Embed Size (px)
Citation preview

438
Biochimica et Biophysica Acta, 425 (1976) 438- -450 © Elsevier Scient i f ic Publishing C o m p a n y - - Pr in ted in The Nether lands
BBA 98554
LIGHT-STIMULATED SYNTHESIS OF CHLOROPLAST DNA
LANA STOLARSKY, ALAN M. WALFIELD, RUTH ASCHAUER BIRCH * and CHARLES L. HERSHBERGER
Department of Microbiology, University of Illinois, Urbana, Ill. 61801 (U.S.A.)
(Received May 26th, 1975)
(Revised manuscript received November 27th, 1975)
Summary
Light-stimulated chloroplast DNA synthesis was studied during chloroplast development in the absence of cell division and nuclear DNA synthesis. Incor- poration of 32P i was stimulated 10--15 fold, however, the ratio of chloroplast DNA to nuclear DNA remained constant. Isotope dilution experiments suggest- ed that stimulated labeling of chloroplast DNA represented more efficient uti- lization of exogenously supplied Pi rather than stimulated turnover of chloro- plast DNA. The low level of DNA synthesis and chloroplast development were resistant to nalidixic acid which inhibits semiconservative replication of chloro- plast DNA.
Introduct ion
Euglena gracilis contains gratuitous chloroplasts that utilize light-energy to support phototrophic growth [1]. The cells use exogenous carbon sources for growth in the dark and chloroplasts degenerate into proplastids without chloro- phyll when cultures are grown in the dark. Light exposure of etiolated cultures induces development of proplastids into mature functional chloroplasts. Matu- ration is accompanied by distinct morphological changes, accumulation of chloroplast ribosomes, and synthesis of chloroplast-specific proteins and RNAs [ 2,3 ]. Investigations to determine the involvement of chloroplast DNA (ctDNA) replication in development provid e contradictory results. Uzzo and Lyman [4] reported that ctDNA increases 2--4-fold during light-regulated development, however, Stutz et al. [5] observed that the number of chloroplast ribosomal RNA cistrons was not altered by exposing dark-grown cells to light; and inhibi- tots of ctDNA replication do not measurably affect chloroplast maturation in
* Present address: Department of Microbiology, University of Illinois, College of Medicine, Chicago, I11., U.S.A.

439
non-dividing cells [6,7]. However, nalidixic acid bleaches and inhibits chloro- plast development in exponentially growing cultures [8,9]. Mielenz and Hersh- berger [10] postulated that unique segments of ctDNA are preferentially am- plified when light-grown cultures enter stationary phase; however, it was not known whether the differential synthesis of satellite DNAs accompanied chloro- plast maturation.
We have investigated synthesis of ctDNA by carefully measuring incorpora- tion of 32P i and proportions of ctDNA : nuclear DNA (nDNA). ctDNA is the only DNA species labeled in non-dividing etiolated cells in the dark or cells un- dergoing light-induced chloroplast development. The small amount of DNA synthesis does not represent net accumulation and ctDNA replication is not es- sential for chloroplast maturation in non-dividing cells.
Materials and Methods
(a) Organisms and growth. E. gracilis strain Z stock cultures were grown and transferred in the dark every week for 3 weeks in proteose peptone, tryptone, yeast extract broth and thereafter maintained in the same medium [11]. Dark- grown stock cultures were transferred at 3-week intervals for 1 year. All trans- fers of dark-adapted cultures were done in the presence of a blue gas flame and No. 7 green Kodak safelight filter. All tubes and flasks were covered with alu- minum foil and Scotch vinyl plastic electrical tape. An inoculum in proteose peptone, tryptone, yeast extract broth was allowed to grow at room tempera- ture for 1 week starting with a 1/100 dilution from the stationary-phase stock culture. Dark-grown experimental cultures were grown in Hutner's medium [12] at 26°C in a New Brunswick G27 incubator with gyrotory agitation at 200 cycles per min from an initial cell density of l0 s cells/ml to a final cell density of 1 • 106--3 • 106 cells/ml. Cell number was monitored with a Research Model B Electronic Particle Counter from Coulter Electronics Incorporated. Viability was monitored by diluting samples into 0.7% agar and spreading on proteose peptone, tryptone, yeast extract broth that was solidified with 1.5% agar [20]. The cells were concentrated in the dark by centrifuging in a clinical centrifuge for 1 min in sterilized nalgene tubes and resuspended into 0.1 vol. of maturation medium. The maturation medium was modified [13] and con- tained: 12.6/~g/1 MnC12 • 4 H20, 2.202 mg/1 CaC12 • 2 H20, 150 pg]l thiamine, 30 pg/1 vitamin B-12, 35.1 mg/l MgSO4.7 H20, 132 mg]l KH~PO4, 6.0 g/1 NH4C1, 650 mg/1 sodium citrate at pH 7.0. The cultures were shaken for 3 days in the dark, referred to as the dark-rest period.
Chloroplast maturation was effected by (a) changing the cells to fresh matu- ration medium or to maturation medium containing one-third the amount of phosphate as indicated in the figure legends, (b) exposing the cells to 500 ft candles of light, and (c) shaking at 26°C for 72 h. Chlorophyll was measured by the method of Arnon [14]. Control experiments were performed with an identical protocol except the cells remained in the dark.
(b)Isotope incorporation. For uptake studies 2--10 mCi of carrier-free 32P i (International Chemical and Nuclear Corp.) was added per 20 ml of matu- ration medium at the same time the cells were exposed to light. For isotope dilution experiments cultures of 200 ml were pulse labeled with 2 mCi of 32P i

440
for 32 h during exponential growth. The cells were washed with maturation medium before the dark-rest period and before exposure to light.
