Embed Size (px)
Citation preview

*Neuroscience Research, Abbott GmbH and Company KG, Ludwigshafen, Germany
�Department of Neurobiology, Graduate School of Medicine, Chiba University, Chuo-ku, Chiba, Japan
�Department of Molecular Neuroscience, Graduate School of Medicine, Osaka University, Suita, Osaka, Japan
Axons in the adult central nervous system exhibit almost noregeneration after injury. This lack of regeneration is becauseof the presence of neurite outgrowth inhibitors, such asNOGO A, myelin-associated glycoprotein, oligodendrocyte-myelin glyco-protein, chondroitin sulfate proteoglycans andproteins of the repulsive guidance molecule family (RGM)(David and Lacroix 2003; Huber et al. 2003; Mueller et al.2006).
Repulsive guidance molecule was originally described as a33 kDa tectum repellent protein in chicken (Drescher et al.1997; Monnier et al. 2002). In humans, three homologousproteins termed RGM A, RGM B, and RGM C have beenreported. Human RGM A is highly homologous to chickenRGM A (with 82% identity). Up-regulation of RGM A wasobserved in humans suffering from traumatic brain injuryand focal ischemia and in injured or damaged regions of thespinal cord in rats (Schwab et al. 2005a,b; Hata et al. 2006).Blocking of RGM Awith a neutralizing polyclonal antibodyenhances the functional recovery in rats with induced spinalcord injury, demonstrating thatRGMAacts as a neurite growthinhibitor in the adult rat spinal cord (Hata et al. 2006).
Neogenin, a deleted in colorectal cancer (DCC) homo-logue, has been identified as the RGM A receptor
(Rajagopalan et al. 2004). DCC and Neogenin have anidentical domain structure and share 50% amino acididentity. At their cytoplasmatic region both proteins sharethree short, conserved domains (called P1, P2, and P3) withhigh amino acid identity (up to 97%). Beside thesedomains the cytoplasmic part of Neogenin shows nosimilarity with any other known protein (Vielmetter et al.1994; Meyerhardt et al. 1997). RGM A binding toNeogenin activates a protein kinase C, RhoA – Rho kinasepathway (Hata et al. 2006; Conrad et al. 2007). However,
Received February 21, 2008; revised manuscript received May 30, 2008;accepted July 29, 2008.Address correspondence and reprint requests to Bernhard K. Mueller,
Neuroscience Research, Abbott GmbH and Company KG, Knollstrasse50, D-67061 Ludwigshafen, Germany.E-mail: [email protected] used: AD, activation domain; BD, binding domain;
CMV, cytomegalovirus; DCC, deleted in colorectal cancer; DMEM,Dulbecco’s modified Eagle’s medium; FAK, focal adhesion kinase; FBS,fetal bovine serum; HA, hemagglutinin; LMO4, LIM-only protein 4;PBS, phosphate-buffered saline; PITPa, phosphatidylinositol transferprotein a; RA, retinoic acid; RGM, repulsive guidance molecule; siNT,non-targeted control siRNA; siRNA, small interfering RNA.
Abstract
Repulsive guidance molecule A (RGM A) was recently
described as a potent inhibitor of neuroregeneration in a rat
spinal cord injury model. The receptor mediating RGM A’s
repulsive activity was shown to be Neogenin, a member of the
Deleted in Colorectal Cancer (DCC) family of netrin receptors.
Binding of RGM A to Neogenin induces activation of the small
GTPase RhoA and of its effector Rho-kinase by an unknown
mechanism. Here we show, that the cytoplasmic tail of Neo-
genin interacts directly with the transcriptional coactivator LIM
domain only 4 (LMO4) in human SH-SY5Y cells, human Ntera
neurons, and in embryonic rat cortical neurons. RGM A binding
to Neogenin but not binding of Netrin-1, induces release of
LMO4 from Neogenin. Down-regulation of LMO4 neutralizes
the repulsive activity of RGM A in neuronal cell lines and
embryonic rat cortical neurons and prevents RhoA activation.
These results show for the first time that an interaction of
Neogenin with LMO4 is involved in the RGM A – Neogenin
signal transduction pathway for RhoA activation.
Keywords: axon guidance, LIM-only protein 4, neuroregen-
eration, repulsive guidance molecule A, RhoA/Rock signaling.
J. Neurochem. (2008) 107, 418–431.
JOURNAL OF NEUROCHEMISTRY | 2008 | 107 | 418–431 doi: 10.1111/j.1471-4159.2008.05621.x
418 Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431� 2008 The Authors

the detailed activation mechanism of the signaling pathwaybetween Neogenin and RhoA resulting in the inhibitory orrepulsive activity on growing neurites has not yet beenelucidated.
Using a yeast-two-hybrid approach we identified LMO4 asa protein directly interacting with Neogenin. LMO4 mainlyconsist of two zinc-finger LIM domains, which are thought tobe involved in protein-protein interactions and in general actas adaptors for the assembly of multi-protein complexes(Novotny-Diermayr et al. 2005; Lu et al. 2006). LMO4 hasbeen shown earlier to play an important role during nervoussystem development (Chen et al. 2002). A conditionaldeletion of LMO4 in postnatal cortical neurons causesaberrant segregation of thalamocortical afferents and abnor-mal development of the somatosensory barrel cortex(Kashani et al. 2006). Approximately 50% of LMO4knock-out mice die at birth with a defect in cranial neuronaltube closure (Lee et al. 2005). Interestingly, a defect incranial neural tube closure is also observed in approximately50% of homozygous RGM A knock out mice (Niederkofleret al. 2004).
Here we provide evidence that LMO4 directly interacts withcytoplasmic domains of Neogenin and plays an important rolein repulsive axon guidance along with RGM A.
Materials and methods
Reagents and materialsAntibodies against human Neogenin were purchased from Santa
Cruz Biotechnology (H175, C20; Heidelberg, Germany); antibodies
against human LMO4 were from Santa Cruz Biotechnology (C15)
and Chemicon (AB5764; Schwalbach, Germany). Anti c-myc
(9E10) and b-III tubulin (SDL3D10) antibodies were obtained from
Sigma-Aldrich (Munich, Germany) and Covance Inc. (TuJ1;
Princeton, NJ, USA), respectively. Antibodies against human RhoA
were purchased from Cytoskeleton (Denver, CO, USA). All
antibodies used in this study are antigen specific (Fig. S1). Unless
otherwise indicated, conditioned medium containing 5 lg/mL
recombinant human RGM A (R&D Systems, Wiesbaden, Germany)
or recombinant mouse Netrin-1 (R&D Systems) was used for
stimulation of neuronal cells.
Expression vectorsThe cDNA encoding full-length Neogenin was amplified by PCR
and subcloned into the mammalian expression vector pcDNA3.1
under control of a cytomegalovirus (CMV) promotor. LMO4 was
amplified by PCR from a MATCHMAKER� (BD CLONTECH,
Mountain View, CA, USA) cDNA library clone and subcloned
downstream of a c-myc epitope tag under control of a CMV
promotor. The cytoplasmatic domain of Neogenin and the P1, P2
and P3 fragments were generated by PCR amplification from the
human Neogenin cDNA clone and subcloned downstream of the
Gal4 DNA-binding domain (GAL-BD) and a c-myc tag into
pGBKT7. Human full length LMO4 and fragments containing the
LIM domains were amplified by PCR and subcloned downstream of
the Gal4 activation domain (GAL4-AD) and a hemagglutinin (HA)
tag into pGADT7.
Yeast-two-hybrid systemThe yeast two-hybrid MATCHMAKER� 3 system (BD CLON-
TECH) was used to screen for potential interacting proteins of the
cytoplasmatic tail of humanNeogenin (aa 1158-1493). A human brain
MATCHMAKER� cDNA library (BD CLONTECH) was cloned
into pACT2 such that it contains a GAL4-AD and an amino-terminal
HA-tag and was pre-transformed into the yeast strain Y187 (MATa,ura3-52, his3-200, ade2-101, trp1-901, leu2-3, 112, gal4D, met–,gal80D, URA3::GAL1UAS-GAL1TATA-lacZ). The cDNA library was
screened by mating with AH109 yeast (MATa, trp1-901, leu2-3, 112,ura3-52, his3-200, gal4D, gal80 D, LYS2::GAL1UAS-GAL1TATA-HIS3,GAL2UAS-GAL2TATA-ADE2,URA3::MEL1UAS-MEL1TATA-lacZ) expressing the Neogenin bait construct. Yeast was initially
selected on quadruple-dropout media that lacked the vector-selective
markers leucine and tryptophan and histidine and adenine; positive
colonies were restreaked on quadruple-dropout media that contained
20 lg/mL X-a-galactosidase (X-a-gal) for blue/white selection.
