Embed Size (px)
Citation preview

Lateral Development and Florogenesis of a Tropical Shallot: A Comparison with Bulb OnionAuthor(s): Yoram Krontal, Rina Kamenetsky and Haim D. RabinowitchSource: International Journal of Plant Sciences, Vol. 159, No. 1 (Jan., 1998), pp. 57-64Published by: The University of Chicago PressStable URL: http://www.jstor.org/stable/2474935 .
Accessed: 15/07/2014 13:43
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access toInternational Journal of Plant Sciences.
http://www.jstor.org
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions

Int. J. Plant Sci. 159(l):57-64. 1998. C) 1998 by The University of Chicago. All rights reserved. 1058-5893/98/590 1 -0007$03.00
LATERAL DEVELOPMENT AND FLOROGENESIS OF A TROPICAL SHALLOT: A COMPARISON WITH BULB ONION
Yoram Krontal,* Rina Kamenetsky,' t and Haim D. Rabinowitch*
*Faculty of Agriculture, Department of Field Crops, Vegetables, and Genetics, Hebrew University of Jerusalem, P.O. Box 12, Rehovot 76100, Israel; and tDepartment of Ornamental Horticulture, ARO,
Volcani Center, P.O. Box 6, Bet Dagan 50250, Israel
The developmental morphology of laterals and flowers was studied in shallot. Growth analyses and microscopic observations revealed strong similarities in organ development between shallots and bulb onions. However, some significant differences were observed, mainly with regard to the minimum physiological age for both laterals and floral differentiation and to the development of the inflorescence. When grown from seed, the initiation of laterals and the first signs of floral differentiation become evident already after the third and sixth true leaves, respectively, as compared to the bulb onion, which first produces 13 or 10-14 leaves, respectively. The shallot inflorescence can be described as an umbel-like flower arrangement, where branches (flower clusters) arise from a common meristem. Leaf formation continues at the axillary meristems in shallots simultaneously with floral development of the original apex. In blooming onion, however, the lateral meristem develops into an adventitious dormant bud. A detailed description of shallot florogenesis and the similarities and differences between blooming shallot and bulb onion are discussed.
Introduction Great morphological variability within the Allium
cepa L. taxon has led to infraspecific groupings of the species. Hence, Helm (1956; cited by Hanelt 1990) made a clear distinction between the following four botanical varieties: var. cepa (common onion); var. vi- viparum (top onion); var. aggregatum (potato or mul- tiplier onion); and the shallot-like var. cepiforme. Lat- er, Jones and Mann (1963) subdivided the onion alliance into three horticultural groups: the common onion group, the aggregatum group (potato onion, ever-ready onion, and shallot), and the proliferum group (top onion).
Shallots intercross freely with the bulb onion to pro- duce fertile hybrids (Hanelt 1990; Messiaen et al. 1993) and there is a strong cytological (Kalkman 1984a, 1984b) and morphological resemblance be- tween the two alliaceous crops. Therefore, Hanelt (1990) reunified the common onion and shallot into one botanical species A. cepa, subdivided into two large groups: common onion (synonyms: A. cepa L. var. cepa; A. cepa L. ssp. cepa and ssp. australe Tro- fim.) and aggregatum (synonyms: A. ascalonicum auct. non Strand; A. cepa ssp. orientale Kazak.; A. cepa var. ascalonicum Baker).
The bulbs of shallot are usually smaller than those of the single-hearted bulb onion. They contain a num- ber of vegetative lateral buds formed in the axils of some of the inner blade-bearing leaves. These laterals remain attached to the original basal plate, thus form- ing a cluster of bulbils (Jones and Mann 1963; Currah and Proctor 1990; Messiaen et al. 1993).
Shallots are cultivated in southeast Asia and some African countries, where onion seed is difficult to pro- duce, onion culture is difficult, and/or the growing sea- son is too short for the production of bulb onion (Cur-
'Author for correspondence and reprints.
Manuscript received December 1996; revised manuscript received August 1997.
rah and Proctor 1990). In addition, because of their unique flavor (Aura 1963; Messiaen et al. 1993), a few long- and intermediate-day selections are popular in some European countries (Messiaen et al. 1993), in the United States (Jones and Mann 1963), and in South America (e.g., Argentina) (H. D. Rabinowitch, person- al observation). Even where seed production is pos- sible, most of these shallot cultivars are clonally prop- agated to maintain the unique quality traits and population homogeneity of the highly heterozygous genotypes. However, in temperate or subtropical regions, bolting is common, and consequently both yield and quality are reduced. Shallot hybrids have re- cently been released by breeders in Israel (H. D. Ra- binowitch) and Holland (Bejo Seed Co.). Therefore, the conditions and processes leading to flowering are of considerable economic importance.
