Embed Size (px)
Citation preview

Lecture 23
– Quiz next Mon. on Pentose Phosphate Pathway
– Metabolic regulation and control of glycolysis/gluconeogenesis
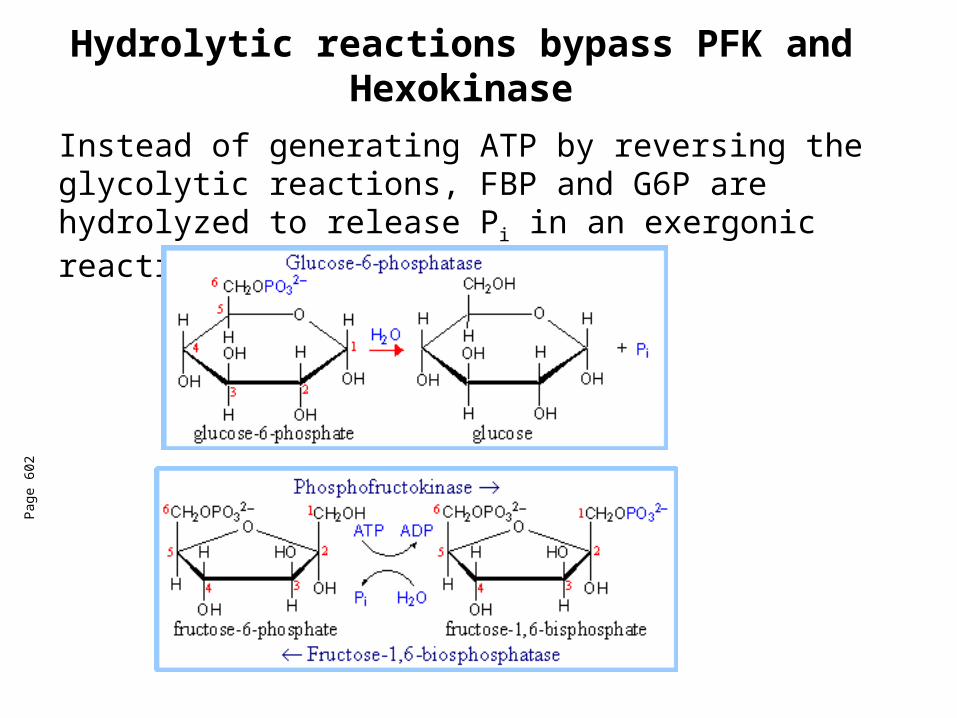
Hydrolytic reactions bypass PFK and Hexokinase
Pag
e 60
2
Instead of generating ATP by reversing the glycolytic reactions, FBP and G6P are hydrolyzed to release Pi in an exergonic reaction.
Pag
e 84
8
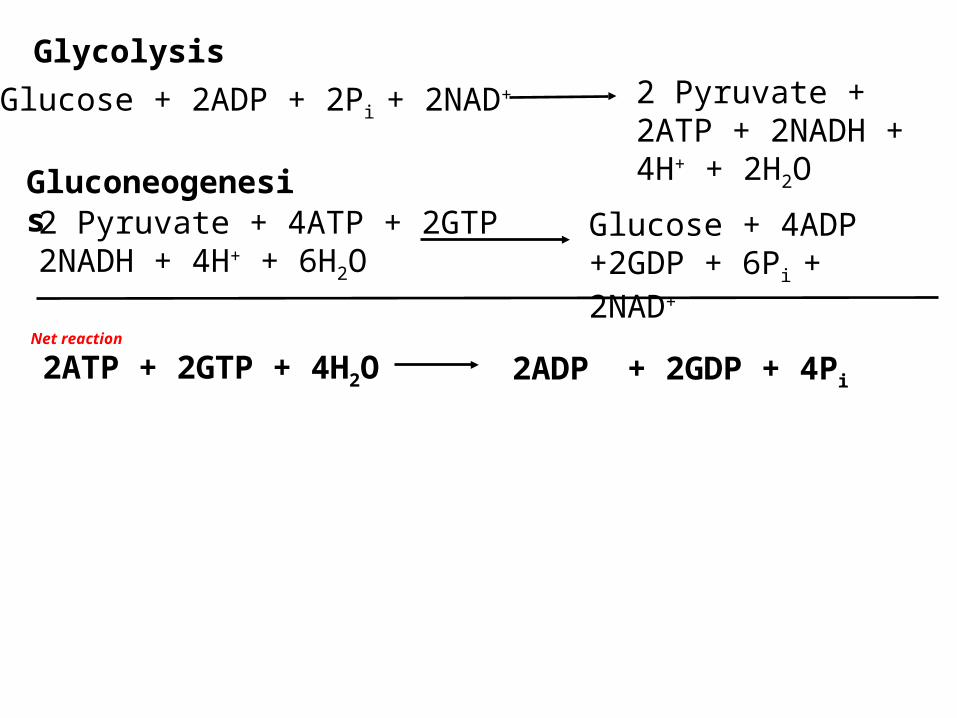
Gluconeogenesis
Glucose + 2ADP + 2Pi + 2NAD+ 2 Pyruvate + 2ATP + 2NADH + 4H+ + 2H2O
Net reaction
2ADP + 2GDP + 4Pi2ATP + 2GTP + 4H2O
Glycolysis
2 Pyruvate + 4ATP + 2GTP 2NADH + 4H+ + 6H2O
Glucose + 4ADP +2GDP + 6Pi + 2NAD+
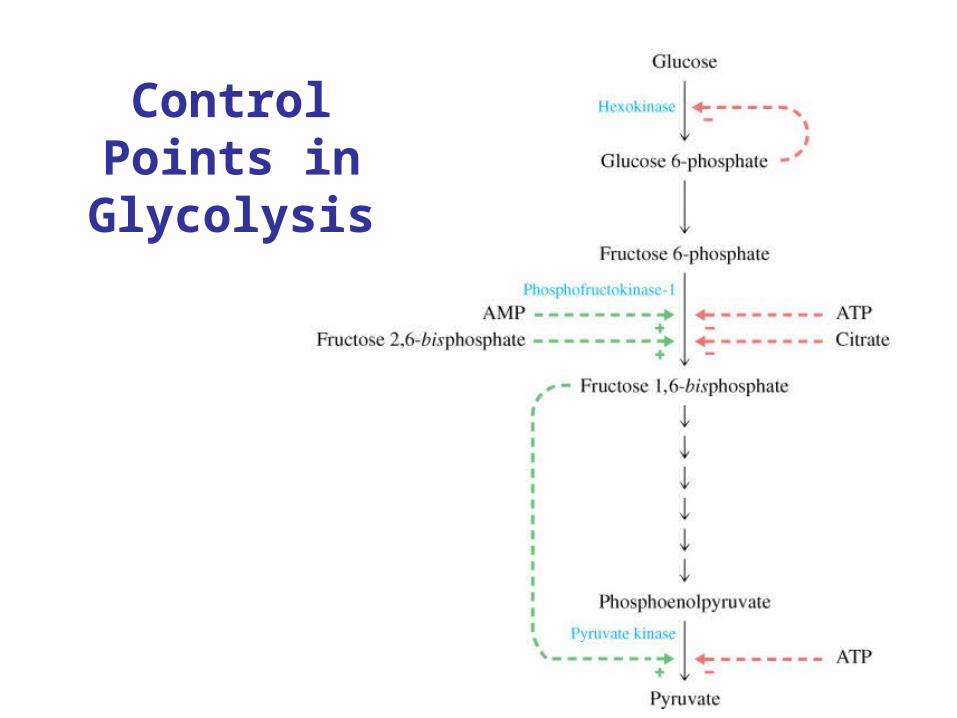
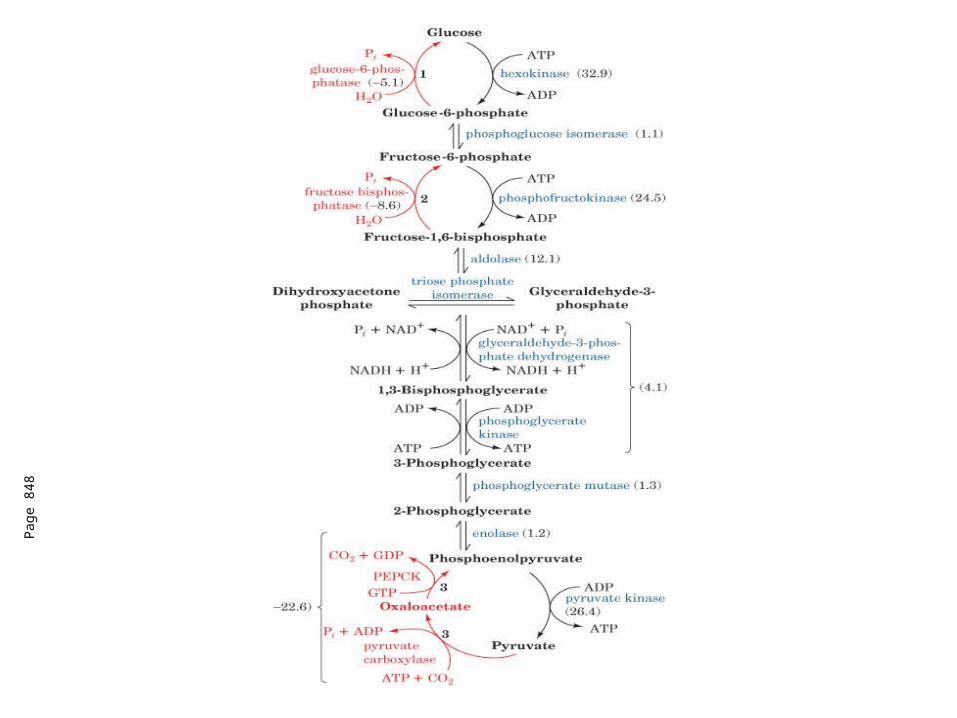
Control Points in Glycolysis
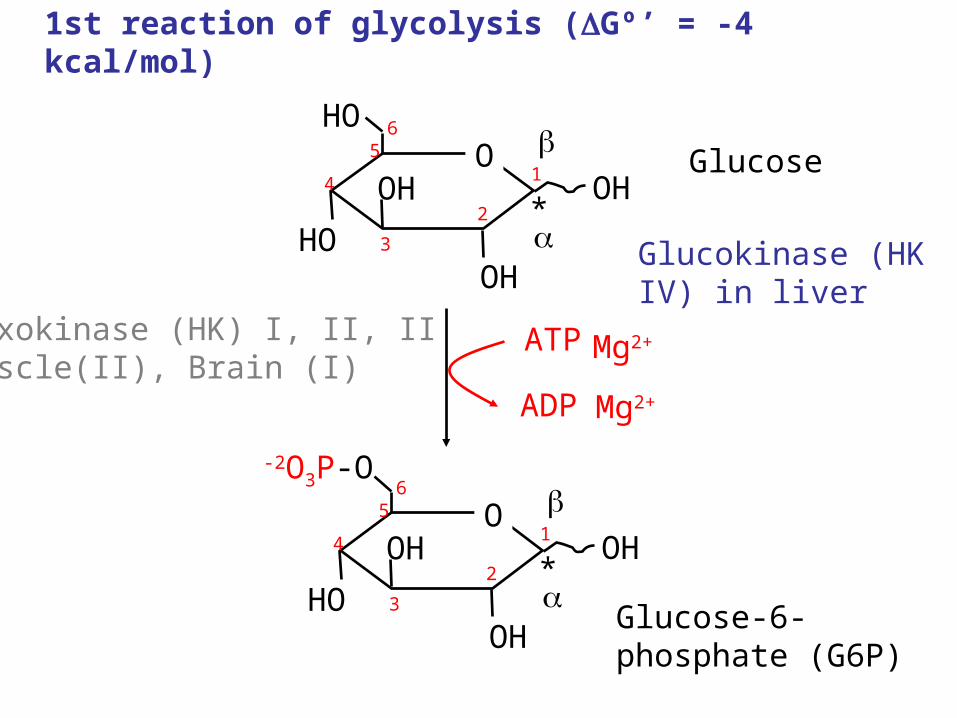
1st reaction of glycolysis (Gº’ = -4 kcal/mol)
OH1
OHO
OH
HOOH
*2
3
4
56
Glucose
OH1
O
-2O3P-O
OH
HOOH
*2
3
4
56
ATP
ADP
Glucose-6-phosphate (G6P)
Hexokinase (HK) I, II, IIMuscle(II), Brain (I)
Mg2+
Mg2+
Glucokinase (HK IV) in liver

