Embed Size (px)
Citation preview

Based on Profs. Kevin Gardner & Reza Khayat 1
Biochemistry - I
Mondays and Wednesdays 9:30-10:45 AM (MR-1307)
SPRING 2017
Lecture 15

Outline
•Bioenergetics •Fates of glucose •Glycolysis •Feeder pathways for glycolysis
• Why learn about Glycolysis and Gluconeogenesis? • Capable of supplying a huge array of metabolites used by many cellular processes,
biofuels, high rate of glycolysis in tumors, lactose intolerance etc.
2Chapter 14 - Glycolysis & Gluconeogenesis
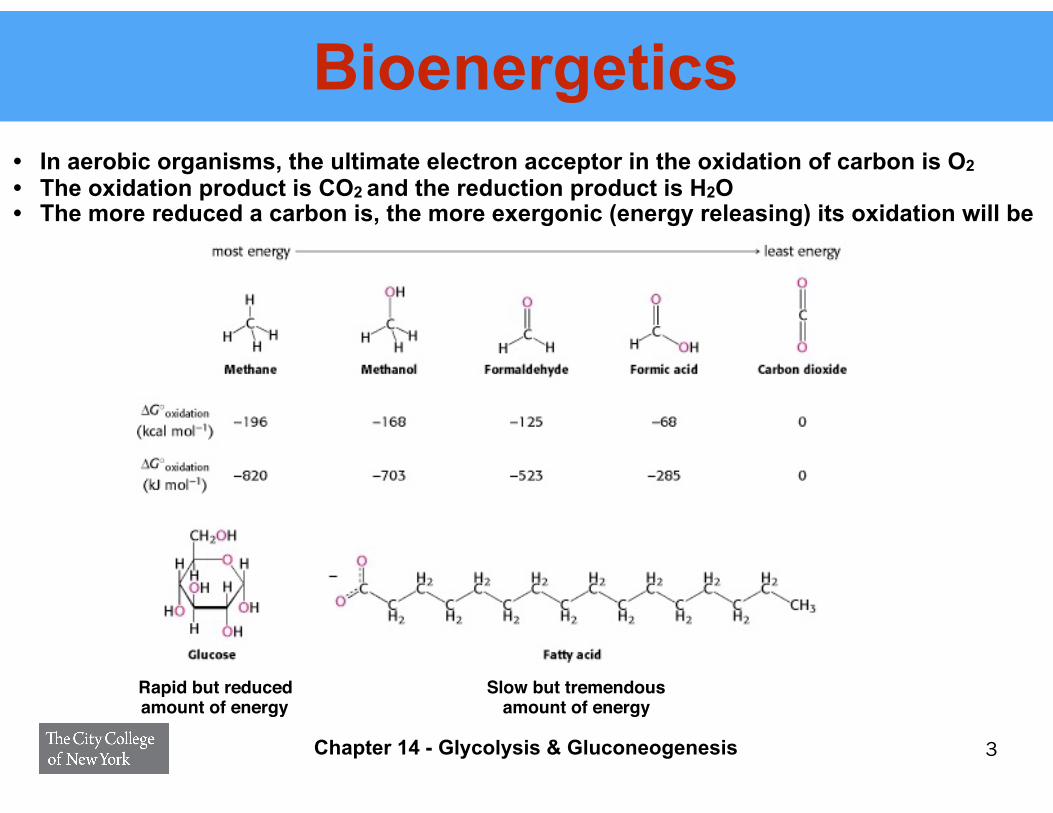
Bioenergetics• In aerobic organisms, the ultimate electron acceptor in the oxidation of carbon is O2 • The oxidation product is CO2 and the reduction product is H2O • The more reduced a carbon is, the more exergonic (energy releasing) its oxidation will be
3
Rapid but reduced amount of energy
Slow but tremendous amount of energy
Chapter 14 - Glycolysis & Gluconeogenesis

Bioenergetics - Two Helpful Pointers
4Chapter 14 - Glycolysis & Gluconeogenesis
Text uses SI units of energy: joules & kilojoules
4.184 J = 1 cal 4.184 kJ = 1 kcal = 1 dietary Cal
note 1 Cal (dietary) = 1000 cal (chemical)

Adenosine Triphosphate (ATP)
5
Reactions written with substrate as ATP, but true substrate is Mg/ATP-2 Mg+2 shields the negative charge of terminal phosphates and allows for nucleophilic attack by enzyme or other substrates Enzymes that bind ATP-4 need Mg+2 present
ATP -> ADP + Pi has ΔG of -30.5 kJ/mol
Chapter 14 - Glycolysis & Gluconeogenesis
Bioenergetics and Biochemical Reaction Types502
phosphoric acid anhydride (phosphoanhydride) bondin ATP separates one of the three negatively chargedphosphates and thus relieves some of the electrostaticrepulsion in ATP; the Pi released is stabilized by the formation of several resonance forms not possible inATP; and ADP2!, the other direct product of hydrolysis,
immediately ionizes, releasing H" into a medium ofvery low [H"] (!10!7 M). Because the concentrationsof the direct products of ATP hydrolysis are, in the cell,far below the concentrations at equilibrium (Table13–5), mass action favors the hydrolysis reaction in the cell.
The free-energy change for ATP hydrolysis is!30.5 kJ/mol under standard conditions, but the ac-tual free energy of hydrolysis (#G) of ATP in livingcells is very different: the cellular concentrations ofATP, ADP, and Pi are not identical and are much lowerthan the 1.0 M of standard conditions (Table 13–5). Fur-thermore, Mg2" in the cytosol binds to ATP and ADP(Fig. 13–12), and for most enzymatic reactions that in-volve ATP as phosphoryl group donor, the true sub-strate is MgATP2!. The relevant #G$% is therefore thatfor MgATP2! hydrolysis. We can calculate #G for ATPhydrolysis using data such as those in Table 13–5. Theactual free energy of hydrolysis of ATP under intracel-lular conditions is often called its phosphorylationpotential, !Gp.
ADP3! " P i2! " H"
#G$% & !30.5 kJ/mol ATP4! " H2O
A
BPO P
!
O
O
B
A!O
O OO
O
O Rib AdenineO
O
OHO
ADP2"
A
B
!O
O
O
O OO Rib Adenine
ADP3"
POPO!O OB
A!
O
O
O
H" "
OPB
AO!
O
O
PO!O OB
A!O
O
OA
OBP
!O
O O
O
O OO Rib Adenine
ATP4"
H
OH
Pi
!
POO OA
O
O
O
POO OB
A!O
O
3!
OH
' !' !
' !
' !
resonancestabilization
AH"
2
ionization3
hydrolysis,with reliefof chargerepulsion
1
FIGURE 13–11 Chemical basis for the large free-energy change as-sociated with ATP hydrolysis. 1 The charge separation that resultsfrom hydrolysis relieves electrostatic repulsion among the four nega-tive charges on ATP. 2 The product inorganic phosphate (Pi) is sta-bilized by formation of a resonance hybrid, in which each of the fourphosphorus–oxygen bonds has the same degree of double-bondcharacter and the hydrogen ion is not permanently associated withany one of the oxygens. (Some degree of resonance stabilization alsooccurs in phosphates involved in ester or anhydride linkages, butfewer resonance forms are possible than for Pi.) 3 The productADP2! immediately ionizes, releasing a proton into a medium ofvery low [H"] (pH 7). A fourth factor (not shown) that favors ATP hy-drolysis is the greater degree of solvation (hydration) of the productsPi and ADP relative to ATP, which further stabilizes the products rel-ative to the reactants.
OP
Mg2"
A!
OPO OB
AO
O
O OO Rib Adenine
MgADP"
OPB
Mg2"
AO!
O
O
PO!O OB
A!O
O
O OBP
!O
O O
O
O OO Rib Adenine
MgATP2"
ø ø
ø ø
O!
O!
O
OB
A
FIGURE 13–12 Mg2" and ATP. Formation of Mg2"
complexes partially shields the negative chargesand influences the conformation of the phosphategroups in nucleotides such as ATP and ADP.
Adenine Nucleotide, Inorganic Phosphate, andPhosphocreatine Concentrations in Some CellsTABLE 13–5
Concentration (mM)*
ATP ADP† AMP Pi PCr
Rat hepatocyte 3.38 1.32 0.29 4.8 0
Rat myocyte 8.05 0.93 0.04 8.05 28
Rat neuron 2.59 0.73 0.06 2.72 4.7
Human erythrocyte 2.25 0.25 0.02 1.65 0
E. coli cell 7.90 1.04 0.82 7.9 0
*For erythrocytes the concentrations are those of the cytosol (human erythrocytes lack a nucleus andmitochondria). In the other types of cells the data are for the entire cell contents, although the cytosol and themitochondria have very different concentrations of ADP. PCr is phosphocreatine, discussed on p. 510.†This value reflects total concentration; the true value for free ADP may be much lower (p. 503).
each of us has about 0.2 mol of ATP at any given time
cannot be stored, usually recycled ~500 cycles per day
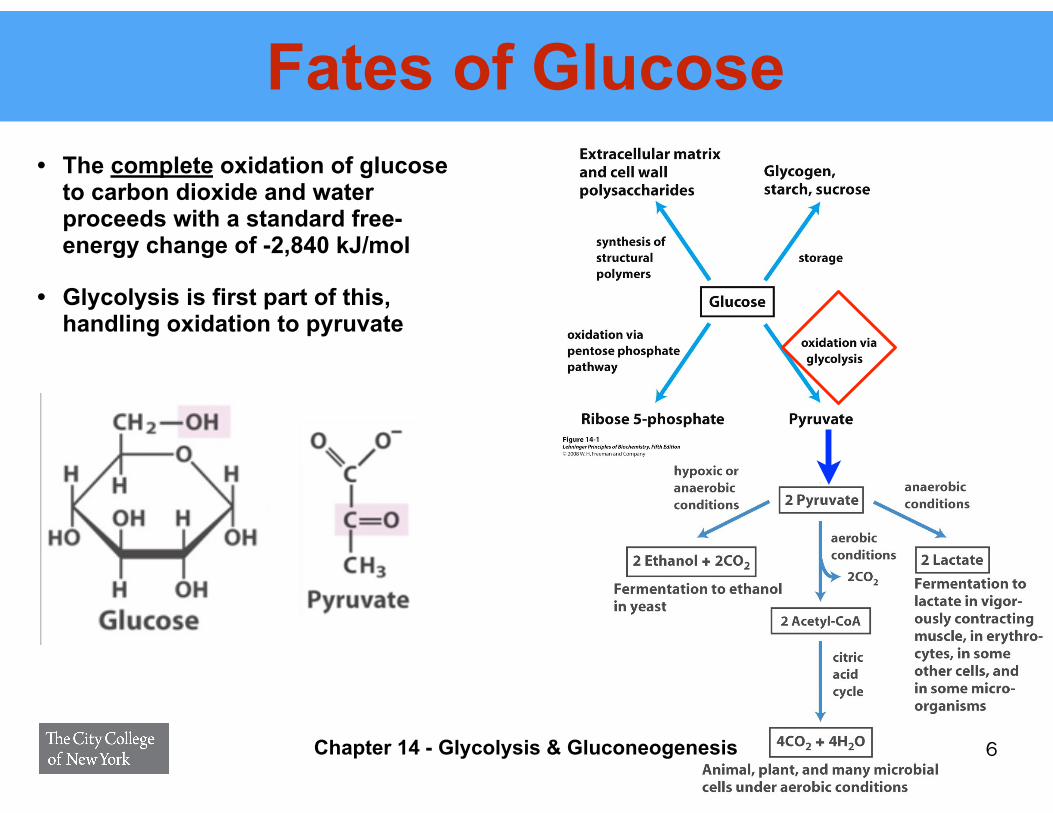
Fates of Glucose• The complete oxidation of glucose
to carbon dioxide and water proceeds with a standard free-energy change of -2,840 kJ/mol
• Glycolysis is first part of this, handling oxidation to pyruvate
6Chapter 14 - Glycolysis & Gluconeogenesis
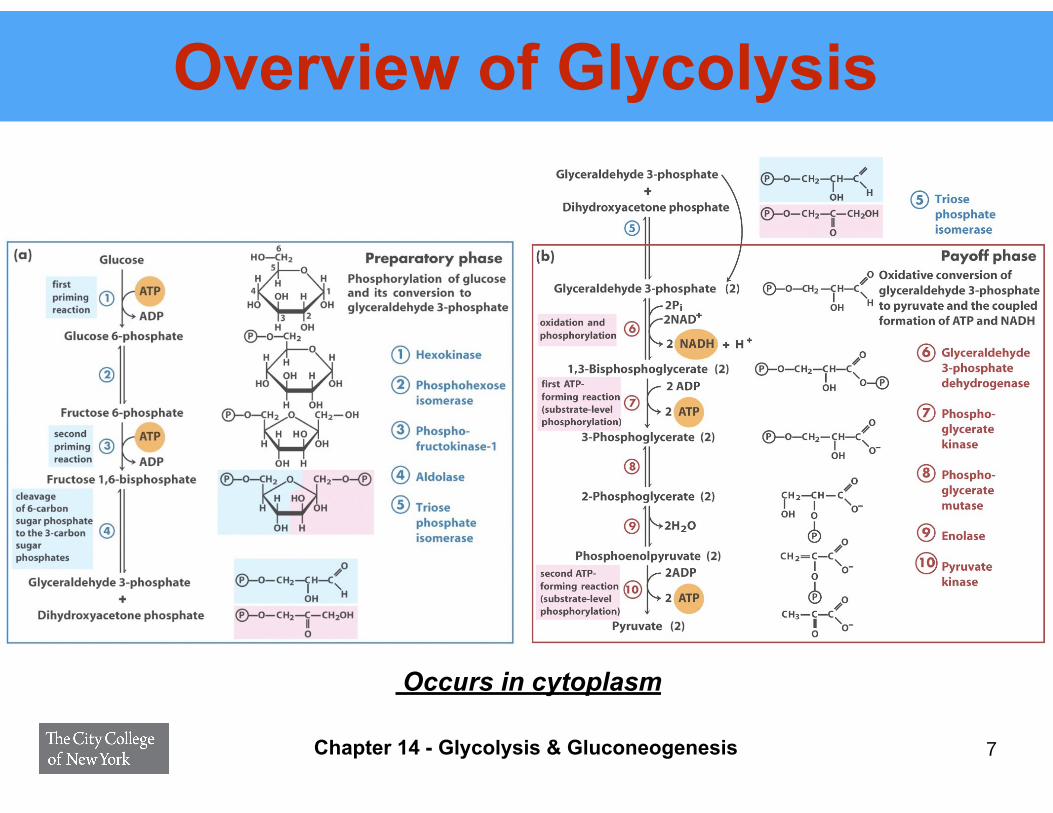
Overview of Glycolysis
7Chapter 14 - Glycolysis & Gluconeogenesis
Occurs in cytoplasm

Terminology
8Chapter 14 - Glycolysis & Gluconeogenesis
Bioenergetics and Biochemical Reaction Types492
KEY CONVENTION: In another simplifying convention usedby biochemists, when H2O, H!, and/or Mg2! are reac-tants or products, their concentrations are not includedin equations such as Equation 13–2 but are insteadincorporated into the constants K"eq and #G"$. ■
Just as K"eq is a physical constant characteristic foreach reaction, so too is #G"$ a constant. As we noted inChapter 6, there is a simple relationship between K"eq
and #G"$:
(13–3)
The standard free-energy change of a chemical reac-tion is simply an alternative mathematical way ofexpressing its equilibrium constant. Table 13–2shows the relationship between #G"$ and K"eq. If theequilibrium constant for a given chemical reaction is 1.0,the standard free-energy change of that reaction is 0.0(the natural logarithm of 1.0 is zero). If K"eq of a reactionis greater than 1.0, its #G"$ is negative. If K"eq is less than1.0, #G"$ is positive. Because the relationship between#G"$ and K"eq is exponential, relatively small changes in#G"$ correspond to large changes in K"eq.