Many researchers described the Allium spp. inflo- rescence as a terminal umbel (Vvedensky 1935; De Mason 1990; Brewster 1994). Louis and Auguste Bra- vais (1887) were the first to study the morphology of Allium inflorescence. They characterized A. moly (sub- genus Amerallium) inflorescences as having two sym- podial flower clusters arranged one on top of the other (Mann 1959). Later, Weber (1929) argued that the in- florescence of A. odorum (subgenus Rhizirideum) be- gins with a terminal flower, which is enveloped by two bracts, each having an axillary flower. This dichasial branching may continue, giving rise to two additional lateral flowers. Modern microscopic studies of A. ro- thii, A. altissimum, and A. karataviense (subgenus Me- lanocrommyum) florogenesis revealed that the differ- entiated apical meristem divides into several peripheral swellings, each of which produces a row of flower primordia (a cluster). This umbel-like inflorescence produces branches that arise from a common meristem (Kamenetsky 1994; Kamenetsky and Japarova 1997).
Floral induction in the bulb onion may occur both during the growing period and during storage of the developed bulbs, provided the plants have reached the
57
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions

58 INTERNATIONAL JOURNAL OF PLANT SCIENCES
Table 1 Sampling Dates and Physiological Stages of Shallot Plants
Sampling date Site and physiological stage
Sept. 11, 1995 ............... Storage/rest period Oct. 5-8, 1995 ............... Field/initial sprouting Oct. 22-25, 1995 ............ Field/3-4 leaves Nov. 26-Dec. 2, 1995 ...... Field/initial flowering, only a few
floral buds visible Dec. 31, 1995 ................ Field/mass flowering, all plants pro-
duce at least one visible floral bud
physiological age of 12 (range of 10-14) leaves (Ra- binowitch 1990). Jones and Emsweller (1936) and Hartsema (1947) described the umbel of A. cepa (sub- genus Rhizirideum) as having a single external spathe containing an enlarged shoot apex and two alternating bracts. The flat apical surface is subdivided into many centers, each made of a spirally arranged cluster of flowers. Thus, the inflorescence, with 50-2000 flowers (Jones and Mann 1963), is made up of a great number of clusters. The few studies on the flowering process in shallot indicate a morphological resemblance to the bulb onion (Messiaen et al. 1993). However, Hartsema (1947) reported that floral initiation and development are scarce in stored shallot bulbs and occur mainly in growing plants. A similar response was reported for A. sphaerocephalon (subgenus Allium) (Berghoef and Zevenbergen 1992).
Little is known about the morphological events dur- ing apex differentiation of shallots and the formation of laterals or about floral initiation and differentiation. Here we describe the sequence of morphological pro- cesses occurring during branching and floral develop- ment of a short-day shallot selection from the tropics.
Material and Methods
Plant Material
Bulbs of a clonally propagated shallot landrace from Thai- land (accession no. 66-1004), and seeds of this clone (har- vested from open-pollinated umbels of ca. 100 caged bulbs), were received from the Field Gene Bank for Vegetatively Propagated Short-Day Allium spp., in Rehovot, Israel.
Growth Conditions
In May 1995, bulb clusters were harvested after their tops had fallen. The dried leaves were trimmed and the bulbs stored through the summer in an open shed under ambient conditions. Prior to planting, the clusters were separated, and large bulbs 20-35 mm in diameter were planted on October 2 on raised beds at a density of 20 plants m-2, at the Faculty of Agriculture Experimental Farm, in Rehovot. Seeds were sown on October 15 to a final stand of 50 seedlings m-2.
Standard agricultural practices were employed throughout.
Morphological Studies
Five to 10 random plants were harvested periodically for the morphogenetic studies, as detailed in table 1.
Preparations for SEM
As described by Kamenetsky (1994), freshly harvested plants were carefully stripped of their leaves, and spathes
were removed from young floral buds. Meristems were iso- lated under a stereomicroscope in small petri dishes contain- ing distilled water to prevent dehydration. The excised mer- istems were fixed in a 5: 5: 90 mixture of glacial acetic acid: formalin (40%): ethanol (70%) and dehydrated in a graded acetone series (35%, 70%, 90%, 100%) and freon. Imme- diately thereafter, tissues were dried using liquid CO, in a Biorad 750 (England) critical-point dryer. Samples were then mounted on SEM stubs with double-sided tape, sputter-coat- ed with ca. 10 nm gold, and studied in a JSM-35C SEM (JEOL, Japan) using an accelerating potential of 15 kV.