Single (blue) colonies were picked and restreaked on quadruple-
dropout media containing X-a-gal and 25 mM 3-aminotriazole two
more times. Yeast colonies were grown up for DNA extraction
[Yeast� plasmid kit (Genotech, Montana, USA)]. Purified DNAwas
transformed back into electro-competent Escherichia coli (Stbl2) forDNA amplification. For control experiments, positive clones were
reintroduced into yeast strain Y187 and mated with yeast strain
AH109 transformed either with pGBKT7-Neo-Cyt or with pGBKT7
expressing onlyGAL4-BD.DNA frompositive cloneswas sequenced
and blasted against the GenBank database.
Protein analysis and protein-protein interaction assaysCells were lysed in modified RIPA lysis-buffer [50 mM Tris pH 7.5,
150 mM NaCl, 1 mM EDTA and Complete� protease inhibitor
(Roche, Mannheim, Germany)] containing 0.5% Triton X-100 and
0.5% Deoxycholat (Sigma, Munich, Germany) for 30 min on ice.
Lysates were centrifuged for 10 min at 13 000 g at 4�C and
incubated with the indicated antibodies (1 lg) at 4�C for 5 h in a
final volume of 500 lL lysis buffer with constant rocking. After
addition of protein A-sepharose beads (50% slurry), reactions were
incubated at 4�C for 1 h with constant rocking. Beads were washed
three times with lysis buffer and once with phosphate-buffered
saline (PBS) before boiling them in sodium dodecyl sulfate-sample
buffer (Schaffar et al. 2004). The immune complexes were resolved
by SDS–polyacrylamide gel electrophoresis (SDS–PAGE) (4–20%)
and transferred to nitrocellulose. After immunostaining, western-
blots were developed using ECL (Enhanced chemiluminescent
detection system, GE Healthcare, Munich, Germany).
Rho-activation assayFor RhoA activation analysis, cells were stimulated after 2 h serum
starvation in OptiMEM-1 medium with 5 lg/mL recombinant RGM
A or Netrin-1 (R&D Systems). Cells were washed twice in cell-lysis
buffer (50 mM Tris-HCl, pH 7.5, 10 mM MgCl2, 300 mM NaCl,
2% IGEPAL, including protease inhibitors). Thereon lysates were
incubated with 50 lg GST-tagged Rhotekin-RBD beads (Cytoskel-
eton). After extensive washing in lysis buffer, beads were analyzed
by SDS–PAGE and immunoblotting.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431
LMO4 mediates RGM A signaling in neuronal growth cones | 419

Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431� 2008 The Authors
420 | G. Schaffar et al.

Cell culture and immunostainingHuman SH-SY5Y cells were cultivated in Earle’s balanced salt
solution/F12 (EBSS/F12) medium supplemented with 10% fetal
bovine serum (FBS) and 1% non-essential amino acids (MEM-
NEAA). For induction of neurite outgrowth cells were cultured in
medium supplemented with 10 lM retinoic acid (RA). Human
Ntera2 cells were cultured in Dulbecco’s modified Eagle’s medium
(DMEM) medium supplemented with 10% FBS and 5% horse
serum. For neuronal differentiation, Ntera2 cells were seeded at a
density of 2.5 · 106 cells/175 cm2 in differentiation medium
(DMEM medium supplemented with 10 lM RA) for 3 weeks.
Differentiated neuronal Ntera2 cells were separated from other cells
by tapping and seeded on poly-L-lysine and laminin coated plates
[100 lg/mL and 20 lg/mL, respectively (both Sigma-Aldrich)] in
neurobasal medium supplemented with B27 and glutamine. Rat
cortical neurons were prepared from cerebral cortex of embryonic
day 18 rats. For this, the cortex was digested with papain for 30 min
at 37�C, followed by further dissociation. Dissociated neurons were
plated on dishes pre-coated with poly-L-lysine (100 lg/mL) in
DMEM containing 10% FBS, penicillin, and streptomycin. After
culturing for 1 day, the medium was replaced with DMEM
supplemented with B27. Two days after dissociation, cortical
neuron cultures were either directly analyzed (for LMO4 expres-
sion) or used in transfection and outgrowth assays. HEK293 cells
stable, expressing Neogenin, were cultured in DMEM containing
10% FBS and 200 lg/mL hygromycin B.
Cells were transfected using the non-liposomal lipid transfection
reagent Effectene (Qiagen, Hilden, Germany) following the manu-
facturer’s protocol. In short, cells were seeded the day before
transfection at 50% confluence. DNA was diluted in EC buffer plus
Enhancer (ratio 30 : 1) before mixing with Effectene. After 10 min
incubation at 21�C, the whole mixture was added to the cells. All
transfections were carried out in growth medium. Transfections of
small interfering RNA (siRNA) (Dharmacon, SMARTpools) were
carried out according to the manufacturer’s recommended procedures
using Dharmacon-1 as transfection reagent (Dharmacon, Lafayette,
CO, USA). Cells were routinely analyzed 48–72 h after transfection.
For immunofluorescence analysis, cells were fixed in 2%
paraformaldehyde for 2 h at 21�C and permeabilized with PBS
containing 0.1% Triton X-100 (15 min at 21�C). After blocking
with 1% bovine serum albumin, cells were incubated with the
indicated primary and secondary antibodies (diluted in PBS with
0.1% bovine serum albumin) for 1 h each. Nuclei were counter-
stained using bisbenzimide (H33258) and embedded in ProLong
antifade mounting medium (Invitrogen, Karlsruhe, Germany). Cells
were analyzed using a fluorescence microscope (Axioplan2, Zeiss,
Gottingen, Germany).
Neurite outgrowth assayFor outgrowth analysis of differentiated human SH-SY5Y, Ntera2
cells and rat cortical neurons, cells were incubated with or without
recombinant RGM A 25 lg/mL for 24 h. RGM A stripes were
prepared as described previously (Rajagopalan et al. 2004) using
poly-L-lysine/laminin coated glass slides and 25 lg/mL RGM A,
respectively. For neurite outgrowth analysis, cells were either
stained with b-III tubulin (SH-SY5Y) or TuJ1 (cortical neurons)
antibodies. Length of neurites was measured and analysed using
the AxioVision software (Zeiss).
Results
Interaction of LMO4 with NeogeninTo map the RGM A – Neogenin signaling pathway, wesearched for new interacting proteins of human Neogenin(Neo). For this, we used the C-terminal cytoplasmic domain[Neo-Cyt (aa1158-1493)] as the bait in a yeast two-hybridscreen. Screening a human brain cDNA library, we isolatedthe LIM-only protein 4 (LMO4) as a potential interactingpartner (Fig. 1a). For further analysis, we tested variousfragments of both proteins. Interaction analysis in yeast cellsusing a-galactosidase and 3-AT sensitivity as markers
Fig. 1 LMO4 – Neogenin interaction in a yeast-two hybrid system.
(a) LMO4 interacts with the cytoplasmatic domain of Neogenin in
yeast. AH109 yeast cells co-transformed either with the AD-Gal-
LMO4, the BD-Gal-Neogenin (Neo-Cyt) or the respective Gal-domains
alone (empty vector) were grown on SD-His/-Leu/-Trp/-Ade/3-AT
selective plates with a-galactosidase. (b) LMO4 interaction with the
cytoplasmic domain of Neogenin is mediated by the conserved P2 and
P3 domains of Neogenin. Interaction of LMO4 with Neogenin domains
was analysed and scored by a-galactosidase activity [()) no blue
coloring after 48 h incubation, (+/)) blue coloring between 24 and 48 h
incubation, (+) blue coloring within 24 h incubation, (++) blue coloring
within 12 h incubation]. LMO4 binding to the P3 domain alone is
weaker than binding to a P2 domain containing protein. The fusion
protein containing both domains (P2down) is binding as efficient as the
full length cytoplasmic Neogenin protein. (c) LMO4 LIM domain
binding to Neogenin. Both LIM domains of LMO4 can separately
interact with Neogenin, but the binding efficiency is decreased com-
pared to full length LMO4. (d) Co-immunoprecipitation of HA-tagged
LMO4 and c-myc-tagged Neogenin in yeast. The cytoplasmic, c-myc-
tagged domain of Neogenin (c-myc-BD-Neo-Cyt) was immunopre-
cipitated from yeast cell lysates, over-expressing either HA-tagged
AD-LMO4, c-myc-tagged BD-Neo-Cyt or both proteins together using
c-myc conjugated agarose beads. Precipitates were analyzed by
SDS–PAGE and western blotting using the indicated antibodies. (e–f)
LMO4/Neogenin interaction in HEK293 cells stable over-expressing
full length Neogenin. Cells were transiently transfected with a c-myc-
LMO4 over-expression plasmid. Lysates were incubated with Neo-
genin- or c-myc antibodies. The resulting immunocomplexes were
subjected to SDS–PAGE and western blotting and analyzed with the
indicated antibodies. (g–i) LMO4/Neogenin interaction in neuronal
cells. Cell lysates of SH-SY5Y cells (g), differentiated Ntera2 cells (h),
and rat cortical neurons (i) were incubated with Neogenin or control
antibodies (IgG). The resulting immunocomplexes were analyzed by
SDS–PAGE and western blotting using LMO4 antibodies (*, unspecific
antibody bands after immunoprecipitation).
