an optimal growth rate other factors were apparently needed, but nevertheless, under these con- ditions at least, UDPG or less effectively uridylic acid initiated growth in a synthetic medium. It appears that either another adaptation or the selection of a variant found still less frequently was necessary for the inoculum to grow in the presence of uridine or vitamin Blz. Since uracil itself did not initiate growth in the synthetic medium, vitamin B1z may be operative, at least in part, in synthesis beyond the pyrimidine moiety.
It is not clear whether the function of UDPG or compounds which have close structural similarity is restricted to the providing of the coenzyme for the conversion of galactose-l-phosphate to glucose-l-phosphate, or whether these compounds have a more general significance in this microorganism. The fact that UDPG is also required by this organism when adapted to and grown on glucose could indicate the importance of galactose or lactose or mixtures of the two even in the adapted organism, or another function of UDPG and related compounds in this organism, or both. Since the uridine-diphosphate derivatives isolated by Park in XtaphyZococcus uureus (25-27) contained peptides, the possibility is allowed that these compounds may also operate in peptide linkage synthesis. The recent reports of Kihara et al. (28) and Kihara and Snell (29) on the requirements of peptides for growth of LactobaciEZus casei make it tempting to wonder whether this or- ganism has a peptide requirement which could alleviate the apparent re- quirement of UDPG and related compounds of initiation of growth.
SUMMARY
1. Resting cell suspensions of cultures of Lactobacillus bulgaricus strain Gere A, which would grow on lactose but not glucose and galactose, actively produced acid when incubated with glucose or galactose, or mixtures of the two, or ar-galactose-l-phosphate, but not from cy- or B-glucose-l-phos- phate, ,&galactose-l-phosphate, or glucose-6-phosphate. Unadapted cul- tures of this organism could not utilize any of the QI- or ,!%galactose-l-phos- phates or Q- or P-glucose-l-phosphates for growth.
2. Attempts to identify an enzyme in these extracts which could phos- phorylize lactose were unsuccessful. Cell-free extracts of unadapted L. buZguricus (growth on lactose only) contained lactase activity, galactokin- ase activity, and hexokinase activity.
3. Hydrolysates of alcohol-precipitable carbohydrates extracted from various cultures of L. bulguricus Gere A contained ribose, arabinose, glu- cose, galactose, and lactose.
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from

W. .J. RUTTER AND R. 0. HANSEN 321
4. UDPG stimulated the production of acid from galactose by resting cell suspensions of L. bulgaricus Gere A which had been modified to grow on glucose or galactose. UDPG stimulated growth of cultures of this microorganism adapted to grow on glucose when grown in a partially puri- fied medium with galactose as the carbohydrate source.
5. UDPG or less effectively uridylic acid, especially in the presence of pyridoxal phosphate, initiated growth in a completely synthetic medium but only after a lag period. After a longer period of incubation, uridine, erotic acid, or vitamin Blz elicited growth of this organism. Uracil was without effect under the conditions tested.
BIBLIOGRAPHY
1. Snell, E. E., Xitay, E., and Hoff-JGrgensen, E., Arch. Biochem., 18, 495 (1948). 2. Leibowitz, J., and Hestrin, S., Advances in Enzymol., 5,87 (1945). 3. Hassid, W. Z., and Doudoroff, M., Advances in Enzymol., 10, 123 (1950). 4. Reithel, F. J., J. Am. Chem. Xoc., 67, 1056 (1945). 5. Kosterlite, H. W., Biochem. J., 33, 1087 (1939). 6. Levene, P. A., and Raymond, A. L., J. Biol. Chem., 92, 765 (1931). 7. Caputto, R., Leloir, L. F., Cardini, C. E., and Paladini, A. C., J. Biol. Chem.,
184, 333 (1956). 8. Paladini, A. C., and Leloir, L. F., Biochem. J., 51, 426 (1952). 9. Najjar, V. A., J. Biol. Chem., 175, 281 (1948).
10. Kornberg, A., J. Biol. Chem., 183,805 (1950). 11. Fiske, C. H., and Subbarow, Y., J. Biol. Chem., 66, 375 (1925). 12. Caputto, R., Leloir, L. F., and Trucco, R. E., Enxymologiu, 14, 350 (1948). 13. Somogyi, M., J. BioZ. Chem., 196, 19 (1952). 14. Nelson, N., J. Biol. Chem., 153, 375 (1944). 15. Trevelyan, W. E., and Harrison, J. S., Biochem. J., 60, 298 (1951). 16. Jermyn, M. A., and Isherwood, F. A., Biochem. J., 44,402 (1949). 17. Partridge, S. M., Biochemical Society Symposia, Cambridge, 3, 57 (1950). 18. Doudoroff, M., Barker, H. A., and Hassid, W. Z., J. BioZ. Chem., 168,725 (1947). 19. Fitting, C., and Doudoroff, M., Federation PTOC., 11, 212 (1952). 20. Caputto, R., Leloir, L. F., Trucco, R. E., Cardini, C. E., and Paladini, A. C.,
J. Biol. Chem., 179, 497 (1949). 21. Reithel, F. J., Horowitz, M. G., Davidson, H. M., and Kittinger, G. W., J. Biol.
Chem., 194, 839 (1952). 22. Caputto, R., and Trucco, R., Nature, 169, 1061 (1952). 23. Malpress, F. H., and Morrison, A. B., Nature, 169, 1103 (1952). 24. Rutter, W. J., and Hansen, R. G., J. Dairy SC., in press. 25. Park, J. T., J. BioZ. Chem., 194, 877 (1952). 26. Park, J. T., J. BioZ. Chem., 194,885 (1952). 27. Park, J. T., J. BioZ. Chem., 194, 897 (1952). 28. Kihara, H., McCullough, W. G., and Snell, E. E., J. Bid. Chem., 197,783 (1952). 29. Kihara, H., and Snell, E. E., J. BioZ. Chem., 197, 791 (1952).
by guest on August 31, 2018
http://ww
w.jbc.org/
Dow
nloaded from

W. J. Rutter and R. G. HansenSTRAIN GERE A
LACTOBACILLUS BULGARICUSCARBOHYDRATE METABOLISM OF
LACTOSE METABOLISM: I.
1953, 202:311-321.J. Biol. Chem.
http://www.jbc.org/content/202/1/311.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/202/1/311.citation.full.haccessed free atThis article cites 0 references, 0 of which can be by guest on A
ugust 31, 2018http://w
ww
.jbc.org/D
ownloaded from





