Regulation of Hexokinase
• Glucose-6-phosphate is an allosteric inhibitor of hexokinase.
• Levels of glucose-6-phosphate increase when downstream steps are inhibited.
• This coordinates the regulation of hexokinase with other regulatory enzymes in glycolysis.
• Hexokinase is not necessarily the first regulatory step inhibited.

Types of regulation
1. Availability of substrate Glucokinase (KM 12 mM) vs. HK (KM = 0.01 - 0.03 mM)
2. Compartmentalization -Brain vs. Liver vs. Muscle (type I mitochondrial membrane, type II cytoplasmic)
3. Allosteric regulation - feedback inhibition by G-6-P, overcome by Pi in type I (Brain/ mitochondrial controlled by Pi levels)
4. Hormonal regulation. Liver has HK as fetal tissue. Changes to glucokinase after about 2 weeks. If there is no dietary carbohydrate, no glucokinase. Must have both insulin and carbohydrates to induce.

2 places where there is no net reaction
1. ATP + F-6-P F-1,6-P2 + ADP
2. F-1,6-P2 F-6-P + Pi
PFK
F-phosphataseMg2+
Mg2+
Net: ATP ADP + Pi + heat
Similar reaction occurs with hexokinase and G-6-phosphatase.Generally regulated so this does not occur (futile cycle).
May function in hibernating animals to generate heat.
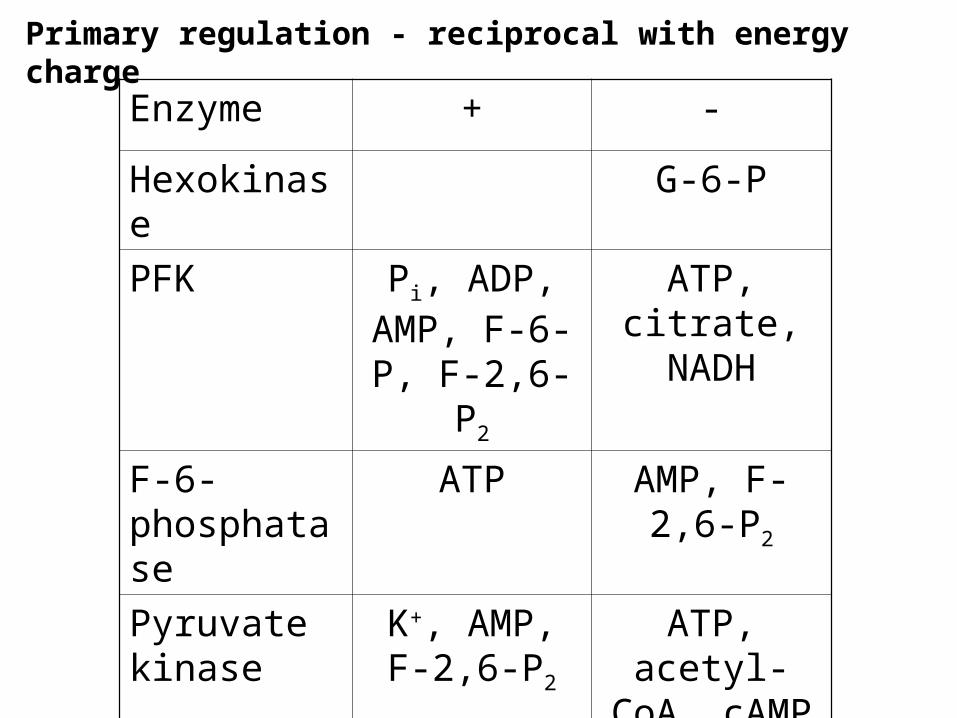
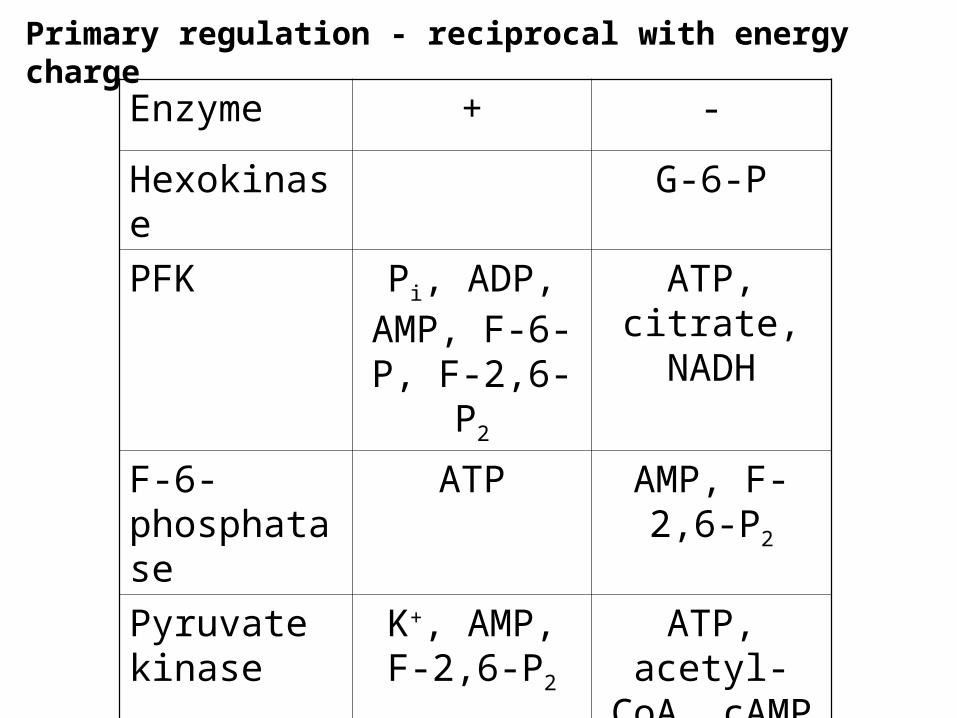
Primary regulation - reciprocal with energy charge
Enzyme + -
Hexokinase G-6-P
PFK Pi, ADP, AMP, F-6-P,
F-2,6-P2
ATP, citrate, NADH
F-6-phosphatase
ATP AMP, F-2,6-P2
Pyruvate kinase
K+, AMP, F-2,6-P2
ATP, acetyl-CoA, cAMP
Pyruvate carboxylase
Acetyl-CoA