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431
LMO4 mediates RGM A signaling in neuronal growth cones | 421

demonstrated that the interaction did not require the P1domain of Neogenin (Fig. 1b). P1 deletion had no effect onLMO4 binding; furthermore the P1 domain alone failed tointeract with LMO4. The P3 domain alone, which has beenshown to be important and sufficient for the interaction ofDCC and Neogenin with focal adhesion kinase (FAK) andthe phosphatidylinositol transfer protein a (PITPa), onlyweakly interacted with LMO4. In contrast the interactionbetween the P2 domain containing fragment was clearlystronger compared to the binding to the P3 domain, butweaker compared to a protein spanning both the P2 and theP3 domain, indicating that both conserved regions ofNeogenin are probably contributing to a stable LMO4/Neogenin interaction. LMO4 in turn mainly consists of twoconserved LIM domains of approximately 55 amino acids(amino acids 23–82 and 87–144) containing highly con-served cysteines and histidines. To investigate the role of thesingle LIM domains regarding the Neogenin interaction, wetested both LIM domains separately in the yeast two-hybridsystem. Both LIM domains alone were able to interact withthe cytoplasmic region of Neogenin, although the interactionwas clearly stronger using the full length protein (Fig. 1c).
To confirm the LMO4/Neogenin interaction, we performedpull down experiments from yeast lysates, after over-expressing c-myc-tagged DNA-BD Neogenin fusion protein,HA-tagged DNA-AD LMO4 fusion protein or both proteinstogether. Using anti-c-myc conjugated agarose beads wewere able to precipitate the c-myc-tagged Neogenin fusionprotein (data not shown). The HA-tagged LMO4 co-precip-itated in lysates from yeast over-expressing both fusionproteins, but not from yeast over-expressing only the LMO4fusion protein (Fig. 1d).
For analysis of the LMO4/Neogenin interaction in mam-malian cells, we performed immunoprecipitation experimentswith lysates from HEK293 cells stable over-expressing fulllength human Neogenin and transiently expressing c-myc-tagged LMO4. Using an anti-Neogenin antibody (H175) wecould co-precipitate the over-expressed LMO4, demonstrat-ing that LMO4 also interacts with Neogenin in mammaliancells (Fig. 1e). In a reverse experiment, we could alsoco-immunoprecipitate Neogenin by using an anti-c-mycantibody (9E10, Sigma) (Fig. 1f). We regarded the associ-ation specific, as neither LMO4 nor Neogenin was detectedin immunoprecipitates from non-(LMO4)transfected cells(mock). To analyze the association between endogenouslyexpressed LMO4 and Neogenin in neuronal cells, wetested cell lysates from SH-SY5Y cells (Fig. 1g), fromdifferentiated neuronal Ntera2 cells (Fig. 1h) and fromprimary rat cortical neurons (Fig. 1i). In all three cases,LMO4 was detected in immunoprecipitates using an anti-Neogenin antibody. LMO4 could not be detected in non-specific immunocomplexes. These results indicate thatLMO4 interacts with Neogenin in yeast, in human neuronalcells and in rat primary neurons.
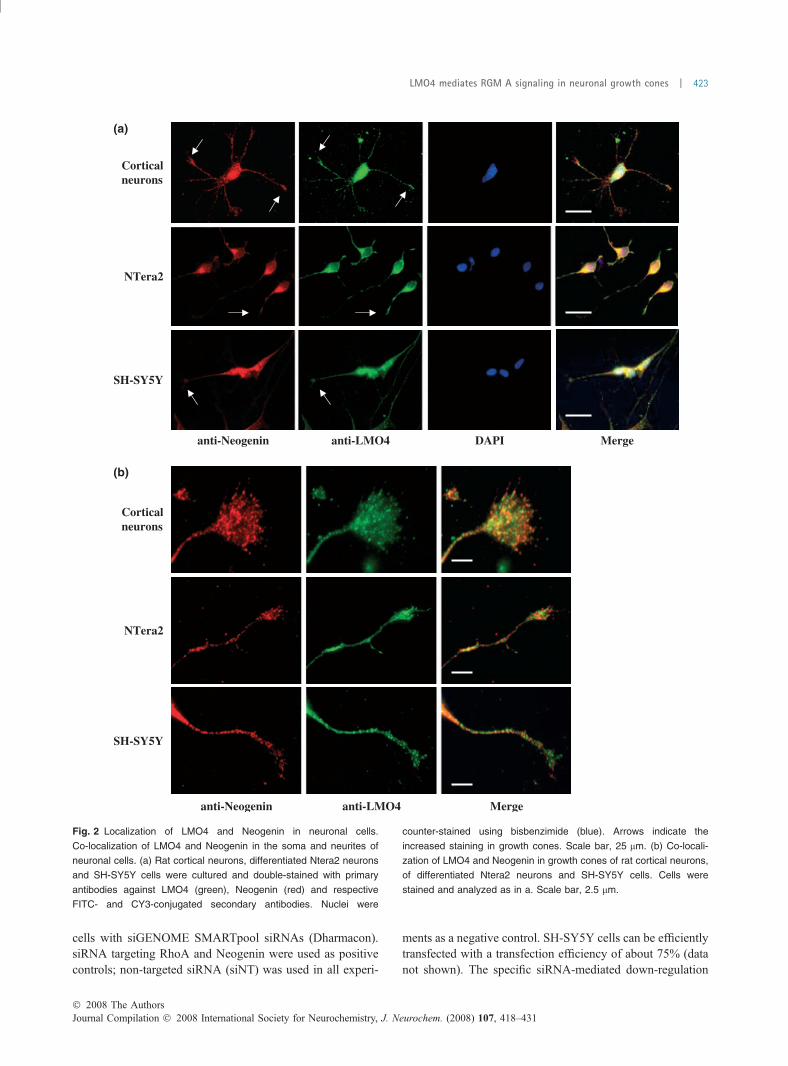
Co-localization of LMO4 and Neogenin in neuronal cellsTo analyze the localization and distribution of LMO4 andNeogenin in SH-SY5Y cells, differentiated Ntera2 cells andrat cortical neurons we performed immunofluorescenceexperiments. Cells were cultured on glass plates for 24 hor 48 h (cortical neurons), respectively, fixed withparaformaldehyde and double stained with LMO4- andNeogenin-specific antibodies. The resulting immunostainingconfirmed data from others (Novotny-Diermayr et al. 2005).LMO4 was not only expressed in the nucleus but also in thesoma and the neurites. We found the highest expression forLMO4 in all tested neurons in the nucleus and the growthcones (Fig. 2a, arrows). Neogenin was detected primarily inthe soma, a strong expression was found in neurites and thegrowth cone, represented by a dotted membrane staining,similar to that described for DCC in neuronal cells.
Merging the Neogenin and LMO4 immunoreactivitydemonstrates a significant co-localization in growth conesand neurites of SH-SY5Y cells, Ntera2 and rat corticalneurons. We found an overlapping pattern, especially in thecentral and peripheral domains of the growth cones, in thefilopodia and at the base of lamellipodia (Fig. 2b).
Regulation of neurite outgrowth by LMO4To analyze the role of LMO4 in neurite outgrowth, weinvestigated the effect of over-expressing and deleting LMO4in the human neuroblastoma cell line SH-SY5Y. Treatment ofSH-SY5Ycellswith retinoic acid (RA) can be easilymonitoredby changes in the cell morphology as it results in a rapidformation of neurite-like extensions (Encinas et al. 2000),which we name neurite in the further course of the text. Foranalysis of LMO4 over-expression, we co-transfectedSH-SY5Y cells with a LMO4 and an EGFP over-expressingplasmid under the control of CMV promotors and inducedneurite outgrowth by RA treatment for 48 h. The analysis oftransfected (green) cells by fluorescence microscopy showed,that over-expression of LMO4 clearly induced a change in cellmorphology and decreased the number of cells with neurite-like extensions (Fig. 3a). EGFP/LMO4 double transfectedcells had a flat and more outspread morphology. In contrast,EGFP/mock transfected cells typically had a spindle-likemorphology with two (or more) neurites (Fig. 3a). Quantifi-cation of the number of cells with neurites in control and inLMO4 transfected cells confirmed these observations. Over-expression of LMO4 completely reversed the neurite out-growth effect of RA on SH-SY5Y cells (Fig. 3b). Accordingly,we found that the average neurite length (described insupporting information) in cells transfected with LMO4 wassignificantly reduced compared to control cells (Fig. 3c). Thesedata demonstrate that LMO4plays a role in neurite growth, andcan reverse or neutralize neurite outgrowth promoting signals.