Observations and Discussion Bulb Development
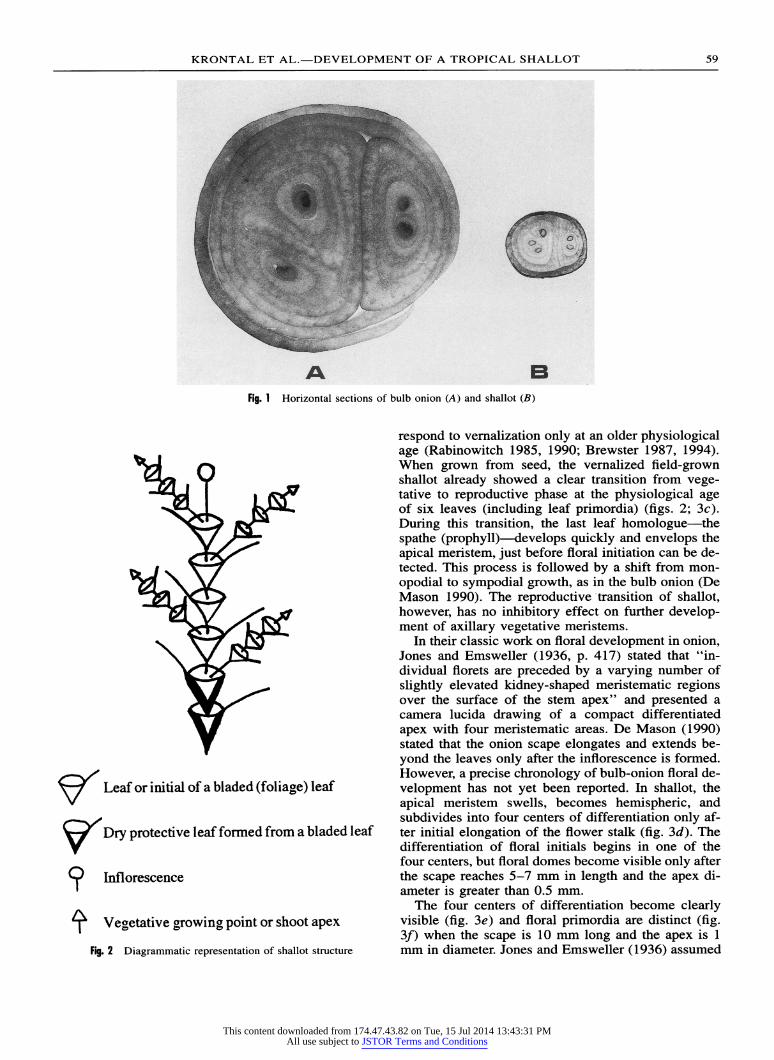
Shallots develop bulbs in response to long photo- periods, and different cultivars can be distinguished by the minimum daylength needed to induce them to bulb. Bulb development of shallots commences with the elongation and swelling of both leaf sheaths and young leaf buds, probably due to cell expansion as in the bulb onion (Brewster 1990, 1994). A single shallot bulb contains several shoot initials (figs. 1, 2), which resemble those of doubled onion (Rabinowitch 1979). During ripening, one to three of the outermost sheaths of both plants develop into dry, protective skins.
Leaf Formation
In shallot, leaf formation begins as a protrusion of the apical dome, which develops as a crescent. Each shallot leaf primordium differentiates at the apical meristem and subsequently develops into a partly lift- ed, complete ring (fig. 3a); the older leaf envelops the successively younger one in a way that is very similar to the development of leaves in the bulb onion (Jones and Mann 1963; De Mason 1990; Brewster 1994). When the meristematic buds elongate, they form al- ternate tubular leaves in an opposite position, as in the bulb onion (Heath and Holdsworth 1948).
Formation of Laterals
Initiation of the first lateral in the meristematic apex of shallot seedlings was already evident after the dif- ferentiation of the third leaf (figs. 2; 3b). This branch- ing is similar to that of chives (Allium schoenopras- um), where the initiation of lateral occurs every two or three leaves (Poulsen 1990). It is, however, mark- edly different from the bulb onion. When grown from seeds, onions often produce 13 or more leaves prior to doubling (Eto 1956; Rabinowitch 1979), and the lat- eral growing point develops into a dormant adventi- tious bud between the original shoot apex and the youngest leaf (Brewster 1994). In contrast, in shallot, lateral shoots and inflorescences develop in tandem with the main apex and terminal (principal) inflores- cence.