Major regulation is through energy charge
ATP
ATP
Gluconeogenesis
GlycolysisADP
Same reactions make AMP or ADP (primarily in lipid and nucleotide metabolism)
AMP + ATP 2 ADPAdenylate kinase
[ATP] +1/2[ADP]
[AMP] + [ADP] + [ATP]Energy charge
1.0 = 100% ATP Body generally likes it close to 0.90.5 = 100% ADP0 = 100% AMP
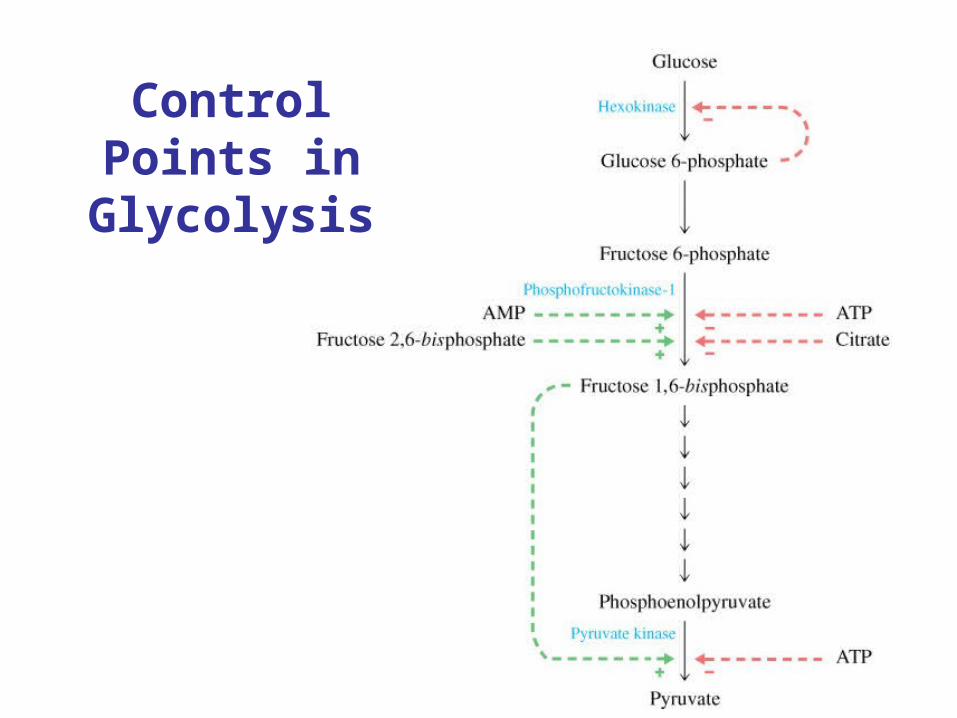
Control Points in Glycolysis
Primary regulation - reciprocal with energy charge
Enzyme + -
Hexokinase G-6-P
PFK Pi, ADP, AMP, F-6-P,
F-2,6-P2
ATP, citrate, NADH
F-6-phosphatase
ATP AMP, F-2,6-P2
Pyruvate kinase
K+, AMP, F-2,6-P2
ATP, acetyl-CoA, cAMP
Pyruvate carboxylase
Acetyl-CoA

Regulation of PhosphoFructokinase (PFK-1)
• PKF-1 has quaternary structure• Inhibited by ATP and Citrate• Activated by AMP and Fructose-2,6-
bisphosphate• Regulation related to energy status of cell.

PFK-1 regulation by adenosine nucleotides
• ATP is substrate and inhibitor. Binds to active site and allosteric site on PFK. Binding of ATP to allosteric site increase Km for ATP
• AMP and ADP are allosteric activators of PFK. • AMP relieves inhibition by ATP.
• ADP decreases Km for ATP
• Glucagon (a pancreatic hormone) produced in response to low blood glucose triggers cAMP signaling pathway that ultimately results in decreased glycolysis.
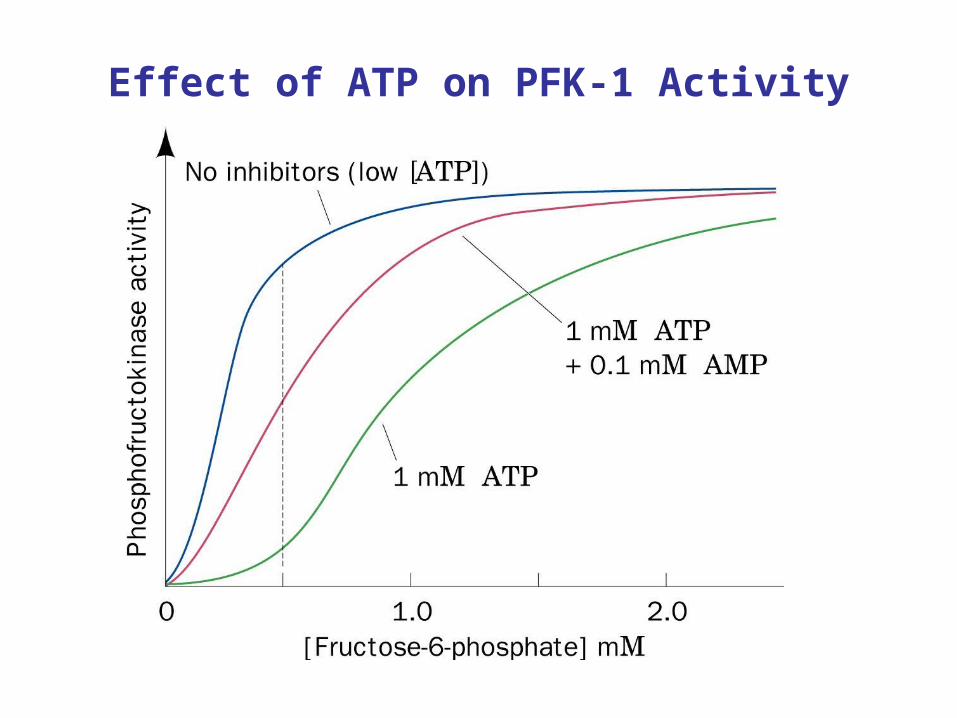
Effect of ATP on PFK-1 Activity
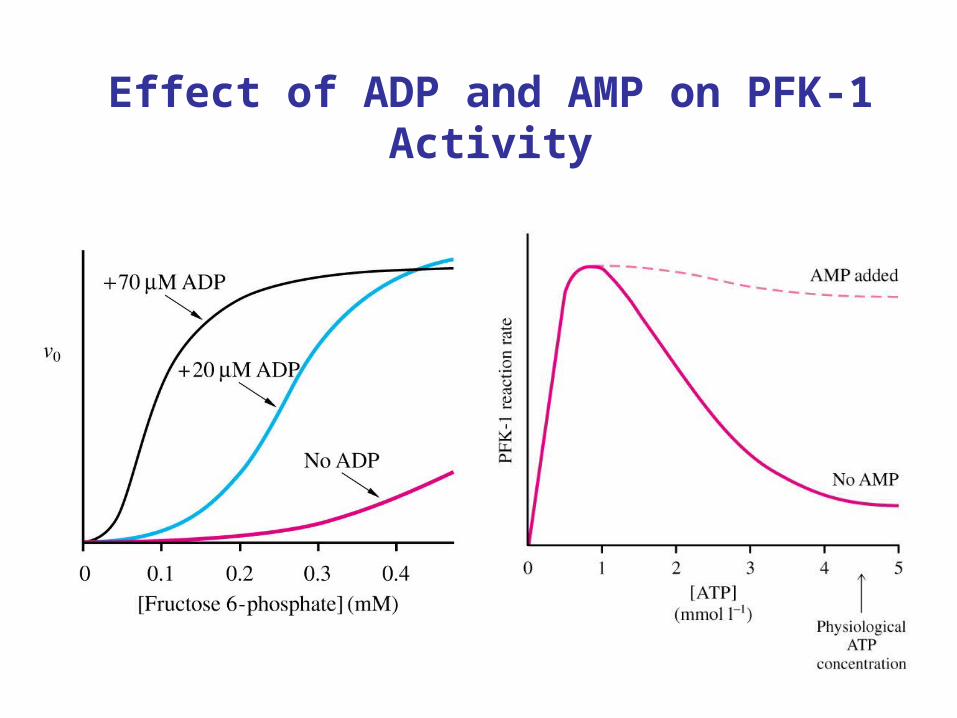
Effect of ADP and AMP on PFK-1 Activity