(c) 32p043- uptake into trichloroacetic acid-precipitable cell components. Cell aliquots of 10 pl were applied to Whatman 3 MM filter paper circles of 2.3 cm diameter or squares of 2.3 cm that were previously dipped into acetone. They were washed three times with cold 5% trichloroacetic acid (containing pyrophosphate at 10 mg/ml), two times with cold ethanol/ether (1 : 1, v/v), one time with 70% ethanol, one time with ether, and dried under an infrared heat lamp. Treated filters were suspended and counted in scintillation fluid containing 0.1 g of 2,2'op-phenylenebis(5-phenyloxazole) and 4.0 g of 2,5- diphenyloxazole per 1 of toluene. Total radioactivity in the medium was deter- mined by applying samples to filters without the subsequent wash procedure.
(d) DNA isolations for measuring 32P i in ctDNA. Samples of 2.0 ml were di- luted with 8.0 ml of cold acetone, washed once with 80% cold acetone and once with cold distilled water. The cells were suspended in 1.0 ml of lysis mix- ture containing: 0.1 M Tris • HC1 buffer at pH 8.0, 0.2 ml of pronase solution with 640 proteolytic units (defined by Calbiochem) of pronase per ml and 0.08 ml of 25% sodium dodecyl sulfate. NaC1 (0.12 ml of a 10% solution} and 2.0 ml of 99% ethanol were added after incubation for 3 h at 37°C. The precipitat- ed nucleic acids were collected by centrifugation at 13 500 X g for 10 min, washed twice with 95% ethanol and dissolved in 1.0 ml of buffer containing 0.015 M NaC1 and 0.0015 M sodium citrate at pH 7.0. RNA was hydrolyzed at 37°C for 30 min with 0.4 unit of ribonuclease T1 and 40 pg of ribonuclease A. After t reatment with 1.5 mg ofpronase for 30 min, the solution was mixed with 0.1 ml of 3.0 M sodium acetate, precipitated with 0.64 ml of isopropyl alcohol and DNA was wound onto a glass rod [15]. The DNA was dissolved in 0.5 ml of the above saline/citrate buffer and quanti tated by colorimetric assay with di- phenylamine [16].
0.2 ml of DNA samples were mixed with 6.57 ml of CsCl solution to a final isopycnic density of 1.706 g/cm 3 and containing 10 pg each of Micrococcus luteus DNA and Tetrahymena pyriformis DNA with buoyant densities of 1.731 [17] and 1.684 g/cm 3 [18], respectively. Samples were centrifuged in a Spinco type 65 rotor for 18 h at 54 000 rev./min and 20°C [19]. Gradients were frac- t ionated from the top with an Isco Model 640 density gradient fractionator and Model UA-2 analyzer. Fractions of 0.1 ml were collected on Whatman 3 MM filter paper discs. The filters from each gradient were washed as described in section c for liquid scintillation counting. Samples for additional characteriza- t ion of DNA were prepared in a similar manner except that 0.1-ml fractions were collected in test tubes and 10-pl aliquots were applied to Whatman 3 MM filters for radioactivity measurements.
(e) DNA specific activity measurements. Aliquots were added to acetone and washed as in the uptake experiments. The DNA was solubilized by suspending the cells in 2.2 ml of solution containing 0.1 M Tris • HC1 buffer at pH 8.0, 320 proteolytic units (as defined by Calbiochem) of pronase, and 0.5% sodium dodecyl sulfate then incubating for 1.5--3.0 h at 37°C. The sodium dodecyl sulfate and particulate mat ter were separated by adding 0.136 ml of cold satu- rated CsC1, incubating at 0.4°C for 30 min then centrifuging for 10 min at 13 500 X g. Samples of 1.0 ml from the supernatant were added to the neutral

441
CsC1 preparative gradients as described in section d except that Tetrahymena DNA was omit ted. The c tDNA fractions were combined, recentrifuged in a sec- ond CsC1 gradient, and the absorbance was recorded with an Isco UA-2 analyzer. The fractions were collected on Whatman 3 MM filters and analyzed as in sec- t ion d. Relative areas of DNA peaks were determined by weighing the peaks from the ultraviolet absorption profiles and specific activities were calculated as cpm per weight of the ultraviolet absorbance peak.
(f) Analytical ultracentrifugation. Analytical ultracentrifugation was per- formed in a Spinco Model E ultracentrifuge. Ratios of c tDNA : nDNA were calculated from results at 260 nm and 290 nm as previously described [20] .
Experimental Results
(a) ctDNA synthesis during chloroplast maturation The incorporation of 32p i into chloroplast DNA in E. gracilis was studied
during maturat ion of chloroplasts wi thout comcomitant synthesis of nDNA. The cells were grown in the dark, concentrated into a medium without a car- bon source for a dark-rest period of 3 days and exposed to light to induce plastid maturat ion to chloroplasts. Viability of cultures remained at 100 + 20% throughout the experiment, therefore cell death did not distort the results. Cell division sometimes continued during the first 12 h of the dark-rest period then cell number remained constant throughout the remainder of the experi- ment. Chloroplast development was monitored by measuring the amount of chlorophyll during the period of exposure to light. The chlorophyll level reach- ed a maximum plateau of 16 #g per 106 cells after 72 h in the light, indicating that chloroplasts completed maturat ion in 72 h, as reported for similar condi- tions [21] . Experimental results were independent of Pi concentrat ion after 24 h in the dark-rest period, however, variable amounts of chlorophyll were synthesized if the Pi concentrat ion was reduced before complet ion of the re- sidual division in the dark-rest period.