It may be helpful to think of the standard free-energy change in another way. #G"$ is the difference be-tween the free-energy content of the products and thefree-energy content of the reactants, under standardconditions. When #G"$ is negative, the products containless free energy than the reactants and the reaction willproceed spontaneously under standard conditions; allchemical reactions tend to go in the direction that re-sults in a decrease in the free energy of the system. A
¢G¿° % &RT ln K¿eq
positive value of #G"$ means that the products of the re-action contain more free energy than the reactants, andthis reaction will tend to go in the reverse direction if westart with 1.0 M concentrations of all components (stan-dard conditions). Table 13–3 summarizes these points.
Table 13–4 gives the standard free-energy changesfor some representative chemical reactions. Note thathydrolysis of simple esters, amides, peptides, and glyco-sides, as well as rearrangements and eliminations, pro-ceed with relatively small standard free-energy changes,whereas hydrolysis of acid anhydrides is accompaniedby relatively large decreases in standard free energy.The complete oxidation of organic compounds such asglucose or palmitate to CO2 and H2O, which in cells re-quires many steps, results in very large decreases instandard free energy. However, standard free-energy
WORKED EXAMPLE 13–1 Calculation of !G"#
Calculate the standard free-energy change of the reac-tion catalyzed by the enzyme phosphoglucomutase
Glucose 1-phosphate ∆ glucose 6-phosphate
given that, starting with 20 mM glucose 1-phosphate andno glucose 6-phosphate, the final equilibrium mixture at25 $C and pH 7.0 contains 1.0 mM glucose 1-phosphateand 19 mM glucose 6-phosphate. Does the reaction inthe direction of glucose 6-phosphate formation proceedwith a loss or a gain of free energy?
Solution: First we calculate the equilibrium constant:
We can now calculate the standard free-energy change:
Because the standard free-energy change is negative,the conversion of glucose 1-phosphate to glucose 6-phosphate proceeds with a loss (release) of freeenergy. (For the reverse reaction, #G"$ has the samemagnitude but the opposite sign.)
% &7.3 kJ/mol % &(8.315 J/mol $K)(298 K)(ln 19)
¢G¿° % &RT ln K¿eq
K¿eq %[glucose 6-phosphate][glucose 1-phosphate]
%19 mM
1.0 mM% 19
Relationship between EquilibriumConstants and Standard Free-EnergyChanges of Chemical Reactions
TABLE 13–2
#G"#
K"eq (kJ/mol) (kcal/mol)*
103 &17.1 &4.1
102 &11.4 &2.7
101 &5.7 &1.4
1 0.0 0.0
10&1 5.7 1.4
10&2 11.4 2.7
10&3 17.1 4.1
10&4 22.8 5.5
10&5 28.5 6.8
10&6 34.2 8.2
*Although joules and kilojoules are the standard units of energy and are used through-out this text, biochemists and nutritionists sometimes express #G"$ values in kilocalo-ries per mole. We have therefore included values in both kilojoules and kilocalories inthis table and in Tables 13–4 and 13–6. To convert kilojoules to kilocalories, divide thenumber of kilojoules by 4.184.
TABLE 13–3
Starting with allcomponents at 1 M,
When K"eq is . . . #G"# is . . . the reaction . . .
'1.0 negative proceeds forward
1.0 zero is at equilibrium
(1.0 positive proceeds in reverse
Relationships among K"eq, #G"#, and the Direction of Chemical Reactions
13.1 Bioenergetics and Thermodynamics 493
changes such as those in Table 13–4 indicate how muchfree energy is available from a reaction under standardconditions. To describe the energy released under theconditions existing in cells, an expression for the actualfree-energy change is essential.
Actual Free-Energy Changes Depend on Reactant and Product Concentrations
We must be careful to distinguish between two differentquantities: the actual free-energy change, !G, and thestandard free-energy change, !G"#. Each chemical reac-tion has a characteristic standard free-energy change,which may be positive, negative, or zero, depending onthe equilibrium constant of the reaction. The standardfree-energy change tells us in which direction and howfar a given reaction must go to reach equilibrium whenthe initial concentration of each component is 1.0 M,the pH is 7.0, the temperature is 25 #C, and the pressureis 101.3 kPa (1 atm). Thus !G"# is a constant: it has acharacteristic, unchanging value for a given reaction.But the actual free-energy change, !G, is a function ofreactant and product concentrations and of the temper-ature prevailing during the reaction, none of which will
necessarily match the standard conditions as definedabove. Moreover, the !G of any reaction proceedingspontaneously toward its equilibrium is always negative,becomes less negative as the reaction proceeds, and iszero at the point of equilibrium, indicating that no morework can be done by the reaction.
!G and !G"# for any reaction aA $ bB ∆ cC $ dDare related by the equation
(13–4)
in which the terms in red are those actually prevailingin the system under observation. The concentrationterms in this equation express the effects commonlycalled mass action, and the term [C]c[D]d/[A]a[B]b is calledthe mass-action ratio, Q. Thus Equation 13–4 can beexpressed as !G % !G"# $ RT ln Q. As an example, let ussuppose that the reaction A $ B ∆ C $ D is taking placeunder the standard conditions of temperature (25 #C) andpressure (101.3 kPa) but that the concentrations of A, B, C, and D are not equal and none of the compo-nents is present at the standard concentration of 1.0 M. To determine the actual free-energy change, !G, underthese nonstandard conditions of concentration as the
¢G % ¢G¿° $ RT ln [C]c[D]d
[A]a[B]b
Standard Free-Energy Changes of Some Chemical ReactionsTABLE 13–4
!G!"
Reaction type (kJ/mol) (kcal/mol)
Hydrolysis reactions
Acid anhydrides
Acetic anhydride $ H2O 88n 2 acetate &91.1 &21.8ATP $ H2O 88n ADP $ Pi &30.5 &7.3ATP $ H2O 88n AMP $ PPi &45.6 &10.9PPi $ H2O 88n 2Pi &19.2 &4.6UDP-glucose $ H2O 88n UMP $ glucose 1-phosphate &43.0 &10.3
Esters
Ethyl acetate $ H2O 88n ethanol $ acetate &19.6 &4.7Glucose 6-phosphate $ H2O 88n glucose $ Pi &13.8 &3.3
Amides and peptides
Glutamine $ H2O 88n glutamate $ NH$4 &14.2 &3.4
Glycylglycine $ H2O 88n 2 glycine &9.2 &2.2
Glycosides
Maltose $ H2O 88n 2 glucose &15.5 &3.7Lactose $ H2O 88n glucose $ galactose &15.9 &3.8
Rearrangements
Glucose 1-phosphate 88n glucose 6-phosphate &7.3 &1.7Fructose 6-phosphate 88n glucose 6-phosphate &1.7 &0.4
Elimination of water
Malate 88n fumarate $ H2O 3.1 0.8
Oxidations with molecular oxygen
Glucose $ 6O2 88n 6CO2 $ 6H2O &2,840 &686Palmitate $ 23O2 88n 16CO2 $ 16H2O &9,770 &2,338
13.1 Bioenergetics and Thermodynamics 493
changes such as those in Table 13–4 indicate how muchfree energy is available from a reaction under standardconditions. To describe the energy released under theconditions existing in cells, an expression for the actualfree-energy change is essential.
Actual Free-Energy Changes Depend on Reactant and Product Concentrations
We must be careful to distinguish between two differentquantities: the actual free-energy change, !G, and thestandard free-energy change, !G"#. Each chemical reac-tion has a characteristic standard free-energy change,which may be positive, negative, or zero, depending onthe equilibrium constant of the reaction. The standardfree-energy change tells us in which direction and howfar a given reaction must go to reach equilibrium whenthe initial concentration of each component is 1.0 M,the pH is 7.0, the temperature is 25 #C, and the pressureis 101.3 kPa (1 atm). Thus !G"# is a constant: it has acharacteristic, unchanging value for a given reaction.But the actual free-energy change, !G, is a function ofreactant and product concentrations and of the temper-ature prevailing during the reaction, none of which will
necessarily match the standard conditions as definedabove. Moreover, the !G of any reaction proceedingspontaneously toward its equilibrium is always negative,becomes less negative as the reaction proceeds, and iszero at the point of equilibrium, indicating that no morework can be done by the reaction.
!G and !G"# for any reaction aA $ bB ∆ cC $ dDare related by the equation
(13–4)
in which the terms in red are those actually prevailingin the system under observation. The concentrationterms in this equation express the effects commonlycalled mass action, and the term [C]c[D]d/[A]a[B]b is calledthe mass-action ratio, Q. Thus Equation 13–4 can beexpressed as !G % !G"# $ RT ln Q. As an example, let ussuppose that the reaction A $ B ∆ C $ D is taking placeunder the standard conditions of temperature (25 #C) andpressure (101.3 kPa) but that the concentrations of A, B, C, and D are not equal and none of the compo-nents is present at the standard concentration of 1.0 M. To determine the actual free-energy change, !G, underthese nonstandard conditions of concentration as the
¢G % ¢G¿° $ RT ln [C]c[D]d
[A]a[B]b
Standard Free-Energy Changes of Some Chemical ReactionsTABLE 13–4
!G!"