Regulation of PFK by Fructose-2,6-bisphosphate
• Fructose-2,6-bisphosphate is an allosteric activator of PFK in eukaryotes, but not prokaryotes
•Formed from fructose-6-phosphate by PFK-2
•Degraded to fructose-6-phosphate by fructose 2,6-bisphosphatase.
•In mammals the 2 activities are on the same enzyme
•PFK-2 inhibited by Pi and stimulated by citrate
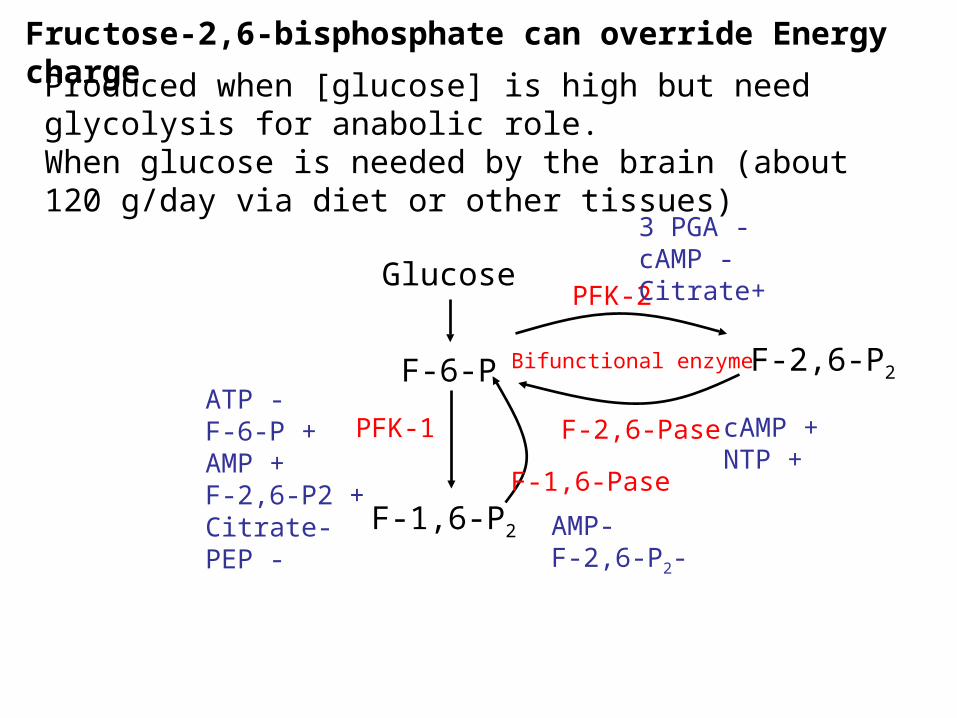
Fructose-2,6-bisphosphate can override Energy chargeProduced when [glucose] is high but need glycolysis for
anabolic role.When glucose is needed by the brain (about 120 g/day via diet or other tissues)
Glucose
F-6-P
F-1,6-P2
F-2,6-P2
F-1,6-Pase
F-2,6-Pase
PFK-2
PFK-1
Bifunctional enzyme
ATP -F-6-P +AMP +F-2,6-P2 +Citrate-PEP -
AMP-F-2,6-P2-
cAMP +NTP +
3 PGA -cAMP -Citrate+

Glucagon Regulation of PFK-1 in Liver•PFK-1 normally inhibited by ATP
•G-Protein mediated cAMP signaling pathway
•Induces protein kinase A that activates phosphatase activity and inhibits kinase activity
•Results in lower F-2,6-P levels decrease PFK-1 activity (less glycolysis)

PFK-2
1. Serves to override ATP inhibition and promote glycolysis once intermediates build up [citrate] [PEP][GAP]
2. Block PFK-2 activity with high [NTP] by stimulating F-2,6-Pase
This will break down F-2,6-P2 and restores energy charge regulation.
cAMP is the hormonal control. The presence of cAMP is indicative of low blood sugar (glucagon) stimulates F-2,6-Pase to increase F-6-P formtion for gluconeogenesis (cAMP also inhibits Pyruvate Kinase).

Regulation of Pyruvate Kinase
• Allosteric enzyme• Activated by Fructose-1,6-bisphosphate (example of
feed-forward regulation)• Inhibited by ATP• When high fructose 1,6-bisphosphate present plot of
[S] vs Vo goes from sigmoidal to hyperbolic.• Increasing ATP concentration increases Km for PEP. • In liver, PK also regulated by glucagon. Protein
kinase A phosphorylates PK and decreases PK acitivty.
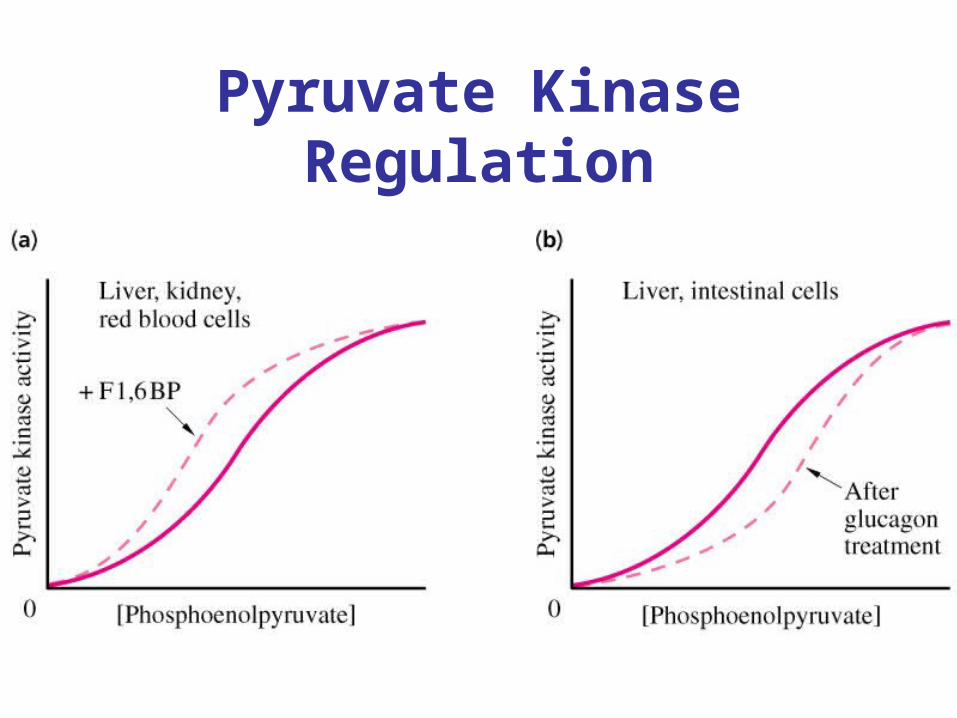
Pyruvate Kinase Regulation

Deregulation of Glycolysis in Cancer Cells
• Glucose uptake and glycolysis is 10X faster in solid tumors than in non-cancerous tissues.
• Tumor cells initally lack connection to blood supply so limited oxygen supply
• Tumor cells have fewer mitochondrial, depend more on glycolysis for ATP
• Increase levels of glycolytic enzymes in tumors (oncogene Ras and tumor suppressor gene p53 involved)