Nucleic acid synthesis during chloroplast maturat ion was examined in detail by comparing the incorporation of 32p i when the cells were exposed to light or dark. The radioactivity in the trichloroacetic acid-precipitable fraction was moni tored during the period of maturat ion as described in Materials and Meth- ods. The percent of the 32pi which became trichloroacetic acid precipitable in- creased both in the presence and iri the absence of light. However, the radio- activity in the light after 24 h was 2.5 times greater than in the absence of light (Fig. 1). This increase was thought mainly due to an increase in RNA synthesis because RNA synthesis is known to be stimulated during chloroplast matura- tion and RNA is the most abundant nucleic acid in the trichloroacetic acid precipitate [22] . The trichloroacetic acid-precipitable fraction may also con- tain polyphosphate; therefore, precludes direct assay of DNA on the filters.
Synthesis of DNA was measured in the dark and in the light. Samples were examined at times from 0 to 24 h after the end of the dark-rest period to mea- sure the radioactivity in DNA. Samples were centrifuged to equilibrium with M. luteus DNA and Tetrahymena DNA as standards in CsC1 density gradients. The two standards and the E. gracilis nDNA gave three peaks which were used as density markers to determine the buoyan t density of radioactive DNA spe-
442
(a) ~" '
~" ,6 ~ o
o (~ 14 7 k.
tO • 6 / "~ B 5
4
2 3 o o o
I I l I I I ~ I ~ I I I I I I0 2 0 30 4 0 50 6 0 2 4 6 8 I0 12 14 ~6 18 2 0 22 24 26 28
Time After Light Exposure (hr) Fraction Number
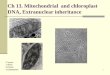
Fig. 1. Incorpozatton of 32p i. Cell suspensions at the end of the dark-rest period weze suspended in matu- rat ion m e d i u m containing 0 . 0 4 4 m g / m l K2HPO4 and 200 /~Ci/rul 32P i. Parallel cultures were exposed to light (o) ox maintained in the dask (I) . The fraction of the total 32p i incorporated into the cells was cal- culated as described in Materials and Methods. TCA, trich]oroacetic acid.
Fig. 2. Equil ibrium ultracentrifugation of 32P i DNA synthes ized in the dark and light. At the end of the dazk-rest period cells were suspended in maturat ion m e d i u m contatnt,~g 0 . 0 4 4 mg/rnl K2HPO 4 and 200 #Ci/ml 32p i. The suspensions were maintained in the dark (Frame A) or l ight (Frame B) and DNA was isolated and centrifuged to equil ibrium as described in Materials and Methods after 24 h in A and 20 h in B. Arrows m a r k the position~ of ultraviolet-absorbing peaks for n D N A with a buoyant density of 1.708 g /cm 3 and the reference DNAs: M. luteus, 1.731 g /cm 3 and T. pyr/form/s, 1.684 g /cm 3.
cies. The incorporated 32P i banded at the density of ctDNA in cultures kept in the dark and in cultures exposed to light (Fig. 2). Radioactive peaks in the gra- dients were identified as DNA because they were completely hydrolyzed by deoxyribonuclease. The radioactive DNA was identified by its density of 1.685 g/cm 3, which was identical to the density of ctDNA from E. gracilis. Nuclear DNA was not normally synthesized in these experiments; however, nDNA sometimes incorporated low levels of radioactivity after prolonged exposure t o 32P i in the light.
Incorporation of 32P i into ctDNA was calculated from gradients similar to Fig. 2. The data were calculated as percentage of radioactivity from the me- dium incorporated into ctDNA per #g of total isolated DNA. The specific ac- tivity measurements indicated that light stimulated the rate of incorporation of 32P i into ctDNA. The extent of stimulation was 10--15-fold and nominally equalled 12.5 for the graph of Fig. 3.