Reaction type (kJ/mol) (kcal/mol)
Hydrolysis reactions
Acid anhydrides
Acetic anhydride $ H2O 88n 2 acetate &91.1 &21.8ATP $ H2O 88n ADP $ Pi &30.5 &7.3ATP $ H2O 88n AMP $ PPi &45.6 &10.9PPi $ H2O 88n 2Pi &19.2 &4.6UDP-glucose $ H2O 88n UMP $ glucose 1-phosphate &43.0 &10.3
Esters
Ethyl acetate $ H2O 88n ethanol $ acetate &19.6 &4.7Glucose 6-phosphate $ H2O 88n glucose $ Pi &13.8 &3.3
Amides and peptides
Glutamine $ H2O 88n glutamate $ NH$4 &14.2 &3.4
Glycylglycine $ H2O 88n 2 glycine &9.2 &2.2
Glycosides
Maltose $ H2O 88n 2 glucose &15.5 &3.7Lactose $ H2O 88n glucose $ galactose &15.9 &3.8
Rearrangements
Glucose 1-phosphate 88n glucose 6-phosphate &7.3 &1.7Fructose 6-phosphate 88n glucose 6-phosphate &1.7 &0.4
Elimination of water
Malate 88n fumarate $ H2O 3.1 0.8
Oxidations with molecular oxygen
Glucose $ 6O2 88n 6CO2 $ 6H2O &2,840 &686Palmitate $ 23O2 88n 16CO2 $ 16H2O &9,770 &2,338
• Note that you’ll see plenty of ΔG’° (“delta G prime naught” = “standard free energy change”) values on next slides; remember that those are determined at standard conditions (pH 7.0, 1 mM MgCl2, 1M other reactantsThese values are obviously not at equilibrium! As such, ΔG’° values tell you which direction a reaction will spontaneously proceed in, and how far this is from equilibrium, when at standard conditions!
• ΔG values (“actual free energy change”) at any specific condition requires knowledge of actual reactant & product concentrations:
Mass action ratioRead section 13.1
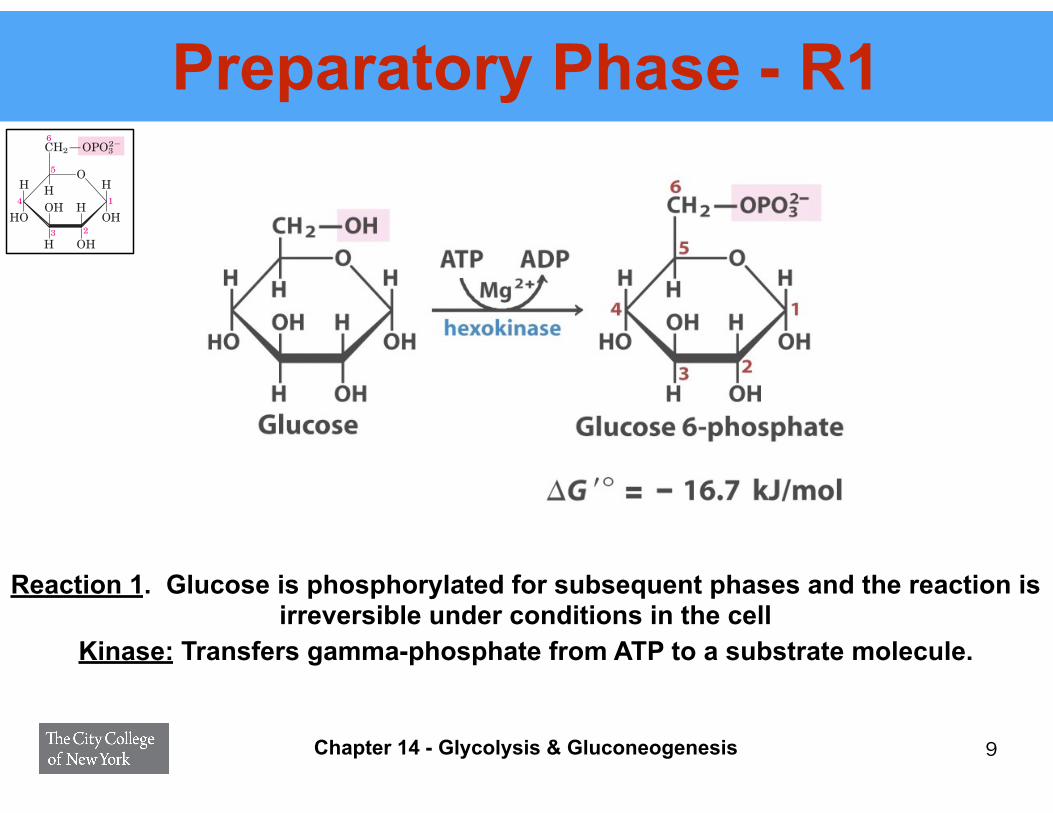
Preparatory Phase - R1
Reaction 1. Glucose is phosphorylated for subsequent phases and the reaction is irreversible under conditions in the cell
9
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Kinase: Transfers gamma-phosphate from ATP to a substrate molecule.
Chapter 14 - Glycolysis & Gluconeogenesis
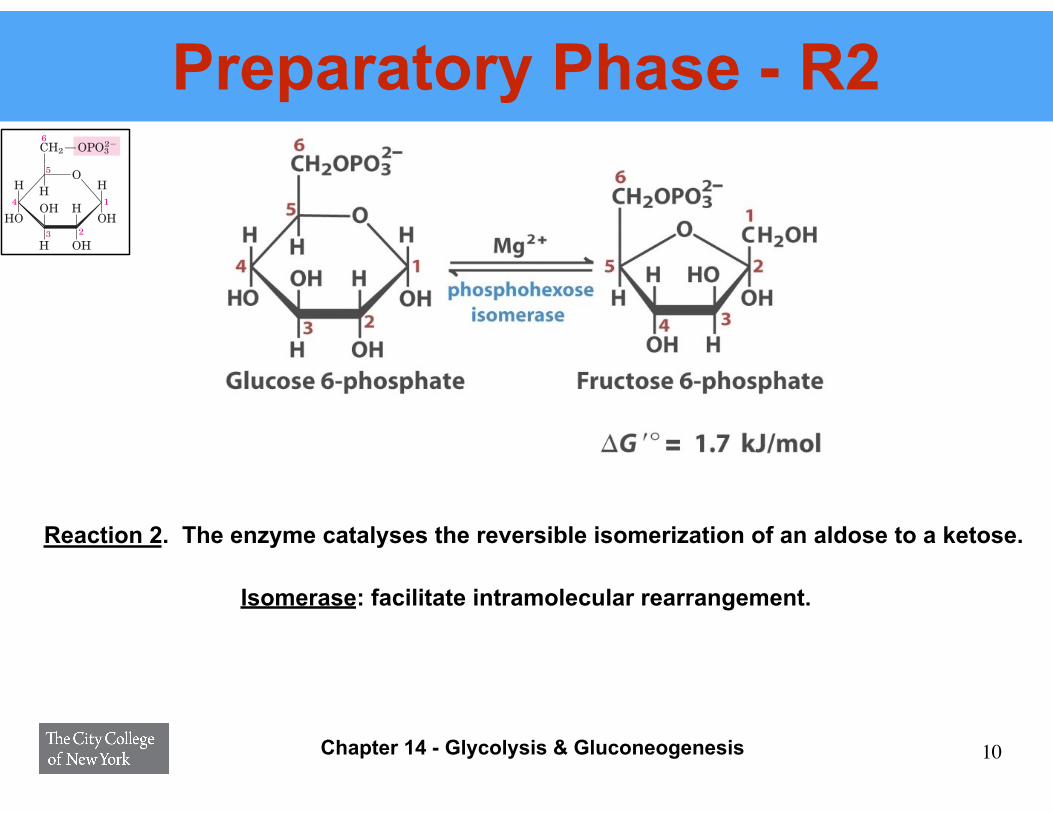
Preparatory Phase - R2
Reaction 2. The enzyme catalyses the reversible isomerization of an aldose to a ketose.
10
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Isomerase: facilitate intramolecular rearrangement.
Chapter 14 - Glycolysis & Gluconeogenesis
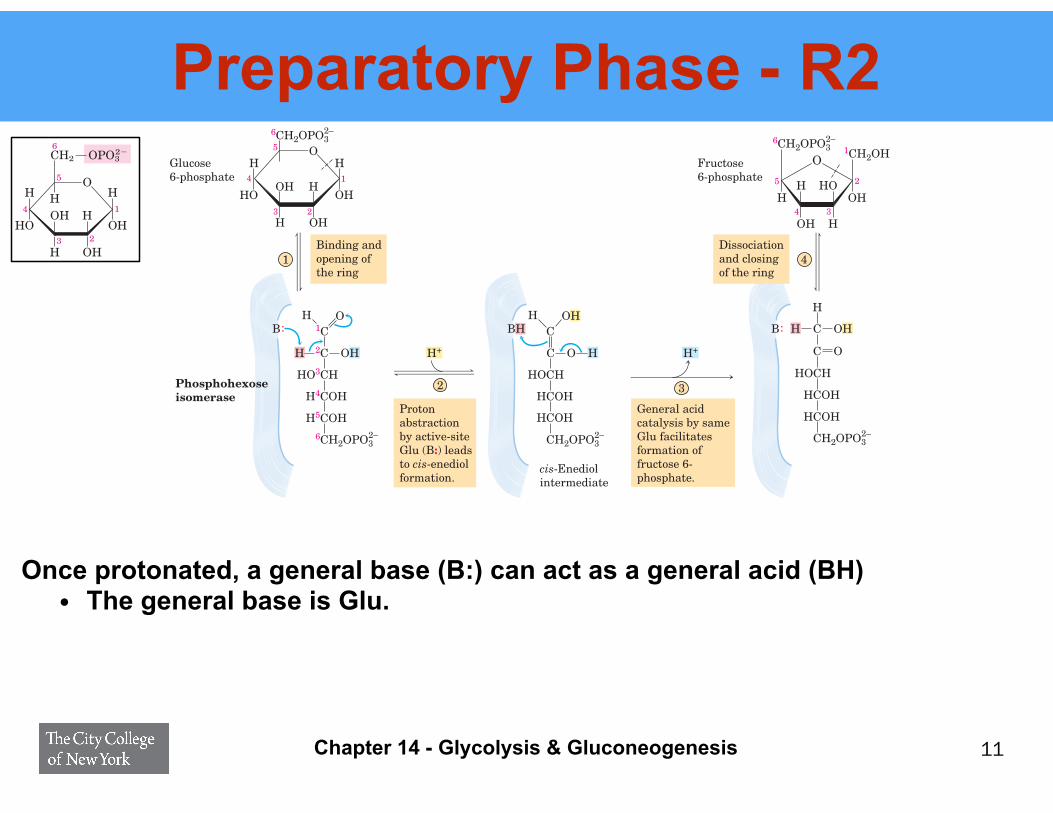
Preparatory Phase - R2
11
14.1 Glycolysis 533
as inositol 1,4,5-trisphosphate; see p. 432) and triphos-phates (such as adenosine triphosphate, ATP). ■
The enzyme that forms fructose 1,6-bisphosphateis called PFK-1 to distinguish it from a second enzyme(PFK-2) that catalyzes the formation of fructose 2,6-bisphosphate from fructose 6-phosphate in a sepa-rate pathway (the roles of PFK-2 and fructose 2,6-bisphosphate are discussed in Chapter 15). The PFK-1reaction is essentially irreversible under cellular con-ditions, and it is the first “committed” step in theglycolytic pathway; glucose 6-phosphate and fructose6-phosphate have other possible fates, but fructose1,6-bisphosphate is targeted for glycolysis.
Some bacteria and protists and perhaps all plantshave a phosphofructokinase that uses pyrophosphate(PPi), not ATP, as the phosphoryl group donor in thesynthesis of fructose 1,6-bisphosphate:
Mg2!
Fructose 6-phosphate ! PPi 88nfructose 1,6-bisphosphate ! Pi
"G#$ % &2.9 kJ/mol
Phosphofructokinase-1 is subject to complex al-losteric regulation; its activity is increased whenever thecell’s ATP supply is depleted or when the ATP breakdownproducts, ADP and AMP (particularly the latter), accu-mulate. The enzyme is inhibited whenever the cell hasample ATP and is well supplied by other fuels such asfatty acids. In some organisms, fructose 2,6-bisphosphate
(not to be confused with the PFK-1 reaction product,fructose 1,6-bisphosphate) is a potent allosteric activatorof PFK-1. Ribulose 5-phosphate, an intermediate in thepentose phosphate pathway discussed later in this chap-ter, also activates phosphofructokinase indirectly. Themultiple layers of regulation of this step in glycolysis arediscussed in greater detail in Chapter 15.