We next examined the effect of LMO4 depletion inSH-SY5Y cells. To specifically silence LMO4 expression,we used RNA interference (RNAi). For this, we transfected
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431� 2008 The Authors
422 | G. Schaffar et al.
cells with siGENOME SMARTpool siRNAs (Dharmacon).siRNA targeting RhoA and Neogenin were used as positivecontrols; non-targeted siRNA (siNT) was used in all experi-
ments as a negative control. SH-SY5Y cells can be efficientlytransfected with a transfection efficiency of about 75% (datanot shown). The specific siRNA-mediated down-regulation
Fig. 2 Localization of LMO4 and Neogenin in neuronal cells.
Co-localization of LMO4 and Neogenin in the soma and neurites of
neuronal cells. (a) Rat cortical neurons, differentiated Ntera2 neurons
and SH-SY5Y cells were cultured and double-stained with primary
antibodies against LMO4 (green), Neogenin (red) and respective
FITC- and CY3-conjugated secondary antibodies. Nuclei were
counter-stained using bisbenzimide (blue). Arrows indicate the
increased staining in growth cones. Scale bar, 25 lm. (b) Co-locali-
zation of LMO4 and Neogenin in growth cones of rat cortical neurons,
of differentiated Ntera2 neurons and SH-SY5Y cells. Cells were
stained and analyzed as in a. Scale bar, 2.5 lm.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431
LMO4 mediates RGM A signaling in neuronal growth cones | 423

on the RNA level was quantified by RT-PCR. We calculatedan mRNA decrease between 80% and 95%, which was in therange of the manufacturer’s specifications. The biochemicalanalysis of protein expression levels demonstrated for all threeproteins analysed a specific down-regulation (Fig. S3). Inaccordance, immunofluorescence analysis of control andsiRNA transfected cells showed a distinct reduction in RhoA,Neogenin and LMO4 staining (data not shown).
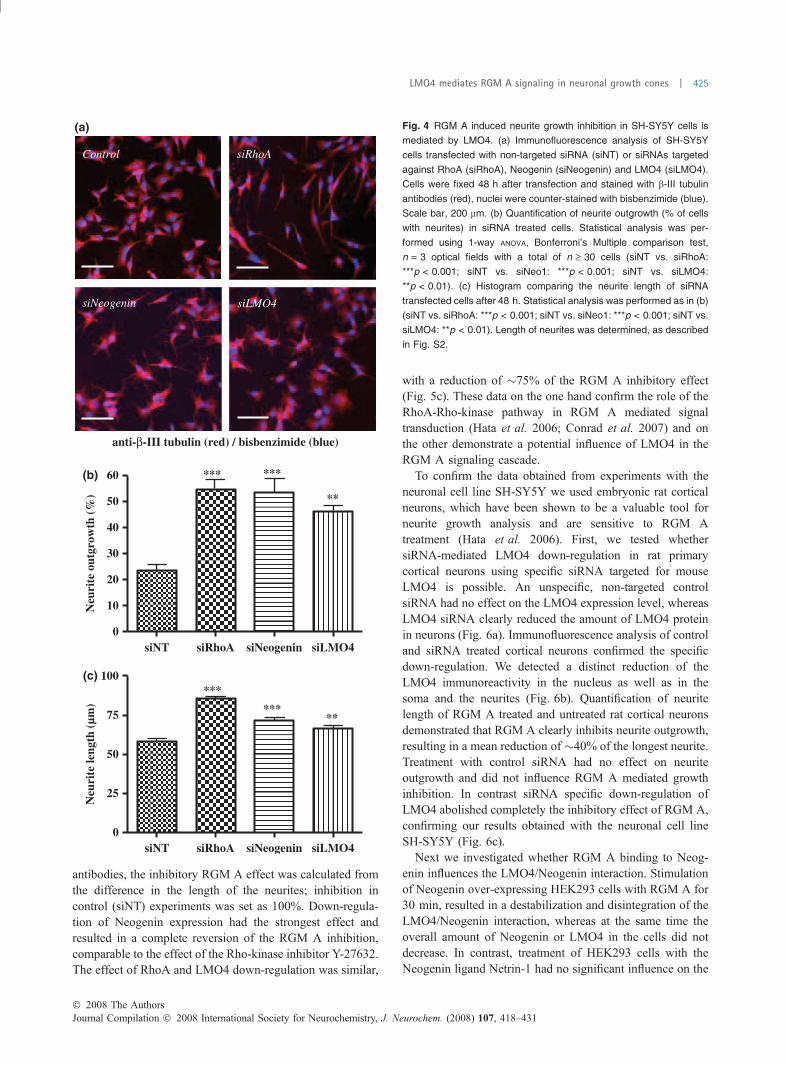
We determined and compared the effect of LMO4depletion with that of RhoA and Neogenin down-regulation.siRNA transfected SH-SY5Y cells were cultured for 48 h,fixed with paraformaldehyde and immunostained with Cy3-conjugated anti b-III tubulin antibodies and bisbenzimide.Down-regulation of RhoA in SH-SY5Y cells clearly inducedneurites and stimulated the outgrowth as documented by thesignificant increases in long neurites, very similar to theeffect observed after RA treatment. A comparable but weakereffect was detected after siRNA-mediated knock-down ofNeogenin or LMO4 (Fig. 4a) Quantification revealed thatthe number of neurite-like extensions doubled after RhoAand Neogenin down-regulation. The effect of LMO4 down-regulation was slightly (but not significantly) smaller com-pared with the effect obtained by down-regulating Neogeninand RhoA (Fig. 4b). We next compared the medium length ofthe longest neurite and found a very robust increase afterRhoA down-regulation. The effect of Neogenin and LMO4down-regulation was comparable, down-regulation of bothproteins resulted in a significant increase of neurite length(Fig. 4c). These data indicate that LMO4 canmodulate neuriteextension and outgrowth. Both LMO4 over-expression anddown-regulation affects the number of SH-SY5Y cells withneurite-like extensions and the length of their extensions.
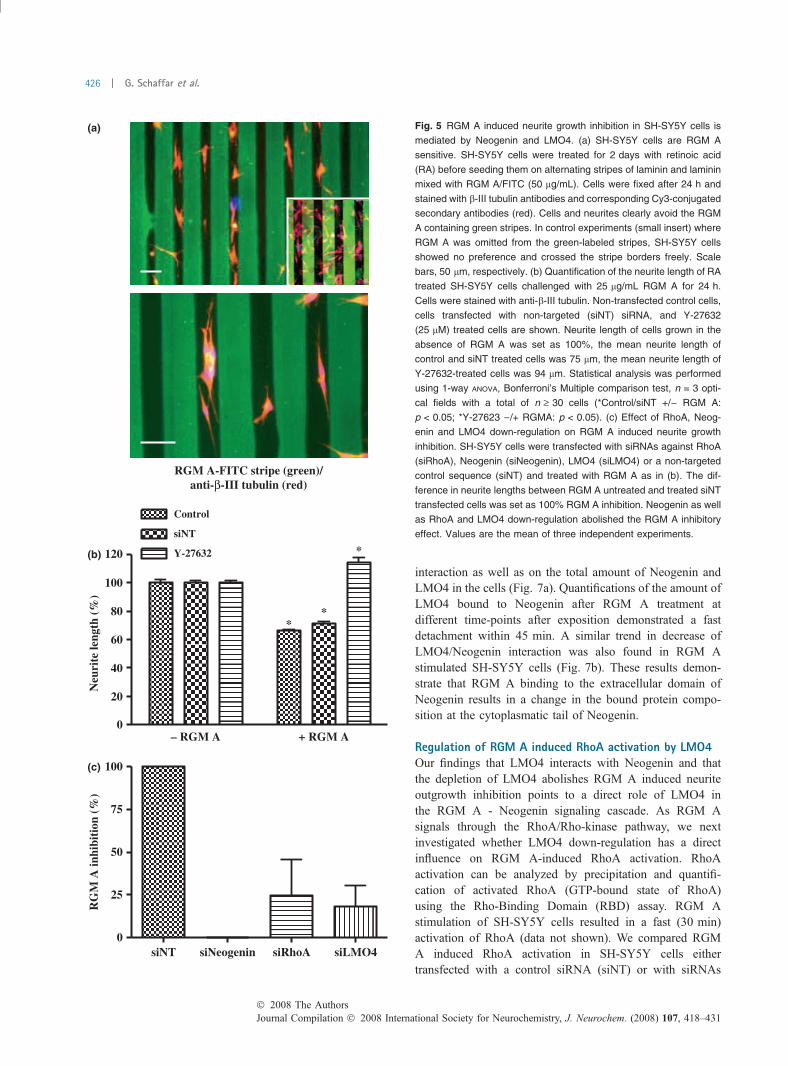
Modulation of RGM A signaling by LMO4We next investigated the role of LMO4 in RGM A mediatedneurite growth inhibition. For this, we first analyzed theSH-SY5Y sensitivity toward RGM A using a stripe assay.RA treated SH-SY5Y cells were seeded onto alternatingstripes of laminin and laminin plus RGM A and analyzed fortheir growth preference after 24 h (Fig. 5a). SH-SY5Y cellspreferentially attached on the laminin stripes and avoided theRGM A stripes (green). In addition, growing neurites avoidthe RGM A stripes and grew parallel along the stripe border.In control experiments (Fig. 5a, small insert), in which RGMA was omitted from the green stripes, we found no selectiveattachment and growth preference of the SH-SY5Y cells.