(b) Ratios o f ctDNA : nDNA to assay synthesis of ctDNA Incorporation of radioactive isotopes does not necessarily indicate net syn-
thesis of ctDNA. Steady-state turnover synthesis or repair synthesis would cause incorporation of radioactive phosphate. Therefore, relative quantities of
4 4 3
"~ .o2
.01
~ 8 5
B '~ B'
g
o o
• J ~ 5 h731 t,708
Time After Light Exposure {hr} Density
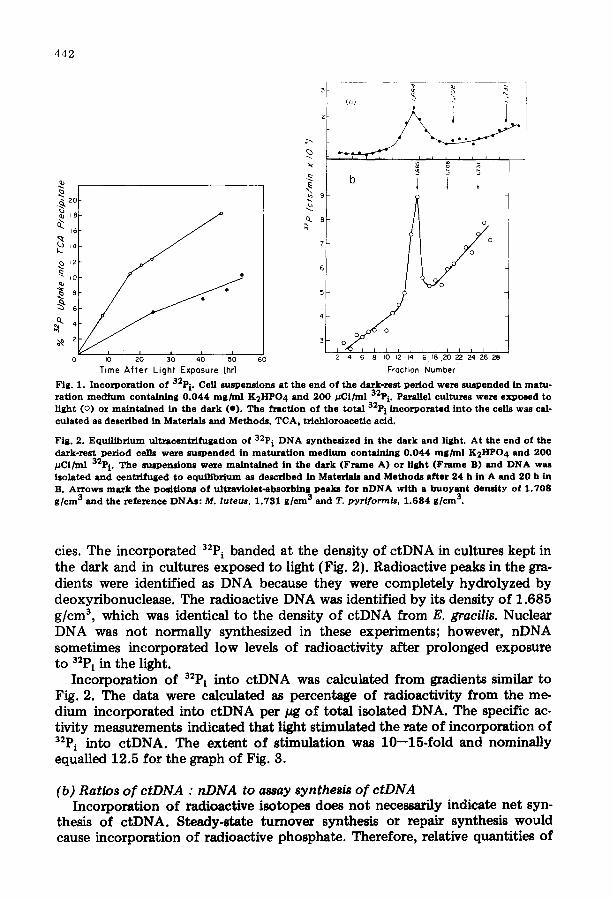
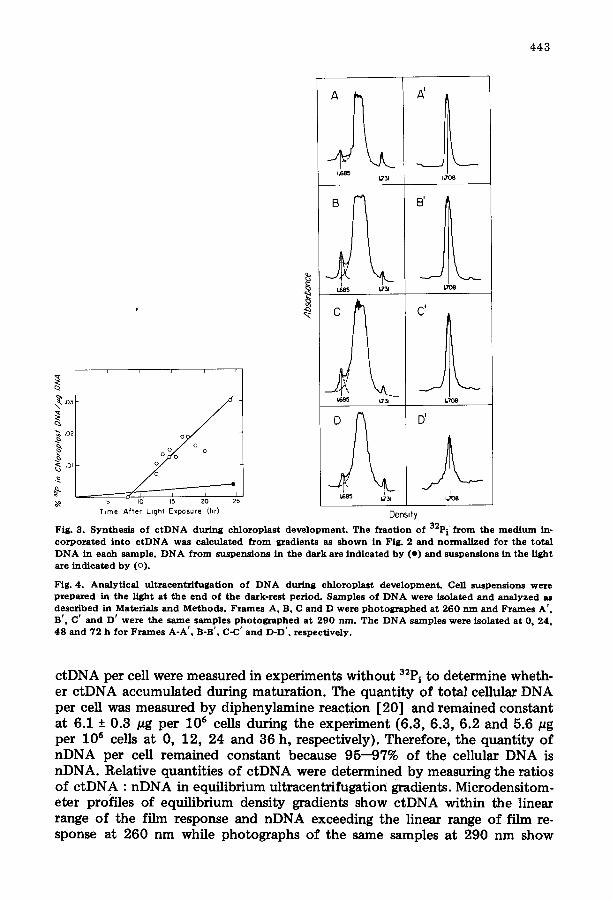
Fig. 3 . S y n t h e s i s o f c t D N A d u r i n g c h l o r o p l a s t d e v e l o p m e n t . T h e f r a c t i o n o f 32p i f r o m t h e m e d i u m in- c o r p o r a t e d i n t o c t D N A w a s c a l c u l a t e d f r o m g r a d i e n t s as s h o w n in Fig . 2 a n d n o r m a l i z e d f o r t h e t o t a l D N A in e a c h s a m p l e . D N A f r o m s u s p e n s i o n s in t h e d a r k a re i n d i c a t e d b y ( e ) a n d s u s p e n s i o n s in t h e l i gh t a re i n d i c a t e d b y (o) .
F ig . 4 . A n a l y t i c a l u l t r a e e n t r i f u g a t i o n o f D N A d u r i n g c h l o r o p l a s t d e v e l o p m e n t . Cell s u s p e n s i o n s w e r e p r e p a r e d in t h e l i g h t a t t h e e n d o f t h e d a r k - r e s t p e r i o d . S a m p l e s o f D N A w e r e i s o l a t e d a n d a n a l y z e d as d e s c r i b e d in Mate r i a l s a n d M e t h o d s . F r a m e s A, B, C a n d D w e r e p h o t o g r a p h e d a t 2 6 0 n m a n d F r a m e s A ' , B ' , C ' a n d D ' w e r e t h e s a m e s a m p l e s p h o t o g r a p h e d a t 2 9 0 n m . T h e D N A s a m p l e s w e r e i s o l a t e d a t 0, 24 , 4 8 a n d 7 2 h f o r F r a m e s A - A ' , B-B' , C-C' a n d D-D ' , r e s p e c t i v e l y .
ctDNA per cell were measured in experiments wi thout 32P i t o determine wheth- er c tDNA accumulated during maturation. The quant i ty of total cellular DNA per cell was measured by diphenylamine reaction [20] and remained constant at 6.1 + 0.3 pg per 106 cells during the experiment (6.3, 6.3, 6.2 and 5.6/~g per l 0 s cells at 0, 12, 24 and 36 h, respectively}. Therefore, the quant i ty of nDNA per cell remained constant because 95--97% of the cellular DNA is nDNA. Relative quantities of c tDNA were determined by measuring the ratios of c tDNA : nDNA in equilibrium ultmcentrifugation gradients. Microdensitom- eter profiles of equilibrium density gradients show c tDNA within the linear range of the film response and nDNA exceeding the linear range of film re- sponse at 260 nm while photographs of the same samples at 290 nm show
4 4 4
nDNA within the linear range of the film response (Fig. 4). The ratio of c tDNA : nDNA was calculated from these profiles and remains constant at 2.8% with a maximum deviation of 0.7% throughout chloroplast maturation (2.1 _+ 0.6, 3.2 + 0.1, 3.4 + 0.6 and 2.5 -+ 0.8% at 0, 24, 48 and 72 h, respective- ly).