4 Cleavage of Fructose 1,6-Bisphosphate The en-zyme fructose 1,6-bisphosphate aldolase, often calledsimply aldolase, catalyzes a reversible aldol condensa-tion (see Fig. 13–4). Fructose 1,6-bisphosphate is cleavedto yield two different triose phosphates, glyceraldehyde3-phosphate, an aldose, and dihydroxyacetone phos-phate, a ketose:
Phosphohexoseisomerase
:OH
B
HO3CH
2C
1C
OH H+H
H4COH
H5COH6CH2OPO3
2–
:B
HOCH
C
C O
H
OHH
HCOH
HCOH
cis-Enediolintermediate
OHHBH
HOCH
C
C
O H H+
HCOH
HCOH
CH2OPO32– CH2OPO3
2–
1 4
2 3
HOHHO
H H
6CH2OPO32–
6CH2OPO32–
H OH
OH
Glucose6-phosphate
O
HOHH
1CH2OH
OH H
OH
Fructose6-phosphate1
2
2
3 34
4
5
5
O
Binding and opening of the ring
Proton abstractionby active-site Glu (B:) leads to cis-enediol formation.
General acid catalysis by same Glu facilitates formation of fructose 6-phosphate.
Dissociation and closing of the ring
MECHANISM FIGURE 14–4 The phosphohexose isomerase reaction.The ring opening and closing reactions (steps 1 and 4 ) are catalyzedby an active-site His residue, by mechanisms omitted here for simplic-ity. The proton (pink) initially at C-2 is made more easily abstractableby electron withdrawal by the adjacent carbonyl and nearby hydroxyl
group. After its transfer from C-2 to the active-site Glu residue (a weak acid), the proton is freely exchanged with the surroundingsolution; that is, the proton abstracted from C-2 in step 2 is notnecessarily the same one that is added to C-1 in step 3 .
Phosphohexose Isomerase Mechanism
aldolase
O
HH
OH H
OH
Fructose 1,6-bisphosphate
HO
1
25
4 3
6CH2OPO3
2& CH2OPO32&
CHOH
Glyceraldehyde3-phosphate
% 23.8 kJ/molDG#$
!
C
O
H
OH
C
CH2
O
Dihydroxyacetonephosphate
e
(1)
(2) (5)
(4)
(3) (6)
CH2OPO32&
CH2OPO32&
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Once protonated, a general base (B:) can act as a general acid (BH) • The general base is Glu.
Chapter 14 - Glycolysis & Gluconeogenesis
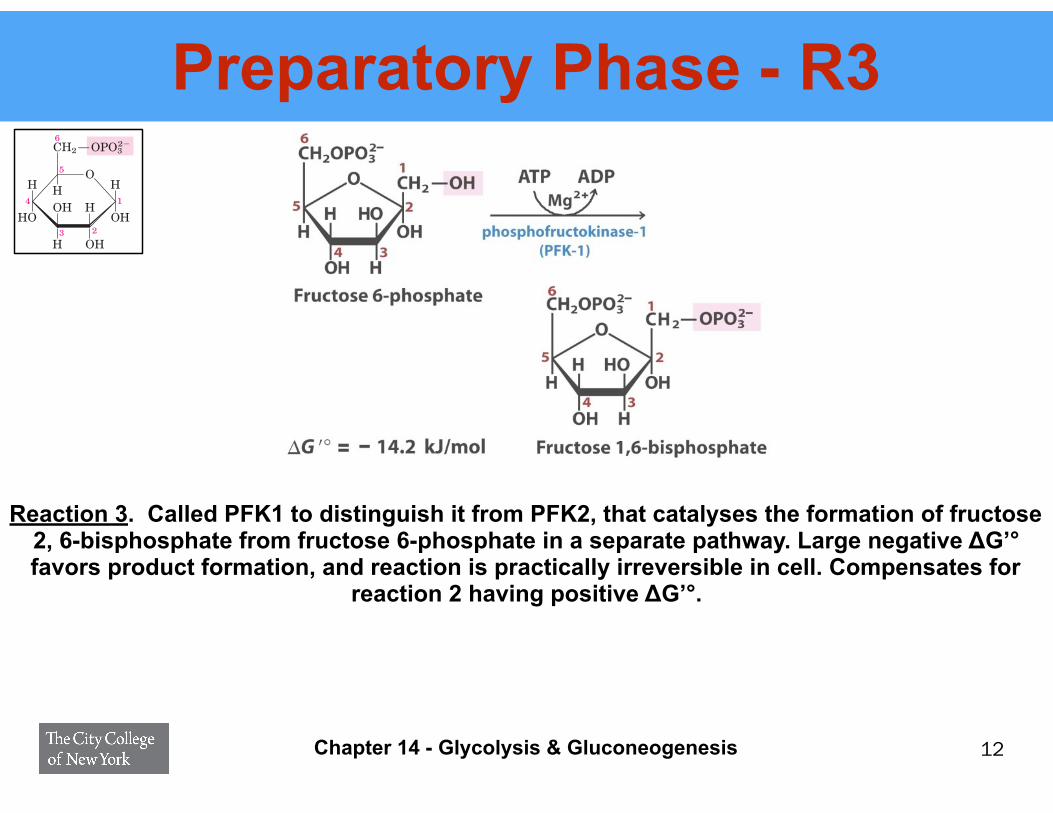
Preparatory Phase - R3
12
Reaction 3. Called PFK1 to distinguish it from PFK2, that catalyses the formation of fructose 2, 6-bisphosphate from fructose 6-phosphate in a separate pathway. Large negative ΔG’° favors product formation, and reaction is practically irreversible in cell. Compensates for
reaction 2 having positive ΔG’°.
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Chapter 14 - Glycolysis & Gluconeogenesis
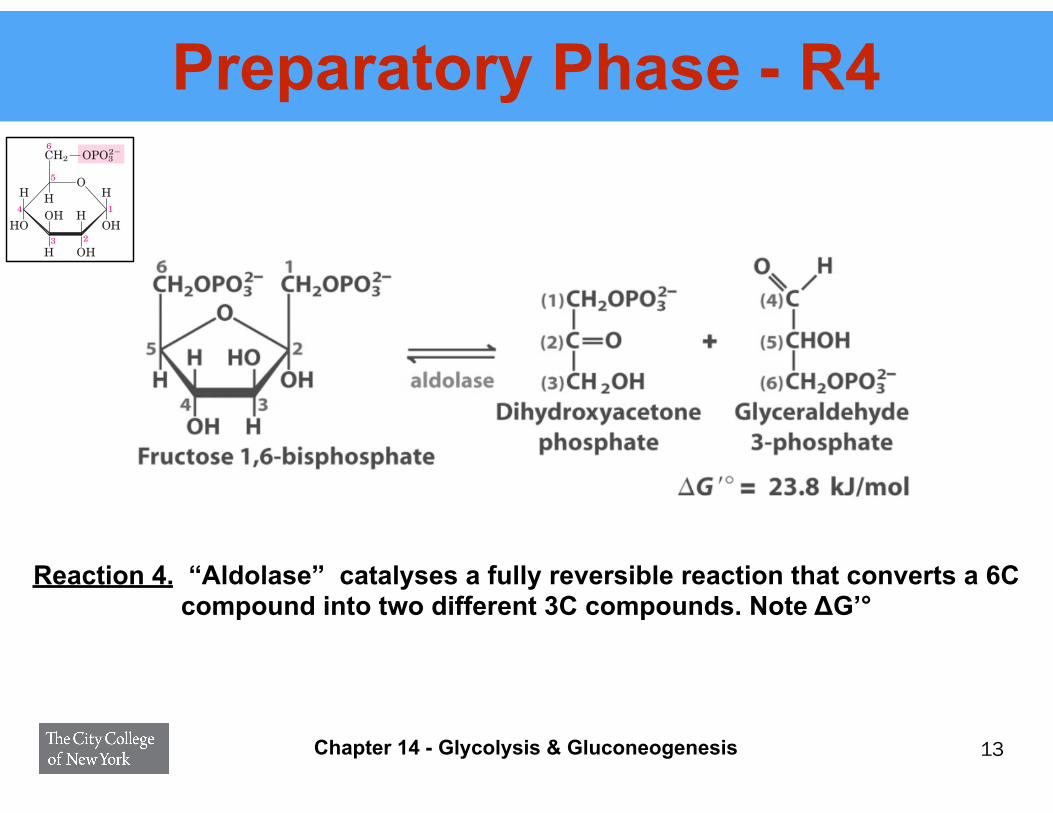
Preparatory Phase - R4
13
Reaction 4. “Aldolase” catalyses a fully reversible reaction that converts a 6C compound into two different 3C compounds. Note ΔG’°
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Chapter 14 - Glycolysis & Gluconeogenesis
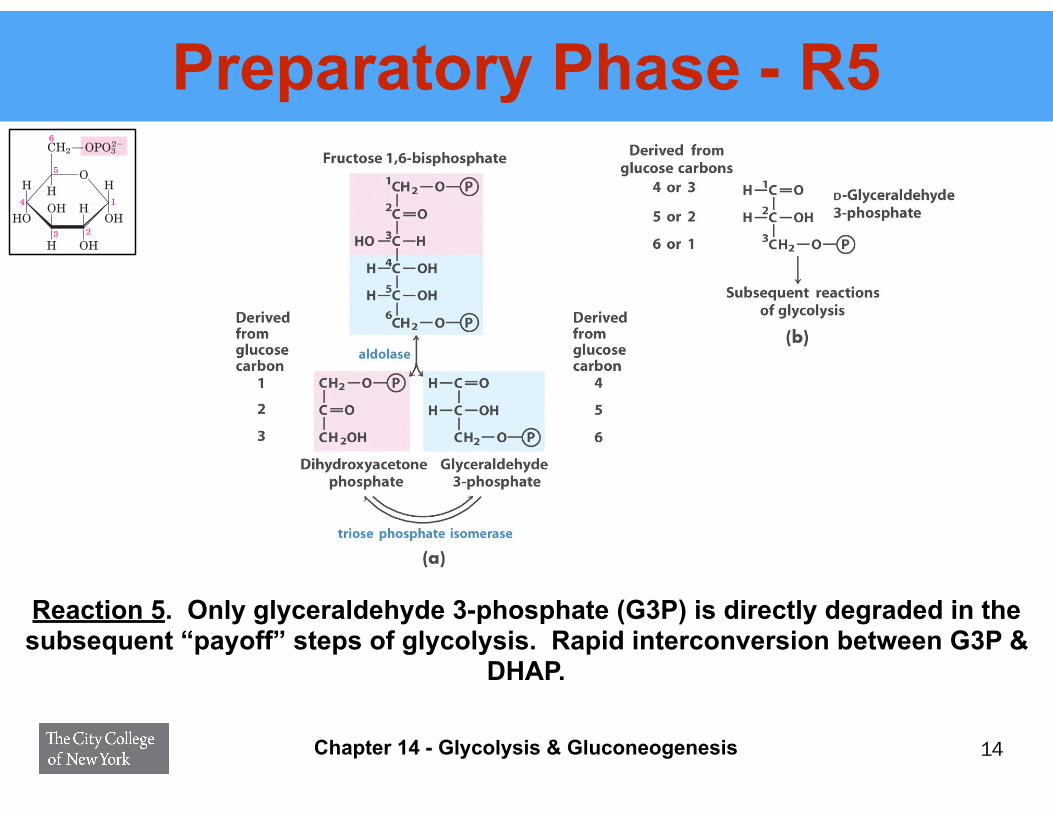
Preparatory Phase - R5
Reaction 5. Only glyceraldehyde 3-phosphate (G3P) is directly degraded in the subsequent “payoff” steps of glycolysis. Rapid interconversion between G3P &
DHAP.