Florogenesis
Numerous experiments indicated a critical physio- logical stage of 12 (10-14) leaves (including leaf buds) prior to onion floral induction, and some bolting- resistant cultivars (e.g., Senshyu Semi-Globe Yellow)
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions
KRONTAL ET AL.-DEVELOPMENT OF A TROPICAL SHALLOT 59
A B Fig. 1 Horizontal sections of bulb onion (A) and shallot (B)
Leaf or initial of a bladed (foliage) leaf
s, Dry protective leaf formed from a bladed leaf
Ci) Inflorescence
L Vegetative growing point or shoot apex
Fig. 2 Diagrammatic representation of shallot structure
respond to vernalization only at an older physiological age (Rabinowitch 1985, 1990; Brewster 1987, 1994). When grown from seed, the vernalized field-grown shallot already showed a clear transition from vege- tative to reproductive phase at the physiological age of six leaves (including leaf primordia) (figs. 2; 3c). During this transition, the last leaf homologue-the spathe (prophyll)-develops quickly and envelops the apical meristem, just before floral initiation can be de- tected. This process is followed by a shift from mon- opodial to sympodial growth, as in the bulb onion (De Mason 1990). The reproductive transition of shallot, however, has no inhibitory effect on further develop- ment of axillary vegetative meristems.
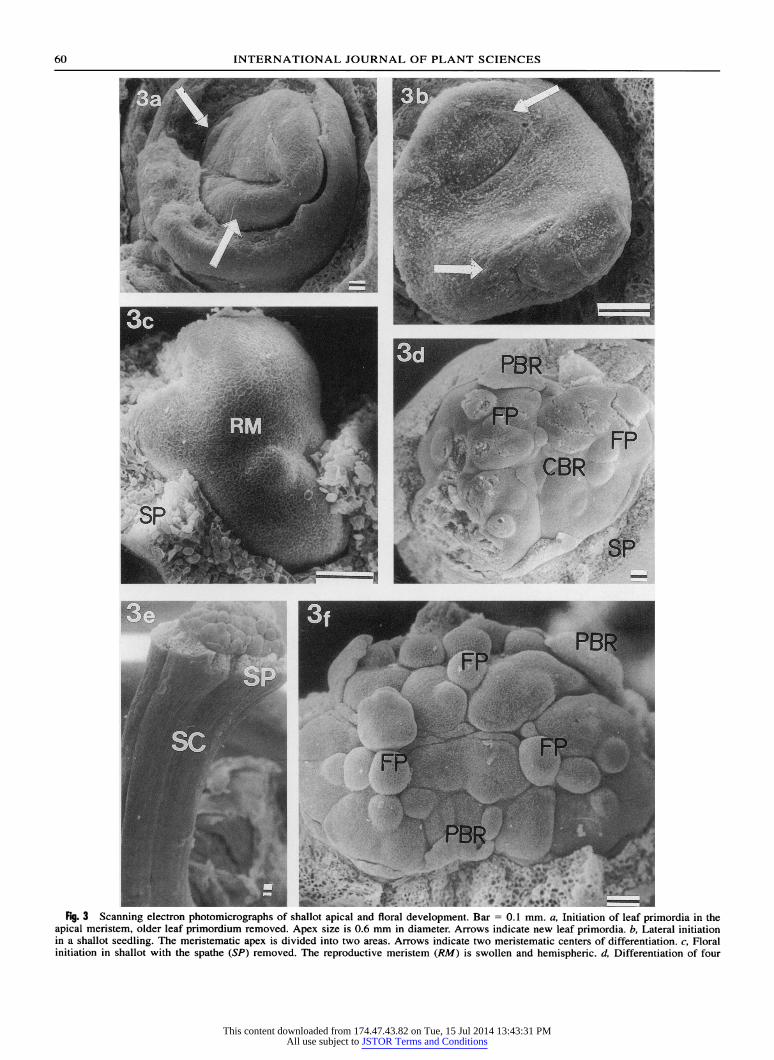
In their classic work on floral development in onion, Jones and Emsweller (1936, p. 417) stated that "in- dividual florets are preceded by a varying number of slightly elevated kidney-shaped meristematic regions over the surface of the stem apex" and presented a camera lucida drawing of a compact differentiated apex with four meristematic areas. De Mason (1990) stated that the onion scape elongates and extends be- yond the leaves only after the inflorescence is formed. However, a precise chronology of bulb-onion floral de- velopment has not yet been reported. In shallot, the apical meristem swells, becomes hemispheric, and subdivides into four centers of differentiation only af- ter initial elongation of the flower stalk (fig. 3d). The differentiation of floral initials begins in one of the four centers, but floral domes become visible only after the scape reaches 5-7 mm in length and the apex di- ameter is greater than 0.5 mm.