To quantify the RGM A growth inhibitory effect in cellcultures, we measured the length of neurites from SH-SY5Ycells, incubated with or without 25 lg/mL RGM A for 24 h.RGM A treatment reduced the medium length of SH-SY5Yneurites by �40% compared with control cells. The inhib-itory effect was abolished by a simultaneous treatment withthe Rho-kinase inhibitor Y-27632 (25 lM), confirming theimportance of the RhoA-Rho kinase pathway in the RGM Ainduced neurite growth inhibition in SH-SY5Y cells and thesuitability of SH-SY5Y neurite-like extensions for analysisof RGM A signaling (Fig. 5b).
To determine whether LMO4 is involved in the RGM Ainduced Neogenin signaling cascade we transfected SH-SY5Y cells with siRNAs targeted against RhoA, Neogenin,LMO4 or a non-targeted control and cultured the cells withor without RGM A for 24 h. After staining with b-III tubulin
Fig. 3 LMO4 over-expression in SH-SY5Y cells. (a) Immunofluores-
cence analysis of SH-SY5Y cells transiently transfected with EGFP or
EGFP and LMO4 over-expressing plasmids. Cells were analyzed 48 h
after transfection. Scale bar, 100 lm. (b) Quantification of neurite
outgrowth (% of cells with neurites) of EGFP/LMO4 over-expressing
cells treated with or without retinoic acid (RA) for 48 h. The length of
neurites was determined using Zeiss AxioVision software, as
described in Fig. S2. (c) Quantification of neurite length of EGFP
(control) or EGFP and LMO4 over-expressing SH-SY5Y cells. Cells
were treated and analyzed as in (b) (*p < 0.05, unpaired, two-tailed
t test, n ‡ 25 cells).
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431� 2008 The Authors
424 | G. Schaffar et al.
antibodies, the inhibitory RGM A effect was calculated fromthe difference in the length of the neurites; inhibition incontrol (siNT) experiments was set as 100%. Down-regula-tion of Neogenin expression had the strongest effect andresulted in a complete reversion of the RGM A inhibition,comparable to the effect of the Rho-kinase inhibitor Y-27632.The effect of RhoA and LMO4 down-regulation was similar,
with a reduction of �75% of the RGM A inhibitory effect(Fig. 5c). These data on the one hand confirm the role of theRhoA-Rho-kinase pathway in RGM A mediated signaltransduction (Hata et al. 2006; Conrad et al. 2007) and onthe other demonstrate a potential influence of LMO4 in theRGM A signaling cascade.
To confirm the data obtained from experiments with theneuronal cell line SH-SY5Y we used embryonic rat corticalneurons, which have been shown to be a valuable tool forneurite growth analysis and are sensitive to RGM Atreatment (Hata et al. 2006). First, we tested whethersiRNA-mediated LMO4 down-regulation in rat primarycortical neurons using specific siRNA targeted for mouseLMO4 is possible. An unspecific, non-targeted controlsiRNA had no effect on the LMO4 expression level, whereasLMO4 siRNA clearly reduced the amount of LMO4 proteinin neurons (Fig. 6a). Immunofluorescence analysis of controland siRNA treated cortical neurons confirmed the specificdown-regulation. We detected a distinct reduction of theLMO4 immunoreactivity in the nucleus as well as in thesoma and the neurites (Fig. 6b). Quantification of neuritelength of RGM A treated and untreated rat cortical neuronsdemonstrated that RGM A clearly inhibits neurite outgrowth,resulting in a mean reduction of �40% of the longest neurite.Treatment with control siRNA had no effect on neuriteoutgrowth and did not influence RGM A mediated growthinhibition. In contrast siRNA specific down-regulation ofLMO4 abolished completely the inhibitory effect of RGM A,confirming our results obtained with the neuronal cell lineSH-SY5Y (Fig. 6c).
Next we investigated whether RGM A binding to Neog-enin influences the LMO4/Neogenin interaction. Stimulationof Neogenin over-expressing HEK293 cells with RGM A for30 min, resulted in a destabilization and disintegration of theLMO4/Neogenin interaction, whereas at the same time theoverall amount of Neogenin or LMO4 in the cells did notdecrease. In contrast, treatment of HEK293 cells with theNeogenin ligand Netrin-1 had no significant influence on the
Fig. 4 RGM A induced neurite growth inhibition in SH-SY5Y cells is
mediated by LMO4. (a) Immunofluorescence analysis of SH-SY5Y
cells transfected with non-targeted siRNA (siNT) or siRNAs targeted
against RhoA (siRhoA), Neogenin (siNeogenin) and LMO4 (siLMO4).
Cells were fixed 48 h after transfection and stained with b-III tubulin
antibodies (red), nuclei were counter-stained with bisbenzimide (blue).
Scale bar, 200 lm. (b) Quantification of neurite outgrowth (% of cells
with neurites) in siRNA treated cells. Statistical analysis was per-
formed using 1-way ANOVA, Bonferroni’s Multiple comparison test,
n = 3 optical fields with a total of n ‡ 30 cells (siNT vs. siRhoA:
***p < 0.001; siNT vs. siNeo1: ***p < 0.001; siNT vs. siLMO4:
**p < 0.01). (c) Histogram comparing the neurite length of siRNA
transfected cells after 48 h. Statistical analysis was performed as in (b)
(siNT vs. siRhoA: ***p < 0.001; siNT vs. siNeo1: ***p < 0.001; siNT vs.
siLMO4: **p < 0.01). Length of neurites was determined, as described
in Fig. S2.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431
LMO4 mediates RGM A signaling in neuronal growth cones | 425
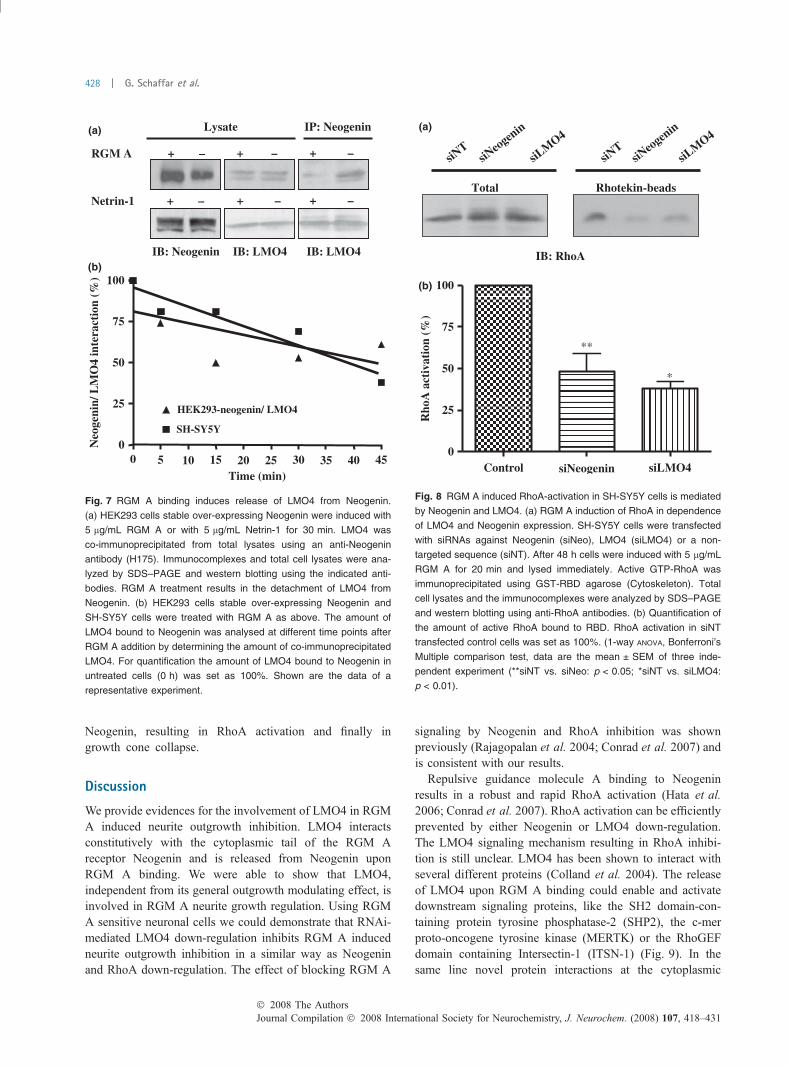
interaction as well as on the total amount of Neogenin andLMO4 in the cells (Fig. 7a). Quantifications of the amount ofLMO4 bound to Neogenin after RGM A treatment atdifferent time-points after exposition demonstrated a fastdetachment within 45 min. A similar trend in decrease ofLMO4/Neogenin interaction was also found in RGM Astimulated SH-SY5Y cells (Fig. 7b). These results demon-strate that RGM A binding to the extracellular domain ofNeogenin results in a change in the bound protein compo-sition at the cytoplasmatic tail of Neogenin.