14
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Chapter 14 - Glycolysis & Gluconeogenesis
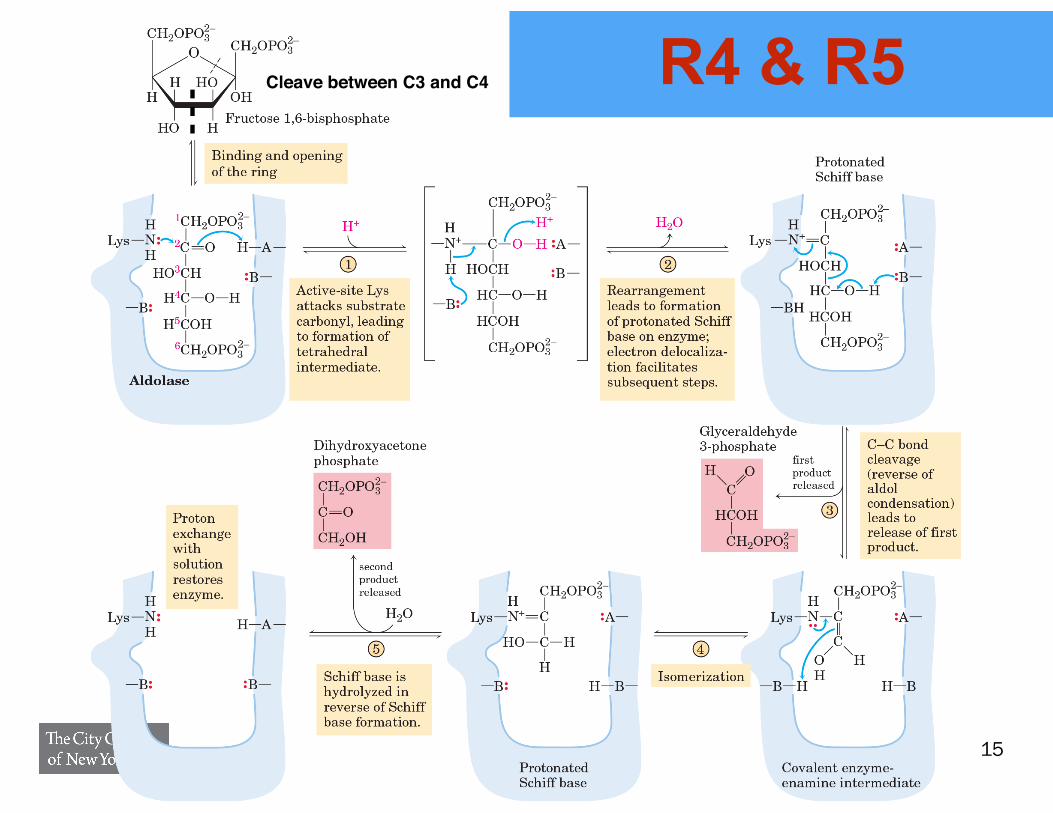
R4 & R5
15
Cleave between C3 and C4
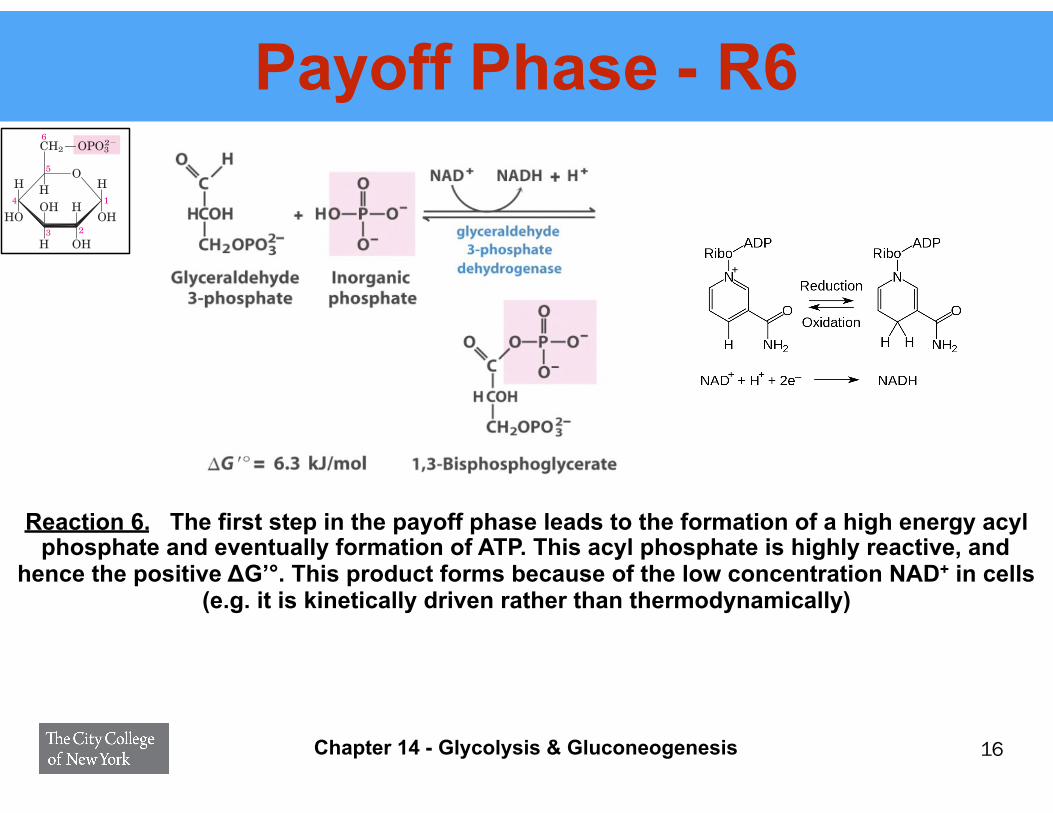
Payoff Phase - R6
Reaction 6. The first step in the payoff phase leads to the formation of a high energy acyl phosphate and eventually formation of ATP. This acyl phosphate is highly reactive, and
hence the positive ΔG’°. This product forms because of the low concentration NAD+ in cells (e.g. it is kinetically driven rather than thermodynamically)
16
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Chapter 14 - Glycolysis & Gluconeogenesis
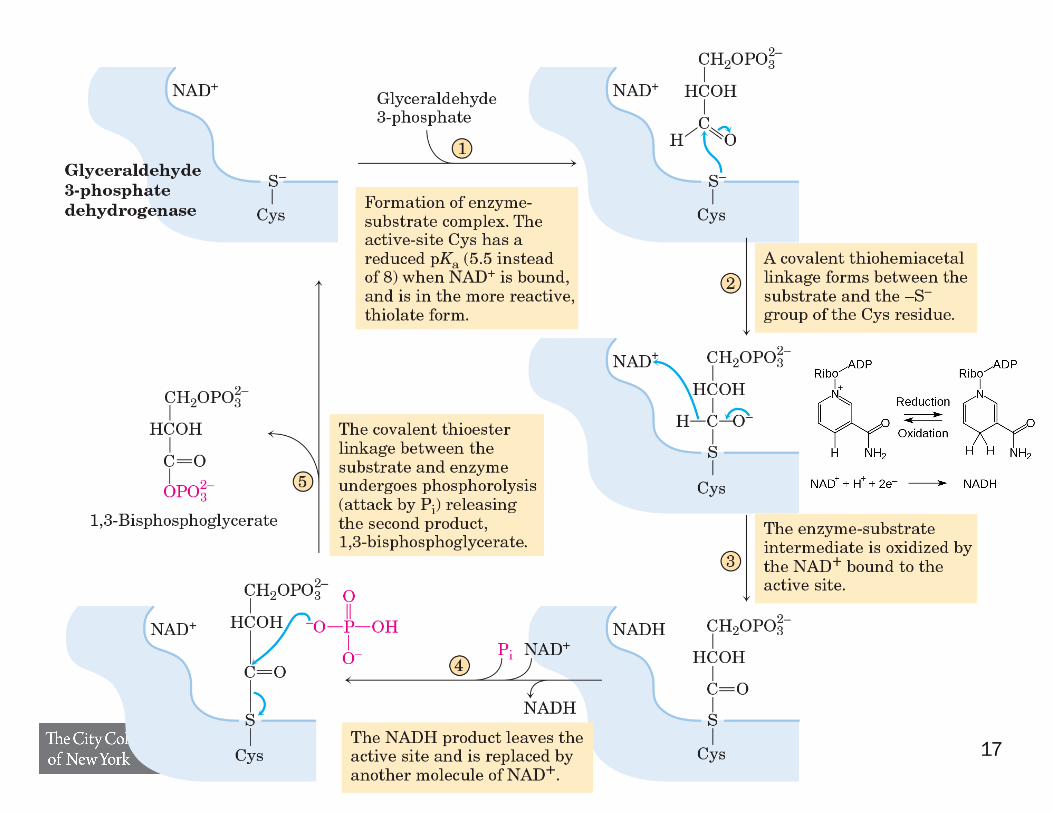
17
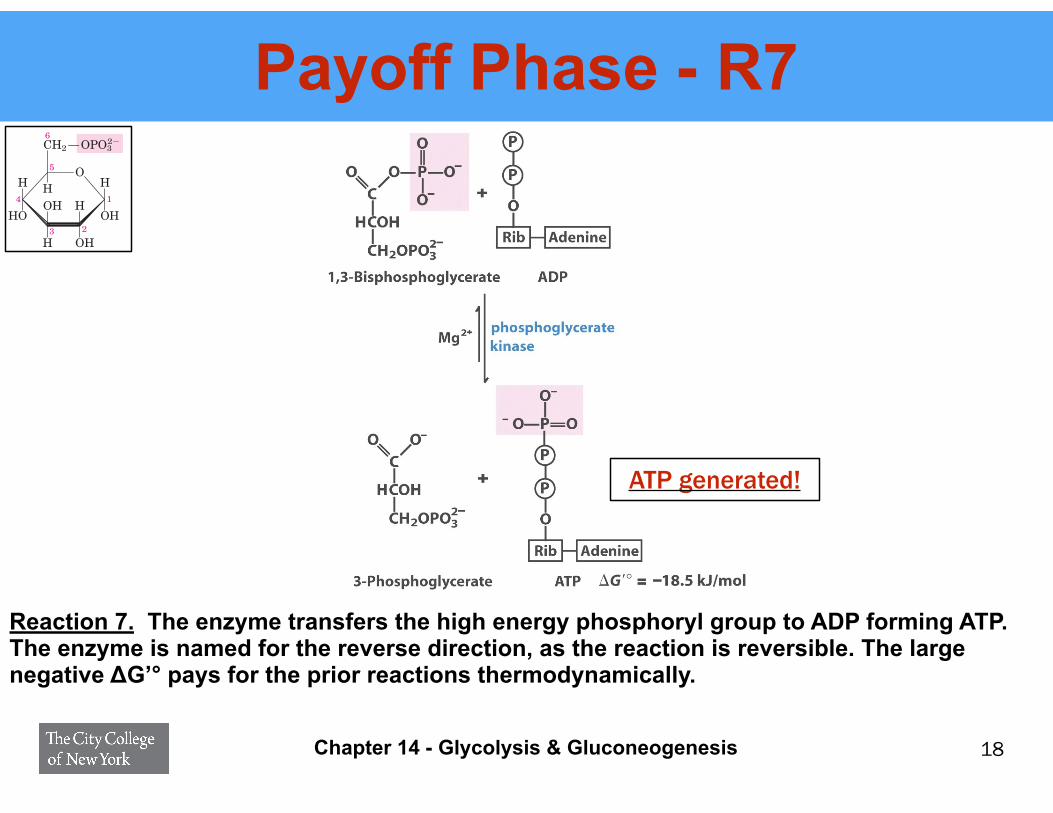
Payoff Phase - R7
Reaction 7. The enzyme transfers the high energy phosphoryl group to ADP forming ATP. The enzyme is named for the reverse direction, as the reaction is reversible. The large negative ΔG’° pays for the prior reactions thermodynamically.
18
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Chapter 14 - Glycolysis & Gluconeogenesis
ATP generated!