The four centers of differentiation become clearly visible (fig. 3e) and floral primordia are distinct (fig. 3f) when the scape is 10 mm long and the apex is 1 mm in diameter. Jones and Emsweller (1936) assumed
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions
60 INTERNATIONAL JOURNAL OF PLANT SCIENCES
a- ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ -M
AS
"* et,tA' Fig. 3 Scanning electron photomicrographs of shallot apical and floral development. Bar = 0.1 mm. a, Initiation of leaf primordia in the
apical meristem, older leaf pnimordium removed. Apex size is 0.6 mm in diameter. Arrows indicate new leaf primordia. b, Lateral initiation
in a shallot seedling. The menistematic apex is divided into two areas. Arrows indicate two meristematic centers of differentiation. c, Floral
initiation in shallot with the spathe (SP) removed. The reproductive meristem (RM) is swollen and hemispheric d. Differentiation of foulr
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions
KRONTAL ET AL.-DEVELOPMENT OF A TROPICAL SHALLOT 61
a fairly definite sequence of floral differentiation in the bulb onion. However, to the best of our knowledge, the exact sequence of development has never been de- termined, and, therefore, we cannot compare the initial morphological events that lead to floral development in these two crop plants.
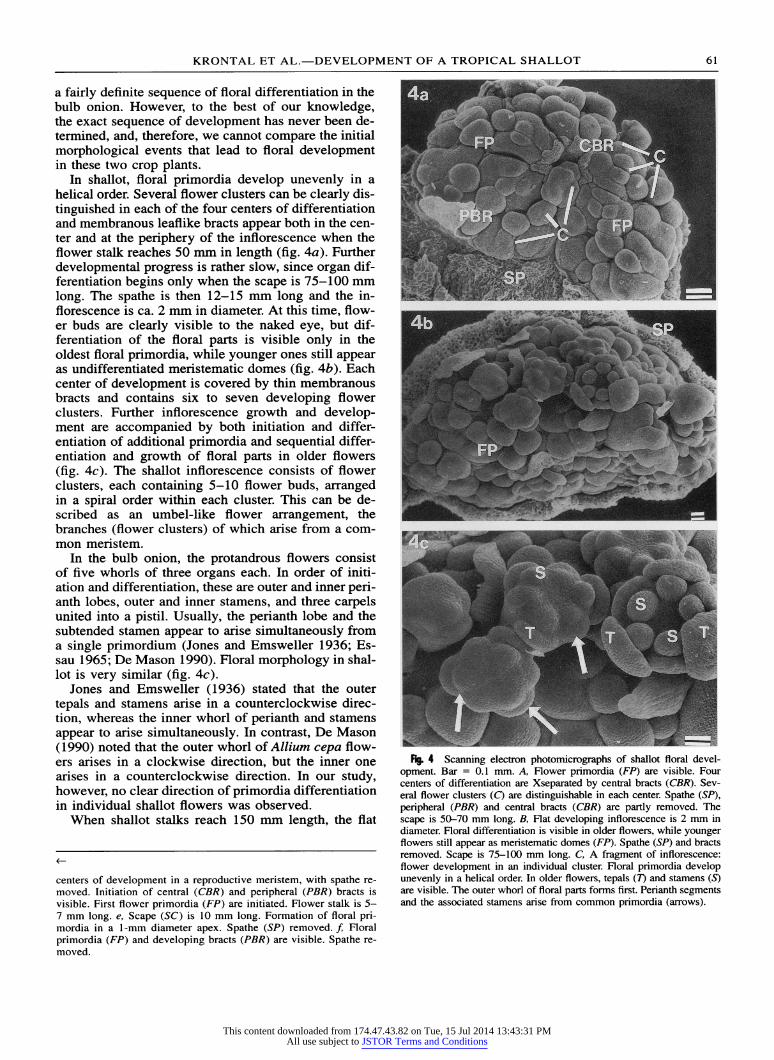
In shallot, floral primordia develop unevenly in a helical order. Several flower clusters can be clearly dis- tinguished in each of the four centers of differentiation and membranous leaflike bracts appear both in the cen- ter and at the periphery of the inflorescence when the flower stalk reaches 50 mm in length (fig. 4a). Further developmental progress is rather slow, since organ dif- ferentiation begins only when the scape is 75-100 mm long. The spathe is then 12-15 mm long and the in- florescence is ca. 2 mm in diameter. At this time, flow- er buds are clearly visible to the naked eye, but dif- ferentiation of the floral parts is visible only in the oldest floral primordia, while younger ones still appear as undifferentiated meristematic domes (fig. 4b). Each center of development is covered by thin membranous bracts and contains six to seven developing flower clusters. Further inflorescence growth and develop- ment are accompanied by both initiation and differ- entiation of additional primordia and sequential differ- entiation and growth of floral parts in older flowers (fig. 4c). The shallot inflorescence consists of flower clusters, each containing 5-10 flower buds, arranged in a spiral order within each cluster. This can be de- scribed as an umbel-like flower arrangement, the branches (flower clusters) of which arise from a com- mon meristem.