Regulation of RGM A induced RhoA activation by LMO4Our findings that LMO4 interacts with Neogenin and thatthe depletion of LMO4 abolishes RGM A induced neuriteoutgrowth inhibition points to a direct role of LMO4 inthe RGM A - Neogenin signaling cascade. As RGM Asignals through the RhoA/Rho-kinase pathway, we nextinvestigated whether LMO4 down-regulation has a directinfluence on RGM A-induced RhoA activation. RhoAactivation can be analyzed by precipitation and quantifi-cation of activated RhoA (GTP-bound state of RhoA)using the Rho-Binding Domain (RBD) assay. RGM Astimulation of SH-SY5Y cells resulted in a fast (30 min)activation of RhoA (data not shown). We compared RGMA induced RhoA activation in SH-SY5Y cells eithertransfected with a control siRNA (siNT) or with siRNAs
Fig. 5 RGM A induced neurite growth inhibition in SH-SY5Y cells is
mediated by Neogenin and LMO4. (a) SH-SY5Y cells are RGM A
sensitive. SH-SY5Y cells were treated for 2 days with retinoic acid
(RA) before seeding them on alternating stripes of laminin and laminin
mixed with RGM A/FITC (50 lg/mL). Cells were fixed after 24 h and
stained with b-III tubulin antibodies and corresponding Cy3-conjugated
secondary antibodies (red). Cells and neurites clearly avoid the RGM
A containing green stripes. In control experiments (small insert) where
RGM A was omitted from the green-labeled stripes, SH-SY5Y cells
showed no preference and crossed the stripe borders freely. Scale
bars, 50 lm, respectively. (b) Quantification of the neurite length of RA
treated SH-SY5Y cells challenged with 25 lg/mL RGM A for 24 h.
Cells were stained with anti-b-III tubulin. Non-transfected control cells,
cells transfected with non-targeted (siNT) siRNA, and Y-27632
(25 lM) treated cells are shown. Neurite length of cells grown in the
absence of RGM A was set as 100%, the mean neurite length of
control and siNT treated cells was 75 lm, the mean neurite length of
Y-27632-treated cells was 94 lm. Statistical analysis was performed
using 1-way ANOVA, Bonferroni’s Multiple comparison test, n = 3 opti-
cal fields with a total of n ‡ 30 cells (*Control/siNT +/) RGM A:
p < 0.05; *Y-27623 )/+ RGMA: p < 0.05). (c) Effect of RhoA, Neog-
enin and LMO4 down-regulation on RGM A induced neurite growth
inhibition. SH-SY5Y cells were transfected with siRNAs against RhoA
(siRhoA), Neogenin (siNeogenin), LMO4 (siLMO4) or a non-targeted
control sequence (siNT) and treated with RGM A as in (b). The dif-
ference in neurite lengths between RGM A untreated and treated siNT
transfected cells was set as 100% RGM A inhibition. Neogenin as well
as RhoA and LMO4 down-regulation abolished the RGM A inhibitory
effect. Values are the mean of three independent experiments.
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431� 2008 The Authors
426 | G. Schaffar et al.

targeted against Neogenin or LMO4. Both Neogenin andLMO4 down-regulation in SH-SY5Y neurons resulted in aclear inhibition of the RGM A induced RhoA activation
by more than 50% compared to the control (Fig. 8). Thisdemonstrates, that LMO4 must be involved in the RGM Asignaling cascade induced after RGM A binding to
Fig. 6 LMO4 mediates the RGM A neurite
outgrowth inhibition in rat cortical neurons.
(a) siRNA mediated down-regulation of
LMO4 in primary rat cortical neurons. Neu-
rons were transfected with non-targeted
control siRNA (siNT) or LMO4 siRNA (siL-
MO4). Cells were lysed 48 h after trans-
fection and total lysates were analyzed by
SDS–PAGE and western blotting using the
indicated antibodies. The neuron-specific
b-III tubulin protein (upper blot) was used as
an unspecific control. A specific reduction of
LMO4 expression was observed in cells
transfected with siRNA targeted LMO4.
(b) Immunofluorescence analysis of siRNA-
mediated LMO4 down-regulation. Rat cor-
tical neurons transfected with control siRNA
(siNT) or siRNA against LMO4 (siLMO4)
were fixed after 48 h and immunostained
for LMO4 (green), the neuron-specific b-III
tubulin protein (Tuj1; red) and respective
FITC- and Cy3-conjugated secondary anti-
bodies. LMO4 is detected in the nucleus as
well as the soma and neurites of rat cortical
neurons. A clear down-regulation of LMO4
in cells transfected with siRNA against
LMO4 is observed (arrow). Scale bar,
50 lm. (c) Quantification of the neurite
length of siRNA treated cells. The medium
length of cortical neurons was significantly
reduced when neurons were treated with
RGM A. However, this neurite growth
inhibitory effect was completely abolished
when the neurons were transfected with
siLMO4. Statistical analysis was performed
using 1-way ANOVA, Bonferroni’s Multiple
comparison test. Data are the mean ± SEM
100 neurites were measured per experi-
ment (*p < 0.05).
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431
LMO4 mediates RGM A signaling in neuronal growth cones | 427
Neogenin, resulting in RhoA activation and finally ingrowth cone collapse.
Discussion
We provide evidences for the involvement of LMO4 in RGMA induced neurite outgrowth inhibition. LMO4 interactsconstitutively with the cytoplasmic tail of the RGM Areceptor Neogenin and is released from Neogenin uponRGM A binding. We were able to show that LMO4,independent from its general outgrowth modulating effect, isinvolved in RGM A neurite growth regulation. Using RGMA sensitive neuronal cells we could demonstrate that RNAi-mediated LMO4 down-regulation inhibits RGM A inducedneurite outgrowth inhibition in a similar way as Neogeninand RhoA down-regulation. The effect of blocking RGM A
signaling by Neogenin and RhoA inhibition was shownpreviously (Rajagopalan et al. 2004; Conrad et al. 2007) andis consistent with our results.
Repulsive guidance molecule A binding to Neogeninresults in a robust and rapid RhoA activation (Hata et al.2006; Conrad et al. 2007). RhoA activation can be efficientlyprevented by either Neogenin or LMO4 down-regulation.The LMO4 signaling mechanism resulting in RhoA inhibi-tion is still unclear. LMO4 has been shown to interact withseveral different proteins (Colland et al. 2004). The releaseof LMO4 upon RGM A binding could enable and activatedownstream signaling proteins, like the SH2 domain-con-taining protein tyrosine phosphatase-2 (SHP2), the c-merproto-oncogene tyrosine kinase (MERTK) or the RhoGEFdomain containing Intersectin-1 (ITSN-1) (Fig. 9). In thesame line novel protein interactions at the cytoplasmic
Fig. 7 RGM A binding induces release of LMO4 from Neogenin.
(a) HEK293 cells stable over-expressing Neogenin were induced with
5 lg/mL RGM A or with 5 lg/mL Netrin-1 for 30 min. LMO4 was
co-immunoprecipitated from total lysates using an anti-Neogenin
antibody (H175). Immunocomplexes and total cell lysates were ana-
lyzed by SDS–PAGE and western blotting using the indicated anti-
bodies. RGM A treatment results in the detachment of LMO4 from
Neogenin. (b) HEK293 cells stable over-expressing Neogenin and
SH-SY5Y cells were treated with RGM A as above. The amount of
LMO4 bound to Neogenin was analysed at different time points after
RGM A addition by determining the amount of co-immunoprecipitated
LMO4. For quantification the amount of LMO4 bound to Neogenin in
untreated cells (0 h) was set as 100%. Shown are the data of a
representative experiment.
Fig. 8 RGM A induced RhoA-activation in SH-SY5Y cells is mediated
by Neogenin and LMO4. (a) RGM A induction of RhoA in dependence
of LMO4 and Neogenin expression. SH-SY5Y cells were transfected
with siRNAs against Neogenin (siNeo), LMO4 (siLMO4) or a non-
targeted sequence (siNT). After 48 h cells were induced with 5 lg/mL
RGM A for 20 min and lysed immediately. Active GTP-RhoA was
immunoprecipitated using GST-RBD agarose (Cytoskeleton). Total
cell lysates and the immunocomplexes were analyzed by SDS–PAGE
and western blotting using anti-RhoA antibodies. (b) Quantification of
the amount of active RhoA bound to RBD. RhoA activation in siNT
transfected control cells was set as 100%. (1-way ANOVA, Bonferroni’s
Multiple comparison test, data are the mean ± SEM of three inde-
pendent experiment (**siNT vs. siNeo: p < 0.05; *siNT vs. siLMO4:
p < 0.01).