Glycogen biosynthesis
Most important storage form of sugar Glycogen - highly branched (1 per 10) polymer of glucose with (1,4) backbone and (1,6) branch points. More branched than starch so more free ends.
Average molecular weight -several million in liver, muscle.1/3 in liver (more concentrated but less overall mass (5-8%)),2/3 in muscle (1%).
Not found in brain - brain requires free glucose (120 g/ day) supplied in diet or from breakdown of glycogen in the liver.
Glucose levels regulated by several key hormones - insulin, glucagon.
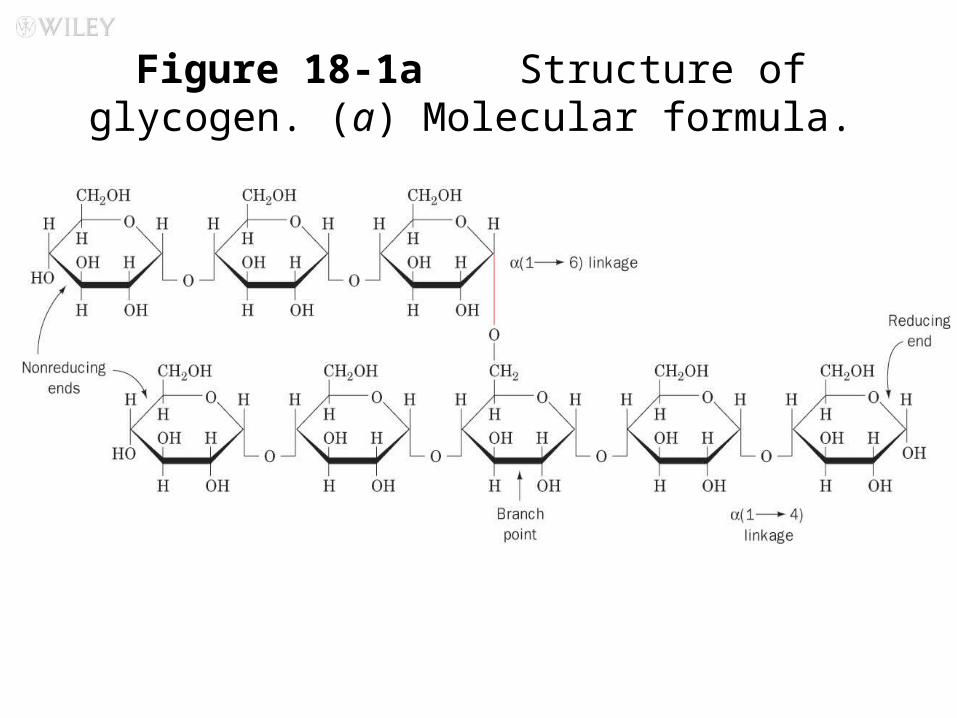
Figure 18-1a Structure of glycogen. (a) Molecular formula.
Pag
e 62
7
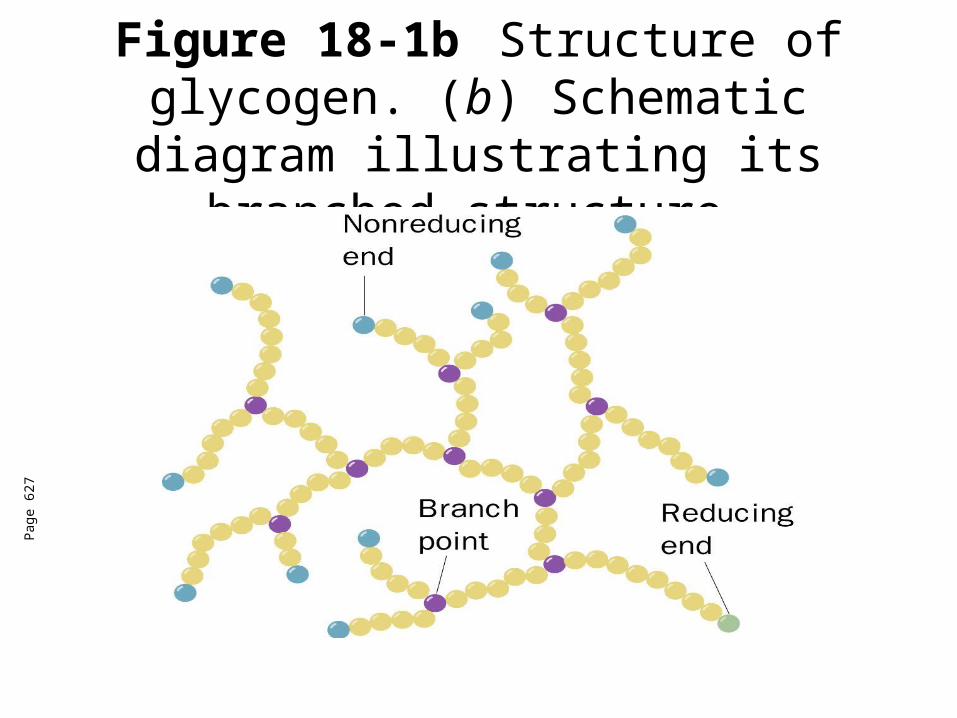
Figure 18-1bStructure of glycogen. (b) Schematic diagram illustrating its
branched structure.
Pag
e 62
7
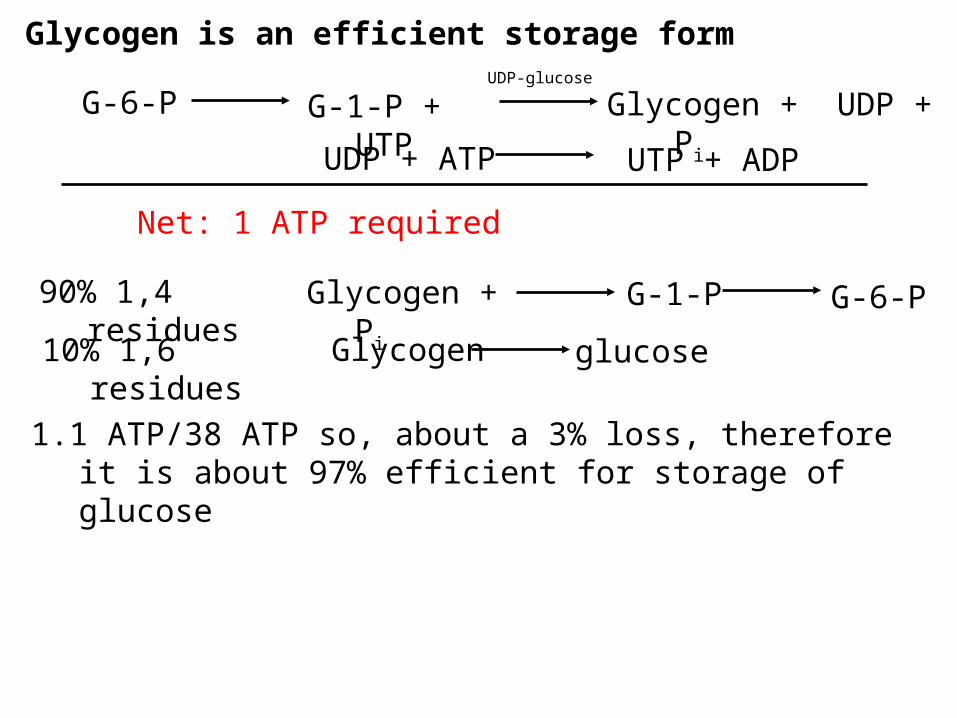
Glycogen is an efficient storage form
G-1-P + UTP Glycogen + UDP + Pi
UDP-glucose
Net: 1 ATP required
Glycogen + Pi
1.1 ATP/38 ATP so, about a 3% loss, therefore it is about 97% efficient for storage of glucose
G-6-P
UDP + ATP UTP + ADP
90% 1,4 residues G-1-P G-6-P
10% 1,6 residues Glycogen glucose

Glycogen biosynthesis
3 enzymes catalyze the steps involved in glycogen synthesis:
UDP-glucose pyrophosphorylaseGlycogen synthaseGlycogen branching enzyme
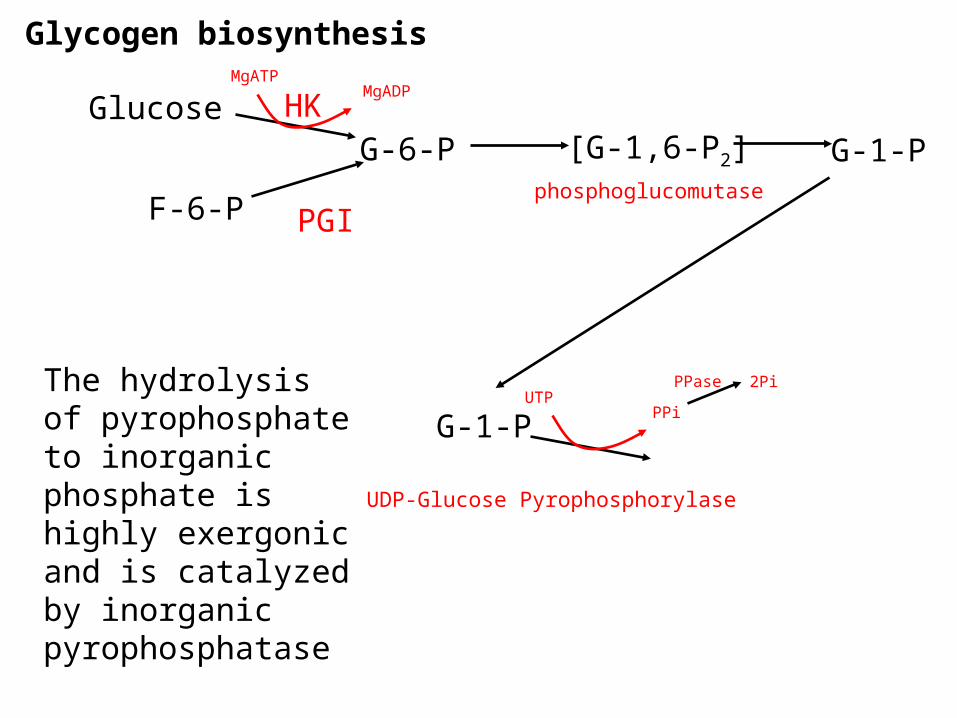
Glycogen biosynthesis
G-6-PGlucose
F-6-P
G-1-P[G-1,6-P2]
PGI
HKMgATP
MgADP
phosphoglucomutase
G-1-PUTP
PPi
UDP-Glucose Pyrophosphorylase
PPase 2PiThe hydrolysis of pyrophosphate to inorganic phosphate is highly exergonic and is catalyzed by inorganic pyrophosphatase
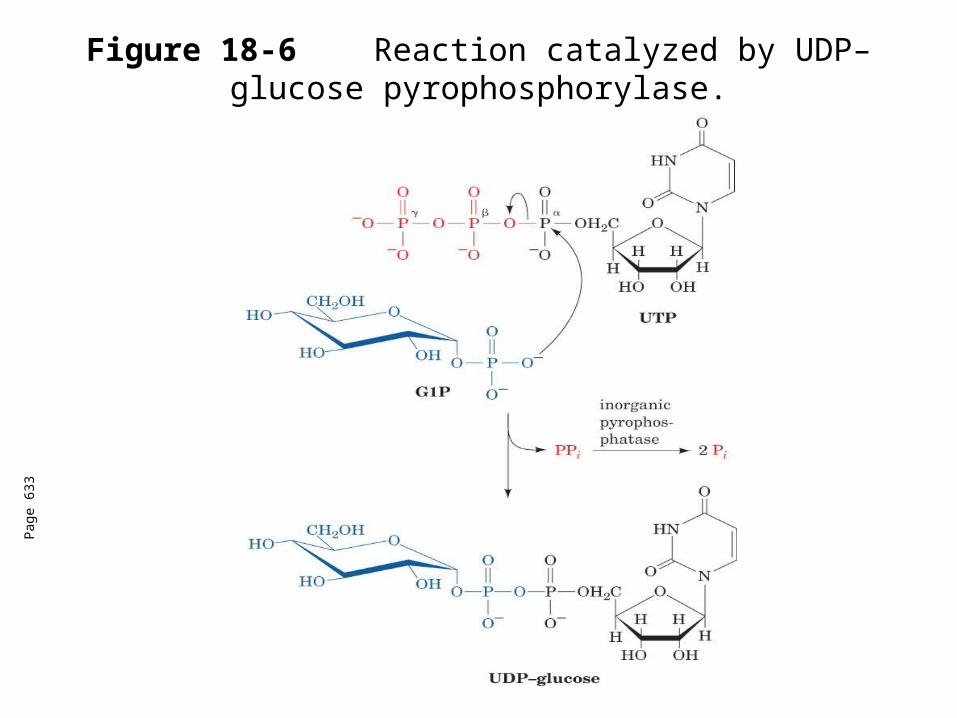
Figure 18-6 Reaction catalyzed by UDP–glucose pyrophosphorylase.
Pag
e 63
3