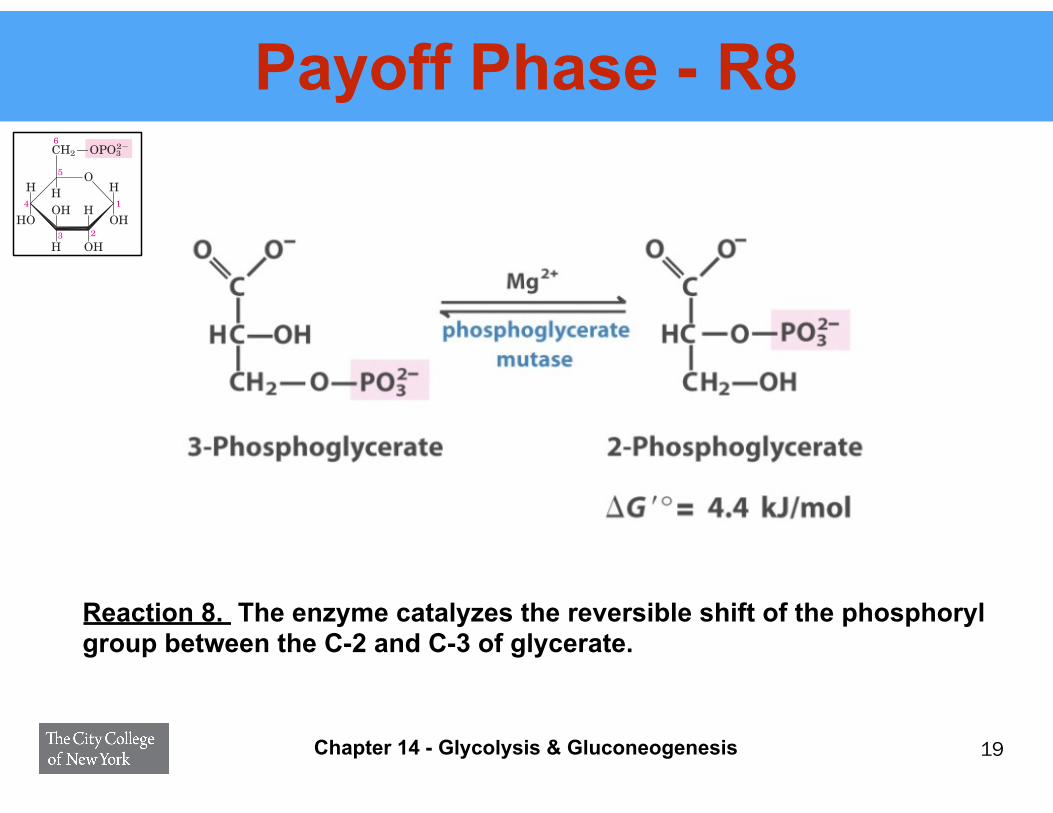
Payoff Phase - R8
Reaction 8. The enzyme catalyzes the reversible shift of the phosphoryl group between the C-2 and C-3 of glycerate.
19
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Chapter 14 - Glycolysis & Gluconeogenesis
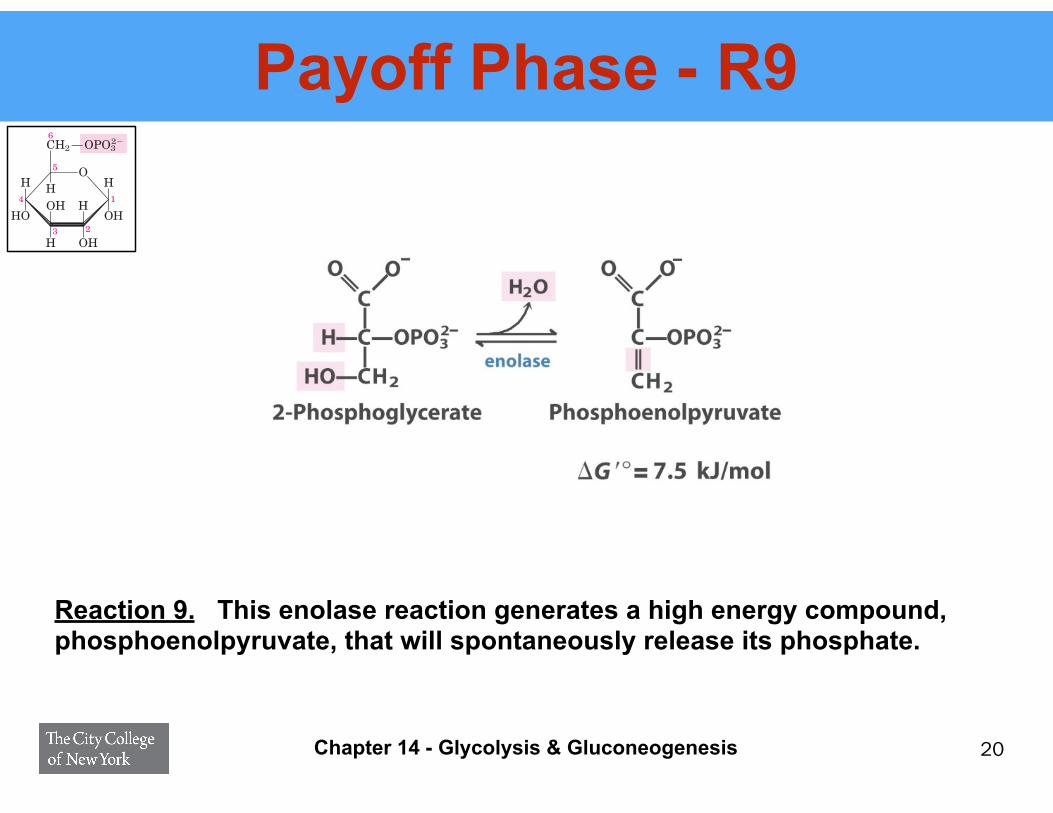
Payoff Phase - R9
Reaction 9. This enolase reaction generates a high energy compound, phosphoenolpyruvate, that will spontaneously release its phosphate.
20
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"
Chapter 14 - Glycolysis & Gluconeogenesis
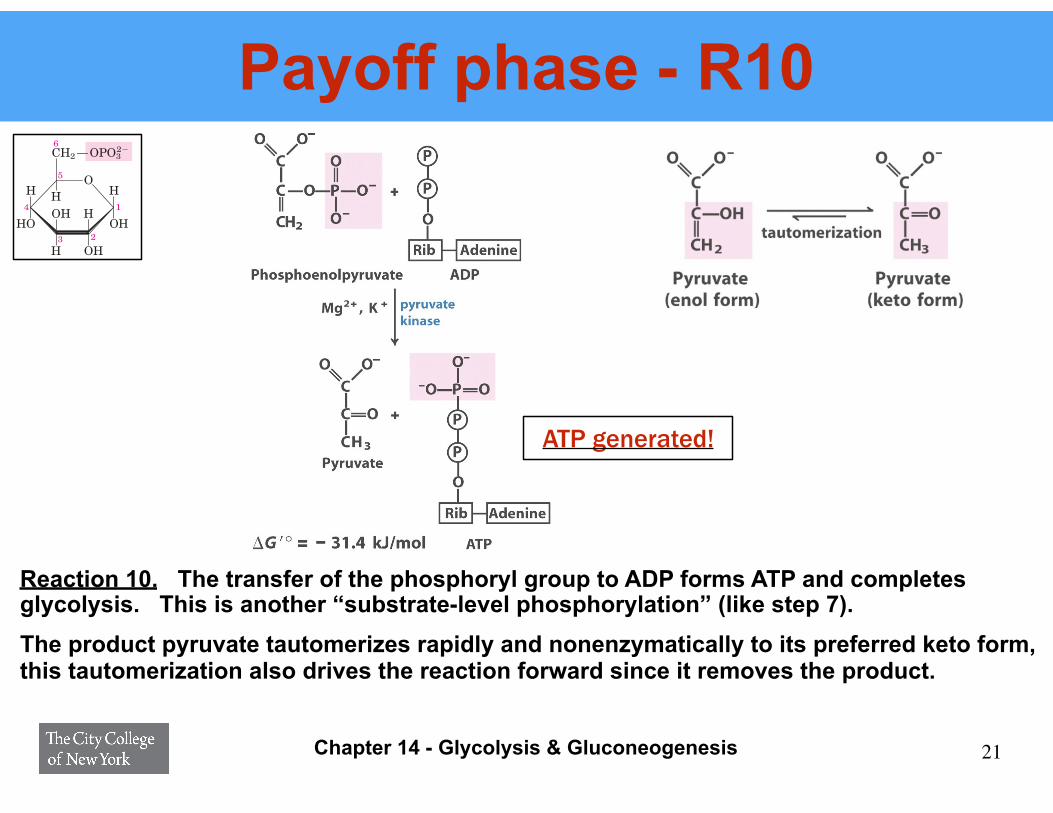
Payoff phase - R10
Reaction 10. The transfer of the phosphoryl group to ADP forms ATP and completes glycolysis. This is another “substrate-level phosphorylation” (like step 7). The product pyruvate tautomerizes rapidly and nonenzymatically to its preferred keto form, this tautomerization also drives the reaction forward since it removes the product.
21
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway532
1 Phosphorylation of Glucose In the first step ofglycolysis, glucose is activated for subsequent reactionsby its phosphorylation at C-6 to yield glucose 6-phos-phate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellularconditions, is catalyzed by hexokinase. Recall that ki-nases are enzymes that catalyze the transfer of the ter-minal phosphoryl group from ATP to an acceptornucleophile (see Fig. 13–20). Kinases are a subclass oftransferases (see Table 6–3). The acceptor in the case ofhexokinase is a hexose, normally D-glucose, althoughhexokinase also catalyzes the phosphorylation of othercommon hexoses, such as D-fructose and D-mannose, insome tissues.
Hexokinase, like many other kinases, requiresMg2! for its activity, because the true substrate of theenzyme is not ATP4" but the MgATP2" complex (seeFig. 13–12). Mg2! shields the negative charges of thephosphoryl groups in ATP, making the terminal phos-phorus atom an easier target for nucleophilic attack byan —OH of glucose. Hexokinase undergoes a profoundchange in shape, an induced fit, when it binds glucose;two domains of the protein move about 8 Å closer toeach other when ATP binds (see Fig. 6–22). Thismovement brings bound ATP closer to a molecule ofglucose also bound to the enzyme and blocks the ac-cess of water (from the solvent), which might other-wise enter the active site and attack (hydrolyze) thephosphoanhydride bonds of ATP. Like the other nineenzymes of glycolysis, hexokinase is a soluble, cytoso-lic protein.
Hexokinase is present in nearly all organisms.The human genome encodes four different hexoki-nases (I to IV), all of which catalyze the same reac-tion. Two or more enzymes that catalyze the samereaction but are encoded by different genes arecalled isozymes (see Box 15–2). One of the isozymespresent in hepatocytes, hexokinase IV (also calledglucokinase), differs from other forms of hexokinasein kinetic and regulatory properties, with importantphysiological consequences that are described inSection 15.3.
2 Conversion of Glucose 6-Phosphate to Fructose6-Phosphate The enzyme phosphohexose isomerase(phosphoglucose isomerase) catalyzes the reversible
The mechanism for this reaction involves an enediol in-termediate (Fig. 14–4). The reaction proceeds readilyin either direction, as might be expected from the rela-tively small change in standard free energy. This isomer-ization has a critical role in the overall chemistry of theglycolytic pathway, as the rearrangement of the car-bonyl and hydroxyl groups at C-1 and C-2 is a necessaryprelude to the next two steps. The phosphorylation thatoccurs in the next reaction (step 3 ) requires that thegroup at C-1 first be converted from a carbonyl to an al-cohol, and in the subsequent reaction (step 4 ) cleav-age of the bond between C-3 and C-4 requires a carbonylgroup at C-2 (p. 497).
3 Phosphorylation of Fructose 6-Phosphate toFructose 1,6-Bisphosphate In the second of the twopriming reactions of glycolysis, phosphofructokinase-1(PFK-1) catalyzes the transfer of a phosphoryl groupfrom ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
KEY CONVENTION: Compounds that contain two phos-phate or phosphoryl groups attached at different po-sitions in the molecule are named bisphosphates(or bisphospho compounds); for example, fructose1,6-bisphosphate and 1,3-bisphosphoglycerate. Com-pounds with two phosphates linked together as a py-rophosphoryl group are named diphosphates; forexample, adenosine diphosphate (ADP). Similarrules apply for the naming of trisphosphates (such
O
OPO32"
H
OHHO
H
H H
OHH
CH2
OH
OO
H
OHHO
H
H H
OHH
CH2
OH
OOH
ATP ADP
Glucose Glucose 6-phosphate
# "16.7 kJ/molDG$%
hexokinase
Mg2!
5
6
4 1
23
O
HO
OH
HH
OH H
CH2
OH
H
OHHO
H
H H
OHH
OH
Glucose 6-phosphate Fructose 6-phosphate
# 1.7 kJ/molDG$%
Mg2!
phosphohexoseisomerase
4
2
1
3
6
5
4 3
2
1O
6
5
CH2OPO32"
CH2OPO32"
isomerization of glucose 6-phosphate, an aldose, to fruc-tose 6-phosphate, a ketose:
ATP ADP
phosphofructokinase-1(PFK-1)
Mg2!
O
HH
OH H
CH2
OH
OH
Fructose 6-phosphate
HO
6
5
4 3
61
1
2
CH2OPO32"
CH OPO2"OPO3
2"
HO
Fructose 1,6-bisphosphate
# "14.2 kJ/molDG$%
O
HH
OH H
CH2
OH
61
5
4 3
2
CH2OPO32"Chapter 14 - Glycolysis & Gluconeogenesis
ATP generated!