(c) Isotope dilution experiments to assay synthesis of ctDNA The light-stimulated c tDNA synthesis was further investigated by isotope
dilution experiments to determine the amount of synthesis. The procedure was to measure the specific activity of the c tDNA after light exposure in cells which had been previously labeled with 3:P i. If the synthesis represented a large in- crease of c tDNA or a large amount of turnover synthesis, the specific activity of the c tDNA after light exposure would decrease because the new synthesis would incorporate non-radioactive phosphate. However, if the synthesis rep- resented a small fraction of the total ctDNA, the specific activity of the c tDNA would remain constant because not enough non-radioactive phosphate would be incorporated to cause a detectible change in the specific activity.
3.0
2.5
2.0
1.5
1.0
~' 0.5
~ o
.~ 0.6
0.5
0.4
0.3
0.1
C
'~ 30
m t~
.a ,c
~ ~ 0 hr$ /
l ~ ~ o.~ I 0.6
O.4
0.2
0 , J I 12 14 16 18 20 22 24 26 28 30 32 34 36 4 6 8
_'9. 5 • e b
C g ~ ~ o h~s
4 6 8 I0 I? 14 16 18 20 22 ?4 26
~ ~ 24 hr$ ~. ~.
I0 12 14 16 18 20 22 ;)4 26 28 30
~ ~ 24 hfs ~. ~_.
8 I0 12 14 16 18 20 22 24 26 28 30 32 34 36
Fract ion N u m b e r
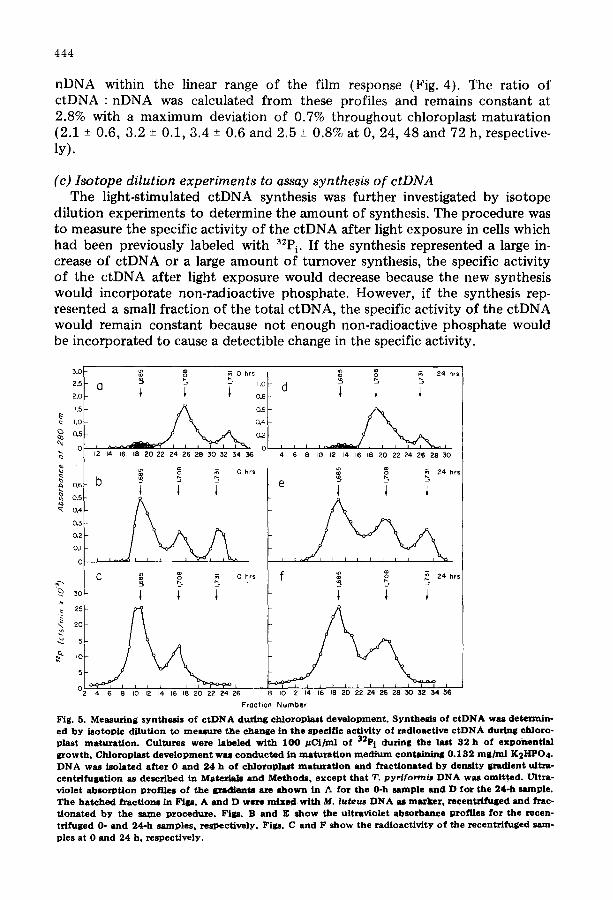
Fig. 5. Measuring s y n t h e ~ s o f c t D N A during ch loroplas t deve lopment . Synthe l i s o f c t D N A was determin- ed b y i so top ic di lut ion to meazure the change in the specif ic act ivity o f radioact ive c t D N A during chloro- plast maturat ion . Cultures were labeled wi th 1 0 0 # C i / m l of 32P i during the last 32 h o f exponent ia l growth. Chloroplast d e v e l o p m e n t was c o n d u c t e d in maturat ion m e d i u m conta in ing 0 . 1 3 2 m g / m l K2I-1PO 4. D N A waS i so lated after 0 and 24 h o f ch loroplas t maturat ion and fTactionated b y dens i ty gradient ulta~a - centr i fugat ion as described in Materials and Methods , e x c e p t that T. pyri for~is D N A was omit ted . Lrltra- v io let absorpt ion prof i les o f the gradiente are s h o w n in ~. for the 0-h sample and D for the 24-h sample. The hatched fraction8 in F i p . A and D were m i x e d with M. luteus D N A as m~rker, recentr l fuged and ~zac- t ionated b y the same procedure . Figs. B and E s h o w the ultraviolet ah lorbance prof i les for the recen- trifuged 0- and 24-h samples , respect ive ly . Figs. C and F s h o w the radioact iv i ty o f the recentri~uged sam- ples at 0 and 2 4 h , respect ively .