UDP-Glucose pyrophosphorylase
Coupling the highly exergonic cleavage of a nucleoside triphosphate to form PPi is a common biosynthetic strategy.
The free energy of the hydrolysis of PPi with the NTP hydrolysis drives the reaction forward.

Glycogen synthase
In this step, the glucosyl unit of UDP-glucose (UDPG) is transferred to the C4-OH group of one of glycogen’s nonreducing ends to form an (1,4) glycosidic bond.
Involves an oxonium ion intermediate (half-chair intermediate)
Each molecule of G1P added to glycogen regenerated needs one molecule of UTP hydrolyzed to UDP and Pi.
UTP is replenished by nucleoside diphosphate kinase
UDP + ATP UTP + ADP
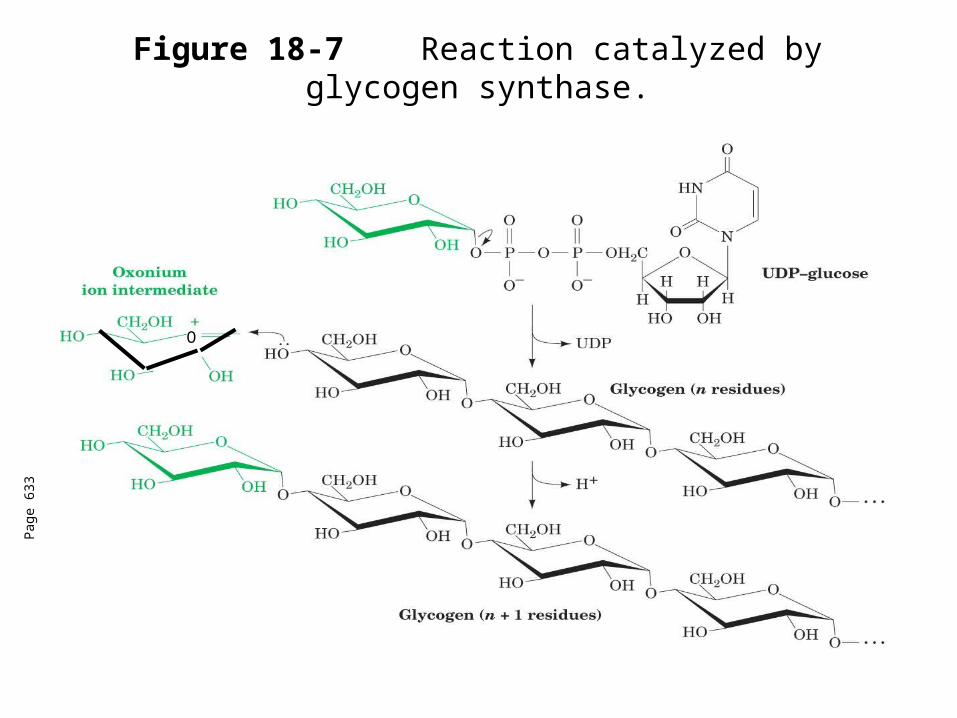
Figure 18-7 Reaction catalyzed by glycogen synthase.
Pag
e 63
3
O

Glycogen synthase
All carbohydrate biosynthesis occurs via UDP-sugars
Can only extend an already (1,4) linked glucan change.
First step is mediated by glycogenin, where glucose is attached to Tyr 194OH group.
The protein dissociates after glycogen reaches a minimum size.

Glycogen branching
Catalyzed by amylo (1,41,6)-transglycosylase (branching enzyme)
Branches are created by the terminal chain segments consisting of 7 glycosyl residues to the C6-OH groups of glucose residues on another chain.
Each transferred segment must be at least 11 residues.
Each new branch point at least 4 residues away from other branch points.
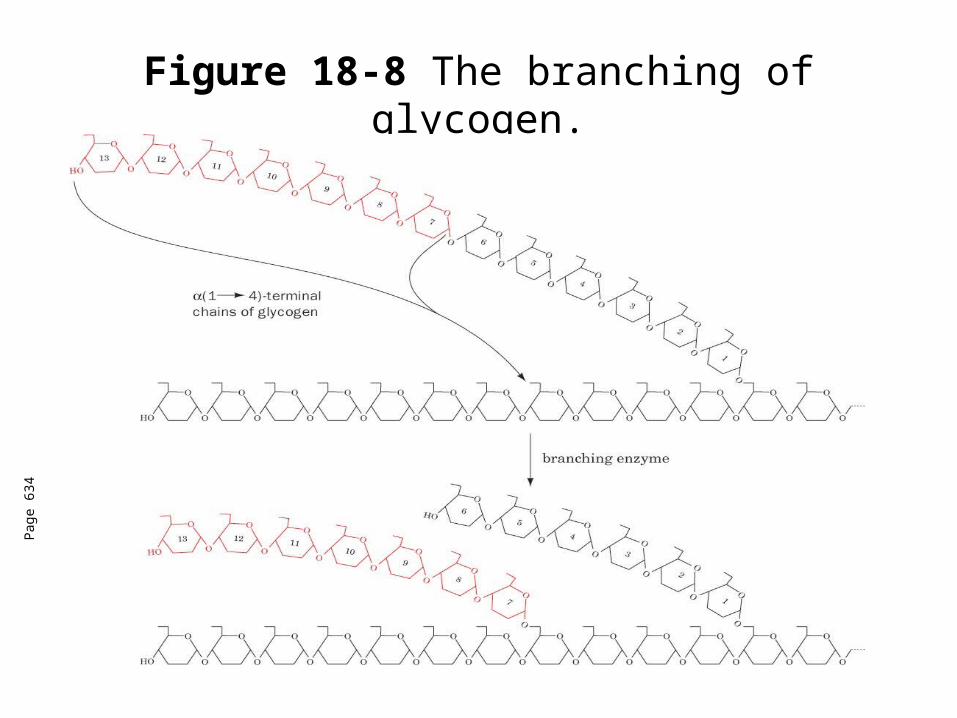
Figure 18-8 The branching of glycogen.
Pag
e 63
4

Glycogen Breakdown
Requires 3 enzymes: 1. Glycogen phosphorylase (phosphorylase) catalyzes
glycogen phosphorylysis (bond cleavage by the substitution of a phosphate group) and yields glucose-1-phosphate (G1P)
2. Glycogen debranching enzyme removes glycogen’s branches, allowing glycogen phosphorylase to complete it’s reactions. It also hydrolyzes a(16)-linked glucosyl units to yield glucose. 92% of glycogen’s glucse residues are converted to G1P and 8% to glucose.
3. Phosphoglucomutase converts G1P to G6P-can either go through glycolysis (muscle cells) or converted to glucose (liver).