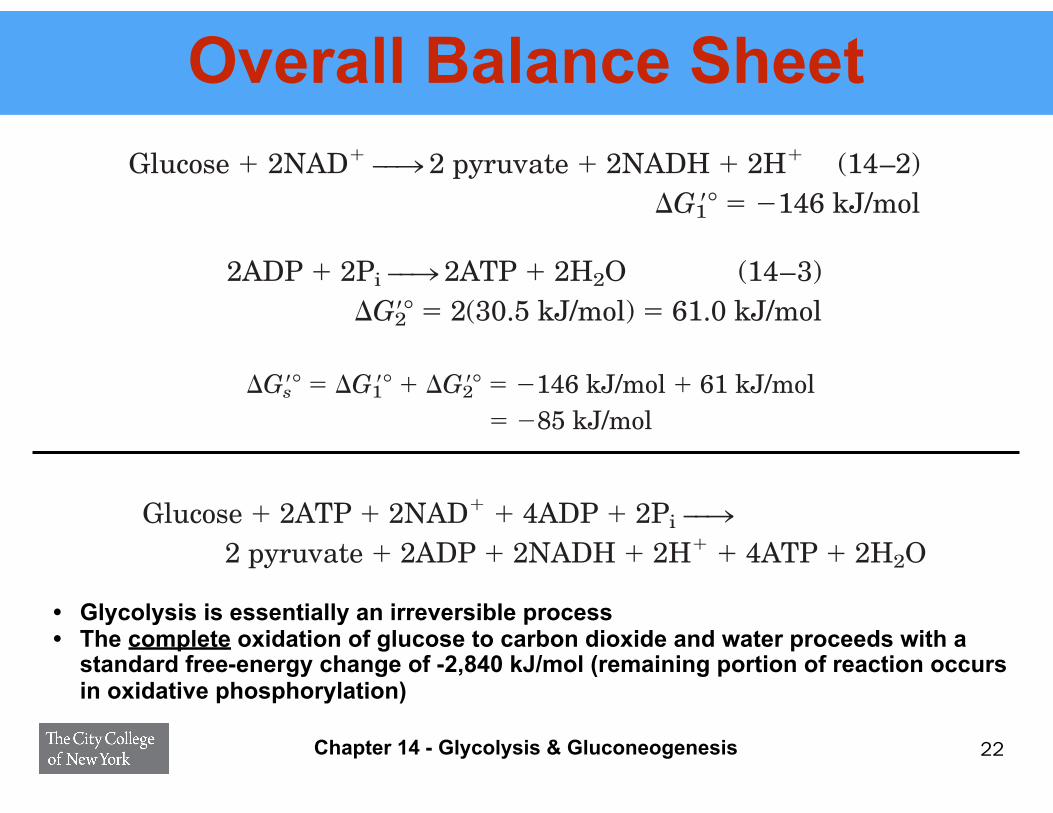
Overall Balance Sheet
22
14.1 Glycolysis 531
For each molecule of glucose degraded to pyruvate, twomolecules of ATP are generated from ADP and Pi, andtwo molecules of NADH are produced by the reductionof NAD!. The hydrogen acceptor in this reaction isNAD! (see Fig. 13–24), bound to a Rossmann fold asshown in Figure 13–25. The reduction of NAD! proceedsby the enzymatic transfer of a hydride ion (:H") from thealdehyde group of glyceraldehyde 3-phosphate to thenicotinamide ring of NAD!, yielding the reduced coen-zyme NADH. The other hydrogen atom of the substratemolecule is released to the solution as H!.
We can now resolve the equation of glycolysis intotwo processes—the conversion of glucose to pyruvate,which is exergonic:
Glucose ! 2NAD! 88n 2 pyruvate ! 2NADH ! 2H! (14–2)#G1$% & "146 kJ/mol
and the formation of ATP from ADP and Pi, which isendergonic:
2ADP ! 2Pi 88n 2ATP ! 2H2O (14–3)#G2$% & 2(30.5 kJ/mol) & 61.0 kJ/mol
The sum of Equations 14–2 and 14–3 gives the overallstandard free-energy change of glycolysis, #G$%s :
#Gs$% & #G1$% ! #G2$% & "146 kJ/mol ! 61 kJ/mol& "85 kJ/mol
Under standard conditions, and under the (nonstan-dard) conditions that prevail in a cell, glycolysis is an es-sentially irreversible process, driven to completion by alarge net decrease in free energy.
Energy Remaining in Pyruvate Glycolysis releasesonly a small fraction of the total available energy of theglucose molecule; the two molecules of pyruvate formedby glycolysis still contain most of the chemical potentialenergy of glucose, energy that can be extracted by ox-idative reactions in the citric acid cycle (Chapter 16)and oxidative phosphorylation (Chapter 19).
Importance of Phosphorylated IntermediatesEach of the nine glycolytic intermediates between glu-cose and pyruvate is phosphorylated (Fig. 14–2). Thephosphoryl groups seem to have three functions.
1. Because the plasma membrane generally lackstransporters for phosphorylated sugars, the phos-phorylated glycolytic intermediates cannot leavethe cell. After the initial phosphorylation, no fur-ther energy is necessary to retain phosphorylatedintermediates in the cell, despite the large differ-ence in their intracellular and extracellularconcentrations.
2. Phosphoryl groups are essential components in theenzymatic conservation of metabolic energy. Energyreleased in the breakage of phosphoanhydridebonds (such as those in ATP) is partially conserved
in the formation of phosphate esters such as glucose6-phosphate. High-energy phosphate compoundsformed in glycolysis (1,3-bisphosphoglycerate andphosphoenolpyruvate) donate phosphoryl groupsto ADP to form ATP.
3. Binding energy resulting from the binding of phos-phate groups to the active sites of enzymes lowersthe activation energy and increases the specificityof the enzymatic reactions (Chapter 6). The phos-phate groups of ADP, ATP, and the glycolytic inter-mediates form complexes with Mg2!, and thesubstrate binding sites of many glycolytic enzymesare specific for these Mg2! complexes. Most gly-colytic enzymes require Mg2! for activity.
The Preparatory Phase of Glycolysis Requires ATP
In the preparatory phase of glycolysis, two molecules ofATP are invested and the hexose chain is cleaved intotwo triose phosphates. The realization that phosphory-lated hexoses were intermediates in glycolysis cameslowly and serendipitously. In 1906, Arthur Harden andWilliam Young tested their hypothesis that inhibitors of proteolytic enzymes would stabilize the glucose-fermenting enzymes in yeast extract. They added bloodserum (known to contain inhibitors of proteolytic en-zymes) to yeast extracts and observed the predictedstimulation of glucose metabolism. However, in a controlexperiment intended to show that boiling the serum de-stroyed the stimulatory activity, they discovered thatboiled serum was just as effective at stimulating glycoly-sis! Careful examination and testing of the contents ofthe boiled serum revealed that inorganic phosphate wasresponsible for the stimulation. Harden and Young soondiscovered that glucose added to their yeast extract wasconverted to a hexose bisphosphate (the “Harden-Young ester,” eventually identified as fructose 1,6-bis-phosphate). This was the beginning of a long series ofinvestigations on the role of organic esters and anhy-drides of phosphate in biochemistry, which has led toour current understanding of the central role of phos-phoryl group transfer in biology.
Arthur Harden,1865–1940
William Young,1878–1942
14.1 Glycolysis 531
For each molecule of glucose degraded to pyruvate, twomolecules of ATP are generated from ADP and Pi, andtwo molecules of NADH are produced by the reductionof NAD!. The hydrogen acceptor in this reaction isNAD! (see Fig. 13–24), bound to a Rossmann fold asshown in Figure 13–25. The reduction of NAD! proceedsby the enzymatic transfer of a hydride ion (:H") from thealdehyde group of glyceraldehyde 3-phosphate to thenicotinamide ring of NAD!, yielding the reduced coen-zyme NADH. The other hydrogen atom of the substratemolecule is released to the solution as H!.
We can now resolve the equation of glycolysis intotwo processes—the conversion of glucose to pyruvate,which is exergonic:
Glucose ! 2NAD! 88n 2 pyruvate ! 2NADH ! 2H! (14–2)#G1$% & "146 kJ/mol
and the formation of ATP from ADP and Pi, which isendergonic:
2ADP ! 2Pi 88n 2ATP ! 2H2O (14–3)#G2$% & 2(30.5 kJ/mol) & 61.0 kJ/mol
The sum of Equations 14–2 and 14–3 gives the overallstandard free-energy change of glycolysis, #G$%s :
#Gs$% & #G1$% ! #G2$% & "146 kJ/mol ! 61 kJ/mol& "85 kJ/mol
Under standard conditions, and under the (nonstan-dard) conditions that prevail in a cell, glycolysis is an es-sentially irreversible process, driven to completion by alarge net decrease in free energy.
Energy Remaining in Pyruvate Glycolysis releasesonly a small fraction of the total available energy of theglucose molecule; the two molecules of pyruvate formedby glycolysis still contain most of the chemical potentialenergy of glucose, energy that can be extracted by ox-idative reactions in the citric acid cycle (Chapter 16)and oxidative phosphorylation (Chapter 19).
Importance of Phosphorylated IntermediatesEach of the nine glycolytic intermediates between glu-cose and pyruvate is phosphorylated (Fig. 14–2). Thephosphoryl groups seem to have three functions.
1. Because the plasma membrane generally lackstransporters for phosphorylated sugars, the phos-phorylated glycolytic intermediates cannot leavethe cell. After the initial phosphorylation, no fur-ther energy is necessary to retain phosphorylatedintermediates in the cell, despite the large differ-ence in their intracellular and extracellularconcentrations.
2. Phosphoryl groups are essential components in theenzymatic conservation of metabolic energy. Energyreleased in the breakage of phosphoanhydridebonds (such as those in ATP) is partially conserved
in the formation of phosphate esters such as glucose6-phosphate. High-energy phosphate compoundsformed in glycolysis (1,3-bisphosphoglycerate andphosphoenolpyruvate) donate phosphoryl groupsto ADP to form ATP.
3. Binding energy resulting from the binding of phos-phate groups to the active sites of enzymes lowersthe activation energy and increases the specificityof the enzymatic reactions (Chapter 6). The phos-phate groups of ADP, ATP, and the glycolytic inter-mediates form complexes with Mg2!, and thesubstrate binding sites of many glycolytic enzymesare specific for these Mg2! complexes. Most gly-colytic enzymes require Mg2! for activity.
The Preparatory Phase of Glycolysis Requires ATP
In the preparatory phase of glycolysis, two molecules ofATP are invested and the hexose chain is cleaved intotwo triose phosphates. The realization that phosphory-lated hexoses were intermediates in glycolysis cameslowly and serendipitously. In 1906, Arthur Harden andWilliam Young tested their hypothesis that inhibitors of proteolytic enzymes would stabilize the glucose-fermenting enzymes in yeast extract. They added bloodserum (known to contain inhibitors of proteolytic en-zymes) to yeast extracts and observed the predictedstimulation of glucose metabolism. However, in a controlexperiment intended to show that boiling the serum de-stroyed the stimulatory activity, they discovered thatboiled serum was just as effective at stimulating glycoly-sis! Careful examination and testing of the contents ofthe boiled serum revealed that inorganic phosphate wasresponsible for the stimulation. Harden and Young soondiscovered that glucose added to their yeast extract wasconverted to a hexose bisphosphate (the “Harden-Young ester,” eventually identified as fructose 1,6-bis-phosphate). This was the beginning of a long series ofinvestigations on the role of organic esters and anhy-drides of phosphate in biochemistry, which has led toour current understanding of the central role of phos-phoryl group transfer in biology.
Arthur Harden,1865–1940
William Young,1878–1942
14.1 Glycolysis 531
For each molecule of glucose degraded to pyruvate, twomolecules of ATP are generated from ADP and Pi, andtwo molecules of NADH are produced by the reductionof NAD!. The hydrogen acceptor in this reaction isNAD! (see Fig. 13–24), bound to a Rossmann fold asshown in Figure 13–25. The reduction of NAD! proceedsby the enzymatic transfer of a hydride ion (:H") from thealdehyde group of glyceraldehyde 3-phosphate to thenicotinamide ring of NAD!, yielding the reduced coen-zyme NADH. The other hydrogen atom of the substratemolecule is released to the solution as H!.