In the bulb onion, the protandrous flowers consist of five whorls of three organs each. In order of initi- ation and differentiation, these are outer and inner peri- anth lobes, outer and inner stamens, and three carpels united into a pistil. Usually, the perianth lobe and the subtended stamen appear to arise simultaneously from a single primordium (Jones and Emsweller 1936; Es- sau 1965; De Mason 1990). Floral morphology in shal- lot is very similar (fig. 4c).
Jones and Emsweller (1936) stated that the outer tepals and stamens arise in a counterclockwise direc- tion, whereas the inner whorl of perianth and stamens appear to arise simultaneously. In contrast, De Mason (1990) noted that the outer whorl of Allium cepa flow- ers arises in a clockwise direction, but the inner one arises in a counterclockwise direction. In our study, however, no clear direction of primordia differentiation in individual shallot flowers was observed.
When shallot stalks reach 150 mm length, the flat
centers of development in a reproductive meristem, with spathe re- moved. Initiation of central (CBR) and peripheral (PBR) bracts is visible. First flower primordia (FP) are initiated. Flower stalk is 5- 7 mm long. e, Scape (SC) is 10 mm long. Formation of floral pri- mordia in a 1-mm diameter apex. Spathe (SP) removed. f Floral primordia (FP) and developing bracts (PBR) are visible. Spathe re- moved.
-~~~~~~~~ L-
-~~~~~~~~~~~ '5
14:
Fi 4 Scanning electron photomicrographs of shallot floral devel- opment. Bar = 0.1 mm. A, Flower primordia (FP) are visible. Four centers of differentiation are Xseparated by central bracts (CBR). Sev- eral flower clusters (C) are distinguishable in each center. Spathe (SP), peripheral (PBR) and central bracts (CBR) are partly removed. The scape is 50-70 mm long. B, Flat developing inflorescence is 2 mm in diameter. Floral differentiation is visible in older flowers, while younger flowers still appear as meristematic domes (FP). Spathe (SP) and bracts removed. Scape is 75-100 mm long. C, A fragment of inflorescence: flower development in an individual cluster. Floral primordia develop unevenly in a helical order. In older flowers, tepals (T) and stamens (S) are visible. The outer whorl of floral parts forms first. Perianth segments and the associated stamens arise from common primordia (arrows).
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions
62 INTERNATIONAL JOURNAL OF PLANT SCIENCES
-.j~ ~ ~ ~ ~ ~ ~~~~~~p
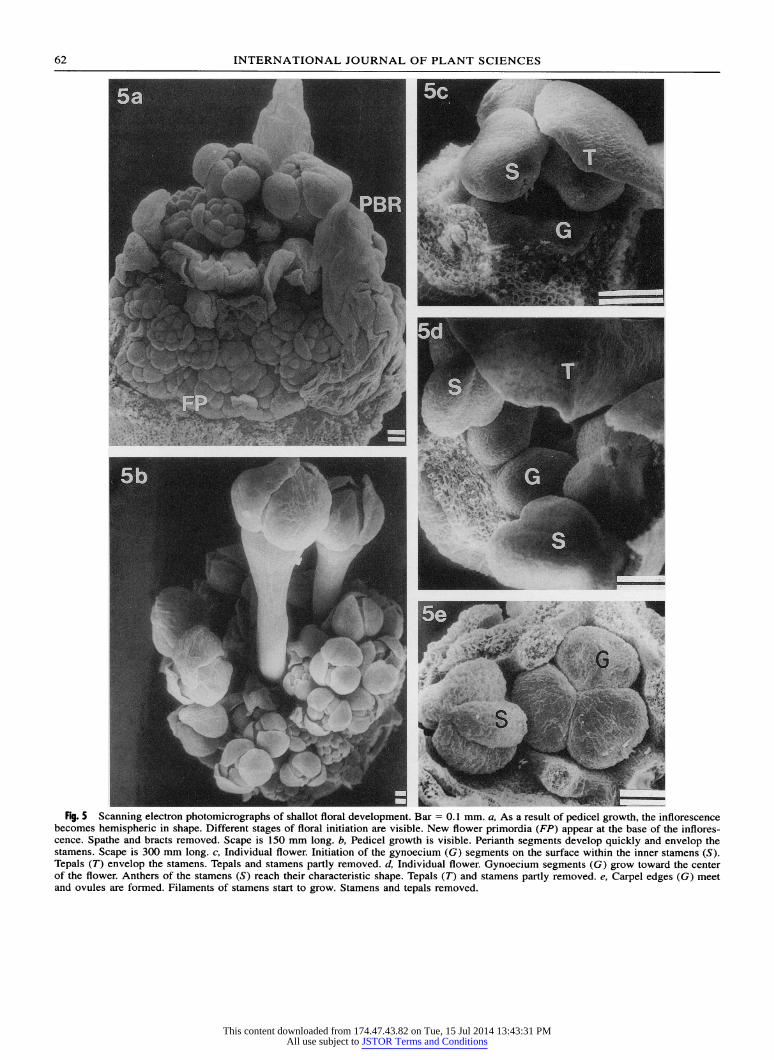
Fig. 5 Scanning electron photomicrographs of shallot floral development. Bar =0.1I mm. a, As a result of pedicel growth, the inflorescence becomes hemispheric in shape. Different stages of floral initiation are visible. New flower primordia (FP) appear at the base of the inflores- cence. Spathe and bracts removed. Scape is 150 mm long. b, Pedicel growth is visible. Perianth segments develop quickly and envelop the stamens. Scape is 300 mm long. c, Individual flower. Initiation of the gynoecium (G) segments on the surface within the inner stamens (S). Tepals (T) envelop the stamens. Tepals and stamens partly removed. d, Individual flower. Gynoecium segments (G) grow toward the center of the flower. Anthers of the stamens (S) reach their characteristic shape. Tepals (T) and stamens partly removed. e, Carpel edges (G) meet and ovuiles are formed. Filame-nts of stame-ns start to gFrow. Stamens andn tepals1 irremvted.