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431� 2008 The Authors
428 | G. Schaffar et al.

domain of Neogenin are thinkable [e.g. FAK, Paxillin (Renet al. 2004; Novotny-Diermayr et al. 2005)]. Additionally,after nuclear entry LMO4, providing a link to the transcrip-tion machinery, could act as a mediator of long-term neuriteoutgrowth changes. This would provide a novel mechanismto explain the complex behavior of axons in response toneurite growth inhibitors like RGM A. LMO4 is an excellentmolecular link for the intimate association and interaction ofaxon guidance and transcriptional regulation. LMO4 plays animportant role in several transcription pathways (Sum et al.2002), e.g. it has been shown that NMDA receptors andvoltage-dependent calcium channels signal via LMO4,inducing its association with the cAMP-response-element-binding protein and LIM homeodomain co-factors (Kashaniet al. 2006). Interestingly we found that the LMO4 releasefrom Neogenin upon RGM A binding is in the same timeframe as the RhoA activation. The activation of RhoA is afast process (within minutes), which excludes a transcrip-tional based LMO4 signaling, but this does not exclude thepossibility that LMO4 in addition acts on a long-termtranscriptional regulatory level (Raivich and Makwana2007).
The closest homolog of the RGM A receptor Neogenin isthe Netrin-1 receptor Deleted in Colorectal Cancer (DCC)(Vielmetter et al. 1994). Netrin-1 binding to DCC results ingrowth cone expansion and neurite outgrowth (Keino-Masu
et al. 1996). Although both receptors share 50% amino acididentity and an identical domain structure, their cytoplasmicregions with the exception of three homologous regions(called P1, P2, and P3) are much less conserved (Muelleret al. 2006). The P3 domain of DCC has been shown to beimportant for Netrin-1 induced neurite outgrowth (Steinet al. 2001). In addition Focal adhesion kinase (FAK) andPhosphatidylinositol transfer protein a (PITPa) specificallyinteract with the P3 domain of DCC and mediate Netrin-1induced neurite outgrowth (Ren et al. 2004; Xie et al. 2005).FAK and PITPa also bind to the P3 domain of Neogenin, buta functional role for this interaction has not yet been proven.
We demonstrate that LMO4 is another interacting partnerof the P3 domain of Neogenin. However in contrast to thedescribed FAK and PITPa interaction we detected a secondLMO4 interaction domain within the cytoplasmatic region ofNeogenin, located at the P2 region. As LMO4 contains twohighly homologous LIM interaction domains (75% aaidentity) with no discrete binding motif (Bach 2000; Xuet al. 2003) it is possible, that both domains bind to separatedomains within Neogenin. Based on the determined bondstrength of the analysed Neogenin fragments, we hypothesizethat the binding of both LIM domains stabilize the overallbinding of LMO4. A similar binding mechanism has beenproposed for the LMO4/LIM-domain binding protein inter-action (Deane et al. 2004). The LIM domains do not contain
Fig. 9 Model of RGM A – Neogenin sig-
naling. LMO4 binds to the cytoplasmatic
domain of Neogenin. Binding of RGM A at
the extracellular site of Neogenin induces
the release of LMO4, probably followed by a
conformational change in the cytoplasmic
domain, which may influence other inter-
action partners of Neogenin like the focal
adhesion kinase (FAK) or Paxillin. Free
LMO4 could interact and activate other
proteins, e.g. the tyrosine phosphatase
SHP2, the tyrosine kinase MERTK, and
Intersectin (ITSN), resulting in a fast RhoA
activation. Simultaneously LMO4, after
nuclear entry, can lead to a long term
change by interacting with transcriptional
regulators as CREB.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431
LMO4 mediates RGM A signaling in neuronal growth cones | 429

an intrinsic catalytic activity, but may modulate the activitiesof their binding partners (Schmeichel and Beckerle 1994).Binding of LMO4 could probably induce a conformationalchange in the cytoplasmic domain resulting in a differentbinding environment for other Neogenin interaction partners.Such a conformational regulation is a common mechanismfor LIM domain based activity control, e.g. it is described forthe LIM dependent autoinhibition of the LIM kinase, aprotein involved in cytoskeletal regulation (Nagata et al.1999).
Although LMO4 has been initially described as a tran-scriptional regulator (Sugihara et al. 1998), interacting withdifferent transcriptional co-regulators and transcription fac-tors (Matthews and Visvader 2003), we demonstrate thatLMO4 in neuronal cells is not restricted to the nucleus.Immunofluorescence of neuronal cells demonstrates thatLMO4 is highly expressed in the nucleus as well asthroughout the cytoplasm and in growth cones. SeveralLIM containing proteins – mostly components of dynamicneurite growth structures – are involved in neuronal path-finding, e.g. LIM kinase, the actin binding LIM protein, andPaxillin (Kadrmas and Beckerle 2004). Some of the LIM-containing proteins have been shown to function both in thecytoplasm and the nucleus (Kadrmas and Beckerle 2004).
LIM-only protein 4 plays an important role in mammaliandevelopment (Visvader et al. 2001; Mousses et al. 2002;Mizunuma et al. 2003). It is also required for proper nervoussystem development (Chen et al. 2002). LMO4 knock-outmice die at birth and approximately 50% of the homozygousknock out mice exhibiting defects in neural tube closure(Hahm et al. 2004; Lee et al. 2005). A very similarphenotype with approximately 50% of the RGM A knock-out mice showing defects in cranial neural tube closure andexencephaly has been reported (Niederkofler et al. 2004),suggesting that both genes act in the same pathway.Interestingly RGM A, Neogenin and LMO4 have verysimilar expression patterns in the adult mouse brain (Fig. S4,Lein et al. 2007). The importance of LMO4 for propernervous system development has recently been suggestedfrom data analysing the formation of thalamocorticalconnections in a lmo4 conditional deletion mouse model(Kashani et al. 2006). Thalamocortical fibers failed tosegregate into distinct barrel specific domains in lmo4 nullmice, resulting in a disturbed organization of the barrel fieldin the somatosensory cortex (Kashani et al. 2006). An earlierstudy already reported that stable over-expression or deletionof LMO4 influences cellular differentiation of neuronal celllines (Vu et al. 2003). These data suggest that LMO4influences neurite growth and is probably involved in severalneuronal pathways.
We show here that LMO4 by interacting with thecytoplasmic domain of Neogenin acts as a signallingmolecule in the RGM A – Neogenin signal transductionpathway, probably linking neurite growth mechanisms and
transcriptional regulation. Although our data support acritical and important role of LMO4 in RGM A inducedneurite growth inhibition, the exact mechanism of the LMO4downstream signaling remains to be elucidated.
Supplementary note
A recent paper published during reviewing of this manuscriptalso reports an interaction between Neogenin and LMO4 in ayeast-two-hybrid system (Goldschneider et al. 2008).
Acknowledgements
We wish to thank Sean Turner for critical reading and manuscript-corrections.
Supporting information
Additional supporting information may be found in the online
version of this article.
Fig. S1 Neogenin, LMO4 and RhoA are expressed in neuronal
cells.
Fig. S2 Definition and measurement of neurite-like extensions in
SH-SY5Y cells.
Fig. S3 siRNA mediated down-regulation of RhoA, Neogenin
and LMO4 in SH-SY5Y cells.
Fig. S4 Expression of LMO4, Neogenin and RGM A in mouse
brain sections.
Please note: Blackwell Publishing are not responsible for the
content or functionality of any supporting information supplied by
the authors. Any queries (other than missing material) should be
directed to the corresponding author for the article.
References
Bach I. (2000) The LIM domain: regulation by association. Mech. Dev.91, 5–17.
Chen H. H., Yip J. W., Stewart A. F. and Frank E. (2002) Differentialexpression of a transcription regulatory factor, the LIM domainonly 4 protein Lmo4, in muscle sensory neurons. Development(Cambridge, England) 129, 4879–4889.
Colland F., Jacq X., Trouplin V., Mougin C., Groizeleau C., HamburgerA., Meil A., Wojcik J., Legrain P. and Gauthier J. M. (2004)Functional proteomics mapping of a human signaling pathway.Genome Res. 14, 1324–1332.
Conrad S., Genth H., Hofmann F., Just I. and Skutella T. (2007)Neogenin-RGMa signaling at the growth cone is bone morpho-genetic protein-independent and involves RhoA, ROCK, and PKC.J. Biol. Chem. 282, 16423–16433.
David S. and Lacroix S. (2003) Molecular approaches to spinal cordrepair. Annu. Rev. Neurosci. 26, 411–440.
Deane J. E., Ryan D. P., Sunde M., Maher M. J., Guss J. M., VisvaderJ. E. and Matthews J. M. (2004) Tandem LIM domains providesynergistic binding in the LMO4:Ldb1 complex. EMBO J. 23,3589–3598.
Drescher U., Bonhoeffer F. and Muller B. K. (1997) The Eph family inretinal axon guidance. Curr. Opin. Neurobiol. 7, 75–80.