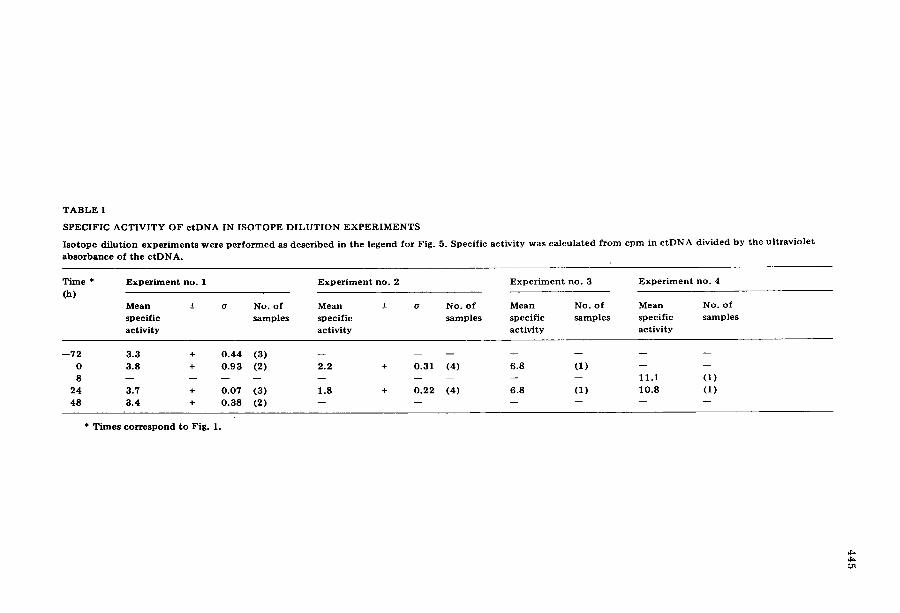
T A B L E I
S P E C I F I C A C T I V I T Y O F c t D N A IN I S O T O P E D I L U T I O N E X P E R I M E N T S
I s o t o p e d i l u t i o n e x p e r i m e n t s w e r e p e r f o r m e d as d e s c r i b e d i n t h e l e g e n d f o r F ig . 5. S p e c i f i c a c t i v i t y w a s c a l c u l a t e d f r o m c p m i n c t D N A d i v i d e d b y t h e u l t r a v i o l e t
a b s o r b a n c e o f t h e c t D N A .
T i m e * E x p e r i m e n t no . 1 E x p e r i m e n t n o . 2 E x p e r i m e n t no . 3 E x p e r i m e n t no . 4
(h) M e a n +- o No . o f M e a n + o No . o f M e a n No. o f M e a n No. o f
s p e c i f i c s a m p l e s s p e c i f i c s a m p l e s s p e c i f i c s a m p l e s s p e c i f i c s a m p l e s
a c t i v i t y a c t i v i t y a c t i v i t y a c t i v i t y
- - 7 2 3 .3 + 0 . 4 4 (3 ) . . . . . . .
0 3 .8 + 0 . 9 3 (2) 2 . 2 + 0 . 3 1 (4) 6 .8 (1 ) - - - - 8 . . . . . . . . . I i . i (1 )
2 4 3.7 + 0 . 0 7 (3 ) 1 .8 + 0 . 2 2 (4 ) 6 .8 (1 ) 1 0 . 8 (1 )
48 3 .4 + 0 . 3 8 (2) . . . . . .
* T i m e s c o r r e s p o n d t o F ig . 1.

446
To incorporate radioactive label into ctDNA in the dark, the cells were pre- labeled with 32P i for 32 h during exponential growth in Hutner 's medium and washed to remove 32P i before the dark-rest period. The total 32P i was the same as used for labeling during maturation, however, the radioactivity per ml was less because the incorporation was completed during exponential growth be- fore concentrat ion of the cells into maturat ion medium. Samples were taken before and after light exposure and the DNA was centrifuged to equilibrium in CsC1 density gradients. The pooled c tDNA fractions were recentrifuged in a second CsC1 density gradient and fractionated onto Whatman (3 MM) paper filters to measure radioactivity. The specific activity was defined as the radio- activity per integrated ultraviolet absorbance area of ctDNA. Typical profiles of gradients are shown in Fig. 5. Specific activities were calculated from similar profiles for several maturat ion times as shown in Table I. The mean specific ac- tivity of ctDNA remained constant (within one standard deviation) between the start of the dark-rest period and 48 h of light exposure.
(d) Effect o f nalidixic acid on light-stimulated incorporation o f 32P i into ctDNA
Semiconservative replication of c tDNA is inhibited by nalidixic acid [23] . Therefore, light-stimulated incorporation of 32P i into c tDNA should be inhibit- ed by nalidixic acid if the synthesis represents normal semiconservative replica- tion. The effect of nalidixic acid was examined with the standard experimental protocol for maturat ion except that the culture was divided in half at the time of exposure to light and nalidixic acid was added to one portion. Maturation medium was adjusted to pH 3.8 for these experiments to match the pH opti- mum for nalidixic acid inhibition of c tDNA replication [23] .
Results at the lower pH were qualitatively the same as previous maturat ion experiments except the chlorophyll levels were reduced approx. 75%. Lower levels of chlorophyll were independent of nalidixic acid or 32P i and seemed to result from the lower pH of the medium. Specific activities of c tDNA were de- termined from equilibrium density gradients as described above.
The mean specific activity for five determinations on the culture wi thout nalidixic acid was 14.02 with a standard deviation of +3.38. The mean specific activity for five determinations on the culture with nalidixic acid was 9.98 with a standard deviation of +2.53. Therefore, light-stimulated incorporation of 3~P i into c tDNA is inhibited only slightly if at all by nalidixic acid.
Discussion
Proplastids and chloroplasts of non-dividing E. gracilis incorporate exogenous 32P i into ctDNA. The radioactive isotope incorporation is stimulated 10--15- fold by light; however, light-stimulated labeling appears to represent more ef- ficient utilization of exogenous 32P l probably by deplet ion of endogenous pools for synthesis of RNA and other products, rather than stimulated synthesis o f ctDNA. The experimental technique would have detected synthesis exceeding 15% replication; therefore, less than 15% of the ctDNA replicates during light~ regulated chloroplast development.

447
Small amounts of net synthes~s would not be detected if less than 15% of the ctDNA were replicated. More precise measurements are not possible by current- ly available techniques. Analytical ultracentrifugation is limited by the accura- cy of measuring the ratio of a small DNA component to a much larger quantity of DNA in the major band, which represents +25% of the minor DNA species. Similarly, the precision of specific activity calculations is limited by the accura- cy of measuring the DNA content of bands in the preparative equilibrium den- sity gradients.