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions

KRONTAL ET AL. DEVELOPMENT OF A TROPICAL SHALLOT 63
inflorescence becomes spherical. The number and size of differentiated shallot flowers increase with time, and new undifferentiated domes of 0.15 mm in diameter become visible at the base of the inflorescence (fig. 5a). Flower formation is almost complete when the stalk is 300 mm long. At this time, pedicels of older flower are 1.6 mm long (fig. Sb), and the gynoecium segments are visible in most flowers. The carpels de- velop as three protruding areas within the inner sta- mens (fig. 5c, 5d) and meet at the heart of the flower to form the trilocular ovary. Concomitantly, the an- thers-0.3 mm in diameter-reach their characteristic shape (fig. Se), and the apex of the fused ovary de- velops into the style, which continues to elongate, even after anthesis.
Under our growth conditions, the flower stalk of the studied shallot becomes 35-50 cm long before the spathe opens, whereas onion scapes at a similar phys- iological stage reach 1-1.8 m in length. At bloom, all pedicels become almost equal in length, hence flower clusters can no longer be recognized in the shallot in- florescence.
The shallot studied appears to differ from the bulb onion in a number of developmental processes related to both branching and florogenesis. The main differ- ences are:
1. Physiological age to branching: When grown from seeds, laterals in the bulb onion begin to form only at the physiological age of 13 leaves, whereas shallot doubling begins much earlier, at or immediately after the three leaf stage of development, as in chives.
2. Minimum physiological age to floral initiation: When grown from seeds, floral initiation only occurs in the bulb onion at the postjuvenile age of 10-14 leaves, whereas the critical leaf number in the shallot studied here was 6.
3. Inflorescence development: The bulb onion inflo- rescence develops from the stem apex as a slightly
raised, kidney-shaped area with several meristematic centers. The exact sequence of flower development is not known. In shallot, flower-stalk elongation precedes the swelling of the apical meristem and its subdivision into four centers of floral development. Within clus- ters, floral primordia develop unevenly in a helical or- der. Shallot inflorescences can be described as an um- bel-like flower arrangement, the branches (flower clusters) of which arise from a common meristem.
4. Individual floral development: The onion flower consists of five whorls of floral organs, each having three members. The floral morphology of shallot is rather similar; however, in our study no clear direction of flower-part differentiation was observed.
5. Flower-stalk development: In the bulb onion, "only when the inflorescence is formed does the true stem elongate through the leaf sheathes and extend be- yond the other parts of the plants" (De Mason 1990, p. 28), whereas in shallot, floral-apex differentiation occurs only after the beginning of flower stalk elon- gation, and flower development is almost complete only when the stalk reaches 300 mm in length.
6. The growth and development of the bulb onion inflorescence is rarely followed by the development of auxiliary vegetative shoot. In shallot, the terminal (principal) inflorescence develops simultaneously with an auxiliary vegetative shoots and additional inflores- cences.
Conclusions There are many strong morphological and devel-
opmental similarities between the bulb onion and shal- lot. Nevertheless, we observed a number of distinct differences between the two alliaceous crops. It is not clear whether these differences reflect normal variation within the Allium cepa gene pool or whether they jus- tify the species or subspecies division suggested by Kazakova (1978) and Hanelt (1990).
Literature Cited
Aura K 1963 Studies of the vegetatively propagated onions culti- vated in Finland, with special reference to flowering and storage. Ann Agric Fenn 5(suppl):1-74
Berghoef J, AP Zevenbergen 1992 Effects of environmental con- ditions on flower initiation and development of Allium sphaero- cephalon L. Horticulture 325:91-96.