We can now resolve the equation of glycolysis intotwo processes—the conversion of glucose to pyruvate,which is exergonic:
Glucose ! 2NAD! 88n 2 pyruvate ! 2NADH ! 2H! (14–2)#G1$% & "146 kJ/mol
and the formation of ATP from ADP and Pi, which isendergonic:
2ADP ! 2Pi 88n 2ATP ! 2H2O (14–3)#G2$% & 2(30.5 kJ/mol) & 61.0 kJ/mol
The sum of Equations 14–2 and 14–3 gives the overallstandard free-energy change of glycolysis, #G$%s :
#Gs$% & #G1$% ! #G2$% & "146 kJ/mol ! 61 kJ/mol& "85 kJ/mol
Under standard conditions, and under the (nonstan-dard) conditions that prevail in a cell, glycolysis is an es-sentially irreversible process, driven to completion by alarge net decrease in free energy.
Energy Remaining in Pyruvate Glycolysis releasesonly a small fraction of the total available energy of theglucose molecule; the two molecules of pyruvate formedby glycolysis still contain most of the chemical potentialenergy of glucose, energy that can be extracted by ox-idative reactions in the citric acid cycle (Chapter 16)and oxidative phosphorylation (Chapter 19).
Importance of Phosphorylated IntermediatesEach of the nine glycolytic intermediates between glu-cose and pyruvate is phosphorylated (Fig. 14–2). Thephosphoryl groups seem to have three functions.
1. Because the plasma membrane generally lackstransporters for phosphorylated sugars, the phos-phorylated glycolytic intermediates cannot leavethe cell. After the initial phosphorylation, no fur-ther energy is necessary to retain phosphorylatedintermediates in the cell, despite the large differ-ence in their intracellular and extracellularconcentrations.
2. Phosphoryl groups are essential components in theenzymatic conservation of metabolic energy. Energyreleased in the breakage of phosphoanhydridebonds (such as those in ATP) is partially conserved
in the formation of phosphate esters such as glucose6-phosphate. High-energy phosphate compoundsformed in glycolysis (1,3-bisphosphoglycerate andphosphoenolpyruvate) donate phosphoryl groupsto ADP to form ATP.
3. Binding energy resulting from the binding of phos-phate groups to the active sites of enzymes lowersthe activation energy and increases the specificityof the enzymatic reactions (Chapter 6). The phos-phate groups of ADP, ATP, and the glycolytic inter-mediates form complexes with Mg2!, and thesubstrate binding sites of many glycolytic enzymesare specific for these Mg2! complexes. Most gly-colytic enzymes require Mg2! for activity.
The Preparatory Phase of Glycolysis Requires ATP
In the preparatory phase of glycolysis, two molecules ofATP are invested and the hexose chain is cleaved intotwo triose phosphates. The realization that phosphory-lated hexoses were intermediates in glycolysis cameslowly and serendipitously. In 1906, Arthur Harden andWilliam Young tested their hypothesis that inhibitors of proteolytic enzymes would stabilize the glucose-fermenting enzymes in yeast extract. They added bloodserum (known to contain inhibitors of proteolytic en-zymes) to yeast extracts and observed the predictedstimulation of glucose metabolism. However, in a controlexperiment intended to show that boiling the serum de-stroyed the stimulatory activity, they discovered thatboiled serum was just as effective at stimulating glycoly-sis! Careful examination and testing of the contents ofthe boiled serum revealed that inorganic phosphate wasresponsible for the stimulation. Harden and Young soondiscovered that glucose added to their yeast extract wasconverted to a hexose bisphosphate (the “Harden-Young ester,” eventually identified as fructose 1,6-bis-phosphate). This was the beginning of a long series ofinvestigations on the role of organic esters and anhy-drides of phosphate in biochemistry, which has led toour current understanding of the central role of phos-phoryl group transfer in biology.
Arthur Harden,1865–1940
William Young,1878–1942
• Glycolysis is essentially an irreversible process • The complete oxidation of glucose to carbon dioxide and water proceeds with a
standard free-energy change of -2,840 kJ/mol (remaining portion of reaction occurs in oxidative phosphorylation)
Glycolysis, Gluconeogenesis, and the Pentose Phosphate Pathway538
enzyme. Phosphoglycerate mutase is initially phospho-rylated by phosphoryl transfer from 2,3-BPG, which isrequired in small quantities to initiate the catalytic cycleand is continuously regenerated by that cycle.
9 Dehydration of 2-Phosphoglycerate to Phos-phoenolpyruvate In the second glycolytic reactionthat generates a compound with high phosphoryl grouptransfer potential (the first was step 6 ), enolasepromotes reversible removal of a molecule of water from2-phosphoglycerate to yield phosphoenolpyruvate(PEP):
The mechanism of the enolase reaction involves an eno-lic intermediate stabilized by Mg2! (see Fig. 6–23). Thereaction converts a compound with a relatively lowphosphoryl group transfer potential ("G#$ for hydrolysisof 2-phosphoglycerate is %17.6 kJ/mol) to one with highphosphoryl group transfer potential ("G#$ for PEP hy-drolysis is %61.9 kJ/mol) (see Fig. 13–13, Table 13–6).
10 Transfer of the Phosphoryl Group from Phos-phoenolpyruvate to ADP The last step in glycolysisis the transfer of the phosphoryl group from phospho-enolpyruvate to ADP, catalyzed by pyruvate kinase,which requires K! and either Mg2! or Mn2!:
The overall reaction has a large, negative standard free-energy change, due in large part to the spontaneousconversion of the enol form of pyruvate to the keto form(see Fig. 13–13). About half of the energy released byPEP hydrolysis ("G#$ & %61.9 kJ/mol) is conserved inthe formation of the phosphoanhydride bond of ATP("G#$ & %30.5 kJ/mol), and the rest (%31.4 kJ/mol)constitutes a large driving force pushing the reaction to-ward ATP synthesis. We discuss the regulation of pyru-vate kinase in Chapter 15.
The Overall Balance Sheet Shows a Net Gain of ATP
We can now construct a balance sheet for glycolysis toaccount for (1) the fate of the carbon skeleton of glu-cose, (2) the input of Pi and ADP and output of ATP, and(3) the pathway of electrons in the oxidation-reductionreactions. The left-hand side of the following equationshows all the inputs of ATP, NAD!, ADP, and Pi (consultFig. 14–2), and the right-hand side shows all the outputs(keep in mind that each molecule of glucose yields twomolecules of pyruvate):
Glucose ! 2ATP ! 2NAD! ! 4ADP ! 2Pi 88n2 pyruvate ! 2ADP ! 2NADH ! 2H! ! 4ATP ! 2H2O
Canceling out common terms on both sides of the equa-tion gives the overall equation for glycolysis under aero-bic conditions:
Glucose ! 2NAD! ! 2ADP ! 2Pi 88n2 pyruvate ! 2NADH ! 2H! ! 2ATP ! 2H2O
The two molecules of NADH formed by glycolysis inthe cytosol are, under aerobic conditions, reoxidized toNAD! by transfer of their electrons to the electron-transfer chain, which in eukaryotic cells is located in themitochondria. The electron-transfer chain passes theseelectrons to their ultimate destination, O2:
Electron transfer from NADH to O2 in mitochondria pro-vides the energy for synthesis of ATP by respiration-linked phosphorylation (Chapter 19).
In the overall glycolytic process, one molecule ofglucose is converted to two molecules of pyruvate (thepathway of carbon). Two molecules of ADP and two ofPi are converted to two molecules of ATP (the pathway
2NADH ! 2H! ! O2 ¡ 2NAD! ! 2H2O
In this substrate-level phosphorylation, the productpyruvate first appears in its enol form, then tauto-merizes rapidly and nonenzymatically to its keto form,which predominates at pH 7:
O% %
C
CH2
H2O
enolase
O
CH
CH2
7.5 kJ/mol
2-Phosphoglycerate Phosphoenolpyruvate
O O
HO
C C
&"G#$
OPO32% OPO3
2%
P
O
P
O
O
P
%
%
%
%
O
C
C
CH3
!
O O
O
P
AdenineRib
, K! pyruvatekinase
O
O
C
C
CH2
P
P
AdenineRib
O%
!
ATP
Phosphoenolpyruvate
Pyruvate
ADP
O% O
Mg2!
O
O
& %31.4 kJ/mol"G#°
O% %
C
CH3
Otautomerization
O
C
CH2
Pyruvate(enol form)
Pyruvate(keto form)
O O
OH
C C
Chapter 14 - Glycolysis & Gluconeogenesis

Why Many Phosphorylated Intermediates?
23
All products/reactants after glucose are phosphorylated
• Glucose is transported into cell via facilitated passive diffusion by a carrier (GLUT). This carrier is reversible.
• Phosphorylation changes charge and structure of glucose. This inhibits its transport through GLUT and out of cell.
• Energy released in hydrolysis of phosphodiester bond of ATP is partially retained in the phosphodiester bond of a product/reactant.
• High energy intermediates (BPG and PEP) can transfer phosphate to ADP to make ATP.
• Binding energy of phosphate intermediates to enzymes is increased because of phosphate. This helps drive catalysis.
Chapter 14 - Glycolysis & Gluconeogenesis
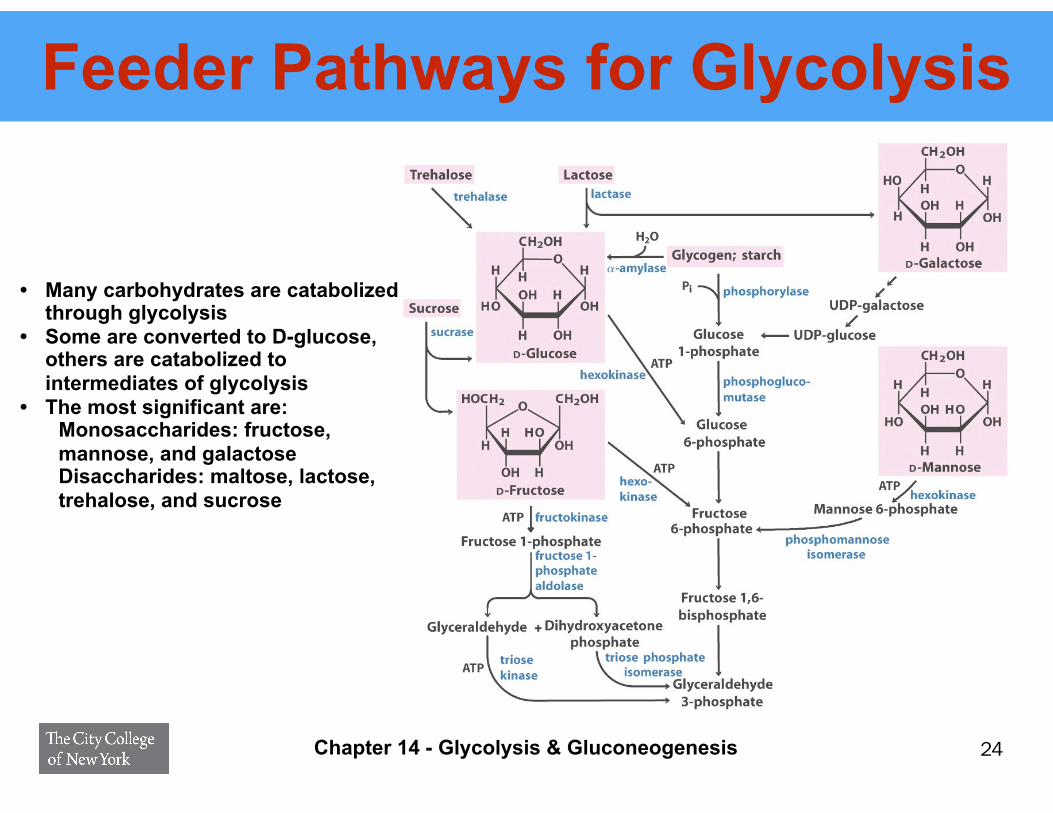
Feeder Pathways for Glycolysis
• Many carbohydrates are catabolized through glycolysis
• Some are converted to D-glucose, others are catabolized to intermediates of glycolysis
• The most significant are: Monosaccharides: fructose, mannose, and galactose Disaccharides: maltose, lactose, trehalose, and sucrose
24Chapter 14 - Glycolysis & Gluconeogenesis










![Biochem [Gluconeogenesis]](https://img.pdfslide.net/doc/110x75/577c82b31a28abe054b1e4af/biochem-gluconeogenesis.jpg)









![Gluconeogenesis [Compatibility Mode]](https://img.pdfslide.net/doc/110x75/577ce5671a28abf103908ef8/gluconeogenesis-compatibility-mode.jpg)