High levels of radioactive isotope were employed to obtain easily measured incorporation into DNA. However, the results were not affected by radiation poisoning of the cells. E. gracilis exhibits tolerance to high levels of radioac- tive isotopes. Cell growth, viability and synthesis of chlorophyll were not al- tered by 32Pi in these studies. In typical experiments the chlorophyll levels after 24 h of maturation were 93.2% of the non-radioactive control when 0.1 mCi of 32pi was present during maturation and 93.5% of the non-radioactive control when 10.5 mCi of 32p~ was present during maturation. Viabilities were 100% + 20% in the same experiments. In a typical experiment the chlorophyll level after 24 h of maturation was 130% of the non-radioactive control for experiment No. 1 of Table I, when 32P i was incorporated during exponential growth. Viability in the experiment was not distinguishable from experiments without 3~P i. Analysis of ctDNA by analytical ultracentrifugation was perform- ed in experiments without the radioactive isotope and confirmed the interpre- tations derived from measuring incorporation of 32P i.
Conclusions from this study differ from the results of Uzzo and Lyman [4], who reported that ctDNA increased 2--4-fold during chloroplast maturation. These observations may reflect experimental conditions because the previous investigation employed maturation medium with a different chemical composi- tion, a different light intensity, and a 10-fold lower number of cells/ml. Alter- nately the difference between observations may result from the assays for ct- DNA. The previous study quantitated ctDNA by measuring the ratio of ctDNA to a satellite DNA species with a buoyant density of 1.691 g/cm 3 and assumed that the quantity of satellite DNA remained constant. Species of DNA with similar buoyant densities have been reported as mitochondrial DNA [24], fragments of ctDNA [25] and covalently closed circular DNA of unknown origin in E. gracilis [10]. It should not be assumed that the amount of satellite DNA remains constant during chloroplast development because the quantity of DNA with a buoyant density of 1.691 g/cm 3 changes in different stages of cul- ture growth [20]. Our studies measured the ratio of ctDNA : nDNA and the specific activity of ctDNA in isotope dilution experiments, therefore, the mea- surements did not depend on assumptions about other species of satellite DNA.
The ratios of ctDNA : nDNA in this study were approximately one-half the ratios observed during balanced exponential growth of cultures growing in the light [20]. Ratios of ctDNA : nDNA were approx. 5% in exponential cultures growing in the dark {unpublished observation). Therefore, the relative amount of ctDNA should double during the transition from chloroplast maturation to exponential growth. Ratios of ctDNA : nDNA approximately double during the transition from exponential growth to stationary phase of cultures growing in the light [20]. Therefore, stationary phase cultures contain about 4--5-fold

448
higher proportions of ctDNA than observed during our maturation experi- ments. These results indicate that ctDNA synthesis during maturation must be measured under carefully controlled conditions to avoid coincidence of changes associated with transitions between different phases of growth.
Anaerobically grown yeast exhibit stimulated synthesis of mitochondrial DNA when mitochondrial development is induced by exposure to oxygen. The mitochondrial DNA synthesis represents 10--30% net replication as measured by isotope dilution experiments [26]. Inhibition of mitochondrial develop- ment by nalidixic acid suggests that the observed replication is essential for nor- mal development.
Results in this study do not distinguish whether the observed ctDNA synthe- sis is required for chloroplast development. Nalidixic acid does not inhibit chlo- roplast development or light-stimulated incorporation of 32Pi into ctDNA. Re- sistance to nalidixic acid suggests that the observed labeling does not represent normal ctDNA replication because semiconservative replication of ctDNA in exponentially growing cultures is inhibited by nalidixic acid [23]. This inter- pretation must be qualified because the observation could be explained if nali- dixic fails to reach its site of action in developing cells suspended in maturation medium. However, the medium employed for testing nalidixic acid in exponen- tially growing cultures contains the same chemical components as maturation medium with glucose added as a carbon source. Nalidixic acid requires cell division to induce bleaching of E. gracilis [6]. However, it is not known wheth- er bleaching requires a process of cell growth or semiconservative replication of ctDNA. Inhibition of bacterial replication by nalidixic acid causes degradation of the partially replicated DNA [27]. Therefore, the resistance of non-dividing E. gracilis to bleaching could reflect the absence of semiconservative ctDNA replication.
The observed 32P i incorporation apparently represents low levels of turnover of ctDNA, and may involve replacement of short segments, long segments or whole molecules. Turnover of ctDNA has been observed in exponentially grow- ing cultures of E. gracilis (ref. 28 and unpublished observations) and represents a frequently encountered prope~y of organelle DNA [29--31]. The synthesis may be a type of repair synthesis; however, the repair must differ from excision repair of thymine dimers caused by ultraviolet irradiation because E. gracilis does not exhibit dark repair of bleaching induced by ultraviolet light [32]. Re- pair replication of ultraviolet-induced lesions is absent in chloroplasts of Chlamydomonas reinhardi [33]. Turnover of ctDNA also may represent abor- tive attempts to initiate normal replication. A specific small segment of mito- chondrial DNA turns-over in mouse L-cells to maintain the DNA in a prerep- licative form for initiation of replication [34,35].