Glycogen Phosphorylase
A dimer - 2 identical 842 residue subunits.Catalyzes the controlling step of glycogen breakdown.Regulated by allosteric interactions and covalent modification.Two forms of phosphorylase made by regulationPhosphorylase a- has a phosphoryl group on Ser14 in each
subunit.Phosphorylase b-lacks the phosphoryl groups.Inhibitors: ATP, G6P, glucoseActivator: AMPGlycogen forms a left-handed helix with 6.5 glucose residues
per turn. Structure can accommodate 4-5 sugar residues only.Pyridoxal phosphate is an essential cofactor for
phosphorylase.Converts glucosyl units of glycogen to G1P
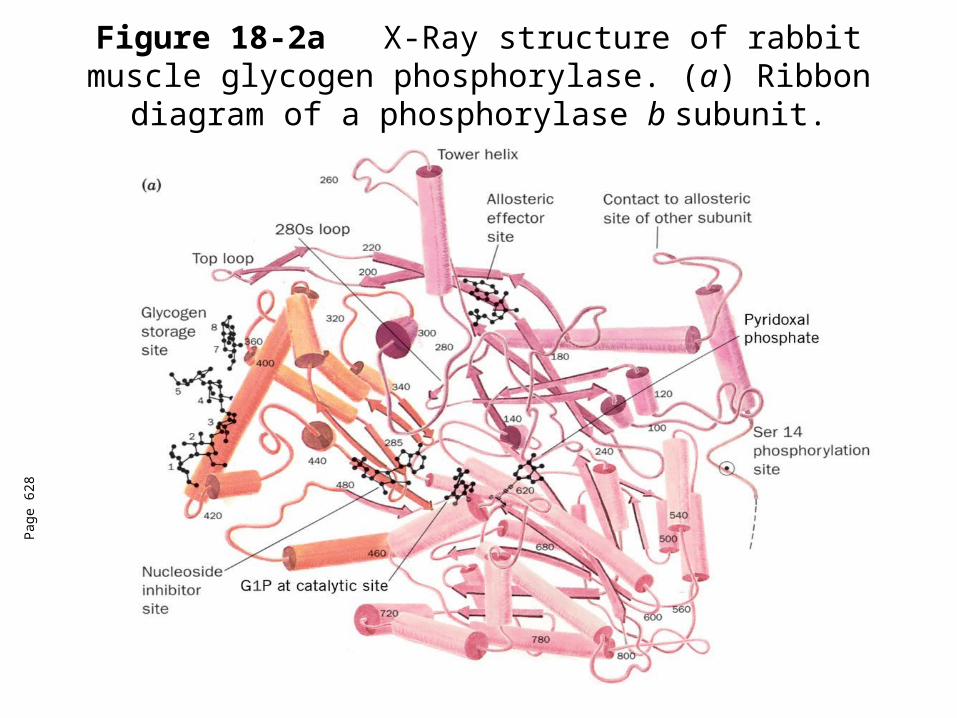
Figure 18-2aX-Ray structure of rabbit muscle glycogen phosphorylase. (a) Ribbon diagram of a phosphorylase
b subunit.
Pag
e 62
8
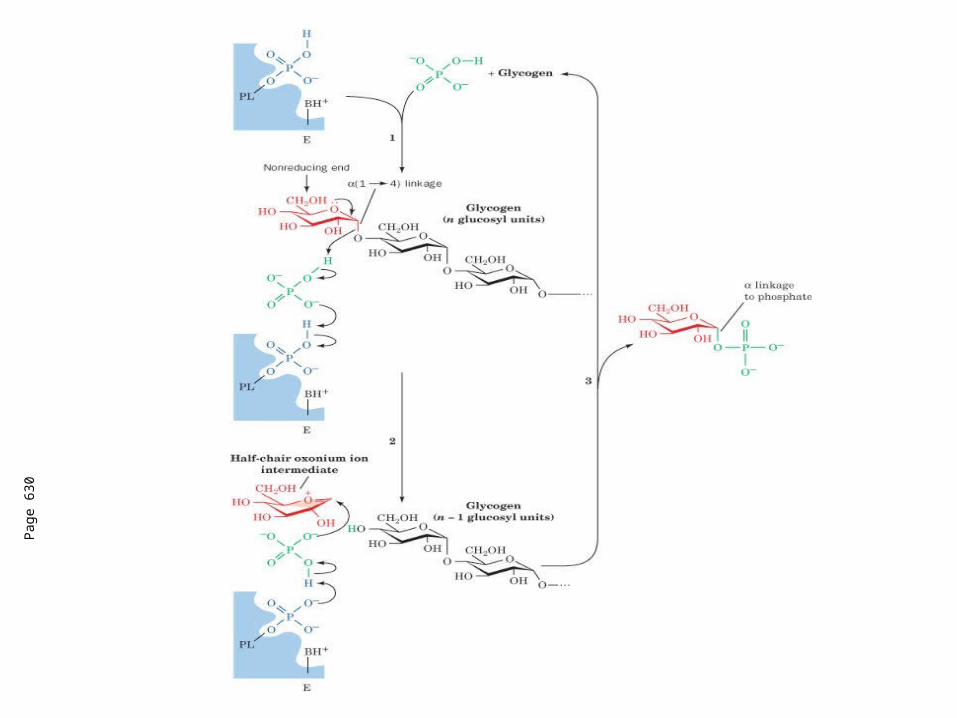
Pag
e 63
0

Phosphoglucomutase
Converts G1P to G6P.
Reaction is similar to that of phosphoglycerate mutase
Difference between phosphoglycerate mutase and phosphoglucomutase is the amino acid residue to which the phosphoryl group is attached.
Serine in phosphoglucomutase as opposed to His imidazole N in phosphoglycerate mutase.
G1,6P occasionally dissociates from the enzyme, so catalytic amounts are necessary for activity. This is supplied by the enzyme phosphoglucokinase.
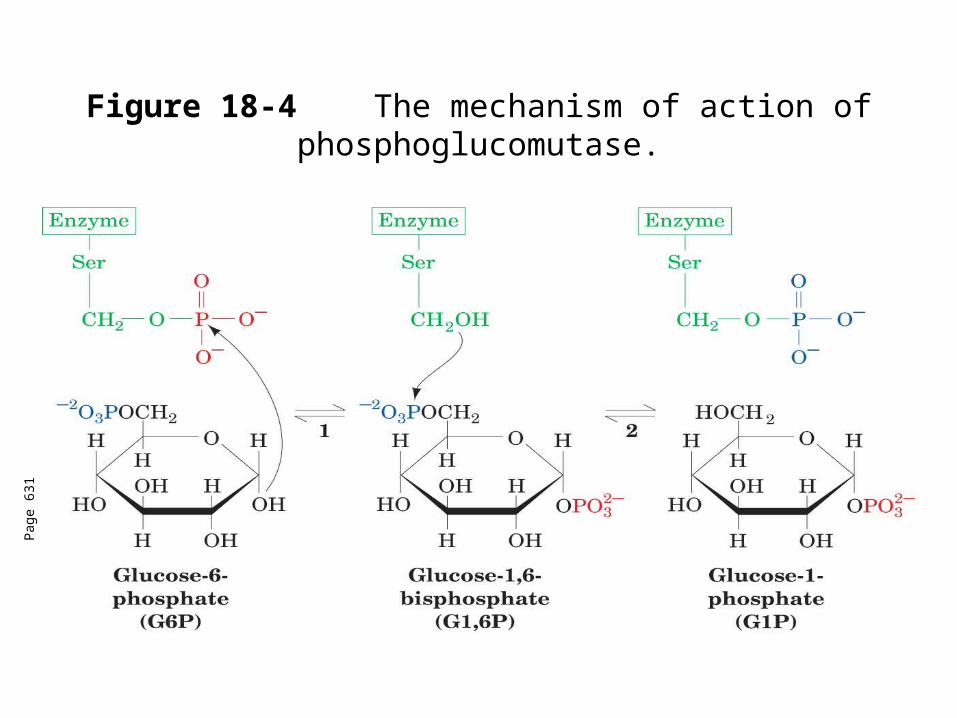
Figure 18-4 The mechanism of action of phosphoglucomutase.
Pag
e 63
1