Brewster JL 1987 Vernalization of the onion: a quantitative ap- proach. Pages 171-183 in JG Atherton, ed. The manipulation of flowering. Butterworths, London.
1990 Physiology of crop growth and bulbing. Pages 53-88 in HD Rabinowitch, JL Brewster, eds. Onions and allied crops. Vol 1. Botany, physiology, and genetics. CRC Press, Boca Raton, Fla.
1994 Onions and other vegetable alliums. CAB Interna- tional, Wallingford, U.K. 236 pp.
Currah L, FS Proctor 1990 Onions in tropical regions. Natural Re- sources Institute Bulletin, Chatham Maritime, Kent, U.K. 232 pp.
De Mason DA 1990 Morphology and anatomy of Allium. Pages 27-51 in HD Rabinowitch, JL Brewster, eds. Onions and allied crops. Vol 1. Botany, physiology and genetics. CRC Press, Boca Raton, Fla.
Essau K 1965 Plant anatomy. 2d ed. Wiley, New York. 767 pp.
Eto K 1956 Studies of bolting onion. I. Relation between the flower bud formation and the bulb division. J Jpn Soc Hortic Sci 25:187- 193.
Hanelt P 1990 Taxonomy, evolution and history. Pages 1-26 in HD Rabinowitch, JL Brewster, eds. Onions and allied crops. Vol 1. Botany, physiology and genetics. CRC Press, Boca Raton, Fla.
Hartsema AM 1961 Influence of temperatures on flower formation and flowering of bulbous and tuberous plants. Pages 123-167 in W Ruhland, ed. Handout deer pflanzenphysiologie. Vol 16. Springer, Berlin.
1947 De periodieke ontwikkeling van Allium cepa L. vaz. Zittauer Riesen. Mededeel Landbouwhoogeschool 48:265-300.
Heath OVS, M Holdsworth 1948 Morphogenic factors as exempli- fied by the onion plant. Symp Soc Exp Biol 2:326-350.
Jones HA, SL Emsweller 1936 Development of the flower and ma- crogametophyte of Allium cepa. Hilgardia 10:415-428.
Jones HA, LK Mann 1963 Onions and their allies: botany, culti- vation, and utilization. Interscience, New York. 286 pp.
Kalkman ER 1984a Analysis of the C-banded karyotype of Alliunm cepa L. Standard system of nomenclature and polymorphism. Ge- netica 65:141-148.
1984b Cytotaxonomic studies in the genus Allium. useful-
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions

64 INTERNATIONAL JOURNAL OF PLANT SCIENCES
ness of C-banding for description and classification. Eucarpia 3d Allium Symposium. Wageningen, 74-77.
Kamenetsky R 1994 Life cycle, flower initiation and propagation of the desert geophyte Allium rothii. Int J Plant Sci 155:597-605.
Kamenetsky R, N Japarova 1997 Relationship between annual cy- cle and floral development of three Allium species from subgenus Melanocrommyum. J Arid Environ 35:473-485.
Mann L 1959 The Allium inflorescence: some species of the section Molium. Am J Bot 46:730-739.
Messiaen CM, J Cohat, JP Leroux, M Pichon, A Beyries 1993 Les Allium alimentary reproduce par vie vegetative. INRA, Paris. 228 pp.
Poulsen NI 1990 Chives Allium schoenoprasum L. Pages 213-250 in JL Brewster, HD Rabinowitch, eds. Onions and allied crops.
Vol 3. Biochemistry, food science, and minor crops. CRC Press, Boca Raton, Fla.
Rabinowitch HD 1979 Doubling of onion bulbs as affected by size and planting date of sets. Ann Appl Biol 93:63-65.
1985 Onions and other edible Alliums. Pages 398-409 in AH Halevy, ed. Handbook of flowering. Vol 1. CRC Press, Boca Raton, Fla.
1990 Physiology of flowering. Pages 113-134 in HD Ra- binowitch, JL Brewster, eds. Onions and allied crops. Vol 1. Bot- any, physiology and genetics. CRC Press, Boca Raton, Fla.
Vvedensky Al 1935 Luk: Allium L. Pages 112-141 in VL Koma- rov, ed. Flora SSSR. Vol 4. Akademia Nauk SSSR, Leningrad.
Weber E 1929 Entwicklungsgeschichtliche untersuchungen uber die Gattung Allium. Bot Arch 25:1-44.
This content downloaded from 174.47.43.82 on Tue, 15 Jul 2014 13:43:31 PMAll use subject to JSTOR Terms and Conditions





