Mielenz and Hershberger [10] postulated that unique segments of ctDNA with buoyant densities of 1.691 and 1.700 g/cm 3 are amplified preferentially at different growth stages of E. gracilis. Preferential synthesis of the unique satellites is not observed in this study during the last 48 h of the dark-rest pe- riod or during light-induced chloroplast development. Therefore, transition from exponential growth to stationary phase in heterotrophic light-grown cul- tures remains the only reported occurrence for preferential synthesis of satel- lites with buoyant densities of 1.691 and 1.700 g/cm 3.

449
Acknowledgments
Appreciation is extended to E. Stutz for providing a copy of ref. 5 before publication, and to H. Lyman for helpful discussion and sharing relevant un- published data. Alan Walfield was a predoctoral trainee supported by U.S. Pub- lic Health Service Training Grant GM 00510. Research expenses were paid by National Science Foundation Grant GB 35598 and the University of Illinois Research Board.
Addendum
After our manuscript was submitted for publication Mahler et al. (H.R. Mahler, K. Assimos and C.C. Lin (1975) J. Bacteriol. 123,637--641) indepen- dently reinvestigated the requirement of DNA synthesis f~r mitochondrial maturation. They demonstrated that DNA synthesis was not required for devel- opment. Therefore, net synthesis of DNA is not necessary for development of chloroplast or mitochondria and differential gene expression must be the major factor regulating organelle development.
References
1 Schiff, J.A. and Epstein, H.T. (1968) The Biology of Euglena (Buetow, D.E., ed.), Vol. II, pp. 285-- 333, Academic Press, New York
2 Schiff, J.A. (1973) Advances in Morphogenesis (Abererombie, M., Braehet, J. and King, T.J., eds.), Vo]. X, pp. 265---312, Academic Press, New York
3 Reger, B.J., Smillie, R.M. and Fuller, R.C. (1972) Plant Physiol. 50, 24--27 4 Uzzo, A. and Lyman, H. (1972) Second International Congress on Photosynthesis (Forti, G., Avron,
M. and Melandri, A., eds.), Vol. III, pp. 2585--2599, W. Junk, The Hague 5 Stutz, E., Crouse, E.J. and Graf, L. (1975) International Colloquium on Cell Cycles, in the press,
Yvettes 6 Lyman, H. (1967) J. Cell Biol. 35, 726--730 7 Diamond, J. and Schiff, J.A. (1970) Can. J. Bot. 48, 1277--1283 8 Parthier, B., Krauspe, R. and Samlteben, S. (1972) Biochim. Biophys. Acta 277, 335--341 9 Neuman, D. and Parthier, B. (1973) Exp. Cell Res. 81, 255--268
10 Mielenz, J.R. and Hershberger, C.L. (1974) Biochem. Biophys, Res. Commun. 58, 769--777 11 Mego, J.L. (1964) Biochim. Biophys. Acta 79, 221--225 12 Hutner, S.H., Zahalsky, A.C., Aaronson, S., Baker, H. and Frank, O. (1966) Methods in Cell Physiolo-
gy (Prescott, D.M., ed.), Vol. II, pp. 217--228, Academic Press, New York 13 Kempner, E.S. and Miller, J.H. (1972) J. ProtozooL 19, 343--346 14 Arnon, D.I. (1949) Plant Physiol. 24, 1--15 15 Marmur, J. (1961) J. Mol. Biol. 3, 208--218 16 Burton, D. (1956) Biochem. J. 62, 315--323 17 Schildkraut, C.L., Marmur, J. and Dory, P. (1962) J. Mol. Biol. 4,430---443 18 Sueoka, N. (1961) Cold Spring Harbour Symp. Quant. Biol. 26, 35---43 19 Flamm, W.G., Bond, H.E. and Burr, H.E. (1966) Biochim. Biophys. Acta 129, 310--317 20 Gibson, W.H. and Hershberger, C.L. (1975) Arch. Biochem. Biophys. 168, 8--14 21 Stem, A.I., Schiff, J.A. and Epstein, H.T. (1964) Plant Physiol. 39, 220--226 22 Gnanam, A. and Kahn, J.S. (1967) Biochim. Biophys. Aeta 142, 475--485 23 Pienkos, P., Walfleld, A. and Hershberger, C. (1974) Arch. Biochem. Biophys. 165, 548--553 24 Nass, M.M.K., Schori, L., Ben-Shaul, Y. and Edelman, M. (1974) Biochim. Biophys. Aeta 374, 283--
291 25 Vandrey, J.P. and Stutz, E. (1973) FEBS Lett. 37, 174--177 26 Rabinowitz, H., Getz, G.S., Casey, J. and Swift, H. (1969) J. Mol. Biol. 41, 381---400 27 Cook, T. M., Brown, K.G., Boyle, J.V. and Goss, W.A. (1966) J. BacterioL 92, 1510--1514 28 Manning, J.E. and Richards, O.C. (1972) Biochemistry 11, 2036--2043 29 Ch'ih, J.J. and Devlin, T.M. (1973) Arch. Biochem. Biophys. 156, 26--33 30 Richards, O.C. and Ryan, R.S. (1974) J. MoL Biol. 82, 57--75

450
31 Gross, N.J., Getz, G.S. and Rabinowitz, M. (1969) J. Biol. Chem. 244, 1552--1562 32 Schiff, J.A., Lyman, H. and Epstein, H.T. (1961) Biochim. Biophys. Acta 50, 310--316 33 Swinton, D.C. and Hanawalt, P.C. (1973) Biochim. Biophys. Acta 294. 385--395 34 Berk0 A.J. and Clayton, D.A. (1974) J. Mol. Biol. 66,801--824 35 Robberson, D.L, and Clayton, D.A. (1973) J. Biol. Chem. 248, 4512--4514