Glycogen debranching enzyme
(14) transglycosylase (glycosyl transferase) transfers a (14) linked trisaccharide unit from a limit branch to a nonreducing end of another branch.
Forms a new (14) linkage with three more units available for phosphorylase.
The (16) bond linking the remaining linkage is hydrolyzed by the same enzyme to yield glucose.
2 active sites on the same enzyme.
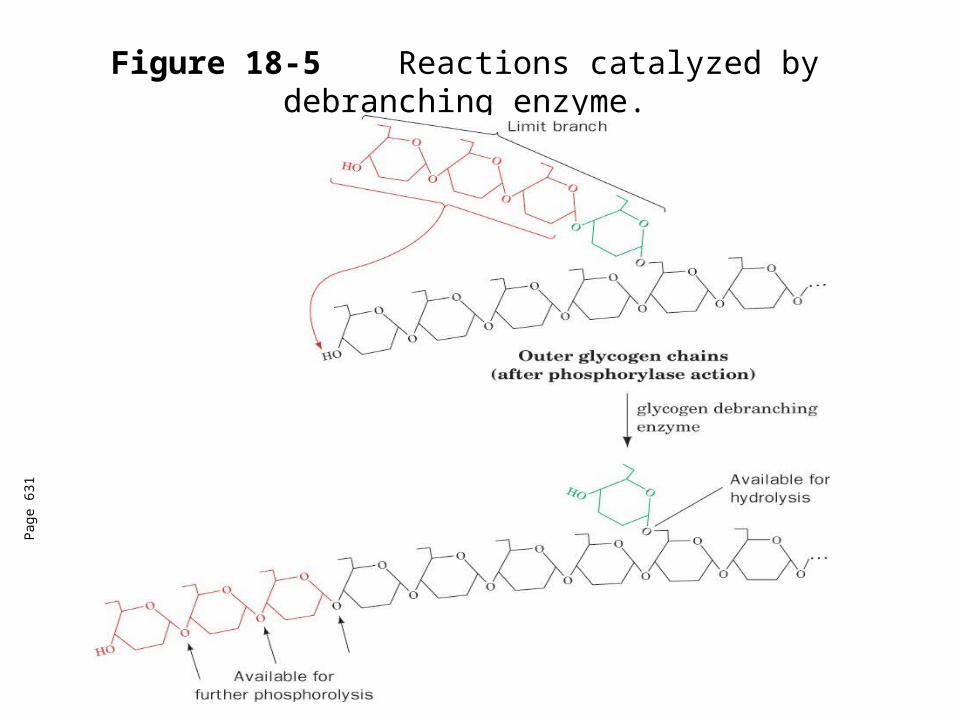
Figure 18-5 Reactions catalyzed by debranching enzyme.
Pag
e 63
1
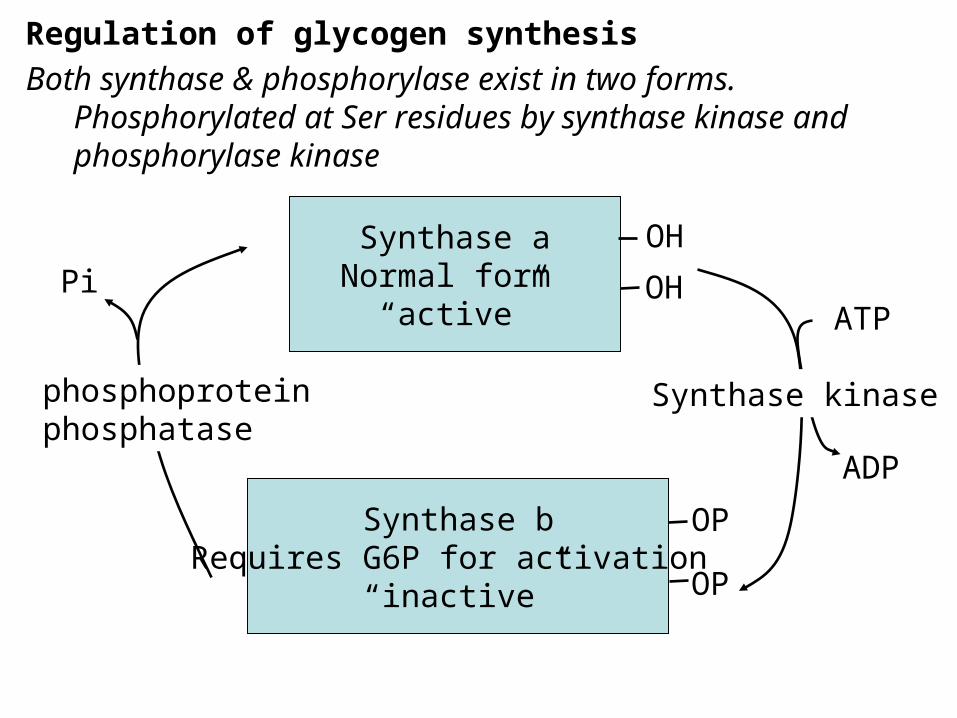
Regulation of glycogen synthesisBoth synthase & phosphorylase exist in two forms.
Phosphorylated at Ser residues by synthase kinase and phosphorylase kinase
Synthase aNormal form
“active”
Synthase bRequires G6P for activation
“inactive”
OH
OH
OP
OP
ATP
ADP
Pi
Synthase kinasephosphoproteinphosphatase
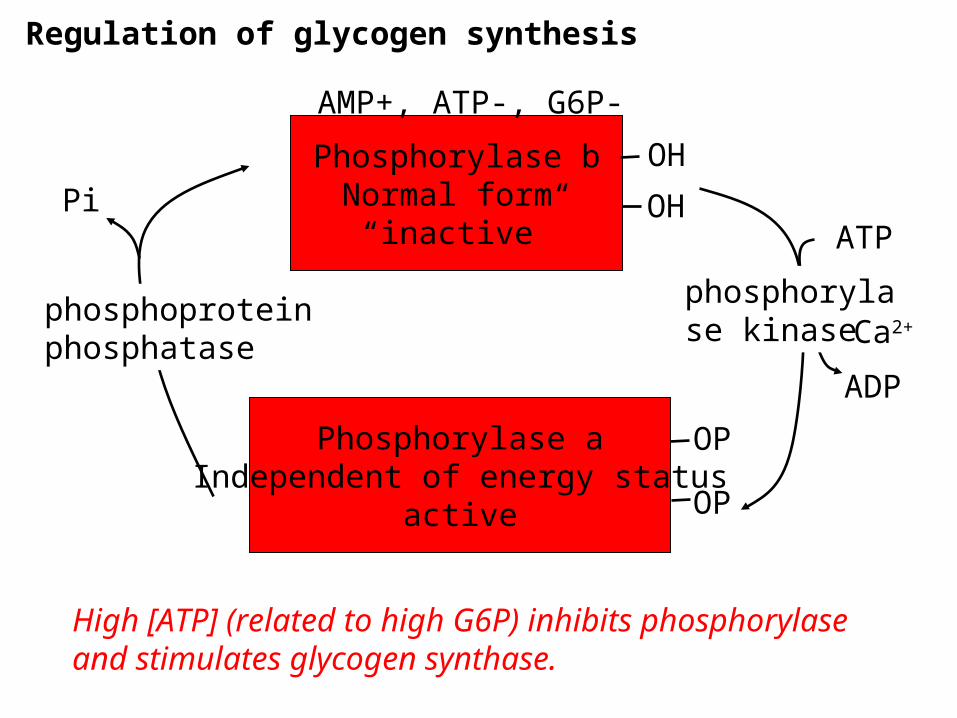
Regulation of glycogen synthesis
Phosphorylase bNormal form
“inactive”
Phosphorylase aIndependent of energy status
active
OH
OH
OP
OP
ATP
ADP
Pi
phosphorylase kinase
phosphoproteinphosphatase
AMP+, ATP-, G6P-
Ca2+
High [ATP] (related to high G6P) inhibits phosphorylase and stimulates glycogen synthase.

Regulation of glycogen synthesis
Process is also under hormonal controlAdrenaline (epinephrine) can regulate glycogen
synthesis/breakdown by stimulating adenylate cyclase
1. External stimulus Adrenaline Adenylate cyclase
ATP
cAMP + PPi
2. R2C2 [C]2 + [R-AMP]2cAMP dependent protein kinase
cAMP
“inactive” “active”
3a. Glycogen synthase a(active)
Glycogen synthase b (inactive)ATP ADP
[C]2
3b. Inactive phosphorylase kinase
Active phosphorylase kinaseATP ADP
[C]2
ATP ADP
Phosphorylase b(inactive)
Phosphorylase a (active)

Consider the whole systemResting muscle
Glycolytic pathway pyruvate ATP
cAMP
O2 respirationInactive phosphorylase b, active synthase aMuscle lacks G6 Pase, Liver PFK inhibited by ATP unless F2,6P2 present
Upon stressEpinephrine Synthase/phosphorylase kinase
Phosphorylse b Phosphorylse a
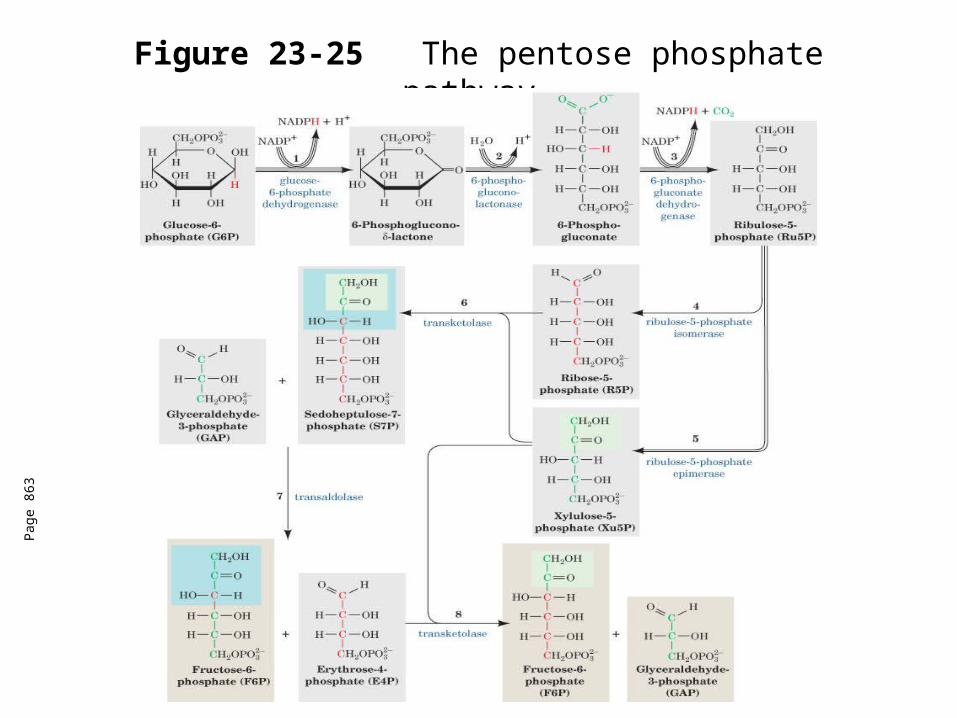
Figure 23-25The pentose phosphate pathway.
Pag
e 86
3