Encinas M., Iglesias M., Liu Y., Wang H., Muhaisen A., Cena V.,Gallego C. and Comella J. X. (2000) Sequential treatment of
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431� 2008 The Authors
430 | G. Schaffar et al.

SH-SY5Y cells with retinoic acid and brain-derived neurotrophicfactor gives rise to fully differentiated, neurotrophic factor-dependent, human neuron-like cells. J. Neurochem. 75, 991–1003.
Goldschneider D., Rama M., Guix C. and Mehlen P. (2008) The neo-genin intracellular domain regulates gene transcription via nucleartranslocation. Mol. Cell Biol. 28, 4068–4079.
Hahm K., Sum E. Y., Fujiwara Y., Lindeman G. J., Visvader J. E. andOrkin S. H. (2004) Defective neural tube closure and anteropos-terior patterning in mice lacking the LIM protein LMO4 or itsinteracting partner Deaf-1. Mol. Cell. Biol. 24, 2074–2082.
Hata K., Fujitani M., Yasuda Y., Doya H., Saito T., Yamagishi S.,Mueller B. K. and Yamashita T. (2006) RGMa inhibition promotesaxonal growth and recovery after spinal cord injury. J. Cell Biol.173, 47–58.
Huber A. B., Kolodkin A. L., Ginty D. D. and Cloutier J. F. (2003)Signaling at the growth cone: ligand-receptor complexes and thecontrol of axon growth and guidance. Annu. Rev. Neurosci. 26,509–563.
Kadrmas J. L. and Beckerle M. C. (2004) The LIM domain: from thecytoskeleton to the nucleus. Nat. Rev. 5, 920–931.
Kashani A. H., Qiu Z., Jurata L., Lee S. K., Pfaff S., Goebbels S., NaveK. A. and Ghosh A. (2006) Calcium activation of the LMO4transcription complex and its role in the patterning of thalamo-cortical connections. J. Neurosci. 26, 8398–8408.
Keino-Masu K., Masu M., Hinck L., Leonardo E. D., Chan S. S., CulottiJ. G. and Tessier-Lavigne M. (1996) Deleted in Colorectal Cancer(DCC) encodes a netrin receptor. Cell 87, 175–185.
Lee S. K., Jurata L. W., Nowak R., Lettieri K., Kenny D. A., Pfaff S. L.and Gill G. N. (2005) The LIM domain-only protein LMO4 isrequired for neural tube closure. Mol. Cell. Neurosci. 28, 205–214.
Lein E. S., Hawrylycz M. J., Ao N. et al. (2007) Genome-wide atlas ofgene expression in the adult mouse brain. Nature 445, 168–176.
Lu Z., Lam K. S., Wang N., Xu X., Cortes M. and Andersen B. (2006)LMO4 can interact with Smad proteins and modulate transforminggrowth factor-beta signaling in epithelial cells. Oncogene 25,2920–2930.
Matthews J. M. and Visvader J. E. (2003) LIM-domain-binding protein1: a multifunctional cofactor that interacts with diverse proteins.EMBO Rep. 4, 1132–1137.
Meyerhardt J. A., Look A. T., Bigner S. H. and Fearon E. R. (1997)Identification and characterization of neogenin, a DCC-relatedgene. Oncogene 14, 1129–1136.
Mizunuma H., Miyazawa J., Sanada K. and Imai K. (2003) The LIM-only protein, LMO4, and the LIM domain-binding protein, LDB1,expression in squamous cell carcinomas of the oral cavity. Br.J. Cancer 88, 1543–1548.
Monnier P. P., Sierra A., Macchi P. et al. (2002) RGM is a repulsiveguidance molecule for retinal axons. Nature 419, 392–395.
Mousses S., Bubendorf L., Wagner U. et al. (2002) Clinical validation ofcandidate genes associated with prostate cancer progression in theCWR22 model system using tissue microarrays. Cancer Res. 62,1256–1260.
Mueller B. K., Yamashita T., Schaffar G. and Mueller R. (2006) The roleof repulsive guidance molecules in the embryonic and adult ver-tebrate central nervous system. Philos. Trans. R. Soc. Lond. 361,1513–1529.
Nagata K., Ohashi K., Yang N. and Mizuno K. (1999) The N-terminalLIM domain negatively regulates the kinase activity of LIM-kinase1. Biochem. J. 343 Pt 1, 99–105.
Niederkofler V., Salie R., Sigrist M. and Arber S. (2004) Repulsiveguidance molecule (RGM) gene function is required for neural tube
closure but not retinal topography in the mouse visual system.J. Neurosci. 24, 808–818.
Novotny-Diermayr V., Lin B., Gu L. and Cao X. (2005) Modulation ofthe interleukin-6 receptor subunit glycoprotein 130 complex and itssignaling by LMO4 interaction. J. Biol. Chem. 280, 12747–12757.
Raivich G. and Makwana M. (2007) The making of successful axonalregeneration: genes, molecules and signal transduction pathways.Brain Res. Rev. 53, 287–311.
Rajagopalan S., Deitinghoff L., Davis D., Conrad S., Skutella T.,Chedotal A., Mueller B. K. and Strittmatter S. M. (2004) Neogeninmediates the action of repulsive guidance molecule. Nat. Cell Biol.6, 756–762.
Ren X. R., Ming G. L., Xie Y. et al. (2004) Focal adhesion kinase innetrin-1 signaling. Nat. Neurosci. 7, 1204–1212.
Schaffar G., Breuer P., Boteva R., Behrends C., Tzvetkov N., StrippelN., Sakahira H., Siegers K., Hayer-Hartl M. and Hartl F. U.(2004) Cellular toxicity of polyglutamine expansion proteins:mechanism of transcription factor deactivation. Mol. Cell 15,95–105.
Schmeichel K. L. and Beckerle M. C. (1994) The LIM domain is amodular protein-binding interface. Cell 79, 211–219.
Schwab J. M., Conrad S., Monnier P. P., Julien S., Mueller B. K. andSchluesener H. J. (2005a) Spinal cord injury-induced lesionalexpression of the repulsive guidance molecule (RGM). Eur. J.Neurosci. 21, 1569–1576.
Schwab J. M., Monnier P. P., Schluesener H. J., Conrad S.,Beschorner R., Chen L., Meyermann R. and Mueller B. K. (2005b)Central nervous system injury-induced repulsive guidancemolecule expression in the adult human brain. Arch. Neurol. 62,1561–1568.
Stein E., Zou Y., Poo M. and Tessier-Lavigne M. (2001) Binding ofDCC by netrin-1 to mediate axon guidance independent of ade-nosine A2B receptor activation. Science 291, 1976–1982.
Sugihara T. M., Bach I., Kioussi C., Rosenfeld M. G. and Andersen B.(1998) Mouse deformed epidermal autoregulatory factor 1 recruitsa LIM domain factor, LMO-4, and CLIM coregulators. Proc. NatlAcad. Sci. USA 95, 15418–15423.
Sum E. Y., Peng B., Yu X., Chen J., Byrne J., Lindeman G. J. andVisvader J. E. (2002) The LIM domain protein LMO4 interactswith the cofactor CtIP and the tumor suppressor BRCA1 andinhibits BRCA1 activity. J. Biol. Chem. 277, 7849–7856.
Vielmetter J., Kayyem J. F., Roman J. M. and Dreyer W. J. (1994)Neogenin, an avian cell surface protein expressed during terminalneuronal differentiation, is closely related to the human tumorsuppressor molecule deleted in colorectal cancer. J. Cell Biol. 127,2009–2020.
Visvader J. E., Venter D., Hahm K., Santamaria M., Sum E. Y., O’ReillyL., White D., Williams R., Armes J. and Lindeman G. J. (2001)The LIM domain gene LMO4 inhibits differentiation of mammaryepithelial cells in vitro and is overexpressed in breast cancer. Proc.Natl Acad. Sci. USA 98, 14452–14457.
Vu D., Marin P., Walzer C., Cathieni M. M., Bianchi E. N., Saidji F.,Leuba G., Bouras C. and Savioz A. (2003) Transcription regulatorLMO4 interferes with neuritogenesis in human SH-SY5Y neuro-blastoma cells. Brain Res. 115, 93–103.
Xie Y., Ding Y. Q., Hong Y. et al. (2005) Phosphatidylinositol transferprotein-alpha in netrin-1-induced PLC signalling and neurite out-growth. Nat. Cell Biol. 7, 1124–1132.
Xu Z., Huang S., Chang L. S., Agulnick A. D. and Brandt S. J. (2003)Identification of a TAL1 target gene reveals a positive role for theLIM domain-binding protein Ldb1 in erythroid gene expressionand differentiation. Mol. Cell. Biol. 23, 7585–7599.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 107, 418–431
LMO4 mediates RGM A signaling in neuronal growth cones | 431





























